
A Novel ceRNET Relying on the lncRNA JPX, miR-378a-3p, and Its mRNA Targets in Lung Cancer

 ,
,  , , and
, , and 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. DNA Constructs
2.2. Cell Cultures, Transfections, and Luciferase Assays
2.3. Cell Proliferation, Cell Migration, and Invasion Assays
2.4. 3D-Sheroid Formation Assay
2.5. RNA Purification and Real-Time PCR Analyses
2.6. Statistical and Bioinformatic Analyses
3. Results
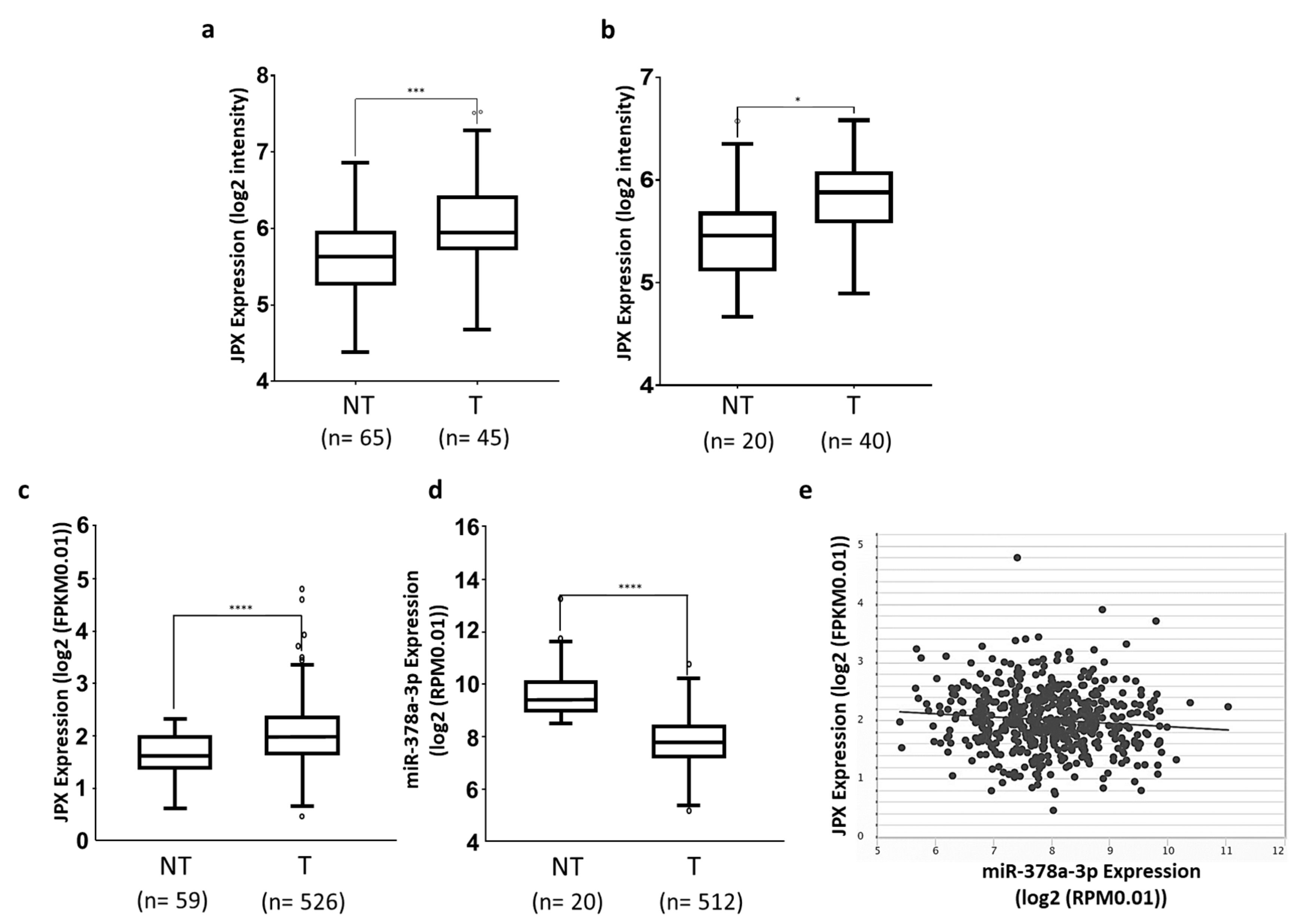
3.1. JPX Is Upregulated in Lung Tumor Tissues
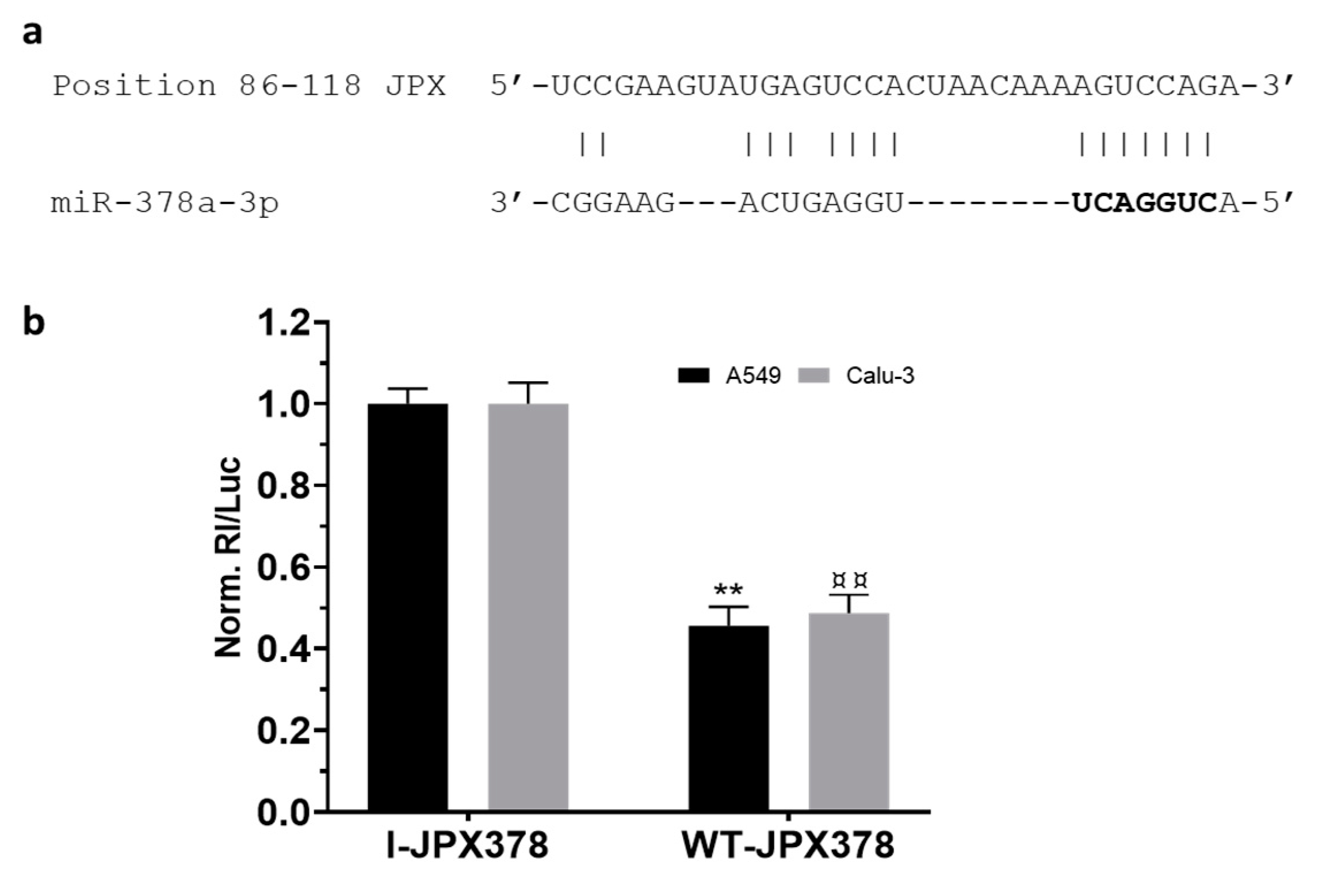
3.2. miR-378a Is Inversely Correlated to JPX Expression in Lung Tumor and Interacts with JPX
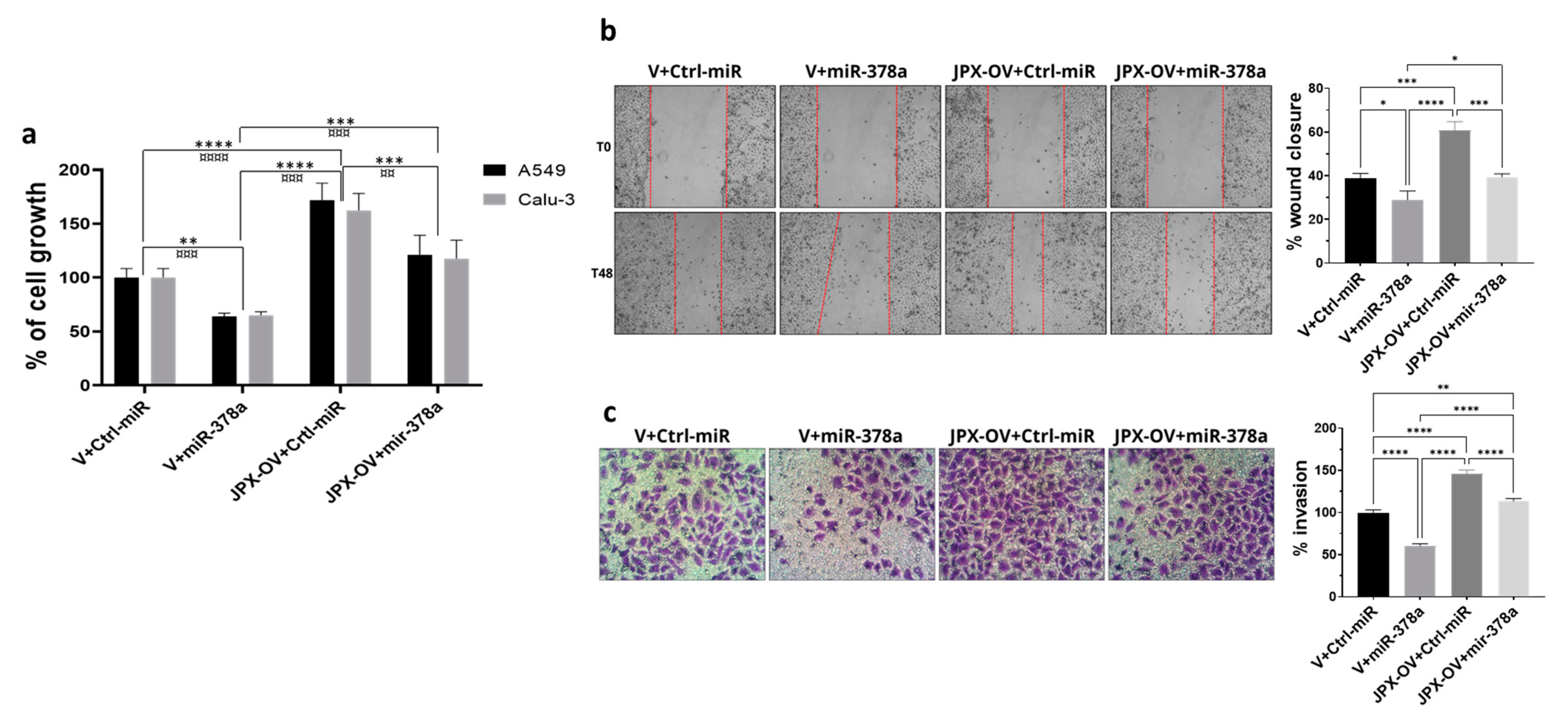
3.3. JPX Promotes Cell Proliferation, Migration, and Invasion by Sponging miR-378a
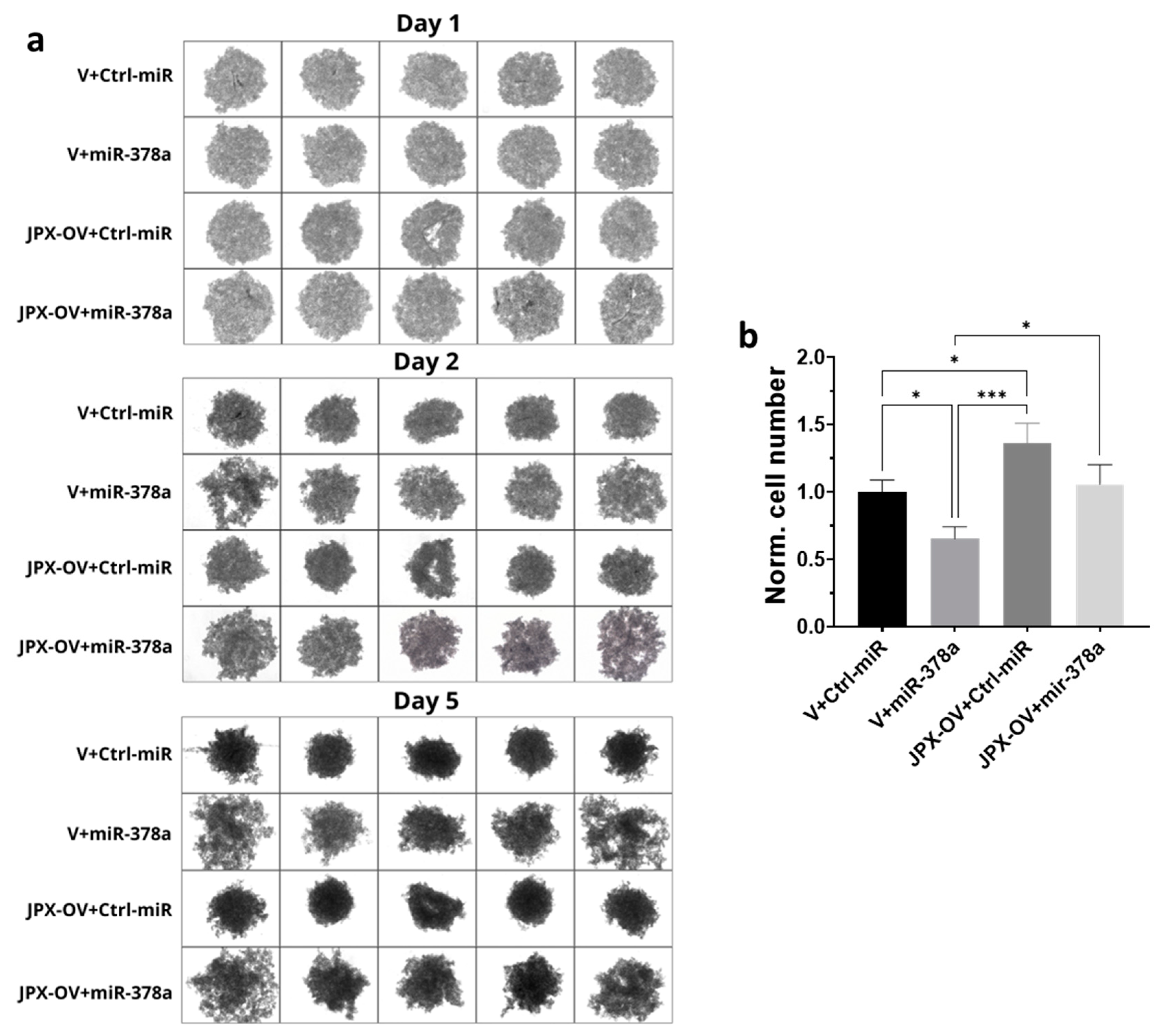
3.4. Functional Impact of JPX, miR-378a, and Their Combination on 3D-Spheroid Formation
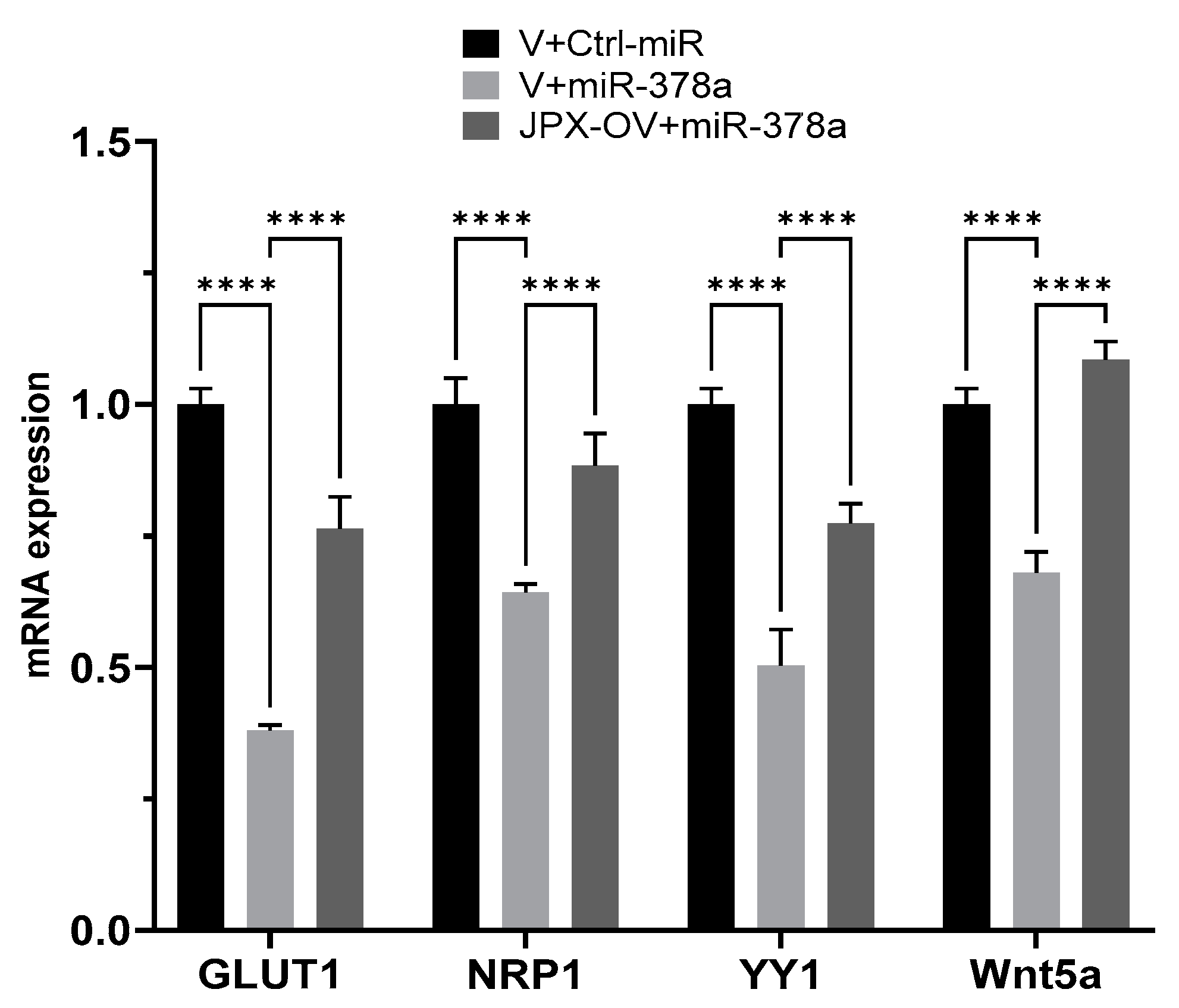
3.5. JPX Counteracts miR-378a Silencing Activity toward Its Oncogenic Targets
4. Discussion
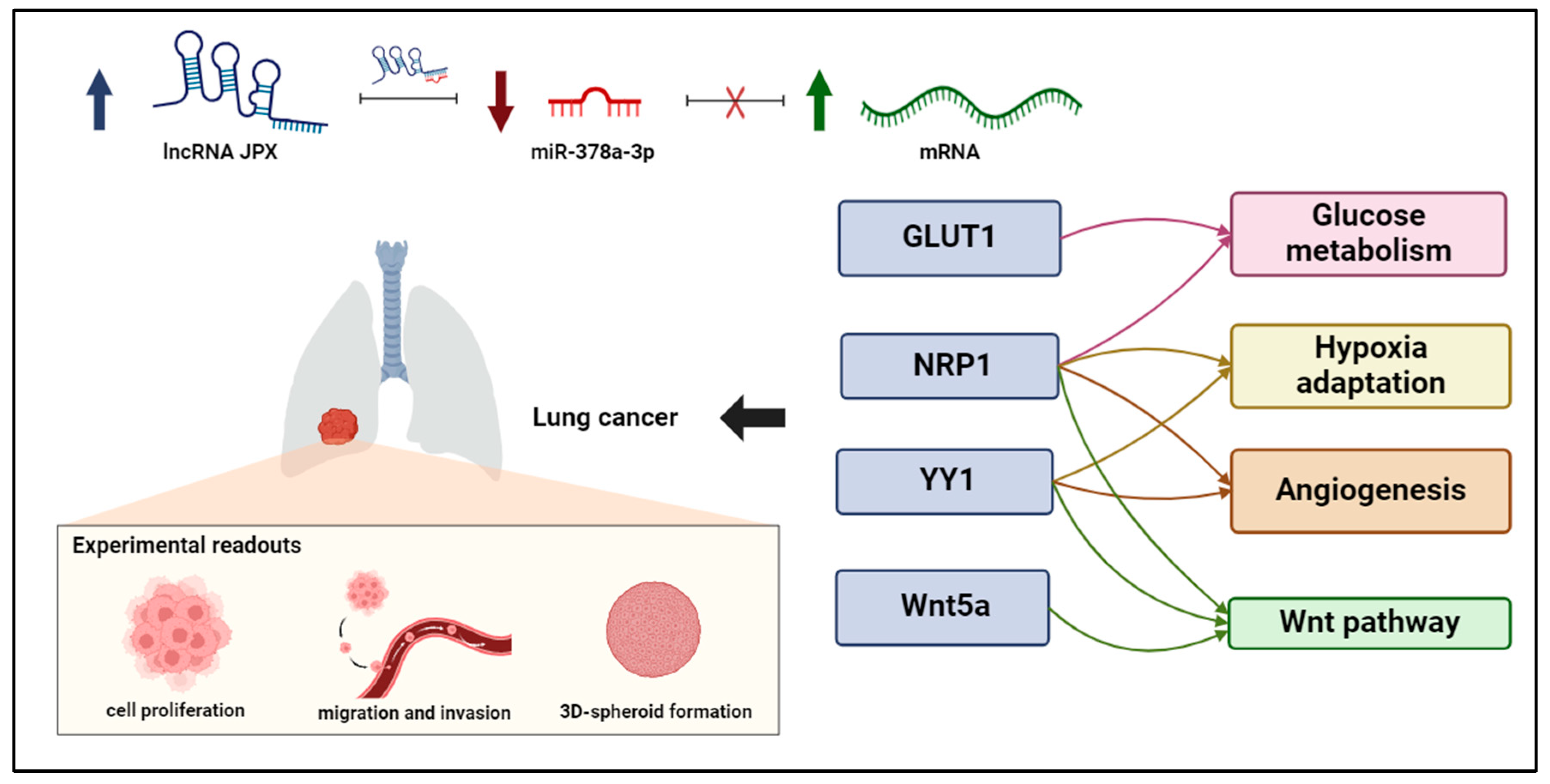
5. Conclusions
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Thai, A.A.; Solomon, B.J.; Sequist, L.V.; Gainor, J.F.; Heist, R.S. Lung Cancer. Lancet 2021, 398, 535–554. [Google Scholar] [CrossRef] [PubMed]
- Rudin, C.M.; Brambilla, E.; Faivre-Finn, C.; Sage, J. Small-Cell Lung Cancer. Nat. Rev. Dis. Primers 2021, 7, 3. [Google Scholar] [CrossRef] [PubMed]
- Swanton, C.; Govindan, R. Clinical Implications of Genomic Discoveries in Lung Cancer. N. Engl. J. Med. 2016, 374, 1864–1873. [Google Scholar] [CrossRef]
- Gilyazova, I.; Gimalova, G.; Nizamova, A.; Galimova, E.; Ishbulatova, E.; Pavlov, V.; Khusnutdinova, E. Non-Coding RNAs as Key Regulators in Lung Cancer. Int. J. Mol. Sci. 2024, 25, 560. [Google Scholar] [CrossRef]
- Fatima, R.; Akhade, V.S.; Pal, D.; Rao, S.M. Long Noncoding RNAs in Development and Cancer: Potential Biomarkers and Therapeutic Targets. Mol. Cell Ther. 2015, 3, 5. [Google Scholar] [CrossRef]
- Le, P.; Romano, G.; Nana-Sinkam, P.; Acunzo, M. Non-Coding RNAs in Cancer Diagnosis and Therapy: Focus on Lung Cancer. Cancers 2021, 13, 1372. [Google Scholar] [CrossRef]
- Chen, B.; Dragomir, M.P.; Yang, C.; Li, Q.; Horst, D.; Calin, G.A. Targeting Non-Coding RNAs to Overcome Cancer Therapy Resistance. Signal Transduct. Target. Ther. 2022, 7, 121. [Google Scholar] [CrossRef]
- Chang, T.-C.; Mendell, J.T. microRNAs in Vertebrate Physiology and Human Disease. Annu. Rev. Genom. Hum. Genet. 2007, 8, 215–239. [Google Scholar] [CrossRef]
- Lewis, B.P.; Burge, C.B.; Bartel, D.P. Conserved Seed Pairing, Often Flanked by Adenosines, Indicates that Thousands of Human Genes Are microRNA Targets. Cell 2005, 120, 15–20. [Google Scholar] [CrossRef]
- Mosca, N.; Russo, A.; Potenza, N. Making Sense of Antisense lncRNAs in Hepatocellular Carcinoma. Int. J. Mol. Sci. 2023, 24, 8886. [Google Scholar] [CrossRef]
- Salmena, L.; Poliseno, L.; Tay, Y.; Kats, L.; Pandolfi, P.P. A ceRNA Hypothesis: The Rosetta Stone of a Hidden RNA Language? Cell 2011, 146, 353–358. [Google Scholar] [CrossRef] [PubMed]
- Tay, Y.; Rinn, J.; Pandolfi, P.P. The Multilayered Complexity of ceRNA Crosstalk and Competition. Nature 2014, 505, 344–352. [Google Scholar] [CrossRef] [PubMed]
- Chan, J.J.; Tay, Y. Noncoding RNA: RNA Regulatory Networks in Cancer. Int. J. Mol. Sci. 2018, 19, 1310. [Google Scholar] [CrossRef] [PubMed]
- Di Palo, A.; Siniscalchi, C.; Mosca, N.; Russo, A.; Potenza, N. A Novel ceRNA Regulatory Network Involving the Long Non-Coding Antisense RNA SPACA6P-AS, miR-125a and Its mRNA Targets in Hepatocarcinoma Cells. Int. J. Mol. Sci. 2020, 21, 5068. [Google Scholar] [CrossRef] [PubMed]
- Siniscalchi, C.; Di Palo, A.; Russo, A.; Potenza, N. The lncRNAs at X Chromosome Inactivation Center: Not Just a Matter of Sex Dosage Compensation. Int. J. Mol. Sci. 2022, 23, 611. [Google Scholar] [CrossRef] [PubMed]
- Tian, D.; Sun, S.; Lee, J.T. The Long Noncoding RNA, Jpx, Is a Molecular Switch for X Chromosome Inactivation. Cell 2010, 143, 390–403. [Google Scholar] [CrossRef] [PubMed]
- Han, X.; Liu, Z. Long Non-coding RNA JPX Promotes Gastric Cancer Progression by Regulating CXCR6 and Autophagy via Inhibiting miR-197. Mol. Med. Rep. 2021, 23, 60. [Google Scholar] [CrossRef] [PubMed]
- Yao, Y.; Chen, S.; Lu, N.; Yin, Y.; Liu, Z. LncRNA JPX Overexpressed in Oral Squamous Cell Carcinoma Drives Malignancy via miR-944/CDH2 Axis. Oral. Dis. 2021, 27, 924–933. [Google Scholar] [CrossRef]
- Pan, J.; Fang, S.; Tian, H.; Zhou, C.; Zhao, X.; Tian, H.; He, J.; Shen, W.; Meng, X.; Jin, X.; et al. lncRNA JPX/miR-33a-5p/Twist1 Axis Regulates Tumorigenesis and Metastasis of Lung Cancer by Activating Wnt/β-Catenin Signaling. Mol. Cancer 2020, 19, 9. [Google Scholar] [CrossRef]
- Jin, M.; Ren, J.; Luo, M.; You, Z.; Fang, Y.; Han, Y.; Li, G.; Liu, H. Long Non-Coding RNA JPX Correlates with Poor Prognosis and Tumor Progression in Non-Small-Cell Lung Cancer by Interacting with miR-145-5p and CCND2. Carcinogenesis 2020, 41, 634–645. [Google Scholar] [CrossRef]
- Li, G.; Li, X.; Yuan, C.; Zhou, C.; Li, X.; Li, J.; Guo, B. Long Non-Coding RNA JPX Contributes to Tumorigenesis by Regulating miR-5195-3p/VEGFA in Non-Small Cell Lung Cancer. Cancer Manag. Res. 2021, 13, 1477–1489. [Google Scholar] [CrossRef]
- Di Palo, A.; Siniscalchi, C.; Polito, R.; Nigro, E.; Russo, A.; Daniele, A.; Potenza, N. microRNA-377-3p Downregulates the Oncosuppressor T-Cadherin in Colorectal Adenocarcinoma Cells. Cell Biol. Int. 2021, 45, 1797–1803. [Google Scholar] [CrossRef]
- Chen, X.; Yang, J.; Wang, Y. LncRNA JPX Promotes Cervical Cancer Progression by Modulating miR-25-3p/SOX4 Axis. Cancer Cell Int. 2020, 20, 441. [Google Scholar] [CrossRef]
- Cannarella, R.; Condorelli, R.A.; La Vignera, S.; Bellucci, C.; Luca, G.; Calafiore, R.; Calogero, A.E. IGF2 and IGF1R mRNAs Are Detectable in Human Spermatozoa. World J. Mens. Health 2020, 38, 545–551. [Google Scholar] [CrossRef]
- Xia, H.; Zhang, Z.; Yuan, J.; Niu, Q. The lncRNA PVT1 Promotes Invasive Growth of Lung Adenocarcinoma Cells by Targeting miR-378c to Regulate SLC2A1 Expression. Hum. Cell 2021, 34, 201–210. [Google Scholar] [CrossRef]
- Zhou, Q.; Feng, J.; Yin, S.; Ma, S.; Wang, J.; Yi, H. LncRNA FAM230B Promotes the Metastasis of Papillary Thyroid Cancer by Sponging the miR-378a-3p/WNT5A Axis. Biochem. Biophys. Res. Commun. 2021, 546, 83–89. [Google Scholar] [CrossRef]
- Rong, D.; Dong, Q.; Qu, H.; Deng, X.; Gao, F.; Li, Q.; Sun, P. m6A-Induced LINC00958 Promotes Breast Cancer Tumorigenesis via the miR-378a-3p/YY1 Axis. Cell Death Discov. 2021, 7, 27. [Google Scholar] [CrossRef]
- Niu, M.; Shan, M.; Liu, Y.; Song, Y.; Han, J.-G.; Sun, S.; Liang, X.-S.; Zhang, G.-Q. DCTPP1, an Oncogene Regulated by miR-378a-3p, Promotes Proliferation of Breast Cancer via DNA Repair Signaling Pathway. Front. Oncol. 2021, 11, 641931. [Google Scholar] [CrossRef] [PubMed]
- Ikeda, K.; Horie-Inoue, K.; Ueno, T.; Suzuki, T.; Sato, W.; Shigekawa, T.; Osaki, A.; Saeki, T.; Berezikov, E.; Mano, H.; et al. miR-378a-3p Modulates Tamoxifen Sensitivity in Breast Cancer MCF-7 Cells through Targeting GOLT1A. Sci. Rep. 2015, 5, 13170. [Google Scholar] [CrossRef] [PubMed]
- Potenza, N.; Mosca, N.; Zappavigna, S.; Castiello, F.; Panella, M.; Ferri, C.; Vanacore, D.; Giordano, A.; Stiuso, P.; Caraglia, M.; et al. MicroRNA-125a-5p Is a Downstream Effector of Sorafenib in Its Antiproliferative Activity Toward Human Hepatocellular Carcinoma Cells. J. Cell Physiol. 2017, 232, 1907–1913. [Google Scholar] [CrossRef] [PubMed]
- Kalitin, N.; Dudina, G.; Kostritsa, N.; Sivirinova, A.; Karamysheva, A. Evaluation of VEGF and VEGFR Gene Expression as Prognostic Markers in Low and Intermediate-1 Risk Patients with Myelodysplastic Syndromes. Oncol. Lett. 2023, 25, 95. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Luo, M. NORAD-Sponged miR-378c Alleviates Malignant Behaviors of Stomach Adenocarcinoma via Targeting NRP1. Cancer Cell Int. 2022, 22, 79. [Google Scholar] [CrossRef] [PubMed]
- Li, J.-H.; Liu, S.; Zhou, H.; Qu, L.-H.; Yang, J.-H. starBase v2.0: Decoding miRNA-ceRNA, miRNA-ncRNA and Protein-RNA Interaction Networks from Large-Scale CLIP-Seq Data. Nucleic Acids Res. 2014, 42, D92–D97. [Google Scholar] [CrossRef] [PubMed]
- Krüger, J.; Rehmsmeier, M. RNAhybrid: microRNA Target Prediction Easy, Fast and Flexible. Nucleic Acids Res. 2006, 34, W451–W454. [Google Scholar] [CrossRef] [PubMed]
- Kuang, Y.; Shen, W.; Zhu, H.; Huang, H.; Zhou, Q.; Yin, W.; Zhou, Y.; Cao, Y.; Wang, L.; Li, X.; et al. The Role of lncRNA Just Proximal to XIST (JPX) in Human Disease Phenotypes and RNA Methylation: The Novel Biomarker and Therapeutic Target Potential. Biomed. Pharmacother. 2022, 155, 113753. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Bai, H.; Jiang, M.; Zhou, C.; Gong, Z. Emerging Role of Long Non-Coding RNA JPX in Malignant Processes and Potential Applications in Cancers. Chin. Med. J. 2023, 136, 757–766. [Google Scholar] [CrossRef]
- Garnique, A.D.M.B.; Machado-Santelli, G.M. Characterization of 3D NSCLC Cell Cultures with Fibroblasts or Macrophages for Tumor Microenvironment Studies and Chemotherapy Screening. Cells 2023, 12, 2790. [Google Scholar] [CrossRef]
- Roman, V.; Mihaila, M.; Radu, N.; Marineata, S.; Diaconu, C.C.; Bostan, M. Cell Culture Model Evolution and Its Impact on Improving Therapy Efficiency in Lung Cancer. Cancers 2023, 15, 4996. [Google Scholar] [CrossRef]
- Wang, Y.; Du, J. miR-378a-3p Regulates Glioma Cell Chemosensitivity to Cisplatin through IGF1R. Open Life Sci. 2021, 16, 1175–1181. [Google Scholar] [CrossRef]
- Lin, Z.; Xia, S.; Liang, Y.; Ji, L.; Pan, Y.; Jiang, S.; Wan, Z.; Tao, L.; Chen, J.; Lin, C.; et al. LXR Activation Potentiates Sorafenib Sensitivity in HCC by Activating microRNA-378a Transcription. Theranostics 2020, 10, 8834–8850. [Google Scholar] [CrossRef] [PubMed]
- Megiorni, F.; Cialfi, S.; McDowell, H.P.; Felsani, A.; Camero, S.; Guffanti, A.; Pizer, B.; Clerico, A.; De Grazia, A.; Pizzuti, A.; et al. Deep Sequencing the microRNA Profile in Rhabdomyosarcoma Reveals Down-Regulation of miR-378 Family Members. BMC Cancer 2014, 14, 880. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhang, X.; Wang, Z.; Hu, Q.; Wu, J.; Li, Y.; Ren, X.; Wu, T.; Tao, X.; Chen, X.; et al. LncRNA-P23154 Promotes the Invasion-Metastasis Potential of Oral Squamous Cell Carcinoma by Regulating Glut1-Mediated Glycolysis. Cancer Lett. 2018, 434, 172–183. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Yu, J.; Tian, H.; Shan, Z.; Liu, W.; Pan, Z.; Ren, J. Circle RNA hsa_circRNA_100290 Serves as a ceRNA for miR-378a to Regulate Oral Squamous Cell Carcinoma Cells Growth via Glucose Transporter-1 (GLUT1) and Glycolysis. J. Cell Physiol. 2019, 234, 19130–19140. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Wang, R.; Li, M.; Yang, Q. Long Non-coding RNA BLACAT2/miR-378a-3p/YY1 Feedback Loop Promotes the Proliferation, Migration and Invasion of Uterine Corpus Endometrial Carcinoma. Oncol. Rep. 2023, 49, 108. [Google Scholar] [CrossRef] [PubMed]
- Fu, H.; Zhang, J.; Pan, T.; Ai, S.; Tang, L.; Wang, F. miR-378a Enhances the Sensitivity of Liver Cancer to Sorafenib by Targeting VEGFR, PDGFRβ and c-Raf. Mol. Med. Rep. 2018, 17, 4581–4588. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Luo, X.; Terp, M.G.; Li, Q.; Li, Y.; Shen, L.; Chen, Y.; Jacobsen, K.; Bivona, T.G.; Chen, H.; et al. DDX56 Modulates Post-Transcriptional Wnt Signaling through miRNAs and Is Associated with Early Recurrence in Squamous Cell Lung Carcinoma. Mol. Cancer 2021, 20, 108. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Sun, X.; Yang, Y.; Jiao, W. Long Non-Coding RNA OIP5-AS1 Promotes Proliferation of Lung Cancer Cells and Leads to Poor Prognosis by Targeting miR-378a-3p. Thorac. Cancer 2018, 9, 939–949. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Chai, B.; Wang, X.; Wu, Z.; Gu, Z.; Liu, X.; Zhao, Y.; Chen, T.; Ma, Z.; Sun, Q. miRNA-199a-5p/SLC2A1 Axis Regulates Glucose Metabolism in Non-Small Cell Lung Cancer. J. Cancer 2022, 13, 2352–2361. [Google Scholar] [CrossRef]
- Wang, Y.-B.; Zheng, K.-W.; Hu, Y.-Y.; Salameen, H.; Zhu, Z.-Y.; Wu, F.-F.; Ding, X. VEGF/Nrp1/HIF-1α Promotes Proliferation of Hepatocellular Carcinoma through a Positive Feedback Loop. Med. Oncol. 2023, 40, 339. [Google Scholar] [CrossRef]
- Wu, S.; Kasim, V.; Kano, M.R.; Tanaka, S.; Ohba, S.; Miura, Y.; Miyata, K.; Liu, X.; Matsuhashi, A.; Chung, U.; et al. Transcription Factor YY1 Contributes to Tumor Growth by Stabilizing Hypoxia Factor HIF-1α in a P53-Independent Manner. Cancer Res. 2013, 73, 1787–1799. [Google Scholar] [CrossRef]
- de Nigris, F.; Crudele, V.; Giovane, A.; Casamassimi, A.; Giordano, A.; Garban, H.J.; Cacciatore, F.; Pentimalli, F.; Marquez-Garban, D.C.; Petrillo, A.; et al. CXCR4/YY1 Inhibition Impairs VEGF Network and Angiogenesis during Malignancy. Proc. Natl. Acad. Sci. USA 2010, 107, 14484–14489. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.; Zheng, M.; Yang, Y.; Chen, Y.; Zhang, X.; Zhu, L.; Zhang, H. Knockdown of lncRNA ACTA2-AS1 Reverses Cisplatin Resistance of Ovarian Cancer Cells via Inhibition of miR-378a-3p-Regulated Wnt5a. Bioengineered 2022, 13, 9829–9838. [Google Scholar] [CrossRef] [PubMed]
- Yu, Q.-Y.; Han, Y.; Lu, J.-H.; Sun, Y.-J.; Liao, X.-H. NRP1 Regulates Autophagy and Proliferation of Gastric Cancer through Wnt/β-Catenin Signaling Pathway. Aging 2023, 15, 8613–8629. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.; Li, X.; Wu, C.W.; Dong, Y.; Cai, M.; Mok, M.T.S.; Wang, H.; Chen, J.; Ng, S.S.M.; Chen, M.; et al. microRNA-7 Is a Novel Inhibitor of YY1 Contributing to Colorectal Tumorigenesis. Oncogene 2013, 32, 5078–5088. [Google Scholar] [CrossRef]
- Kang, W.; Tong, J.H.M.; Chan, A.W.H.; Zhao, J.; Dong, Y.; Wang, S.; Yang, W.; Sin, F.M.C.; Ng, S.S.M.; Yu, J.; et al. Yin Yang 1 Contributes to Gastric Carcinogenesis and Its Nuclear Expression Correlates with Shorter Survival in Patients with Early Stage Gastric Adenocarcinoma. J. Transl. Med. 2014, 12, 80. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene ID | TCGA Log2 Fold Change (p-Value) | GSE19188 Log2 Fold Change (p-Value) | GSE33532 Log2 Fold Change (p-Value) |
|---|---|---|---|
| JPX | 1.38 (2.0 × 10−7) | 1.45 (0.0001) | 1.21 (0.0103) |
| miR-378a-3p | 0.18 (5.2 × 10−19) | - | - |
| GLUT1 | 12.33 (1.8 × 10−45) | 1.36 (5.61 × 10−21) | 1.39 (3.44 × 10−11) |
| NRP1 | 1.02 (ns) | 0.93 (9.81 × 10−6) | 0.96 (2.59 × 10−3) |
| YY1 | 1.14 (0.0014) | 1.04 (6.76 × 10−11) | 1.06 (9.10 × 10−13) |
| Wnt5a | 1.19 (ns) | 1.00 (ns) | 1.06 (0.012) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mosca, N.; Pezzullo, M.; De Leo, I.; Truda, A.; Marchese, G.; Russo, A.; Potenza, N. A Novel ceRNET Relying on the lncRNA JPX, miR-378a-3p, and Its mRNA Targets in Lung Cancer. Cancers 2024, 16, 1526. https://doi.org/10.3390/cancers16081526
Mosca N, Pezzullo M, De Leo I, Truda A, Marchese G, Russo A, Potenza N. A Novel ceRNET Relying on the lncRNA JPX, miR-378a-3p, and Its mRNA Targets in Lung Cancer. Cancers. 2024; 16(8):1526. https://doi.org/10.3390/cancers16081526
Chicago/Turabian StyleMosca, Nicola, Mariaceleste Pezzullo, Ilenia De Leo, Anna Truda, Giovanna Marchese, Aniello Russo, and Nicoletta Potenza. 2024. "A Novel ceRNET Relying on the lncRNA JPX, miR-378a-3p, and Its mRNA Targets in Lung Cancer" Cancers 16, no. 8: 1526. https://doi.org/10.3390/cancers16081526
APA StyleMosca, N., Pezzullo, M., De Leo, I., Truda, A., Marchese, G., Russo, A., & Potenza, N. (2024). A Novel ceRNET Relying on the lncRNA JPX, miR-378a-3p, and Its mRNA Targets in Lung Cancer. Cancers, 16(8), 1526. https://doi.org/10.3390/cancers16081526