Protein Kinase C Epsilon and Genetic Networks in Osteosarcoma Metastasis


Abstract
:1. Introduction
2. Results
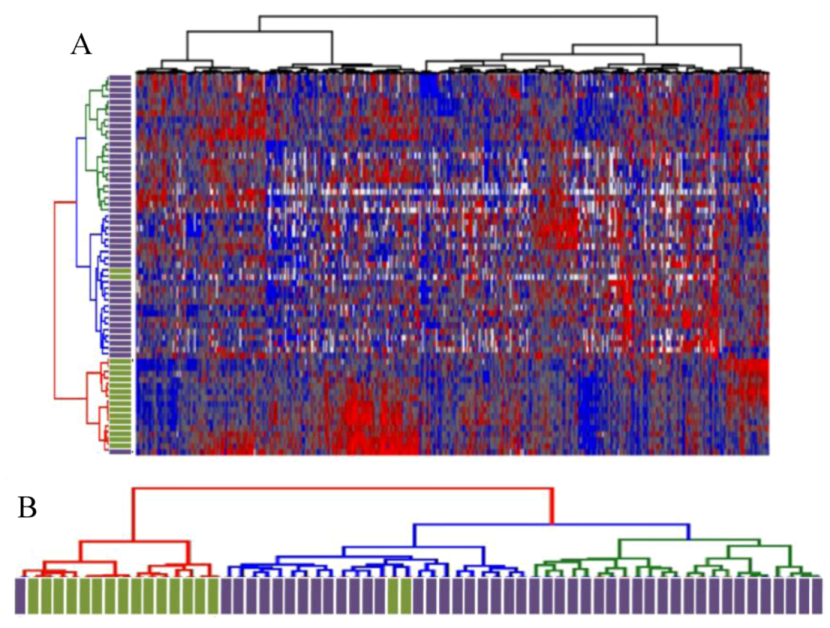
2.1. Unsupervised Hierarchical Clustering of Expression Profiles Reveals Distinct Subtypes of Osteosarcomas

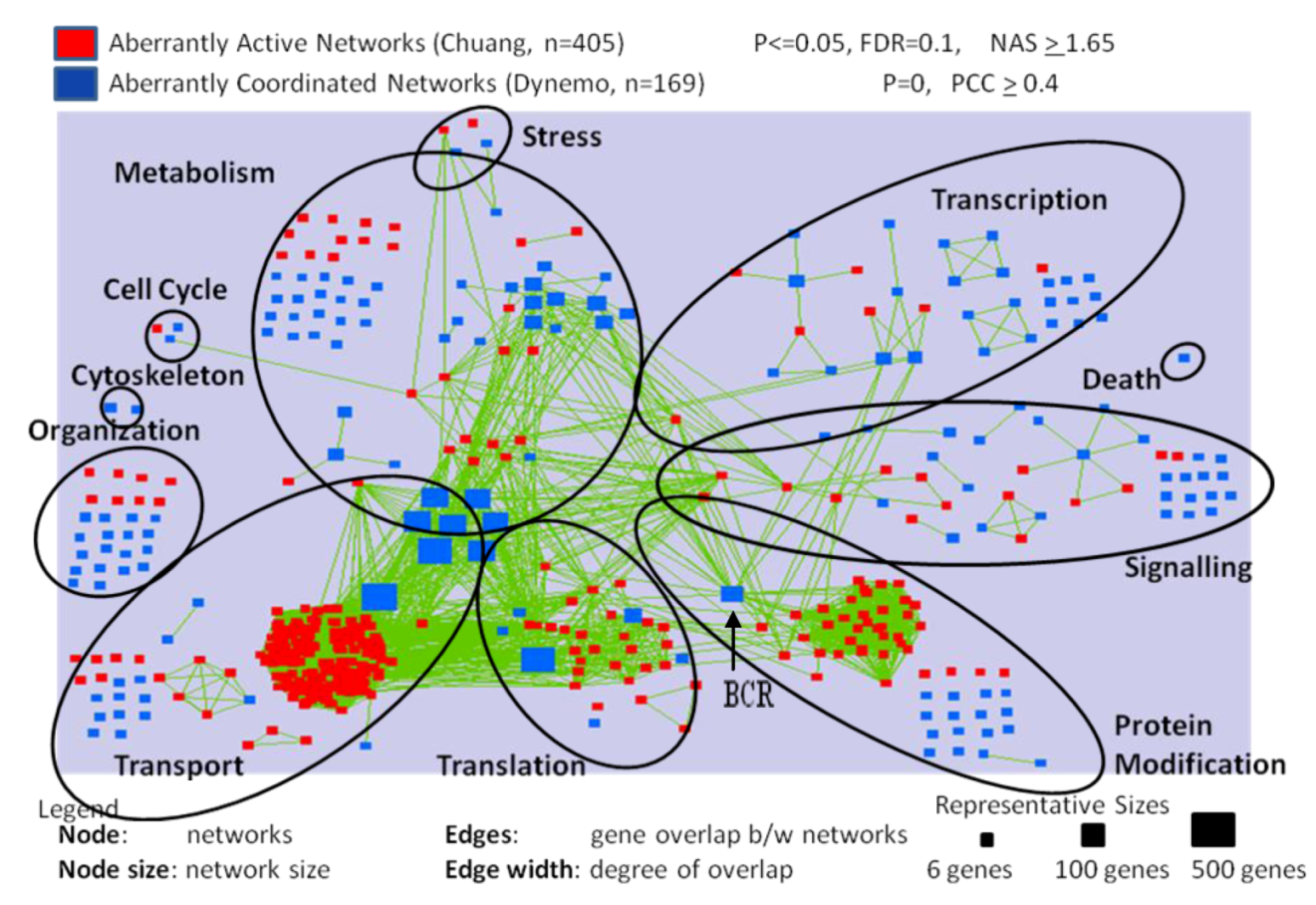
2.2. Differentially Activated and Organized Networks in Metastatic Osteosarcomas

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
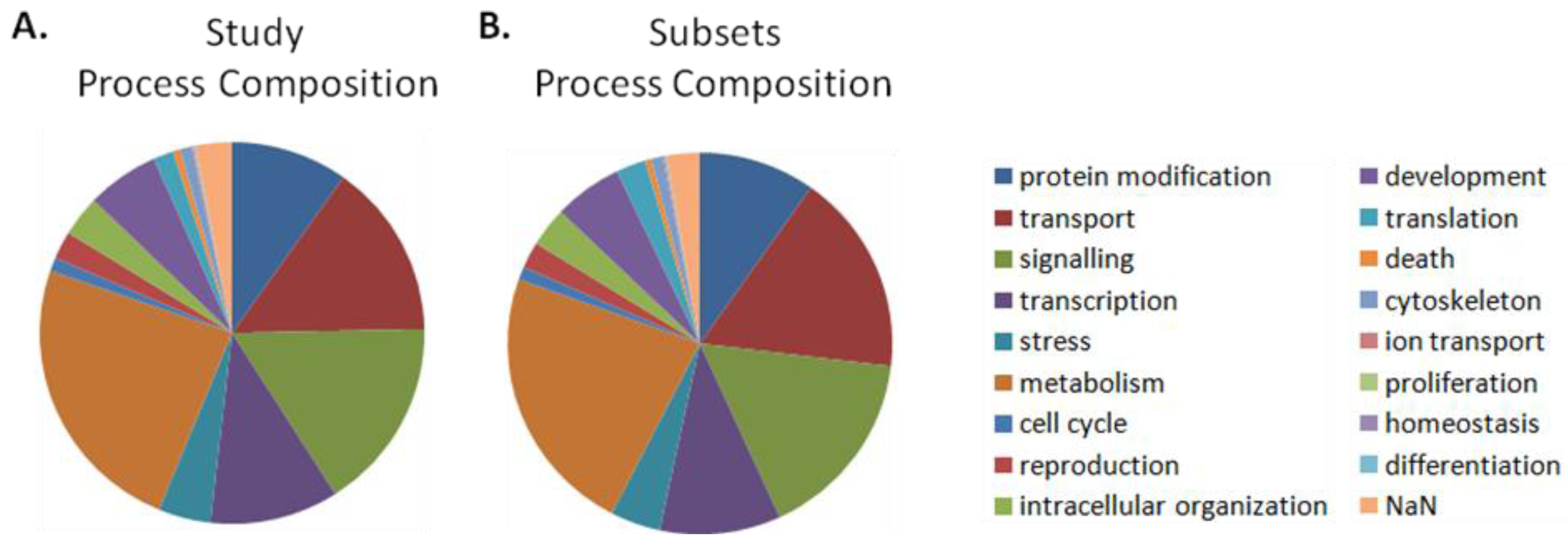
| Cellular Processes | All Networks in Study | Significant Networks: Permissive Cutoffs | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Subsets | Entire Networks | Differentially Activated | Differentially Organized | |||||||
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 |
| # | % of all | # | % of all | # | % of all | p-value | # | % of all | p-value | |
| protein modification | 573 | 9.8 | 575 | 9.8 | 52 | 10.5 * | 5.3E-02 | 49 | 7.2 | |
| transport | 1002 | 17.1 | 869 | 14.8 | 194 | 39 * | 1.1E-34 Ϯ | 133 | 19.5 * | 9.0E-05 Ϯ |
| signaling | 954 | 16.3 | 955 | 16.3 | 69 | 13.9 | 87 | 12.7 | ||
| transcription | 595 | 10.2 | 635 | 10.8 | 17 | 3.4 | 48 | 7 | ||
| stress | 247 | 4.2 | 257 | 4.4 | 15 | 3 | 9 | 1.3 | ||
| metabolism | 1340 | 22.9 | 1410 | 24.1 | 58 | 11.7 | 275 | 40.3 * | 5.7E-24 Ϯ | |
| cell cycle | 65 | 1.1 | 68 | 1.2 | 6 | 1.2 * | 1.6E-01 | 7 | 1 | |
| reproduction | 130 | 2.2 | 137 | 2.3 | 6 | 1.2 | 0 | 0 | ||
| intracellular organization | 195 | 3.3 | 201 | 3.4 | 27 | 5.4 * | 3.4E-03 Ϯ | 54 | 7.9 * | 9.1E-10 Ϯ |
| development | 338 | 5.8 | 357 | 6.1 | 0 | 0 | 0 | 0 | ||
| translation | 146 | 2.5 | 99 | 1.7 | 50 | 10.1 * | 4.4E-19 Ϯ | 17 | 2.5 * | 3.9E-2 Ϯ |
| death | 31 | 0.5 | 36 | 0.6 | 0 | 0 | 3 | 0.4 | ||
| cytoskeleton | 57 | 1 | 59 | 1 | 0 | 0 | 0 | 0 | ||
| ion transport | 10 | 0.2 | 8 | 0.1 | 2 | 0.4 * | 1.6E-01 | 0 | 0 | |
| proliferation | 3 | 0.1 | 2 | 0 | 1 | 0.2 * | 2.1E-01 | 0 | 0 | |
| homeostasis | 2 | 0 | 2 | 0 | 0 | 0 | 0 | 0 | ||
| differentiation | 6 | 0.1 | 6 | 0.1 | 0 | 0 | 0 | 0 | ||
| NaN | 161 | 2.7 | 179 | 3.1 | 0 | 0 | 1 | 0.1 | ||
| Totals | 5855 | 100 | 5855 | 99.8 | 497 | 100 | 683 | 99.9 | ||

2.3. Genes Previously Implicated in Osteosarcoma Metastasis are among Significant Network Results
| Gene | Ref. | Gene | Ref. | Gene | Ref. | |
|---|---|---|---|---|---|---|
| BIRC5 | [16] | IVD | -- | CXCR3 | X | [17] |
| CAV1 | [18] | KIT | [19] | EGF | X | [20,21] |
| CCN1 | [22] | LRP5 | [23] | EGFR2 | X | [24,25,26,27,28,29] |
| CCN3 | [30] | MMP2 | [31] | FADD | X | [32] |
| CD44 | [33] | PDGFRA | [34,35] | HIF1 | X | [36] |
| CDH11 | [37] | PEDF | [38] | IL12A | X | [39] |
| CDH2 | [37] | RECK | [40] | IL12B | X | [39] |
| COL18A1 | [41] | S100A6 | [42] | INS | X | [43,44,45,46,47,48,49,50,51] |
| CXCR4 | [52,53,54,55] | SPARC | [56] | MAML1 | X | [57] |
| DPF2 | -- | TIMP1 | [58] | MIRK | X | [59] |
| EGFR | [60] | PLAUR | [61] | MMP14 | X | [62] |
| EZR | [25,63,64] | VEGFA | [55,65,66,67,68,69,70,71,72] | MMP9 | X | [58,73,74] |
| FAS | [32,75,76,77] | VEGFB | [55,65,66,67,68,69,70,71,72] | PDGFA | X | [35] |
| IGF1 | [43,44,45,46,47,48,49,50,51] | VEGFC | [55,65,66,67,68,69,70,71,72] | VCP | X | [78] |
| IGF2 | [43,44,45,46,47,48,49,50,51] | WIF1 | [79] | |||
| IGF1R | [43,44,45,46,47,48,49,50,51] | YYI | [80] |
| Query | Node | Score | Psample | FDRsample | Pgene | FDRgene |
|---|---|---|---|---|---|---|
| CDH2 | CDH2 | 3.89 | 0 | 0.0017 | 0.037 | 0.019 |
| S100A6 | CACYBP | 3.94 | 0 | 0.00096 | 0.007 | 0.18 |
| TIMP1 | ECH1 | 3.28 | 0 | 0.00096 | 0.003 | 0.17 |
| PLAUR | PGAP1 | 2.54 | 0 | 0.0019 | 0.003 | 0.19 |
| IVD | MECR | 3.94 | 0.001 | 0.01 | 0.046 | 0.0076 |
| Query | Node | ΔPCCTotal | Psample | Query | Node | ΔPCCTotal | Psample |
|---|---|---|---|---|---|---|---|
| S100A6 | S100A6 | 0.41 | 0 | TIMP1 | LRP1 | 0.32 | 0 |
| KIT | KIT | 0.34 | 0 | PLAUR | LRP1 | 0.32 | 0 |
| YY1 | YY1 | 0.32 | 0.001 | BIRC5 | PAFAH1B1 | 0.34 | 0 |
| BIRC5 | BIRC5 | 0.35 | 0.001 | NOV | GIA1 | 0.3 | 0 |
| MMP2 | MMP2 | 0.35 | 0.001 | VEGFA | SPARC | 0.34 | 0 |
| SPARC | SPARC | 0.34 | 0 | YY1 | HDAC | 0.33 | 0 |
| IGF1R | CAMK2B | 0.37 | 0 | IGF1 | PRKCD | 0.36 | 0 |
| IGF2 | CAMK2B | 0.37 | 0 | PDGFRA | JAK1 | 0.33 | 0 |
| EGFR | CAMK2B | 0.37 | 0 | CXCR4 | JAK1 | 0.33 | 0 |
| KIT | CAMK2B | 0.37 | 0 | FAS | BTK | 0.35 | 0 |
| CDH2 | CAMK2B | 0.37 | 0 | VEGFB | RASA1 | 0.32 | 0 |
| EZR | CAMK2B | 0.37 | 0 | MMP2 | ITGB2 | 0.37 | 0 |
| CD44 | CAMK2B | 0.37 | 0 | LRP5 | FZD8 | 0.48 | 0 |
| SPARC | CAMK2B | 0.37 | 0 | SERPINF1 | CSNK2A1 | 0.33 | 0.001 |
| CYR61 | ATP2A2 | 0.33 | 0 | COL18A1 | CTSL1 | 0.33 | 0.001 |
| CAV1 | ATP2A2 | 0.33 | 0 | WIF1 | FZD1 | 0.37 | 0.001 |
| S100A6 | ACTN1 | 0.32 | 0 | IVD | MCCC1 | 0.37 | 0 |
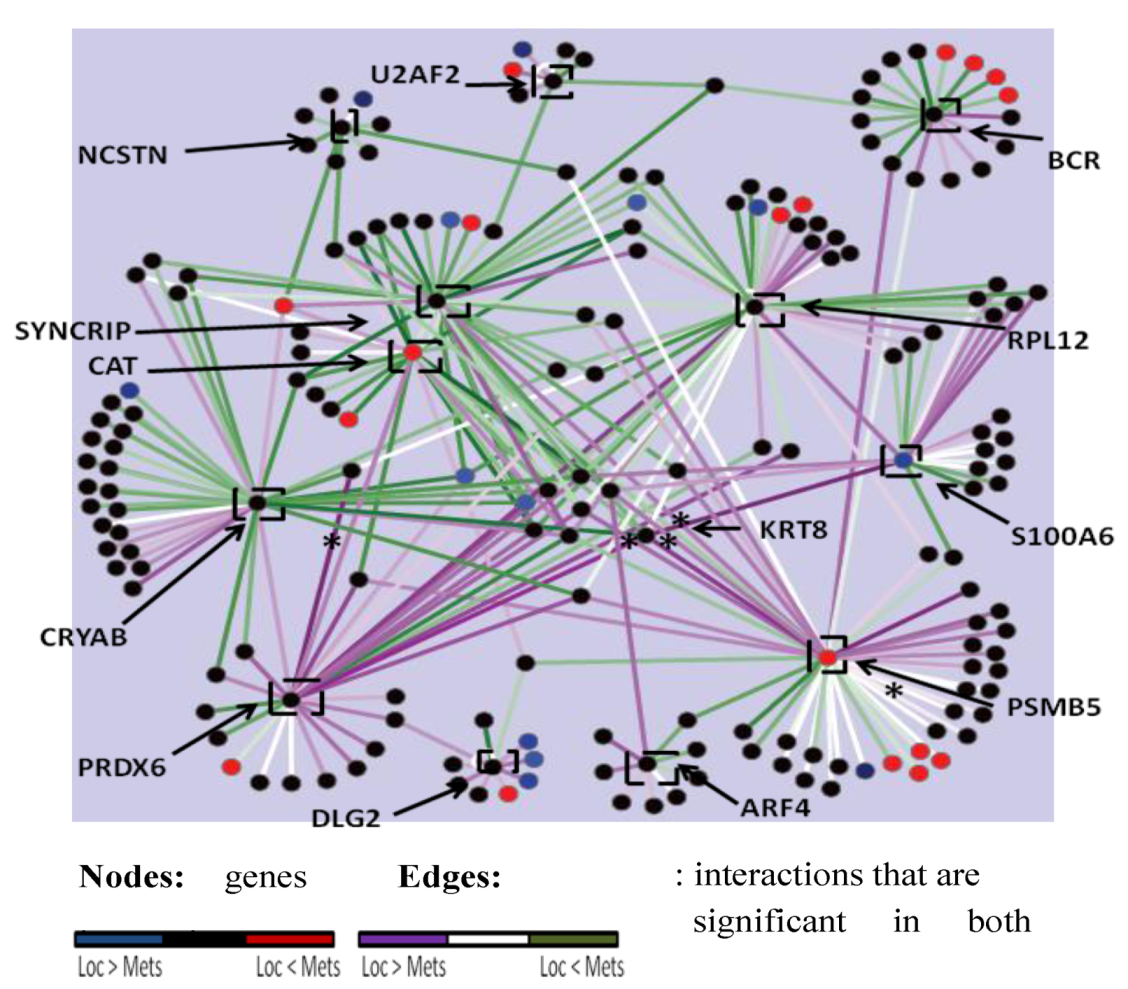
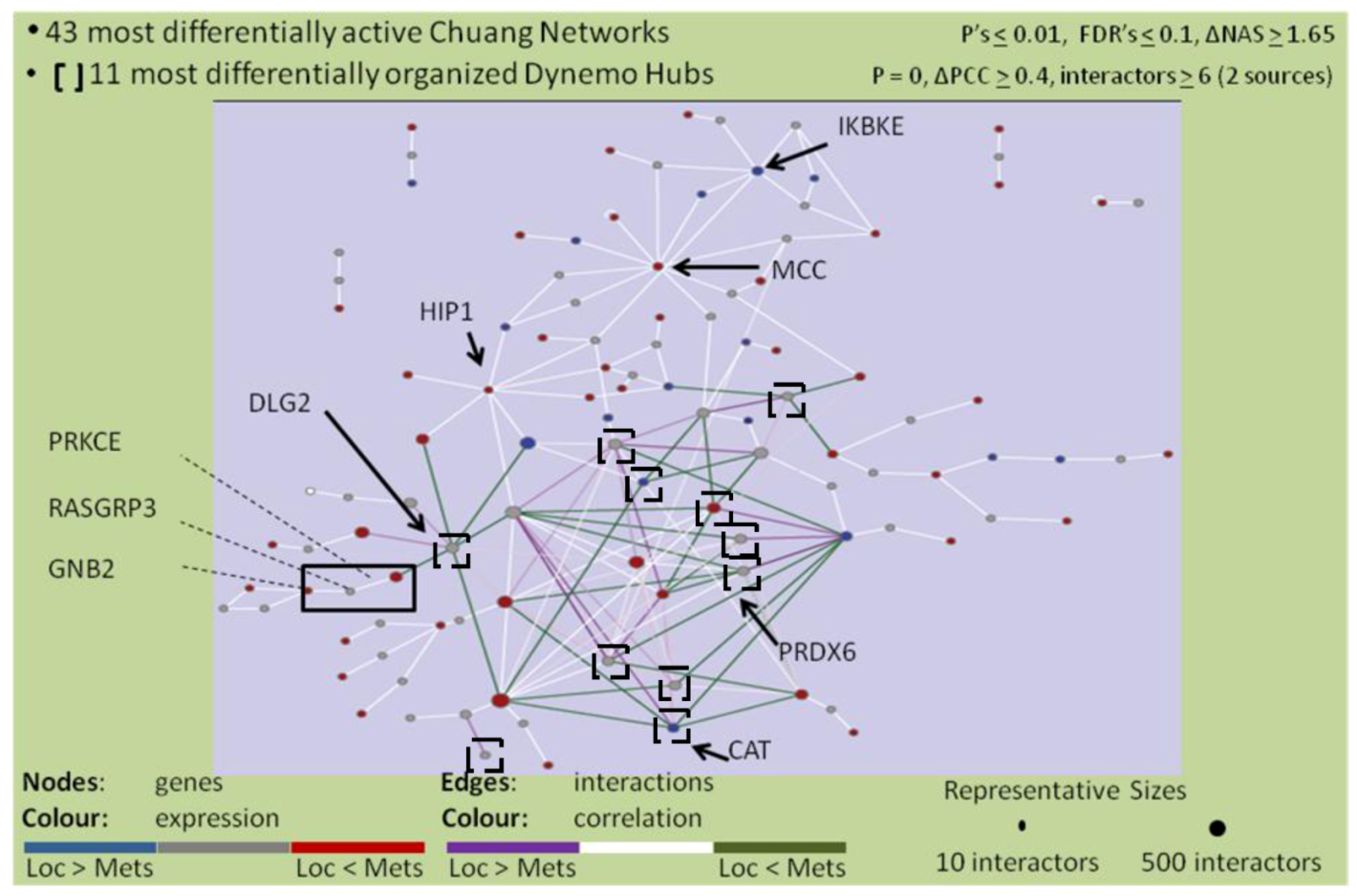
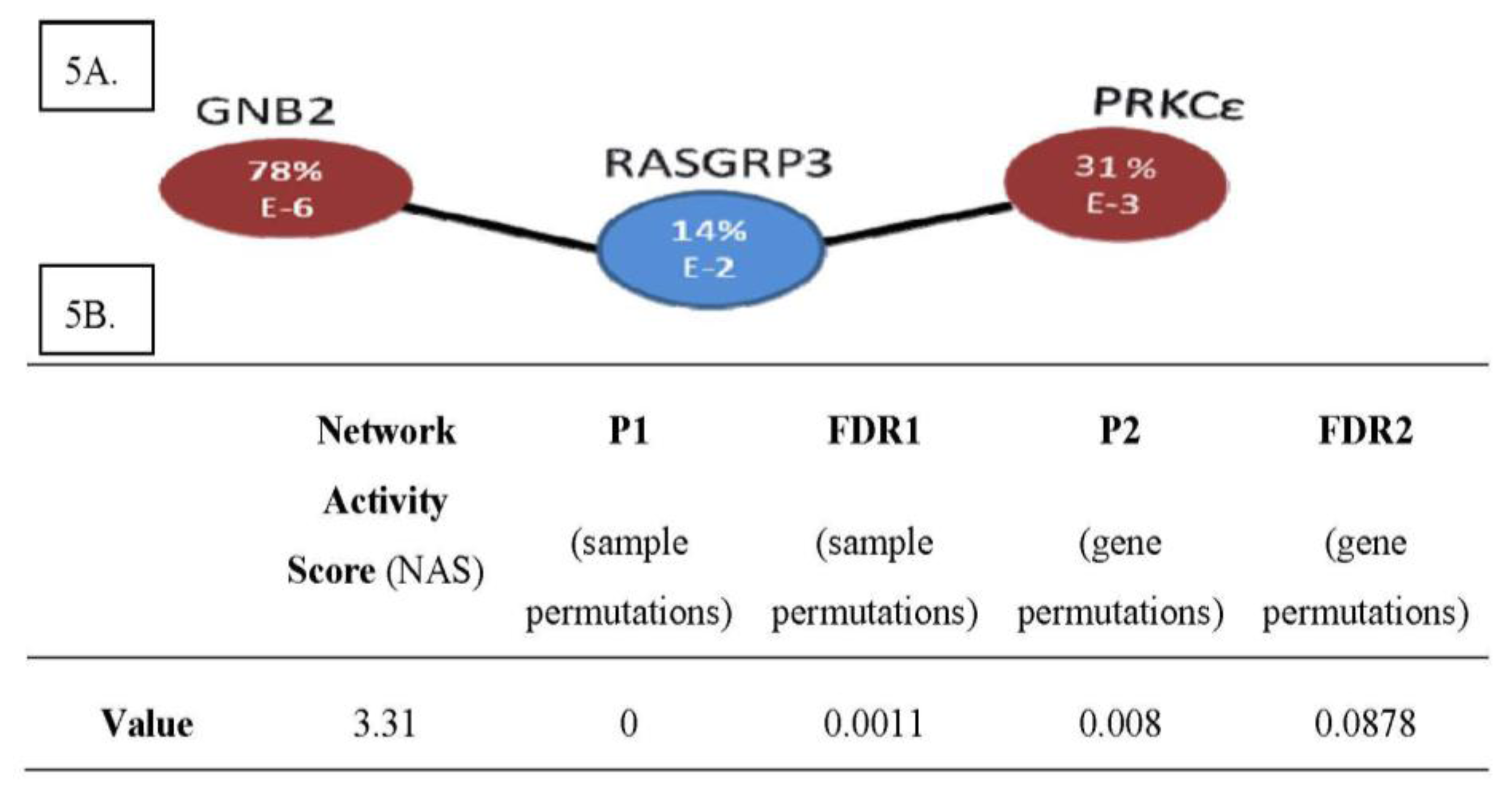
2.4. The PRKCε-RASGRP3-GNB2 Network Is Differentially Activated, and May Interact with the Disorganized DLG2 Hub



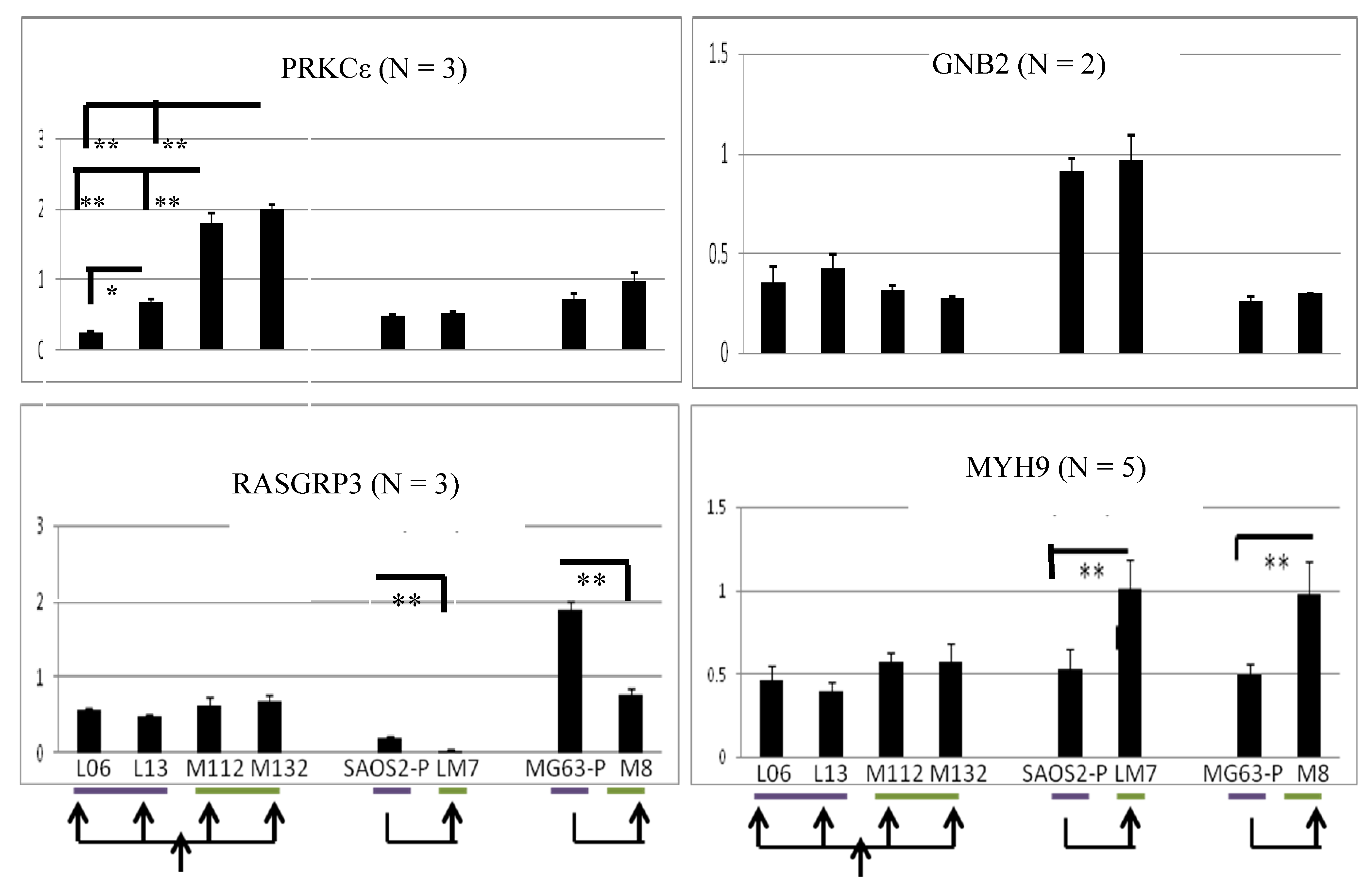
2.5. PRKCε-RASGRP3-GNB2 Network Is Differentially Activated in Vitro


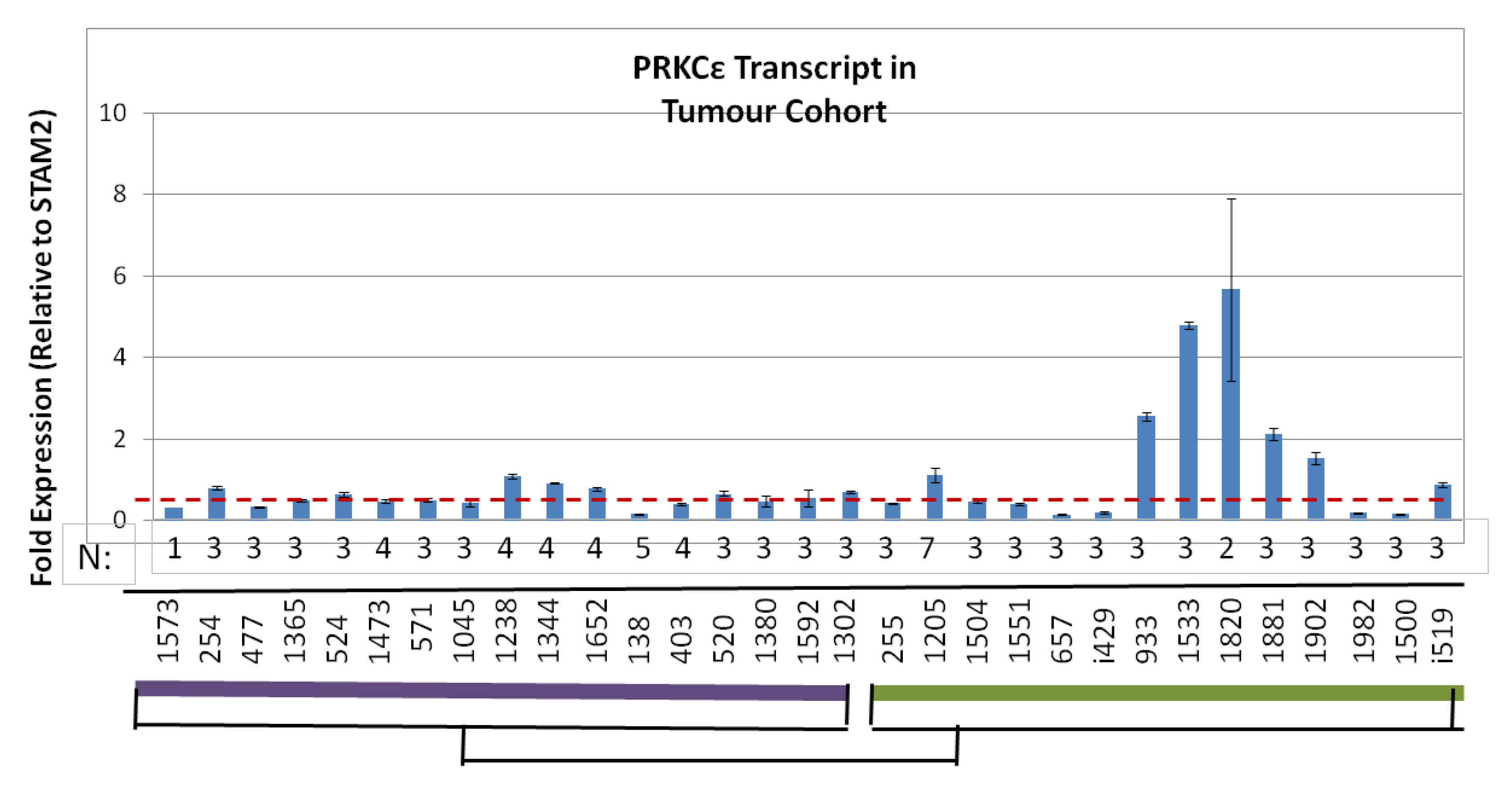
2.6. Human Osteosarcomas That Are Metastatic-at-Diagnosis Are More Likely to Exhibit High Levels of PRKCε mRNA

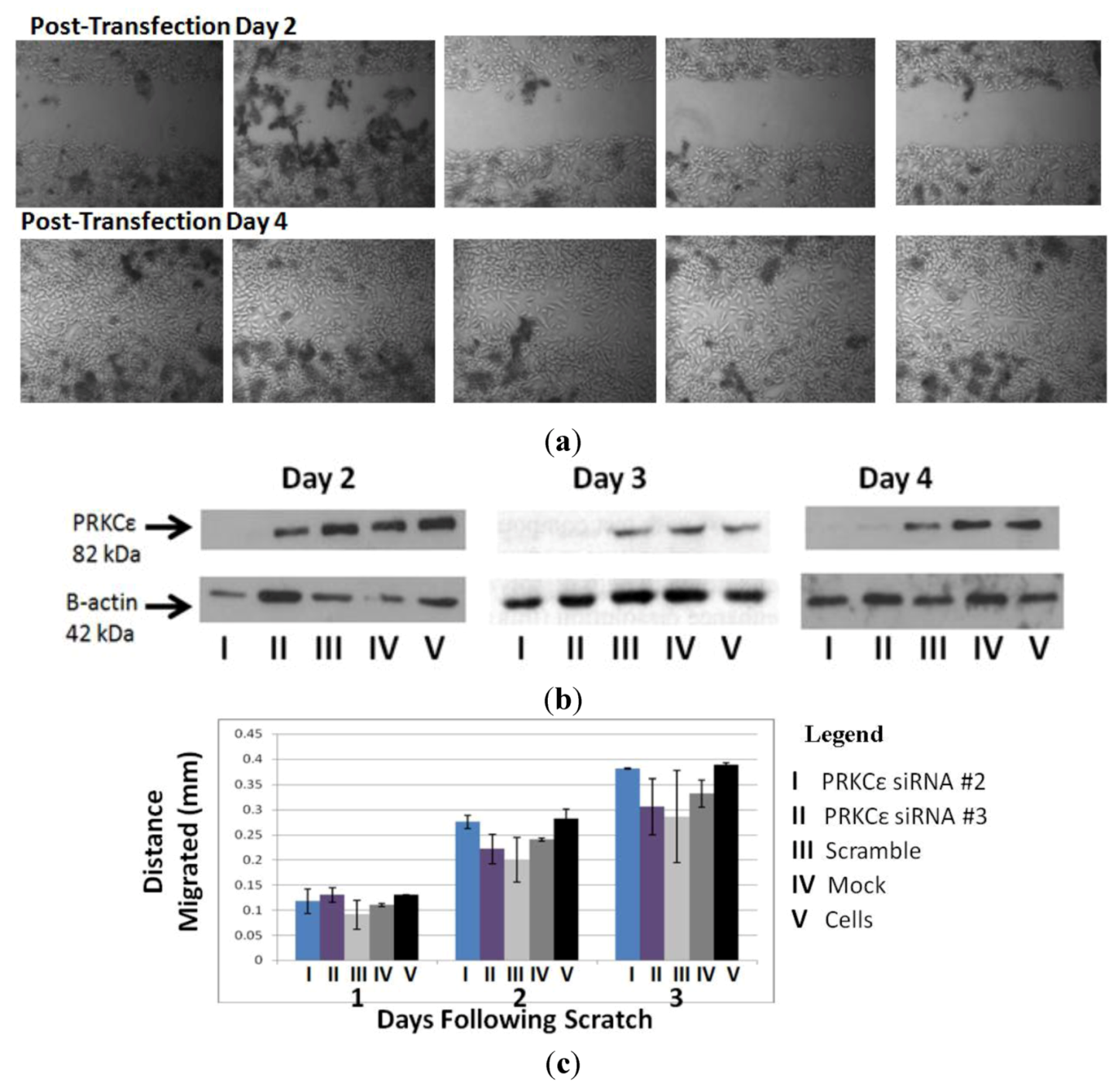
2.7. PRKCε Is Not Required for Migration of Highly Metastatic M132 Cells
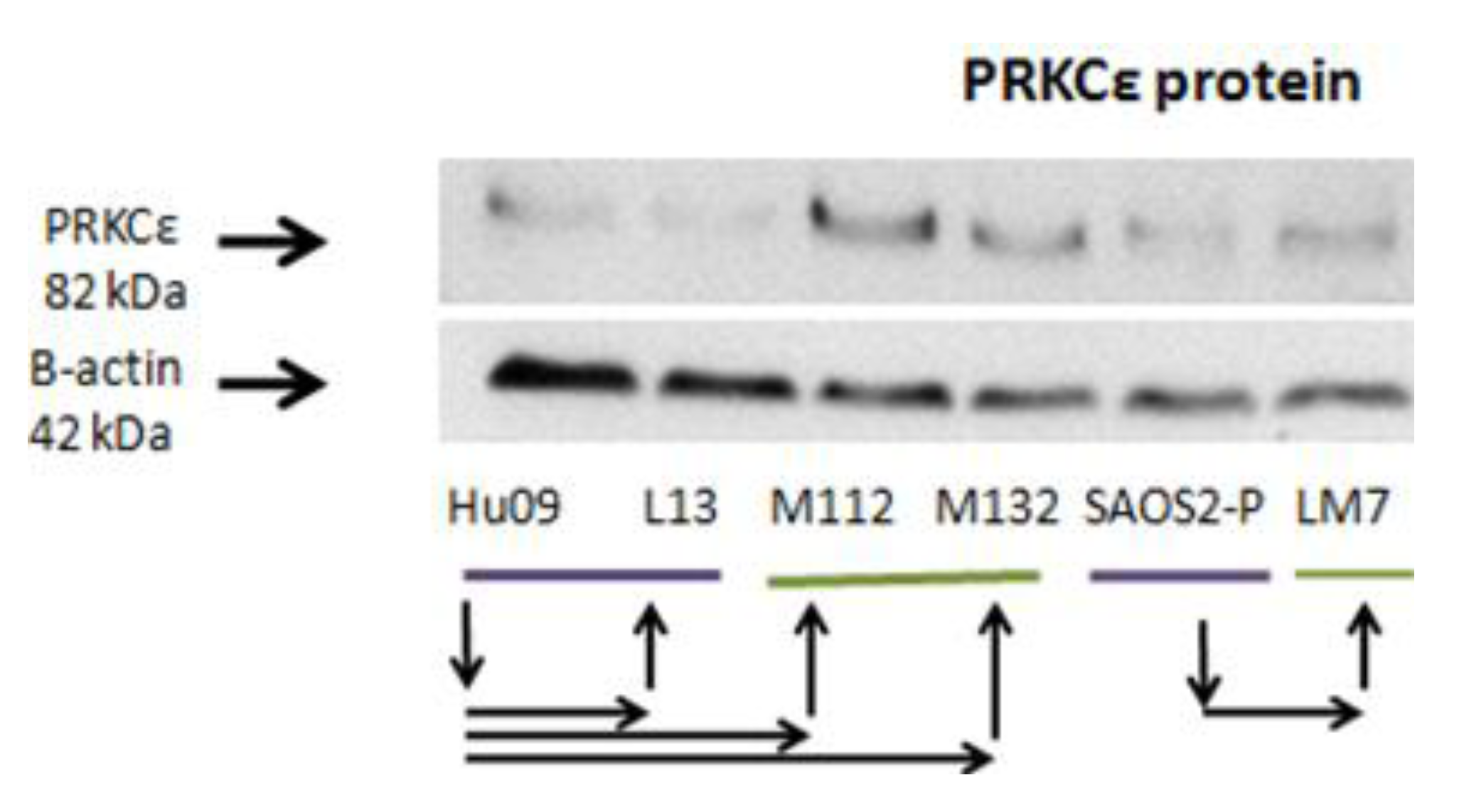
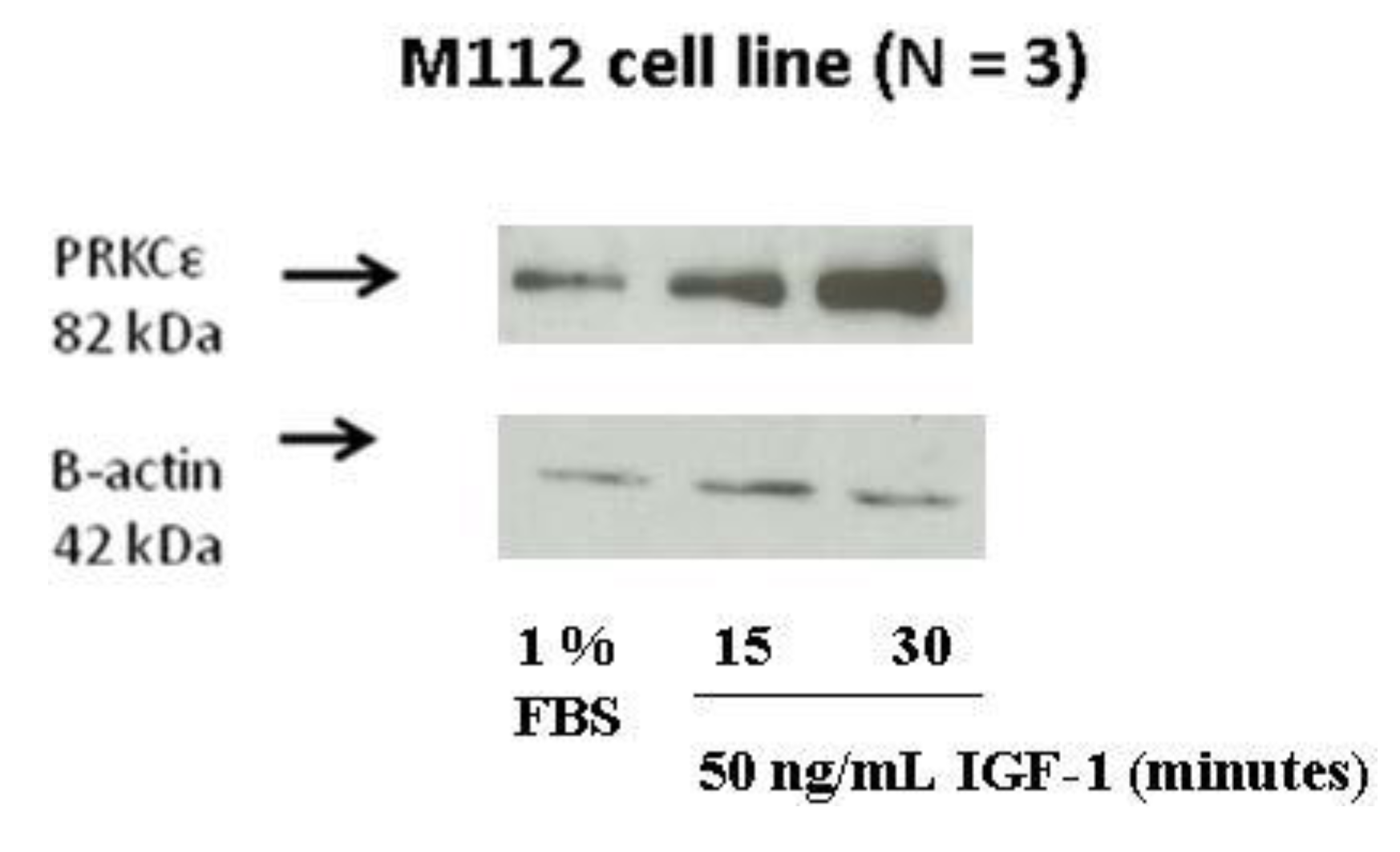
2.8. IGF-1 Stimulation Induces Protein Expression of PRKCε in M112 Osteosarcoma Cells


3. Discussion
4. Experimental Section
4.1. Patient Follow-Up
4.2. Tumor Samples
4.3. Gene Expression Profiling
4.4. Unsupervised Hierarchical Clustering
4.5. Supervised Network Analysis
4.5.1. Differentially Activated Networks
| Trial | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 |
|---|---|---|---|---|---|---|---|---|---|---|
| Differential Expression (%) | 0 | 0 | 10 | 20 | 30 | 40 | 50 | 60 | 70 | 80 |
| P Value Maximum | None (i.e., all genes included) | 0.001 | ||||||||





4.5.2. Differentially Organized Network—Dynemo




4.5.3. Visualization of Network Results
4.5.4. Cellular Process Annotation
| Original Terms | Further Simplified Terms |
|---|---|
| cell death, death | death |
| multicellular organismal development, embryonic development, anatomical structure morphogenesis | development |
| cell differentiation, differentiation | differentiation |
| regulation of gene expression, epigenetic | epigenetics |
| cell growth, growth | growth |
| cellular component organization, organelle organization, mitochondrion organization, cytoplasm organization | intracellular organization |
| metabolic process, cellular amino acid and derivative metabolic process, secondary metabolic process, lipid metabolic process, biosynthetic process, catabolic process, carbohydrate metabolic process, protein metabolic process, nucleobase nucleoside nucleotide and nucleic acid metabolic process, DNA metabolic process, generation of precursor metabolites and energy | metabolism |
| signal transduction, response to biotic stimulus, response to external stimulus, response to abiotic stimulus, cell-cell signaling, cell communication, response to endogenous stimulus, cell recognition | signaling |
| protein transport, transport | transport |
| regulation of biological process, biological process, behavior | NaN |

4.5.5. Cellular Process Enrichment

4.6. Cell Culture
4.7. Quantitative Reverse-Transcription Polymerase Chain Reaction (rt-PCR)
| Gene Symbol | Primer Pairs |
|---|---|
| STAM2 | Forward 5'-TGGATGACAGTGATGCCAATTG-3' |
| Reverse 5'-CGCTGCCTCAGTCTCTATGT-3' | |
| PRKCε | Forward 5'-CACTGCAAGCTGGCTGACT-3' |
| Reverse 5'-TGCAGGATCTCAGGAGCTATG-3' | |
| RASGRP3 | Forward 5'-GGATTTCTCTGGGGCATAATC-3' |
| Reverse 5'-AGGAGGTCTTTGCACTGTTTG-3' | |
| GNB2 | Forward 5'-CTATCAAGCTGTGGGACGTG-3' |
| Reverse 5'-GTAGCCGTTGGGGAAGAAAG-3' | |
| MYH9 | Forward 5'-GCCTACAGGAGTATGATGCAAG-3' |
| Reverse 5'-ACTGGATGACCTTCTTGGTGTT-3' |
4.8. Western Blots
4.9. Knockdown of PRKCε
4.10. Scratch Assay
4.11. IGF-1 Induction Assay
5. Conclusions
Acknowledgments
References
- Kansara, M.; Thomas, D.M. Molecular pathogenesis of osteosarcoma. DNA Cell Biol. 2007, 26, 1–18. [Google Scholar] [CrossRef]
- Murphey, M.D.; Robbin, M.R.; McRae, G.A.; Flemming, D.J.; Temple, H.T.; Kransdorf, M.J. The many faces of osteosarcoma. Radiographics 1997, 17, 1205–1231. [Google Scholar]
- PosthumaDeBoer, J.; Witlox, M.A.; Kaspers, G.J.; van Royen, B.J. Molecular alterations as target for therapy in metastatic osteosarcoma: A review of literature. Clin. Exp. Metastasis 2011, 28, 493–503. [Google Scholar] [CrossRef]
- Clark, J.C.; Dass, C.R.; Choong, P.F. A review of clinical and molecular prognostic factors in osteosarcoma. J. Cancer Res. Clin. Oncol. 2008, 134, 281–297. [Google Scholar] [CrossRef]
- Kong, C.; Hansen, M.F. Biomarkers in Osteosarcoma. Expert Opin. Med. Diagn. 2009, 3, 13–23. [Google Scholar] [CrossRef]
- Schulze, A.; Downward, J. Navigating gene expression using microarrays—A technology review. Nat. Cell Biol. 2001, 3, E190–E195. [Google Scholar] [CrossRef]
- Dupuy, A.; Simon, R.M. Critical review of published microarray studies for cancer outcome and guidelines on statistical analysis and reporting. J. Natl. Cancer Inst. 2007, 99, 147–157. [Google Scholar] [CrossRef]
- Goeman, J.J.; Buhlmann, P. Analyzing gene expression data in terms of gene sets: Methodological issues. Bioinformatics 2007, 23, 980–987. [Google Scholar]
- Song, S.; Black, M.A. Microarray-based gene set analysis: A comparison of current methods. BMC Bioinformatics 2008, 9, 502. [Google Scholar]
- Subramanian, A.; Kuehn, H.; Gould, J.; Tamayo, P.; Mesirov, J.P. GSEA-P: A desktop application for Gene Set Enrichment Analysis. Bioinformatics 2007, 23, 3251–3253. [Google Scholar]
- Chuang, H.Y.; Lee, E.; Liu, Y.T.; Lee, D.; Ideker, T. Network-based classification of breast cancer metastasis. Mol. Syst. Biol. 2007, 3, 140. [Google Scholar]
- Taylor, I.W.; Linding, R.; Warde-Farley, D.; Liu, Y.; Pesquita, C.; Faria, D.; Bull, S.; Pawson, T.; Morris, Q.; Wrana, J.L. Dynamic modularity in protein interaction networks predicts breast cancer outcome. Nat. Biotechnol. 2009, 27, 199–204. [Google Scholar]
- Perou, C.M.; Sorlie, T.; Eisen, M.B.; van de Rijn, M.; Jeffrey, S.S.; Rees, C.A.; Pollack, J.R.; Ross, D.T.; Johnsen, H.; Akslen, L.A.; et al. Molecular portraits of human breast tumours. Nature 2000, 406, 747–752. [Google Scholar]
- Van’t Veer, L.J.; Dai, H.; van de Vijver, M.J.; He, Y.D.; Hart, A.A.; Mao, M.; Peterse, H.L.; van der Kooy, K.; Marton, M.J.; Witteveen, A.T.; et al. Gene expression profiling predicts clinical outcome of breast cancer. Nature 2002, 415, 530–536. [Google Scholar]
- Cerami, E.G.; Gross, B.E.; Demir, E.; Rodchenkov, I.; Babur, O.; Anwar, N.; Schultz, N.; Bader, G.D.; Sander, C. Pathway Commons, a web resource for biological pathway data. Nucleic Acids Res. 2011, 39, D685–D690. [Google Scholar] [CrossRef]
- Osaka, E.; Suzuki, T.; Osaka, S.; Yoshida, Y.; Sugita, H.; Asami, S.; Tabata, K.; Hemmi, A.; Sugitani, M.; Nemoto, N.; et al. Survivin as a prognostic factor for osteosarcoma patients. Acta Histochem. Cytochem. 2006, 39, 95–100. [Google Scholar] [CrossRef]
- Pradelli, E.; Karimdjee-Soilihi, B.; Michiels, J.F.; Ricci, J.E.; Millet, M.A.; Vandenbos, F.; Sullivan, T.J.; Collins, T.L.; Johnson, M.G.; Medina, J.C.; et al. Antagonism of chemokine receptor CXCR3 inhibits osteosarcoma metastasis to lungs. Int. J. Cancer 2009, 125, 2586–2594. [Google Scholar] [CrossRef]
- Cantiani, L.; Manara, M.C.; Zucchini, C.; de Sanctis, P.; Zuntini, M.; Valvassori, L.; Serra, M.; Olivero, M.; di Renzo, M.F.; Colombo, M.P.; et al. Caveolin-1 reduces osteosarcoma metastases by inhibiting c-Src activity and met signaling. Cancer Res. 2007, 67, 7675–7685. [Google Scholar] [CrossRef]
- Entz-Werle, N.; Marcellin, L.; Gaub, M.P.; Guerin, E.; Schneider, A.; Berard-Marec, P.; Kalifa, C.; Brugiere, L.; Pacquement, H.; Schmitt, C.; et al. Prognostic significance of allelic imbalance at the c-kit gene locus and c-kit overexpression by immunohistochemistry in pediatric osteosarcomas. J. Clin. Oncol. 2005, 23, 2248–2255. [Google Scholar] [CrossRef]
- Boulytcheva, I.V.; Soloviev, Y.N.; Kushlinskii, N.E.; Mahson, A.N. Expression of molecular markers in the tumor and survival prognosis in osteosarcoma. Bull. Exp. Biol. Med. 2010, 150, 237–242. [Google Scholar]
- Oda, Y.; Wehrmann, B.; Radig, K.; Walter, H.; Rose, I.; Neumann, W.; Roessner, A. Expression of growth factors and their receptors in human osteosarcomas. Immunohistochemical detection of epidermal growth factor, platelet-derived growth factor and their receptors: Its correlation with proliferating activities and p53 expression. Gen. Diagn. Pathol. 1995, 141, 97–103. [Google Scholar]
- Fromigue, O.; Hamidouche, Z.; Vaudin, P.; Lecanda, F.; Patino, A.; Barbry, P.; Mari, B.; Marie, P.J. CYR61 downregulation reduces osteosarcoma cell invasion, migration, and metastasis. J. Bone Miner. Res. 2011, 26, 1533–1542. [Google Scholar] [CrossRef]
- Hoang, B.H.; Kubo, T.; Healey, J.H.; Sowers, R.; Mazza, B.; Yang, R.; Huvos, A.G.; Meyers, P.A.; Gorlick, R. Expression of LDL receptor-related protein 5 (LRP5) as a novel marker for disease progression in high-grade osteosarcoma. Int. J. Cancer 2004, 109, 106–111. [Google Scholar]
- Boulytcheva, I.V.; Soloviev, Y.N.; Kushlinskii, N.E.; Mahson, A.N. Expression of molecular markers in the tumor and survival prognosis in osteosarcoma. Bull. Exp. Biol. Med. 2010, 150, 237–242. [Google Scholar]
- Ferrari, S.; Bertoni, F.; Zanella, L.; Setola, E.; Bacchini, P.; Alberghini, M.; Versari, M.; Bacci, G. Evaluation of P-glycoprotein, HER-2/ErbB-2, p53, and Bcl-2 in primary tumor and metachronous lung metastases in patients with high-grade osteosarcoma. Cancer 2004, 100, 1936–1942. [Google Scholar] [CrossRef]
- Gorlick, R.; Huvos, A.G.; Heller, G.; Aledo, A.; Beardsley, G.P.; Healey, J.H.; Meyers, P.A. Expression of HER2/erbB-2 correlates with survival in osteosarcoma. J. Clin. Oncol. 1999, 17, 2781–2788. [Google Scholar]
- Onda, M.; Matsuda, S.; Higaki, S.; Iijima, T.; Fukushima, J.; Yokokura, A.; Kojima, T.; Horiuchi, H.; Kurokawa, T.; Yamamoto, T. ErbB-2 expression is correlated with poor prognosis for patients with osteosarcoma. Cancer 1996, 77, 71–78. [Google Scholar] [CrossRef]
- Scotlandi, K.; Manara, M.C.; Hattinger, C.M.; Benini, S.; Perdichizzi, S.; Pasello, M.; Bacci, G.; Zanella, L.; Bertoni, F.; Picci, P.; et al. Prognostic and therapeutic relevance of HER2 expression in osteosarcoma and Ewing's sarcoma. Eur. J. Cancer 2005, 41, 1349–1361. [Google Scholar] [CrossRef]
- Zhou, H.; Randall, R.L.; Brothman, A.R.; Maxwell, T.; Coffin, C.M.; Goldsby, R.E. Her-2/neu expression in osteosarcoma increases risk of lung metastasis and can be associated with gene amplification. J. Pediatr. Hematol. Oncol. 2003, 25, 27–32. [Google Scholar] [CrossRef]
- Perbal, B.; Zuntini, M.; Zambelli, D.; Serra, M.; Sciandra, M.; Cantiani, L.; Lucarelli, E.; Picci, P.; Scotlandi, K. Prognostic value of CCN3 in osteosarcoma. Clin. Cancer Res. 2008, 14, 701–709. [Google Scholar] [CrossRef]
- Bjornland, K.; Flatmark, K.; Pettersen, S.; Aaasen, A.O.; Fodstad, O.; Maelandsmo, G.M. Matrix metalloproteinases participate in osteosarcoma invasion. J. Surg. Res. 2005, 127, 151–156. [Google Scholar] [CrossRef]
- Gordon, N.; Koshkina, N.V.; Jia, S.F.; Khanna, C.; Mendoza, A.; Worth, L.L.; Kleinerman, E.S. Corruption of the Fas pathway delays the pulmonary clearance of murine osteosarcoma cells, enhances their metastatic potential, and reduces the effect of aerosol gemcitabine. Clin. Cancer Res. 2007, 13, 4503–4510. [Google Scholar] [CrossRef]
- Weber, G.F.; Bronson, R.T.; Ilagan, J.; Cantor, H.; Schmits, R.; Mak, T.W. Absence of the CD44 gene prevents sarcoma metastasis. Cancer Res. 2002, 62, 2281–2286. [Google Scholar]
- Kubo, T.; Piperdi, S.; Rosenblum, J.; Antonescu, C.R.; Chen, W.; Kim, H.S.; Huvos, A.G.; Sowers, R.; Meyers, P.A.; Healey, J.H.; et al. Platelet-derived growth factor receptor as a prognostic marker and a therapeutic target for imatinib mesylate therapy in osteosarcoma. Cancer 2008, 112, 2119–2129. [Google Scholar] [CrossRef]
- Sulzbacher, I.; Birner, P.; Trieb, K.; Traxler, M.; Lang, S.; Chott, A. Expression of platelet-derived growth factor-AA is associated with tumor progression in osteosarcoma. Mod. Pathol. 2003, 16, 66–71. [Google Scholar] [CrossRef]
- Mizobuchi, H.; Garcia-Castellano, J.M.; Philip, S.; Healey, J.H.; Gorlick, R. Hypoxia markers in human osteosarcoma: An exploratory study. Clin. Orthop. Relat. Res. 2008, 466, 2052–2059. [Google Scholar] [CrossRef]
- Kashima, T.; Nakamura, K.; Kawaguchi, J.; Takanashi, M.; Ishida, T.; Aburatani, H.; Kudo, A.; Fukayama, M.; Grigoriadis, A.E. Overexpression of cadherins suppresses pulmonary metastasis of osteosarcoma in vivo. Int. J. Cancer 2003, 104, 147–154. [Google Scholar] [CrossRef]
- Ek, E.T.; Dass, C.R.; Contreras, K.G.; Choong, P.F. PEDF-derived synthetic peptides exhibit antitumor activity in an orthotopic model of human osteosarcoma. J. Orthop. Res. 2007, 25, 1671–1680. [Google Scholar] [CrossRef]
- Duan, X.; Jia, S.F.; Koshkina, N.; Kleinerman, E.S. Intranasal interleukin-12 gene therapy enhanced the activity of ifosfamide against osteosarcoma lung metastases. Cancer 2006, 106, 1382–1388. [Google Scholar] [CrossRef]
- Xu, J.; Wu, S.; Shi, X. Expression of matrix metalloproteinase regulator, RECK, and its clinical significance in osteosarcoma. J. Orthop. Res. 2010, 28, 1621–1625. [Google Scholar] [CrossRef]
- Kaya, M.; Wada, T.; Nagoya, S.; Yamashita, T. Prevention of postoperative progression of pulmonary metastases in osteosarcoma by antiangiogenic therapy using endostatin. J. Orthop. Sci. 2007, 12, 562–567. [Google Scholar] [CrossRef]
- Luu, H.H.; Zhou, L.; Haydon, R.C.; Deyrup, A.T.; Montag, A.G.; Huo, D.; Heck, R.; Heizmann, C.W.; Peabody, T.D.; Simon, M.A.; He, T.C. Increased expression of S100A6 is associated with decreased metastasis and inhibition of cell migration and anchorage independent growth in human osteosarcoma. Cancer Lett. 2005, 229, 135–148. [Google Scholar] [CrossRef]
- Braczkowski, R.; Schally, A.V.; Plonowski, A.; Varga, J.L.; Groot, K.; Krupa, M.; Armatis, P. Inhibition of proliferation in human MNNG/HOS osteosarcoma and SK-ES-1 Ewing sarcoma cell lines in vitro and in vivo by antagonists of growth hormone-releasing hormone: Effects on insulin-like growth factor II. Cancer 2002, 95, 1735–1745. [Google Scholar]
- Burrow, S.; Andrulis, I.L.; Pollak, M.; Bell, R.S. Expression of insulin-like growth factor receptor, IGF-1, and IGF-2 in primary and metastatic osteosarcoma. J. Surg. Oncol. 1998, 69, 21–27. [Google Scholar] [CrossRef]
- Duan, Z.; Choy, E.; Harmon, D.; Yang, C.; Ryu, K.; Schwab, J.; Mankin, H.; Hornicek, F.J. Insulin-like growth factor-I receptor tyrosine kinase inhibitor cyclolignan picropodophyllin inhibits proliferation and induces apoptosis in multidrug resistant osteosarcoma cell lines. Mol. Cancer Ther. 2009, 8, 2122–2130. [Google Scholar]
- Kolb, E.A.; Gorlick, R.; Houghton, P.J.; Morton, C.L.; Lock, R.; Carol, H.; Reynolds, C.P.; Maris, J.M.; Keir, S.T.; Billups, C.A.; et al. Initial testing (stage 1) of a monoclonal antibody (SCH 717454) against the IGF-1 receptor by the pediatric preclinical testing program. Pediatr. Blood Cancer 2008, 50, 1190–1197. [Google Scholar] [CrossRef]
- Kolb, E.A.; Kamara, D.; Zhang, W.; Lin, J.; Hingorani, P.; Baker, L.; Houghton, P.; Gorlick, R. R1507, a fully human monoclonal antibody targeting IGF-1R, is effective alone and in combination with rapamycin in inhibiting growth of osteosarcoma xenografts. Pediatr. Blood Cancer 2010, 55, 67–75. [Google Scholar]
- Pinski, J.; Schally, A.V.; Groot, K.; Halmos, G.; Szepeshazi, K.; Zarandi, M.; Armatis, P. Inhibition of growth of human osteosarcomas by antagonists of growth hormone-releasing hormone. J. Natl. Cancer Inst. 1995, 87, 1787–1794. [Google Scholar] [CrossRef]
- Pinski, J.; Schally, A.V.; Halmos, G.; Szepeshazi, K.; Groot, K. Somatostatin analog RC-160 inhibits the growth of human osteosarcomas in nude mice. Int. J. Cancer 1996, 65, 870–874. [Google Scholar] [CrossRef]
- Pollack, R. Suramin Blockade of InsulinLike Growth Factor I-Stimulated Proliferation of Human Osteosarcoma Cells. J. Natl. Cancer Inst. 1990, 82, 1349–1352. [Google Scholar] [CrossRef]
- Sekyi-Otu, A.; Bell, R.S.; Ohashi, C.; Pollak, M.; Andrulis, I.L. Insulin-like growth factor 1 (IGF-1) receptors, IGF-1, and IGF-2 are expressed in primary human sarcomas. Cancer Res. 1995, 55, 129–134. [Google Scholar]
- Kim, S.Y.; Lee, C.H.; Midura, B.V.; Yeung, C.; Mendoza, A.; Hong, S.H.; Ren, L.; Wong, D.; Korz, W.; Merzouk, A.; et al. Inhibition of the CXCR4/CXCL12 chemokine pathway reduces the development of murine pulmonary metastases. Clin. Exp. Metastasis 2008, 25, 201–211. [Google Scholar] [CrossRef]
- Laverdiere, C.; Hoang, B.H.; Yang, R.; Sowers, R.; Qin, J.; Meyers, P.A.; Huvos, A.G.; Healey, J.H.; Gorlick, R. Messenger RNA expression levels of CXCR4 correlate with metastatic behavior and outcome in patients with osteosarcoma. Clin. Cancer Res. 2005, 11, 2561–2567. [Google Scholar]
- Lin, F.; Zheng, S.E.; Shen, Z.; Tang, L.N.; Chen, P.; Sun, Y.J.; Zhao, H.; Yao, Y. Relationships between levels of CXCR4 and VEGF and blood-borne metastasis and survival in patients with osteosarcoma. Med. Oncol. 2011, 28, 649–653. [Google Scholar] [CrossRef]
- Oda, Y.; Yamamoto, H.; Tamiya, S.; Matsuda, S.; Tanaka, K.; Yokoyama, R.; Iwamoto, Y.; Tsuneyoshi, M. CXCR4 and VEGF expression in the primary site and the metastatic site of human osteosarcoma: Analysis within a group of patients, all of whom developed lung metastasis. Mod. Pathol. 2006, 19, 738–745. [Google Scholar] [CrossRef]
- Dalla-Torre, C.A.; Yoshimoto, M.; Lee, C.H.; Joshua, A.M.; de Toledo, S.R.; Petrilli, A.S.; Andrade, J.A.; Chilton-MacNeill, S.; Zielenska, M.; Squire, J.A. Effects of THBS3, SPARC and SPP1 expression on biological behavior and survival in patients with osteosarcoma. BMC Cancer 2006, 6, 237. [Google Scholar] [CrossRef]
- Engin, F.; Bertin, T.; Ma, O.; Jiang, M.M.; Wang, L.; Sutton, R.E.; Donehower, L.A.; Lee, B. Notch signaling contributes to the pathogenesis of human osteosarcomas. Hum. Mol. Genet. 2009, 18, 1464–1470. [Google Scholar] [CrossRef]
- Ferrari, C.; Benassi, S.; Ponticelli, F.; Gamberi, G.; Ragazzini, P.; Pazzaglia, L.; Balladelli, A.; Bertoni, F.; Picci, P. Role of MMP-9 and its tissue inhibitor TIMP-1 in human osteosarcoma: Findings in 42 patients followed for 1–16 years. Acta Orthop. Scand. 2004, 75, 487–491. [Google Scholar] [CrossRef]
- Yang, C.; Ji, D.; Weinstein, E.J.; Choy, E.; Hornicek, F.J.; Wood, K.B.; Liu, X.; Mankin, H.; Duan, Z. The kinase Mirk is a potential therapeutic target in osteosarcoma. Carcinogenesis 2010, 31, 552–558. [Google Scholar] [CrossRef]
- Kersting, C.; Gebert, C.; Agelopoulos, K.; Schmidt, H.; van Diest, P.J.; Juergens, H.; Winkelmann, W.; Kevric, M.; Gosheger, G.; Brandt, B.; et al. Epidermal growth factor receptor expression in high-grade osteosarcomas is associated with a good clinical outcome. Clin. Cancer Res. 2007, 13, 2998–3005. [Google Scholar] [CrossRef]
- Dass, C.R.; Nadesapillai, A.P.; Robin, D.; Howard, M.L.; Fisher, J.L.; Zhou, H.; Choong, P.F. Downregulation of uPAR confirms link in growth and metastasis of osteosarcoma. Clin. Exp. Metastasis 2005, 22, 643–652. [Google Scholar] [CrossRef]
- Uchibori, M.; Nishida, Y.; Nagasaka, T.; Yamada, Y.; Nakanishi, K.; Ishiguro, N. Increased expression of membrane-type matrix metalloproteinase-1 is correlated with poor prognosis in patients with osteosarcoma. Int. J. Oncol. 2006, 28, 33–42. [Google Scholar]
- Khanna, C.; Wan, X.; Bose, S.; Cassaday, R.; Olomu, O.; Mendoza, A.; Yeung, C.; Gorlick, R.; Hewitt, S.M.; Helman, L.J. The membrane-cytoskeleton linker ezrin is necessary for osteosarcoma metastasis. Nat. Med. 2004, 10, 182–186. [Google Scholar]
- Kim, M.S.; Song, W.S.; Cho, W.H.; Lee, S.Y.; Jeon, D.G. Ezrin expression predicts survival in stage IIB osteosarcomas. Clin. Orthop. Relat. Res. 2007, 459, 229–236. [Google Scholar]
- Abdeen, A.; Chou, A.J.; Healey, J.H.; Khanna, C.; Osborne, T.S.; Hewitt, S.M.; Kim, M.; Wang, D.; Moody, K.; Gorlick, R. Correlation between clinical outcome and growth factor pathway expression in osteogenic sarcoma. Cancer 2009, 115, 5243–5250. [Google Scholar] [CrossRef]
- Charity, R.M.; Foukas, A.F.; Deshmukh, N.S.; Grimer, R.J. Vascular endothelial growth factor expression in osteosarcoma. Clin. Orthop. Relat. Res. 2006, 448, 193–198. [Google Scholar] [CrossRef]
- Feng, Y.; Hu, J.; Ma, J.; Feng, K.; Zhang, X.; Yang, S.; Wang, W.; Zhang, J.; Zhang, Y. RNAi-mediated silencing of VEGF-C inhibits non-small cell lung cancer progression by simultaneously down-regulating the CXCR4, CCR7, VEGFR-2 and VEGFR-3-dependent axes-induced ERK, p38 and AKT signalling pathways. Eur. J. Cancer 2011, 47, 2353–2363. [Google Scholar]
- Kaya, M.; Wada, T.; Akatsuka, T.; Kawaguchi, S.; Nagoya, S.; Shindoh, M.; Higashino, F.; Mezawa, F.; Okada, F.; Ishii, S. Vascular endothelial growth factor expression in untreated osteosarcoma is predictive of pulmonary metastasis and poor prognosis. Clin. Cancer Res. 2000, 6, 572–577. [Google Scholar]
- Kaya, M.; Wada, T.; Kawaguchi, S.; Nagoya, S.; Yamashita, T.; Abe, Y.; Hiraga, H.; Isu, K.; Shindoh, M.; Higashino, F.; et al. Increased pre-therapeutic serum vascular endothelial growth factor in patients with early clinical relapse of osteosarcoma. Br. J. Cancer 2002, 86, 864–869. [Google Scholar] [CrossRef]
- Kaya, M.; Wada, T.; Nagoya, S.; Sasaki, M.; Matsumura, T.; Yamashita, T. The level of vascular endothelial growth factor as a predictor of a poor prognosis in osteosarcoma. J. Bone Joint Surg. Br. 2009, 91, 784–788. [Google Scholar] [CrossRef]
- Lee, Y.H.; Tokunaga, T.; Oshika, Y.; Suto, R.; Yanagisawa, K.; Tomisawa, M.; Fukuda, H.; Nakano, H.; Abe, S.; Tateishi, A.; et al. Cell-retained isoforms of vascular endothelial growth factor (VEGF) are correlated with poor prognosis in osteosarcoma. Eur. J. Cancer 1999, 35, 1089–1093. [Google Scholar] [CrossRef]
- Sulzbacher, I.; Birner, P.; Trieb, K.; Lang, S.; Chott, A. Expression of osteopontin and vascular endothelial growth factor in benign and malignant bone tumors. Virchows Arch. 2002, 441, 345–349. [Google Scholar] [CrossRef]
- Foukas, A.F.; Deshmukh, N.S.; Grimer, R.J.; Mangham, D.C.; Mangos, E.G.; Taylor, S. Stage-IIB osteosarcomas around the knee. A study of MMP-9 in surviving tumour cells. J. Bone Joint Surg. Br. 2002, 84, 706–711. [Google Scholar] [CrossRef]
- Himelstein, B.P.; Asada, N.; Carlton, M.R.; Collins, M.H. Matrix metalloproteinase-9 (MMP-9) expression in childhood osseous osteosarcoma. Med. Pediatr. Oncol. 1998, 31, 471–474. [Google Scholar] [CrossRef]
- Gordon, N.; Arndt, C.A.; Hawkins, D.S.; Doherty, D.K.; Inwards, C.Y.; Munsell, M.F.; Stewart, J.; Koshkina, N.V.; Kleinerman, E.S. Fas expression in lung metastasis from osteosarcoma patients. J. Pediatr. Hematol. Oncol. 2005, 27, 611–615. [Google Scholar] [CrossRef]
- Koshkina, N.V.; Khanna, C.; Mendoza, A.; Guan, H.; DeLauter, L.; Kleinerman, E.S. Fas-negative osteosarcoma tumor cells are selected during metastasis to the lungs: The role of the Fas pathway in the metastatic process of osteosarcoma. Mol. Cancer Res. 2007, 5, 991–999. [Google Scholar] [CrossRef]
- Lafleur, E.A.; Koshkina, N.V.; Stewart, J.; Jia, S.F.; Worth, L.L.; Duan, X.; Kleinerman, E.S. Increased Fas expression reduces the metastatic potential of human osteosarcoma cells. Clin Cancer Res. 2004, 10, 8114–8119. [Google Scholar] [CrossRef]
- Asai, T.; Tomita, Y.; Nakatsuka, S.; Hoshida, Y.; Myoui, A.; Yoshikawa, H.; Aozasa, K. VCP (p97) regulates NFkappaB signaling pathway, which is important for metastasis of osteosarcoma cell line. Jpn. J. Cancer Res. 2002, 93, 296–304. [Google Scholar]
- Rubin, E.M.; Guo, Y.; Tu, K.; Xie, J.; Zi, X.; Hoang, B.H. Wnt inhibitory factor 1 decreases tumorigenesis and metastasis in osteosarcoma. Mol. Cancer Ther. 2010, 9, 731–741. [Google Scholar] [CrossRef]
- De Nigris, F.; Rossiello, R.; Schiano, C.; Arra, C.; Williams-Ignarro, S.; Barbieri, A.; Lanza, A.; Balestrieri, A.; Giuliano, M.T.; Ignarro, L.J.; et al. Deletion of Yin Yang 1 protein in osteosarcoma cells on cell invasion and CXCR4/angiogenesis and metastasis. Cancer Res. 2008, 68, 1797–1808. [Google Scholar] [CrossRef]
- Han, J.D.; Bertin, N.; Hao, T.; Goldberg, D.S.; Berriz, G.F.; Zhang, L.V.; Dupuy, D.; Walhout, A.J.; Cusick, M.E.; Roth, F.P.; et al. Evidence for dynamically organized modularity in the yeast protein-protein interaction network. Nature 2004, 430, 88–93. [Google Scholar]
- Aziz, M.H.; Manoharan, H.T.; Verma, A.K. Protein kinase C epsilon, which sensitizes skin to sun’s UV radiation-induced cutaneous damage and development of squamous cell carcinomas, associates with Stat3. Cancer Res. 2007, 67, 1385–1394. [Google Scholar] [CrossRef]
- Gundimeda, U.; Schiffman, J.E.; Gottlieb, S.N.; Roth, B.I.; Gopalakrishna, R. Negation of the cancer-preventive actions of selenium by over-expression of protein kinase Cepsilon and selenoprotein thioredoxin reductase. Carcinogenesis 2009, 30, 1553–1561. [Google Scholar]
- Hafeez, B.B.; Zhong, W.; Weichert, J.; Dreckschmidt, N.E.; Jamal, M.S.; Verma, A.K. Genetic ablation of PKC epsilon inhibits prostate cancer development and metastasis in transgenic mouse model of prostate adenocarcinoma. Cancer Res. 2011, 71, 2318–2327. [Google Scholar] [CrossRef]
- McJilton, M.A.; van Sikes, C.; Wescott, G.G.; Wu, D.; Foreman, T.L.; Gregory, C.W.; Weidner, D.A.; Harris Ford, O.; Morgan Lasater, A.; Mohler, J.L.; et al. Protein kinase Cepsilon interacts with Bax and promotes survival of human prostate cancer cells. Oncogene 2003, 22, 7958–7968. [Google Scholar] [CrossRef]
- Meshki, J.; Caino, M.C.; von Burstin, V.A.; Griner, E.; Kazanietz, M.G. Regulation of prostate cancer cell survival by protein kinase Cepsilon involves bad phosphorylation and modulation of the TNFalpha/JNK pathway. J. Biol. Chem. 2010, 285, 26033–26040. [Google Scholar]
- Wu, D.; Foreman, T.L.; Gregory, C.W.; McJilton, M.A.; Wescott, G.G.; Ford, O.H.; Alvey, R.F.; Mohler, J.L.; Terrian, D.M. Protein kinase cepsilon has the potential to advance the recurrence of human prostate cancer. Cancer Res. 2002, 62, 2423–2429. [Google Scholar]
- Grossoni, V.C.; Todaro, L.B.; Kazanietz, M.G.; de Kier Joffe, E.D.; Urtreger, A.J. Opposite effects of protein kinase C beta1 (PKCbeta1) and PKCepsilon in the metastatic potential of a breast cancer murine model. Breast Cancer Res. Treat. 2009, 118, 469–480. [Google Scholar]
- Pan, Q.; Bao, L.W.; Kleer, C.G.; Sabel, M.S.; Griffith, K.A.; Teknos, T.N.; Merajver, S.D. Protein kinase C epsilon is a predictive biomarker of aggressive breast cancer and a validated target for RNA interference anticancer therapy. Cancer Res. 2005, 65, 8366–8371. [Google Scholar]
- Huang, B.; Cao, K.; Li, X.; Guo, S.; Mao, X.; Wang, Z.; Zhuang, J.; Pan, J.; Mo, C.; Chen, J.; et al. The expression and role of protein kinase C (PKC) epsilon in clear cell renal cell carcinoma. J. Exp. Clin. Cancer Res. 2011, 30, 88. [Google Scholar] [CrossRef]
- Jansen, A.P.; Verwiebe, E.G.; Dreckschmidt, N.E.; Wheeler, D.L.; Oberley, T.D.; Verma, A.K. Protein kinase C-epsilon transgenic mice: A unique model for metastatic squamous cell carcinoma. Cancer Res. 2001, 61, 808–812. [Google Scholar]
- Reddig, P.J.; Dreckschmidt, N.E.; Zou, J.; Bourguignon, S.E.; Oberley, T.D.; Verma, A.K. Transgenic mice overexpressing protein kinase C epsilon in their epidermis exhibit reduced papilloma burden but enhanced carcinoma formation after tumor promotion. Cancer Res. 2000, 60, 595–602. [Google Scholar]
- Sand, J.M.; Aziz, M.H.; Dreckschmidt, N.E.; Havighurst, T.C.; Kim, K.; Oberley, T.D.; Verma, A.K. PKCepsilon overexpression, irrespective of genetic background, sensitizes skin to UVR-induced development of squamous-cell carcinomas. J. Invest. Dermatol. 2010, 130, 270–277. [Google Scholar] [CrossRef]
- Sand, J.M.; Hafeez, B.B.; Aziz, M.H.; Siebers, E.M.; Dreckschmidt, N.E.; Verma, A.K. Ultraviolet radiation and 12-O-tetradecanoylphorbol-13-acetate-induced interaction of mouse epidermal protein kinase C epsilon with Stat3 involve integration with Erk1/2. Mol. Carcinog. 2012, 51, 291–302. [Google Scholar] [CrossRef]
- Wheeler, D.L.; Ness, K.J.; Oberley, T.D.; Verma, A.K. Inhibition of the development of metastatic squamous cell carcinoma in protein kinase C epsilon transgenic mice by alpha-difluoromethylornithine accompanied by marked hair follicle degeneration and hair loss. Cancer Res. 2003, 63, 3037–3042. [Google Scholar]
- Bao, L.; Gorin, M.A.; Zhang, M.; Ventura, A.C.; Pomerantz, W.C.; Merajver, S.D.; Teknos, T.N.; Mapp, A.K.; Pan, Q. Preclinical development of a bifunctional cancer cell homing, PKCepsilon inhibitory peptide for the treatment of head and neck cancer. Cancer Res. 2009, 69, 5829–5834. [Google Scholar] [CrossRef]
- Martinez-Gimeno, C.; Diaz-Meco, M.T.; Dominguez, I.; Moscat, J. Alterations in levels of different protein kinase C isotypes and their influence on behavior of squamous cell carcinoma of the oral cavity: Epsilon PKC, a novel prognostic factor for relapse and survival. Head Neck 1995, 17, 516–525. [Google Scholar]
- Pan, Q.; Bao, L.W.; Teknos, T.N.; Merajver, S.D. Targeted disruption of protein kinase C epsilon reduces cell invasion and motility through inactivation of RhoA and RhoC GTPases in head and neck squamous cell carcinoma. Cancer Res. 2006, 66, 9379–9384. [Google Scholar] [CrossRef]
- Tosi, L.; Rinaldi, E.; Carinci, F.; Farina, A.; Pastore, A.; Pelucchi, S.; Cassano, L.; Evangelisti, R.; Carinci, P.; Volinia, S. Akt, protein kinase C, and mitogen-activated protein kinase phosphorylation status in head and neck squamous cell carcinoma. Head Neck 2005, 27, 130–137. [Google Scholar] [CrossRef]
- Bae, K.M.; Wang, H.; Jiang, G.; Chen, M.G.; Lu, L.; Xiao, L. Protein kinase C epsilon is overexpressed in primary human non-small cell lung cancers and functionally required for proliferation of non-small cell lung cancer cells in a p21/Cip1-dependent manner. Cancer Res. 2007, 67, 6053–6063. [Google Scholar] [CrossRef]
- Caino, M.C.; von Burstin, V.A.; Lopez-Haber, C.; Kazanietz, M.G. Differential regulation of gene expression by protein kinase C isozymes as determined by genome-wide expression analysis. J. Biol. Chem. 2011, 286, 11254–11264. [Google Scholar]
- Ding, L.; Wang, H.; Lang, W.; Xiao, L. Protein kinase C-epsilon promotes survival of lung cancer cells by suppressing apoptosis through dysregulation of the mitochondrial caspase pathway. J. Biol. Chem. 2002, 277, 35305–35313. [Google Scholar] [CrossRef]
- Felber, M.; Sonnemann, J.; Beck, J.F. Inhibition of novel protein kinase C-epsilon augments TRAIL-induced cell death in A549 lung cancer cells. Pathol. Oncol. Res. 2007, 13, 295–301. [Google Scholar]
- Aviel-Ronen, S.; Coe, B.P.; Lau, S.K.; da Cunha Santos, G.; Zhu, C.Q.; Strumpf, D.; Jurisica, I.; Lam, W.L.; Tsao, M.S. Genomic markers for malignant progression in pulmonary adenocarcinoma with bronchioloalveolar features. Proc. Natl. Acad. Sci. USA 2008, 105, 10155–10160. [Google Scholar] [CrossRef]
- Yang, D.; Kedei, N.; Li, L.; Tao, J.; Velasquez, J.F.; Michalowski, A.M.; Toth, B.I.; Marincsak, R.; Varga, A.; Biro, T.; et al. RasGRP3 contributes to formation and maintenance of the prostate cancer phenotype. Cancer Res. 2010, 70, 7905–7917. [Google Scholar] [CrossRef]
- Kimura, K.; Nakano, T.; Park, Y.B.; Tani, M.; Tsuda, H.; Beppu, Y.; Moriya, H.; Yokota, J. Establishment of human osteosarcoma cell lines with high metastatic potential to lungs and their utilities for therapeutic studies on metastatic osteosarcoma. Clin. Exp. Metastasis 2002, 19, 477–485. [Google Scholar]
- Nakano, T.; Tani, M.; Ishibashi, Y.; Kimura, K.; Park, Y.B.; Imaizumi, N.; Tsuda, H.; Aoyagi, K.; Sasaki, H.; Ohwada, S.; et al. Biological properties and gene expression associated with metastatic potential of human osteosarcoma. Clin. Exp. Metastasis 2003, 20, 665–674. [Google Scholar] [CrossRef]
- Jia, S.F.; Worth, L.L.; Kleinerman, E.S. A nude mouse model of human osteosarcoma lung metastases for evaluating new therapeutic strategies. Clin. Exp. Metastasis 1999, 17, 501–506. [Google Scholar] [CrossRef]
- Zhu, W.; Guo, F.; Xu, T.; Chen, A. Establishment of nude mice model of human osteosarcoma cell line MG63 with different potential of metastasis. Chin. Ger. J. Clin. Oncol. 2007, 6, 484–486. [Google Scholar] [CrossRef]
- England, K.; Ashford, D.; Kidd, D.; Rumsby, M. PKC epsilon is associated with myosin IIA and actin in fibroblasts. Cell. Signal. 2002, 14, 529–536. [Google Scholar] [CrossRef]
- Allen, T.R.; Krueger, K.D.; Hunter, K.W.; Agrawal, D.K. Evidence that insulin-like growth factor-1 requires protein kinase C-(epsilon), PI3-kinase and mitogen-activated protein kinase pathways to protect human vascular smooth muscle cells from apoptosis. Immunol. Cell. Biol. 2005, 83, 651–667. [Google Scholar]
- Thommes, K.B.; Hoppe, J.; Vetter, H.; Sachinidis, A. The synergistic effect of PDGF-AA and IGF-1 on VSMC proliferation might be explained by the differential activation of their intracellular signaling pathways. Exp. Cell Res. 1996, 226, 59–66. [Google Scholar]
- Yano, K.; Bauchat, J.R.; Liimatta, M.B.; Clemmons, D.R.; Duan, C. Down-regulation of protein kinase C inhibits insulin-like growth factor I-induced vascular smooth muscle cell proliferation, migration, and gene expression. Endocrinology 1999, 140, 4622–4632. [Google Scholar] [CrossRef]
- Newton, A.C. Regulation of the ABC kinases by phosphorylation: Protein kinase C as a paradigm. Biochem. J. 2003, 370, 361–371. [Google Scholar] [CrossRef]
- Mamane, Y.; Petroulakis, E.; LeBacquer, O.; Sonenberg, N. mTOR, translation initiation and cancer. Oncogene 2006, 25, 6416–6422. [Google Scholar]
- Hua, Y.; Qiu, Y.; Zhao, A.; Wang, X.; Chen, T.; Zhang, Z.; Chi, Y.; Li, Q.; Sun, W.; Li, G.; et al. Dynamic metabolic transformation in tumor invasion and metastasis in mice with LM-8 osteosarcoma cell transplantation. J. Proteome Res. 2011, 10, 3513–3521. [Google Scholar] [CrossRef]
- Huang, B.; Cao, K.; Li, X.; Guo, S.; Mao, X.; Wang, Z.; Zhuang, J.; Pan, J.; Mo, C.; Chen, J.; et al. The expression and role of protein kinase C (PKC) epsilon in clear cell renal cell carcinoma. J. Exp. Clin. Cancer Res. 2011, 30, 88. [Google Scholar] [CrossRef]
- Leask, A.; Shi-Wen, X.; Khan, K.; Chen, Y.; Holmes, A.; Eastwood, M.; Denton, C.P.; Black, C.M.; Abraham, D.J. Loss of protein kinase Cepsilon results in impaired cutaneous wound closure and myofibroblast function. J. Cell. Sci. 2008, 121, 3459–3467. [Google Scholar] [CrossRef]
- Simon, R.M.; Lam, A.; Li, M.C.; Ngan, M.; Menenzes, S.; Zhao, Y. Analysis of Gene Expression Data Using BRB-Array Tools. Cancer Inform. 2007, 3, 11–17. [Google Scholar]
- Merico, D.; Isserlin, R.; Stueker, O.; Emili, A.; Bader, G.D. Enrichment map: A network-based method for gene-set enrichment visualization and interpretation. PLoS One 2010, 5, e13984. [Google Scholar]
- Cline, M.S.; Smoot, M.; Cerami, E.; Kuchinsky, A.; Landys, N.; Workman, C.; Christmas, R.; Avila-Campilo, I.; Creech, M.; Gross, B.; et al. Integration of biological networks and gene expression data using Cytoscape. Nat. Protoc. 2007, 2, 2366–2382. [Google Scholar] [CrossRef]
- Harris, M.A.; Clark, J.; Ireland, A.; Lomax, J.; Ashburner, M.; Foulger, R.; Eilbeck, K.; Lewis, S.; Marshall, B.; Mungall, C.; et al. The Gene Ontology (GO) database and informatics resource. Nucleic Acids Res. 2004, 32, D258–D261. [Google Scholar] [CrossRef]
- Boyle, E.I.; Weng, S.; Gollub, J.; Jin, H.; Botstein, D.; Cherry, J.M.; Sherlock, G. GO::TermFinder—open source software for accessing Gene Ontology information and finding significantly enriched Gene Ontology terms associated with a list of genes. Bioinformatics 2004, 20, 3710–3715. [Google Scholar]
- Eppert, K.; Wunder, J.S.; Aneliunas, V.; Kandel, R.; Andrulis, I.L. von Willebrand factor expression in osteosarcoma metastasis. Mod. Pathol. 2005, 18, 388–397. [Google Scholar] [CrossRef]
- Gorin, M.A.; Pan, Q. Protein kinase C epsilon: An oncogene and emerging tumor biomarker. Mol. Cancer 2009, 8, 9. [Google Scholar]
- Griner, E.M.; Kazanietz, M.G. Protein kinase C and other diacylglycerol effectors in cancer. Nat. Rev. Cancer 2007, 7, 281–294. [Google Scholar] [CrossRef]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Goudarzi, A.; Gokgoz, N.; Gill, M.; Pinnaduwage, D.; Merico, D.; Wunder, J.S.; Andrulis, I.L. Protein Kinase C Epsilon and Genetic Networks in Osteosarcoma Metastasis. Cancers 2013, 5, 372-403. https://doi.org/10.3390/cancers5020372
Goudarzi A, Gokgoz N, Gill M, Pinnaduwage D, Merico D, Wunder JS, Andrulis IL. Protein Kinase C Epsilon and Genetic Networks in Osteosarcoma Metastasis. Cancers. 2013; 5(2):372-403. https://doi.org/10.3390/cancers5020372
Chicago/Turabian StyleGoudarzi, Atta, Nalan Gokgoz, Mona Gill, Dushanthi Pinnaduwage, Daniele Merico, Jay S. Wunder, and Irene L. Andrulis. 2013. "Protein Kinase C Epsilon and Genetic Networks in Osteosarcoma Metastasis" Cancers 5, no. 2: 372-403. https://doi.org/10.3390/cancers5020372
APA StyleGoudarzi, A., Gokgoz, N., Gill, M., Pinnaduwage, D., Merico, D., Wunder, J. S., & Andrulis, I. L. (2013). Protein Kinase C Epsilon and Genetic Networks in Osteosarcoma Metastasis. Cancers, 5(2), 372-403. https://doi.org/10.3390/cancers5020372