Correction: Reschke, R.; Olson, D.J. Leveraging STING, Batf3 Dendritic Cells, CXCR3 Ligands, and Other Components Related to Innate Immunity to Induce a “Hot” Tumor Microenvironment That Is Responsive to Immunotherapy. Cancers 2022, 14, 2458


- 42. Garofalo, C.; De Marco, C.; Cristiani, C.M. NK Cells in the Tumor Microenvironment as New Potential Players Mediating Chemotherapy Effects in Metastatic Melanoma. Front. Oncol. 2021, 12, 754541. https://doi.org/10.3389/fonc.2021.754541.
- 43. McKay, K.; Moore, P.C.; Smoller, B.R.; Hiatt, K.M. Association between natural killer cells and regression in melanocytic lesions. Hum. Pathol. 2011, 42, 1960–1964. https://doi.org/10.1016/j.humpath.2011.02.019.
- 44. Reschke, R.; Dumann, K.; Ziemer, M. Risk Stratification and Clinical Characteristics of Patients with Late Recurrence of Melanoma (>10 Years). J. Clin. Med. 2022, 11, 2026. https://doi.org/10.3390/jcm11072026.
- 45. Mittal, D.; Vijayan, D.; Putz, E.M.; Aguilera, A.R.; Markey, K.A.; Straube, J.; Kazakoff, S.; Nutt, S.L.; Takeda, K.; Hill, G.R.; et al. Interleukin-12 from CD103+ Batf3-Dependent Dendritic Cells Required for NK-Cell Suppression of Metastasis. Cancer Immunol. Res. 2017, 5, 1098–1108. https://doi.org/10.1158/2326-6066.
- 46. Mattei, F.; Schiavoni, G.; Belardelli, F.; Tough, D.F. IL-15 is expressed by dendritic cells in response to type I IFN, double-stranded RNA, or lipopolysaccharide and promotes dendritic cell activation. J. Immunol. 2001, 167, 1179–1187. https://doi.org/ 10.4049/jimmunol.167.3.1179.
- 47. Bergamaschi, C.; Pandit, H.; Nagy, B.A.; Stellas, D.; Jensen, S.M.; Bear, J.; Cam, M.; Valentin, A.; Fox, B.A.; Felber, B.K.; et al. Heterodimeric IL-15 delays tumor growth and promotes intratumoral CTL and dendritic cell accumulation by a cytokine network involving XCL1, IFN--γ, CXCL9 and CXCL10. J. Immunother. Cancer 2020, 8, e000599. https://doi.org/10.1136/jitc-2020-000599.
- 48. Reschke, R.; Gajewski, TF. CXCL9 and CXCL10 bring the heat to tumors. Sci. Immunol. 2022, 7, eabq6509. https://doi.org/10.1126/sciimmunol.abq6509.
- 49. Hoch, T.; Schulz, D.; Eling, N.; Gómez, J.M.; Levesque, M.P.; Bodenmiller, B. Multiplexed imaging mass cytometry of the chemokine milieus in melanoma characterizes features of the response to immunotherapy. Sci. Immunol. 2022, 7, eabk1692. https://doi.org/10.1126/sciimmunol.abk1692.
- 60. Orr, B.; Mahdi, H.; Fang, Y.; Strange, M.; Uygun, I.; Rana, M.; Zhang, L.; Mora, A.S.; Pusateri, A.; Elishaev, E.; et al. Phase I trial combining chemokine-targeting with loco-regional chemoimmunotherapy for recurrent, platinum-sensitive ovarian cancer shows induction of CXCR3 ligands and markers of type 1 immunity. Clin. Cancer Res. 2022, 28, 2038–2049.
- 61. Mansurov, A.; Hosseinchi, P.; Chang, K.; Lauterbach, A.L.; Gray, L.T.; Alpar, A.T.; Budina, E.; Slezak, A.J.; Kang, S.; Cao, S.; et al. Masking the immunotoxicity of interleukin-12 by fusing it with a domain of its receptor via a tumour-protease-cleavable linker. Nat. Biomed. Eng. 2022, 6, 819–829. https://doi.org/10.1038/s41551-022-00888-0.
- 68. Reschke, R.; Gussek, P.; Boldt, A.; Sack, U.; Köhl, U.; Lordick, F.; Gora, T.; Kreuz, M.; Reiche, K.; Simon, J.C.; et al. Distinct Immune Signatures Indicative of Treatment Response and Immune-Related Adverse Events in Melanoma Patients under Immune Checkpoint Inhibitor Therapy. Int. J. Mol. Sci. 2021, 22, 8017. https://doi.org/10.3390/ijms22158017.
- 74. Reschke, R.; Jäger, I.; Mehnert-Theuerkauf, A.; Ziemer, M. Therapy understanding and health related quality of life in stage III/IV melanoma patients treated with novel adjuvant therapies. J. Dtsch. Dermatol. Ges. 2021, 19, 215–221. https://doi.org/10.1111/ddg.14317.
Reference
- Reschke, R.; Olson, D.J. Leveraging STING, Batf3 Dendritic Cells, CXCR3 Ligands, and Other Components Related to Innate Immunity to Induce A “Hot” Tumor Microenvironment That Is Responsive to Immunotherapy. Cancers 2022, 14, 2458. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
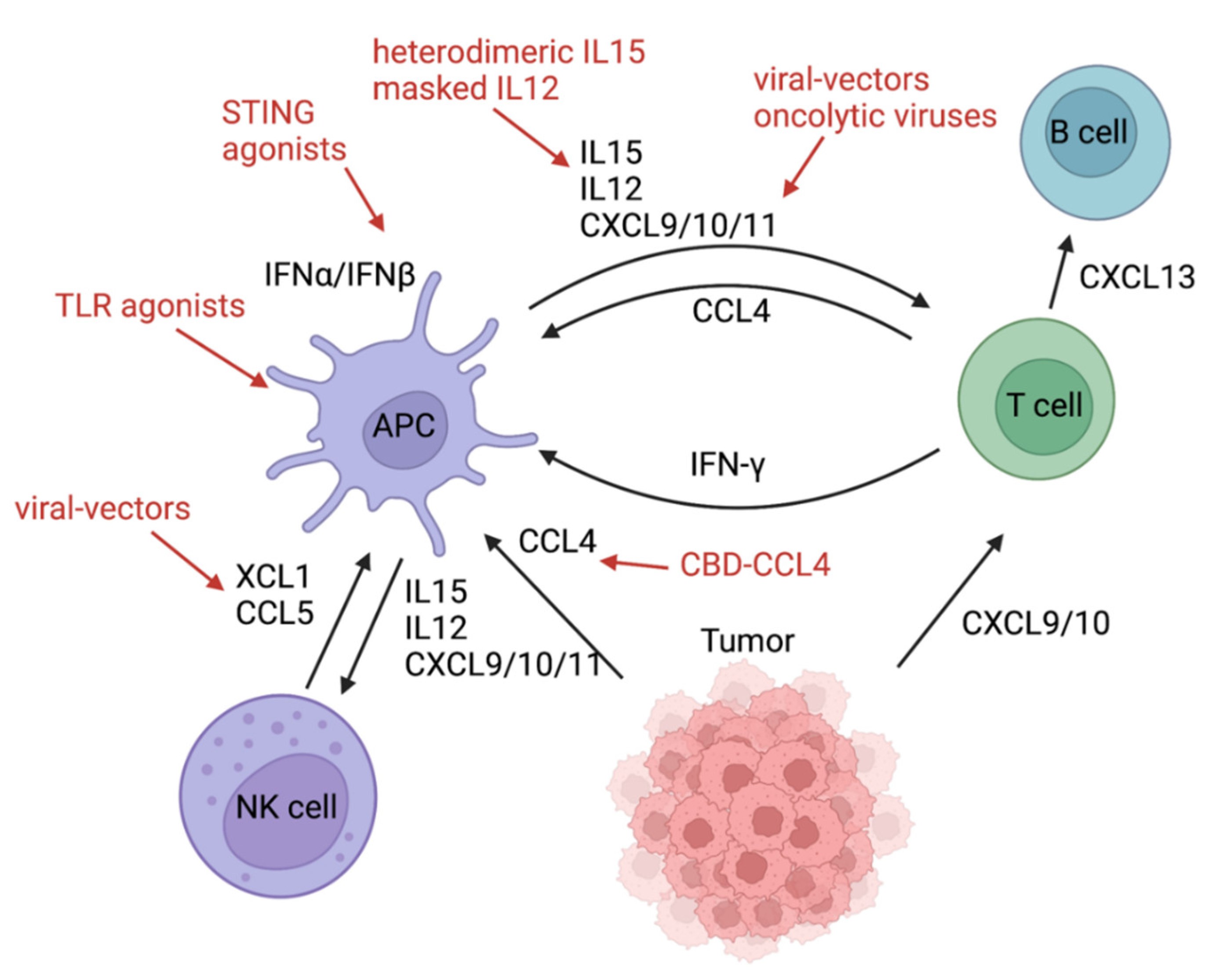
| Components | Function | Species | Delivery Route/Therapeutic Agent | Year/Citation |
|---|---|---|---|---|
| STING | Sensing tumor-derived DNA, DC maturation, and CTL priming | mouse | systemic administration of SR-717 in a “closed” conformation | 2020 [28] |
| mouse | oral administration of MSA-2 in a “closed” conformation | 2020 [29] | ||
| mouse | engineered extracellular vesicle exogenously loaded with cyclic dinucleotide | 2021 [30] | ||
| human | intravenous infusion of TAK-676 | ongoing NCT04420884 | ||
| human | intravenous infusion of SB 11285 | ongoing NCT04096638 | ||
| human | intravenous infusion of SNX281 | ongoing NCT04609579 | ||
| human | intratumoral injection of CDK-002 | ongoing NCT04592484 | ||
| TLR9 | sensing tumor-derived DNA, DC maturation, and CTL priming | human | intratumoral injection of Vidutolimod | 2021 [24] |
| mouse | intratumoral injection CpG oligodeoxynucleotide (TLR9 ligand) and an antibody against OX40 | 2022 [70] | ||
| TLR3 | maturation and CTL priming | human | intraperitoneal injection of rintalomid (TLR3 agonist), celecoxib, and cisplatin | 2022 [60] |
| VEGF | local lymphangiogenesis, immune cell trafficking, and CTL activation | mouse | injection of “VEGFC vax” | 2021 [35] |
| XCL1+ Flt3L | DC recruitment and expansion | mouse | intratumoral injection of XCL1 and SFlt3L encoded in recombinant Semliki Forest virus-derived vectors | 2018 [50] |
| CCL4 | DC recruitment | mouse | intravenous administration of a fusion protein of CCL4 and the collagen-binding domain of von Willebrand factor | 2019 [51] |
| CXCL9/10/11 | CTL recruitment | mouse | CXCL9 and OX40L | 2020 [53] |
| mouse | intravenous injection of oncolytic vesicular stomatitis virus encodes CXCL9 | 2020 [55] | ||
| mouse | genetically engineered mesenchymal stem cells producing CXCL10 | 2018 [71] | ||
| humanized mouse | injection of CXCL10-producing SynNotch T cells | 2021 [72] | ||
| mouse | intravenous delivery of CXCL9/10/11 plasmids by nanoparticles | 2022 [73] | ||
| human | NG-641 is an oncolytic adenoviral vector which expresses a FAP-TAc antibody together with an immune enhancer module (CXCL9/CXCL10/IFNα). | ongoing NCT04053283 | ||
| IL12 | DC activation/IFNγ production of T and NK cells | mouse | intravenous injection of IL12 fused to a domain of the IL12 receptor | 2022 [61] |
| IL15 | DC recruitment/activation and downstream recruitment of CTLs/NK cells | mouse | intraperitoneal injection of heterodimeric IL-15 | 2020 [47] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Reschke, R.; Olson, D.J. Correction: Reschke, R.; Olson, D.J. Leveraging STING, Batf3 Dendritic Cells, CXCR3 Ligands, and Other Components Related to Innate Immunity to Induce a “Hot” Tumor Microenvironment That Is Responsive to Immunotherapy. Cancers 2022, 14, 2458. Cancers 2024, 16, 1234. https://doi.org/10.3390/cancers16061234
Reschke R, Olson DJ. Correction: Reschke, R.; Olson, D.J. Leveraging STING, Batf3 Dendritic Cells, CXCR3 Ligands, and Other Components Related to Innate Immunity to Induce a “Hot” Tumor Microenvironment That Is Responsive to Immunotherapy. Cancers 2022, 14, 2458. Cancers. 2024; 16(6):1234. https://doi.org/10.3390/cancers16061234
Chicago/Turabian StyleReschke, Robin, and Daniel J. Olson. 2024. "Correction: Reschke, R.; Olson, D.J. Leveraging STING, Batf3 Dendritic Cells, CXCR3 Ligands, and Other Components Related to Innate Immunity to Induce a “Hot” Tumor Microenvironment That Is Responsive to Immunotherapy. Cancers 2022, 14, 2458" Cancers 16, no. 6: 1234. https://doi.org/10.3390/cancers16061234
APA StyleReschke, R., & Olson, D. J. (2024). Correction: Reschke, R.; Olson, D.J. Leveraging STING, Batf3 Dendritic Cells, CXCR3 Ligands, and Other Components Related to Innate Immunity to Induce a “Hot” Tumor Microenvironment That Is Responsive to Immunotherapy. Cancers 2022, 14, 2458. Cancers, 16(6), 1234. https://doi.org/10.3390/cancers16061234