
VISTA Ligation Reduces Antitumor T-Cell Activity in Pancreatic Cancer

 , , , and
, , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Results
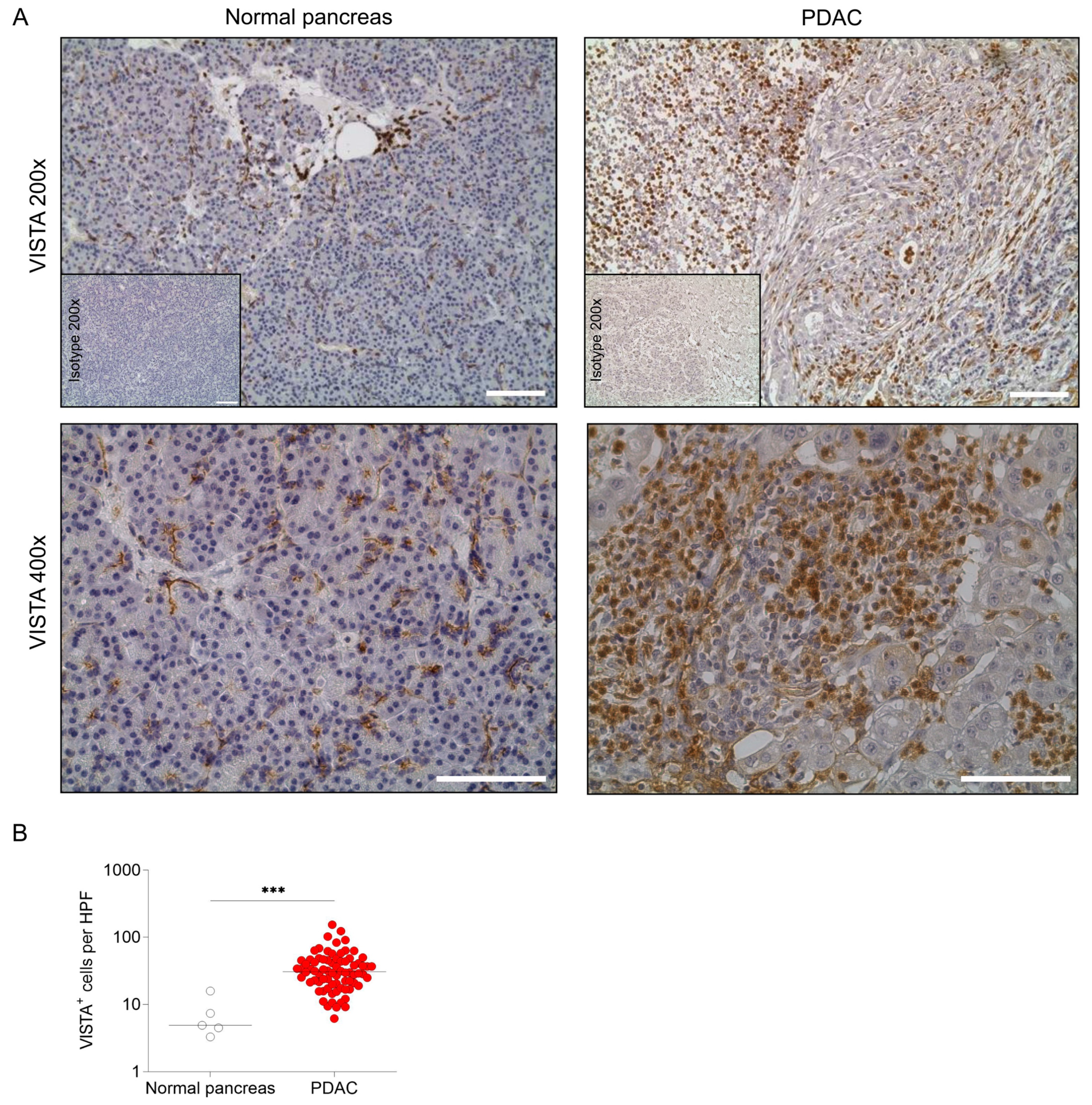
2.1. VISTA Is Expressed in Human PDAC
2.2. VISTA Inhibits Proinflammatory T-Cell Function and Cytokine Production
2.3. VISTA Blockade Reduces Tumor Growth in an Orthotopic PDAC Mouse Model
3. Discussion
4. Materials and Methods
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA A Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef]
- Siegel, R.L.; Miller, K.D.; Fuchs, H.E.; Jemal, A. Cancer Statistics, 2022. CA Cancer J. Clin. 2022, 72, 7–33. [Google Scholar] [CrossRef] [PubMed]
- Ryan, D.P.; Hong, T.S.; Bardeesy, N. Pancreatic Adenocarcinoma. N. Engl. J. Med. 2014, 371, 1039–1049. [Google Scholar] [CrossRef] [PubMed]
- Seifert, A.M.; Reiche, C.; Heiduk, M.; Tannert, A.; Meinecke, A.-C.; Baier, S.; von Renesse, J.; Kahlert, C.; Distler, M.; Welsch, T.; et al. Detection of Pancreatic Ductal Adenocarcinoma with Galectin-9 Serum Levels. Oncogene 2020, 39, 3102–3113. [Google Scholar] [CrossRef] [PubMed]
- Le, D.T.; Durham, J.N.; Smith, K.N.; Wang, H.; Bartlett, B.R.; Aulakh, L.K.; Lu, S.; Kemberling, H.; Wilt, C.; Luber, B.S.; et al. Mismatch Repair Deficiency Predicts Response of Solid Tumors to PD-1 Blockade. Science 2017, 357, 409–413. [Google Scholar] [CrossRef]
- O’Reilly, E.M.; Oh, D.-Y.; Dhani, N.; Renouf, D.J.; Lee, M.A.; Sun, W.; Fisher, G.; Hezel, A.; Chang, S.-C.; Vlahovic, G.; et al. Durvalumab With or Without Tremelimumab for Patients With Metastatic Pancreatic Ductal Adenocarcinoma: A Phase 2 Randomized Clinical Trial. JAMA Oncol. 2019, 5, 1431–1438. [Google Scholar] [CrossRef]
- Hu, Z.I.; Shia, J.; Stadler, Z.K.; Varghese, A.M.; Capanu, M.; Salo-Mullen, E.; Lowery, M.A.; Diaz, L.A.; Mandelker, D.; Yu, K.H.; et al. Evaluating Mismatch Repair Deficiency in Pancreatic Adenocarcinoma: Challenges and Recommendations. Clin. Cancer Res. 2018, 24, 1326–1336. [Google Scholar] [CrossRef]
- Bockorny, B.; Grossman, J.E.; Hidalgo, M. Facts and Hopes in Immunotherapy of Pancreatic Cancer. Clin. Cancer Res. 2022, 28, OF1–OF12. [Google Scholar] [CrossRef]
- Conforti, F.; Pala, L.; Bagnardi, V.; Pas, T.D.; Martinetti, M.; Viale, G.; Gelber, R.D.; Goldhirsch, A. Cancer Immunotherapy Efficacy and Patients’ Sex: A Systematic Review and Meta-Analysis. Lancet Oncol. 2018, 19, 737–746. [Google Scholar] [CrossRef]
- Heiduk, M.; Plesca, I.; Glück, J.; Müller, L.; Digomann, D.; Reiche, C.; von Renesse, J.; Decker, R.; Kahlert, C.; Sommer, U.; et al. Neoadjuvant Chemotherapy Drives Intratumoral T Cells toward a Proinflammatory Profile in Pancreatic Cancer. JCI Insight 2022, 7, e152761. [Google Scholar] [CrossRef]
- Seifert, A.M.; List, J.; Heiduk, M.; Decker, R.; von Renesse, J.; Meinecke, A.-C.; Aust, D.E.; Welsch, T.; Weitz, J.; Seifert, L. Gamma-Delta T Cells Stimulate IL-6 Production by Pancreatic Stellate Cells in Pancreatic Ductal Adenocarcinoma. J. Cancer Res. Clin. Oncol. 2020, 146, 3233–3240. [Google Scholar] [CrossRef]
- Carstens, J.L.; Correa de Sampaio, P.; Yang, D.; Barua, S.; Wang, H.; Rao, A.; Allison, J.P.; LeBleu, V.S.; Kalluri, R. Spatial Computation of Intratumoral T Cells Correlates with Survival of Patients with Pancreatic Cancer. Nat. Commun. 2017, 8, 15095. [Google Scholar] [CrossRef] [PubMed]
- Thyagarajan, A.; Alshehri, M.S.A.; Miller, K.L.R.; Sherwin, C.M.; Travers, J.B.; Sahu, R.P. Myeloid-Derived Suppressor Cells and Pancreatic Cancer: Implications in Novel Therapeutic Approaches. Cancers 2019, 11, 1627. [Google Scholar] [CrossRef]
- Steele, N.G.; Carpenter, E.S.; Kemp, S.B.; Sirihorachai, V.; The, S.; Delrosario, L.; Lazarus, J.; Amir, E.D.; Gunchick, V.; Espinoza, C.; et al. Multimodal Mapping of the Tumor and Peripheral Blood Immune Landscape in Human Pancreatic Cancer. Nat. Cancer 2020, 1, 1097–1112. [Google Scholar] [CrossRef]
- Wartenberg, M.; Cibin, S.; Zlobec, I.; Vassella, E.; Eppenberger-Castori, S.; Terracciano, L.; Eichmann, M.D.; Worni, M.; Gloor, B.; Perren, A.; et al. Integrated Genomic and Immunophenotypic Classification of Pancreatic Cancer Reveals Three Distinct Subtypes with Prognostic/Predictive Significance. Clin. Cancer Res. 2018, 24, 4444–4454. [Google Scholar] [CrossRef] [PubMed]
- Seifert, L.; Plesca, I.; Müller, L.; Sommer, U.; Heiduk, M.; von Renesse, J.; Digomann, D.; Glück, J.; Klimova, A.; Weitz, J.; et al. LAG-3-Expressing Tumor-Infiltrating T Cells Are Associated with Reduced Disease-Free Survival in Pancreatic Cancer. Cancers 2021, 13, 1297. [Google Scholar] [CrossRef]
- Fukunaga, A.; Miyamoto, M.; Cho, Y.; Murakami, S.; Kawarada, Y.; Oshikiri, T.; Kato, K.; Kurokawa, T.; Suzuoki, M.; Nakakubo, Y.; et al. CD8+ Tumor-Infiltrating Lymphocytes Together with CD4+ Tumor-Infiltrating Lymphocytes and Dendritic Cells Improve the Prognosis of Patients with Pancreatic Adenocarcinoma. Pancreas 2004, 28, e26–e31. [Google Scholar] [CrossRef] [PubMed]
- Balli, D.; Rech, A.J.; Stanger, B.Z.; Vonderheide, R.H. Immune Cytolytic Activity Stratifies Molecular Subsets of Human Pancreatic Cancer. Clin. Cancer Res. 2017, 23, 3129–3138. [Google Scholar] [CrossRef]
- Seifert, A.M.; Eymer, A.; Heiduk, M.; Wehner, R.; Tunger, A.; von Renesse, J.; Decker, R.; Aust, D.E.; Welsch, T.; Reissfelder, C.; et al. PD-1 Expression by Lymph Node and Intratumoral Regulatory T Cells Is Associated with Lymph Node Metastasis in Pancreatic Cancer. Cancers 2020, 12, 2756. [Google Scholar] [CrossRef]
- Robert, C. A Decade of Immune-Checkpoint Inhibitors in Cancer Therapy. Nat. Commun. 2020, 11, 3801. [Google Scholar] [CrossRef]
- Wang, L.; Rubinstein, R.; Lines, J.L.; Wasiuk, A.; Ahonen, C.; Guo, Y.; Lu, L.-F.; Gondek, D.; Wang, Y.; Fava, R.A.; et al. VISTA, a Novel Mouse Ig Superfamily Ligand That Negatively Regulates T Cell Responses. J. Exp. Med. 2011, 208, 577–592. [Google Scholar] [CrossRef] [PubMed]
- Flies, D.B.; Han, X.; Higuchi, T.; Zheng, L.; Sun, J.; Ye, J.J.; Chen, L. Coinhibitory Receptor PD-1H Preferentially Suppresses CD4+ T Cell–Mediated Immunity. J. Clin. Investig. 2014, 124, 1966–1975. [Google Scholar] [CrossRef] [PubMed]
- ElTanbouly, M.A.; Croteau, W.; Noelle, R.J.; Lines, J.L. VISTA: A Novel Immunotherapy Target for Normalizing Innate and Adaptive Immunity. Semin. Immunol. 2019, 42, 101308. [Google Scholar] [CrossRef] [PubMed]
- Lines, J.L.; Pantazi, E.; Mak, J.; Sempere, L.F.; Wang, L.; O’Connell, S.; Ceeraz, S.; Suriawinata, A.A.; Yan, S.; Ernstoff, M.S.; et al. VISTA Is an Immune Checkpoint Molecule for Human T Cells. Cancer Res. 2014, 74, 1924–1932. [Google Scholar] [CrossRef]
- Wang, L.; Mercier, I.L.; Putra, J.; Chen, W.; Liu, J.; Schenk, A.D.; Nowak, E.C.; Suriawinata, A.A.; Li, J.; Noelle, R.J. Disruption of the Immune-Checkpoint VISTA Gene Imparts a Proinflammatory Phenotype with Predisposition to the Development of Autoimmunity. Proc. Natl. Acad. Sci. USA 2014, 111, 14846–14851. [Google Scholar] [CrossRef]
- Han, X.; Vesely, M.D.; Yang, W.; Sanmamed, M.F.; Badri, T.; Alawa, J.; López-Giráldez, F.; Gaule, P.; Lee, S.W.; Zhang, J.-P.; et al. PD-1H (VISTA)–Mediated Suppression of Autoimmunity in Systemic and Cutaneous Lupus Erythematosus. Sci. Transl. Med. 2019, 11, eaax1159. [Google Scholar] [CrossRef]
- Mercier, I.L.; Chen, W.; Lines, J.L.; Day, M.; Li, J.; Sergent, P.; Noelle, R.J.; Wang, L. VISTA Regulates the Development of Protective Antitumor Immunity. Cancer Res. 2014, 74, 1933–1944. [Google Scholar] [CrossRef]
- Gao, J.; Ward, J.F.; Pettaway, C.A.; Shi, L.Z.; Subudhi, S.K.; Vence, L.M.; Zhao, H.; Chen, J.; Chen, H.; Efstathiou, E.; et al. VISTA Is an Inhibitory Immune Checkpoint That Is Increased after Ipilimumab Therapy in Patients with Prostate Cancer. Nat. Med. 2017, 23, 551–555. [Google Scholar] [CrossRef]
- Kuklinski, L.F.; Yan, S.; Li, Z.; Fisher, J.L.; Cheng, C.; Noelle, R.J.; Angeles, C.V.; Turk, M.J.; Ernstoff, M.S. VISTA Expression on Tumor-Infiltrating Inflammatory Cells in Primary Cutaneous Melanoma Correlates with Poor Disease-Specific Survival. Cancer Immunol. Immunother. 2018, 67, 1113–1121. [Google Scholar] [CrossRef]
- Villarroel-Espindola, F.; Yu, X.; Datar, I.; Mani, N.; Sanmamed, M.; Velcheti, V.; Syrigos, K.; Toki, M.; Zhao, H.; Chen, L.; et al. Spatially Resolved and Quantitative Analysis of VISTA/PD-1H as a Novel Immunotherapy Target in Human Non–Small Cell Lung Cancer. Clin. Cancer. Res. 2018, 24, 1562–1573. [Google Scholar] [CrossRef]
- Blando, J.; Sharma, A.; Higa, M.G.; Zhao, H.; Vence, L.; Yadav, S.S.; Kim, J.; Sepulveda, A.M.; Sharp, M.; Maitra, A.; et al. Comparison of Immune Infiltrates in Melanoma and Pancreatic Cancer Highlights VISTA as a Potential Target in Pancreatic Cancer. Proc. Natl. Acad. Sci. USA 2019, 116, 1692–1697. [Google Scholar] [CrossRef] [PubMed]
- Deng, J.; Li, J.; Sarde, A.; Lines, J.L.; Lee, Y.-C.; Qian, D.C.; Pechenick, D.A.; Manivanh, R.; Mercier, I.L.; Lowrey, C.H.; et al. Hypoxia-Induced VISTA Promotes the Suppressive Function of Myeloid-Derived Suppressor Cells in the Tumor Microenvironment. Cancer Immunol. Res. 2019, 7, 1079–1090. [Google Scholar] [CrossRef] [PubMed]
- Curis, Inc. Phase 1 Study of CI-8993 Anti-VISTA Antibody in Patients With Advanced Solid Tumor Malignancies; clinicaltrials.gov; Curis, Inc.: Lexington, KY, USA, 2022. [Google Scholar]
- Sasikumar, P.G.; Sudarshan, N.S.; Adurthi, S.; Ramachandra, R.K.; Samiulla, D.S.; Lakshminarasimhan, A.; Ramanathan, A.; Chandrasekhar, T.; Dhudashiya, A.A.; Talapati, S.R.; et al. PD-1 Derived CA-170 Is an Oral Immune Checkpoint Inhibitor That Exhibits Preclinical Anti-Tumor Efficacy. Commun. Biol. 2021, 4, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Nomi, T.; Sho, M.; Akahori, T.; Hamada, K.; Kubo, A.; Kanehiro, H.; Nakamura, S.; Enomoto, K.; Yagita, H.; Azuma, M.; et al. Clinical Significance and Therapeutic Potential of the Programmed Death-1 Ligand/Programmed Death-1 Pathway in Human Pancreatic Cancer. Clin. Cancer Res. 2007, 13, 2151–2157. [Google Scholar] [CrossRef] [PubMed]
- Yarchoan, M.; Albacker, L.A.; Hopkins, A.C.; Montesion, M.; Murugesan, K.; Vithayathil, T.T.; Zaidi, N.; Azad, N.S.; Laheru, D.A.; Frampton, G.M.; et al. PD-L1 Expression and Tumor Mutational Burden Are Independent Biomarkers in Most Cancers. JCI Insight 2019, 4, 126908. [Google Scholar] [CrossRef]
- Timmer, F.E.F.; Geboers, B.; Nieuwenhuizen, S.; Dijkstra, M.; Schouten, E.A.C.; Puijk, R.S.; de Vries, J.J.J.; van den Tol, M.P.; Bruynzeel, A.M.E.; Streppel, M.M.; et al. Pancreatic Cancer and Immunotherapy: A Clinical Overview. Cancers 2021, 13, 4138. [Google Scholar] [CrossRef]
- Mulati, K.; Hamanishi, J.; Matsumura, N.; Chamoto, K.; Mise, N.; Abiko, K.; Baba, T.; Yamaguchi, K.; Horikawa, N.; Murakami, R.; et al. VISTA Expressed in Tumour Cells Regulates T Cell Function. Br. J. Cancer 2019, 120, 115–127. [Google Scholar] [CrossRef]
- Schaafsma, E.; Croteau, W.; ElTanbouly, M.; Nowak, E.C.; Smits, N.C.; Deng, J.; Sarde, A.; Webber, C.A.; Rabadi, D.; Cheng, C.; et al. VISTA Targeting of T-Cell Quiescence and Myeloid Suppression Overcomes Adaptive Resistance. Cancer Immunol. Res. 2023, 11, 38–55. [Google Scholar] [CrossRef]
- Xie, S.; Huang, J.; Qiao, Q.; Zang, W.; Hong, S.; Tan, H.; Dong, C.; Yang, Z.; Ni, L. Expression of the Inhibitory B7 Family Molecule VISTA in Human Colorectal Carcinoma Tumors. Cancer Immunol. Immunother. 2018, 67, 1685–1694. [Google Scholar] [CrossRef]
- Hummingbird Bioscience, Inc. A Phase 1 Study of HMBD-002-V4C26 (HMBD-002), a Monoclonal Antibody Targeting VISTA, as Monotherapy and Combined With Pembrolizumab, in Patients With Advanced Solid Malignancies; clinicaltrials.gov; Hummingbird Bioscience, Inc.: Singapore, 2022. [Google Scholar]
- Olson, B.; Li, Y.; Lin, Y.; Liu, E.T.; Patnaik, A. Mouse Models for Cancer Immunotherapy Research. Cancer Discov 2018, 8, 1358–1365. [Google Scholar] [CrossRef]
- Partecke, L.I.; Sendler, M.; Kaeding, A.; Weiss, F.U.; Mayerle, J.; Dummer, A.; Nguyen, T.D.; Albers, N.; Speerforck, S.; Lerch, M.M.; et al. A Syngeneic Orthotopic Murine Model of Pancreatic Adenocarcinoma in the C57/BL6 Mouse Using the Panc02 and 6606PDA Cell Lines. ESR 2011, 47, 98–107. [Google Scholar] [CrossRef] [PubMed]
- Seifert, L.; Werba, G.; Tiwari, S.; Giao Ly, N.N.; Alothman, S.; Alqunaibit, D.; Avanzi, A.; Barilla, R.; Daley, D.; Greco, S.H.; et al. The Necrosome Promotes Pancreatic Oncogenesis via CXCL1 and Mincle-Induced Immune Suppression. Nature 2016, 532, 245–249. [Google Scholar] [CrossRef] [PubMed]
- Cerami, E.; Gao, J.; Dogrusoz, U.; Gross, B.E.; Sumer, S.O.; Aksoy, B.A.; Jacobsen, A.; Byrne, C.J.; Heuer, M.L.; Larsson, E.; et al. The CBio Cancer Genomics Portal: An Open Platform for Exploring Multidimensional Cancer Genomics Data. Cancer Discov. 2012, 2, 401–404. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Years | ||
|---|---|---|
| Mean age | 67 | |
| Age range | 47–80 | |
| n (%) | ||
| Sex | ||
| Female | 36 | (47.37) |
| Male | 40 | (52.63) |
| pT Stage | ||
| 1 | 5 | (6.58) |
| 2 | 45 | (59.21) |
| 3 | 25 | (32.89) |
| 4 | 0 | (0) |
| Unknown | 1 | (1.32) |
| pN Stage | ||
| 0 | 40 | (52.63) |
| 1 | 27 | (35.53) |
| 2 | 9 | (11.84) |
| M Stage | ||
| 0 | 76 | (100) |
| 1 | 0 | (0) |
| UICC Stage | ||
| I | 27 | (35.53) |
| II | 39 | (51.32) |
| III | 9 | (11.84) |
| IV | 0 | (0) |
| Unknown | 1 | (1.31) |
| Neoadjuvant Treatment | ||
| Yes | 25 | (32.89) |
| No | 51 | (67.11) |
| Years | ||
|---|---|---|
| Mean age | 67 | |
| Age range | 36–79 | |
| n (%) | ||
| Sex | ||
| Female | 36 | (53.73) |
| Male | 31 | (46.27) |
| pT Stage | ||
| 1 | 7 | (10.45) |
| 2 | 39 | (58.21) |
| 3 | 20 | (29.85) |
| 4 | 0 | (0) |
| Unknown | 1 | (1.49) |
| pN Stage | ||
| 0 | 30 | (44.78) |
| 1 | 26 | (38.8) |
| 2 | 11 | (16.42) |
| M Stage | ||
| 0 | 66 | (98.51) |
| 1 | 1 | (1.49) |
| UICC Stage | ||
| I | 7 | (10.45) |
| II | 48 | (71.64) |
| III | 11 | (16.42) |
| IV | 1 | (1.49) |
| Neoadjuvant Treatment | ||
| Yes | 16 | (23.88) |
| No | 51 | (76.12) |
| Years | ||
|---|---|---|
| Mean age | 67 | |
| Age range | 49–83 | |
| n (%) | ||
| Sex | ||
| Female | 5 | (38.5) |
| Male | 8 | (61.5) |
| pT Stage | ||
| 1 | 1 | (7.7) |
| 2 | 2 | (15.4) |
| 3 | 8 | (61.5) |
| 4 | 2 | (15.4) |
| pN Stage | ||
| 0 | 5 | (38.5) |
| 1 | 6 | (46.1) |
| 2 | 2 | (15.4) |
| M Stage | ||
| 0 | 11 | (84.6) |
| 1 | 2 | (15.4) |
| UICC Stage | ||
| I | 2 | (15.4) |
| II | 7 | (53.8) |
| III | 2 | (15.4) |
| IV | 2 | (15.4) |
| Neoadjuvant Treatment | ||
| Yes | 2 | (15.4) |
| No | 11 | (84.6) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Digomann, D.; Strack, J.; Heiduk, M.; Plesca, I.; Rupp, L.; Reiche, C.; Nicolaus, S.; Beer, C.; Sommer, U.; Schmitz, M.; et al. VISTA Ligation Reduces Antitumor T-Cell Activity in Pancreatic Cancer. Cancers 2023, 15, 2326. https://doi.org/10.3390/cancers15082326
Digomann D, Strack J, Heiduk M, Plesca I, Rupp L, Reiche C, Nicolaus S, Beer C, Sommer U, Schmitz M, et al. VISTA Ligation Reduces Antitumor T-Cell Activity in Pancreatic Cancer. Cancers. 2023; 15(8):2326. https://doi.org/10.3390/cancers15082326
Chicago/Turabian StyleDigomann, David, Johannes Strack, Max Heiduk, Ioana Plesca, Luise Rupp, Charlotte Reiche, Simone Nicolaus, Carolin Beer, Ulrich Sommer, Marc Schmitz, and et al. 2023. "VISTA Ligation Reduces Antitumor T-Cell Activity in Pancreatic Cancer" Cancers 15, no. 8: 2326. https://doi.org/10.3390/cancers15082326
APA StyleDigomann, D., Strack, J., Heiduk, M., Plesca, I., Rupp, L., Reiche, C., Nicolaus, S., Beer, C., Sommer, U., Schmitz, M., Distler, M., Weitz, J., Seifert, A. M., & Seifert, L. (2023). VISTA Ligation Reduces Antitumor T-Cell Activity in Pancreatic Cancer. Cancers, 15(8), 2326. https://doi.org/10.3390/cancers15082326