Effects of Alpha Particle and Proton Beam Irradiation as Putative Cross-Talk between A549 Cancer Cells and the Endothelial Cells in a Co-Culture System

Abstract
:1. Introduction
2. Results


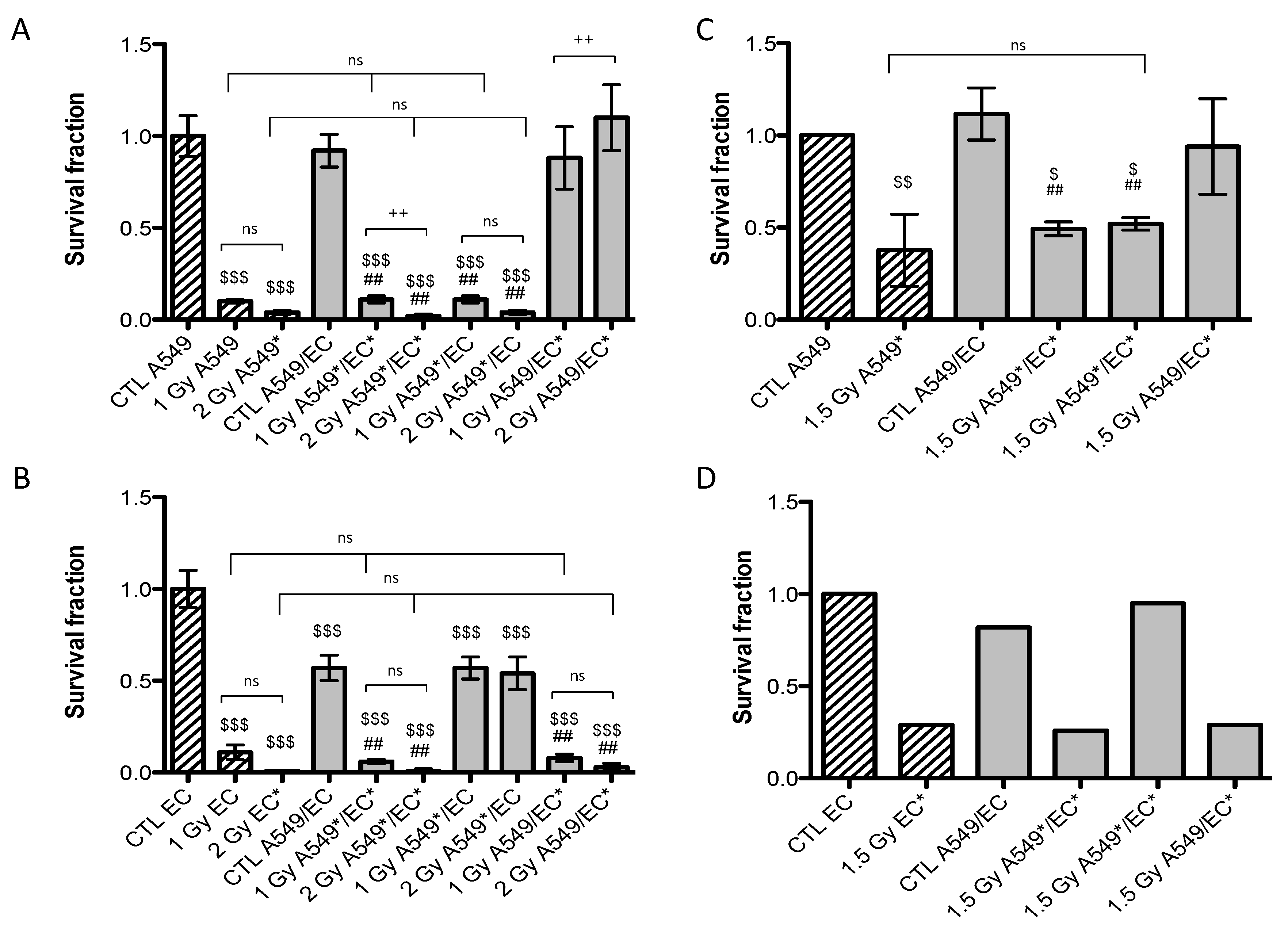
2.1. Survival Fraction of A549 Cells and EC after Particle Irradiation in Mono- or Co-Culture Configurations

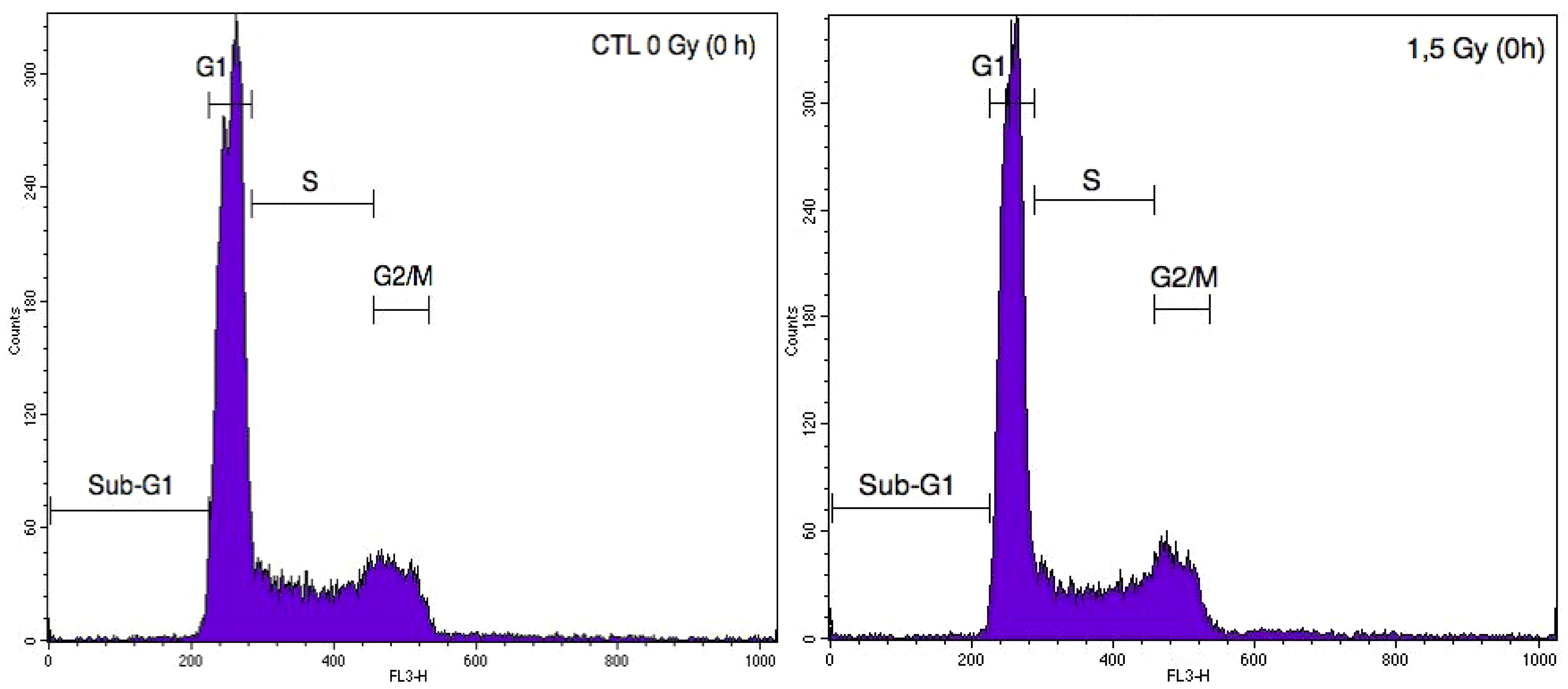
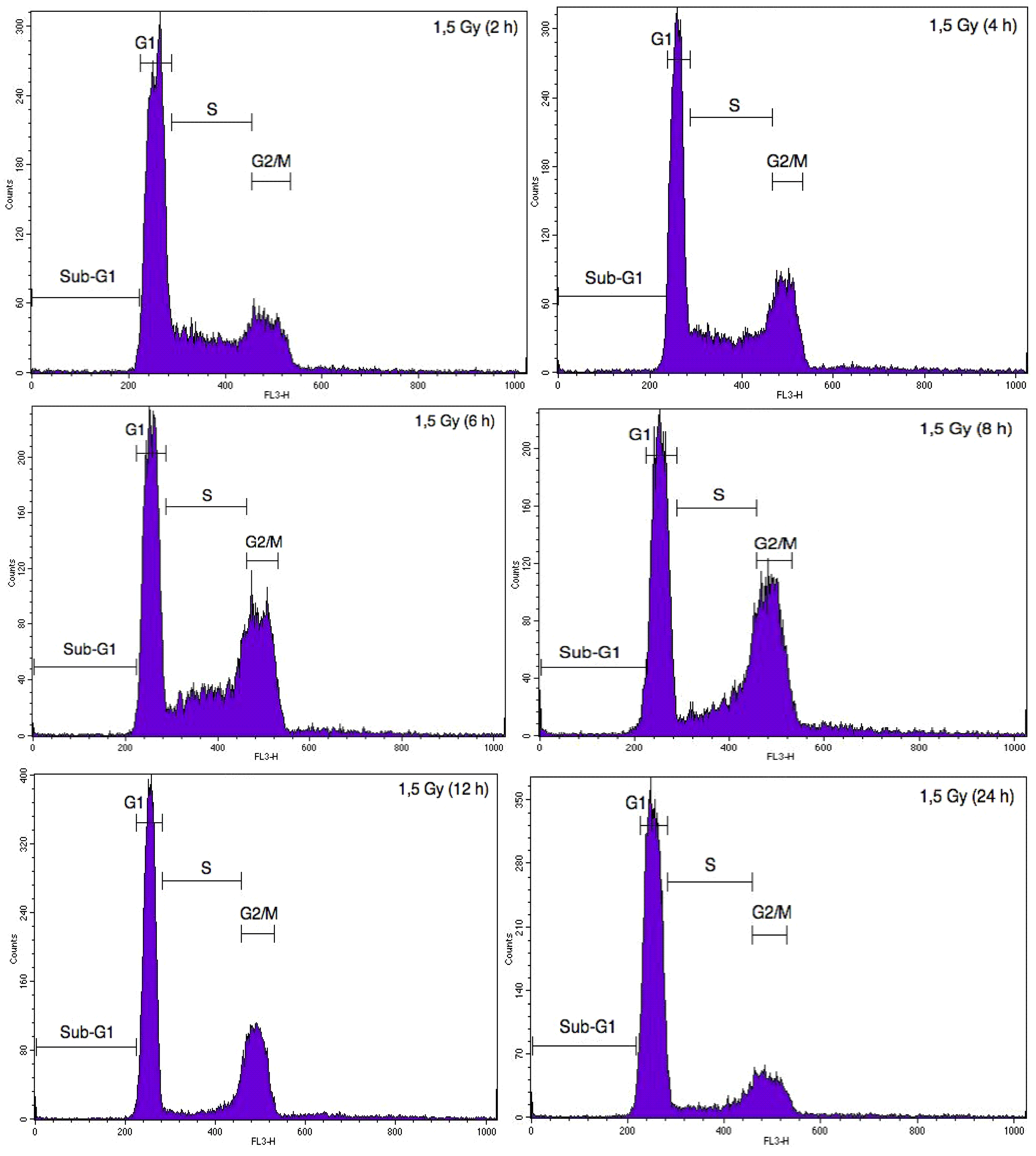
2.2. Cell Cycle Analysis of A549 Cells and EC after Particle Irradiation in Mono- or Co-Culture Configurations

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Alpha particles | G1 (%) | S (%) | G2/M (%) | |
| Mono-culture | CTL A549 | 38.9 | 48.9 | 12.1 |
| 1 Gy A549* | 38.4 | 22.5 | 39.2 | |
| Co-culture | CTL A549/EC | 51.3 | 40.6 | 8.6 |
| 1 Gy A549*/EC* | 44.9 | 32.2 | 22.9 | |
| 1 Gy A549*/EC | 45.4 | 30.2 | 24.1 | |
| 1 Gy A549/EC* | 50.8 | 44.9 | 4.3 | |
| Proton beam | G1 (%) | S (%) | G2/M (%) | |
| Mono-culture | CTL A549 | 46.4 | 51.2 | 2.3 |
| 1 Gy A549* | 22.9 | 46.7 | 30.4 | |
| Co-culture | CTL A549/EC | 50.3 | 45.2 | 4.5 |
| 1 Gy A549*/EC* | 27.9 | 45.8 | 26.3 | |
| 1 Gy A549*/EC | 36 | 50.5 | 13.4 | |
| 1 Gy A549/EC* | 47.6 | 51.1 | 1.3 | |
| Alpha particles | G1 (%) | S (%) | G2/M (%) | |
| Mono-culture | CTL EC | 78 | 20.5 | 1.6 |
| 1 Gy EC* | 62.1 | 31.4 | 6.6 | |
| Co-culture | CTL A549/EC | 66.4 | 30.8 | 2.8 |
| 1 Gy A549*/EC* | 69.8 | 19.2 | 11 | |
| 1 Gy A549*/EC | 68.1 | 28.9 | 3 | |
| 1 Gy A549/EC* | 66.5 | 28.9 | 4.6 | |
| Proton beam | G1 (%) | S (%) | G2/M (%) | |
| Mono-culture | CTL EC | 72.9 | 23.3 | 3.7 |
| 1 Gy EC* | 68.1 | 24.9 | 7 | |
| Co-culture | CTL A549/EC | 63 | 35.6 | 1.4 |
| 1 Gy A549*/EC* | 56.6 | 38 | 5.5 | |
| 1 Gy A549*/EC | 62.4 | 34.8 | 2.9 | |
| 1 Gy A549/EC* | 57 | 39.8 | 3.23 |
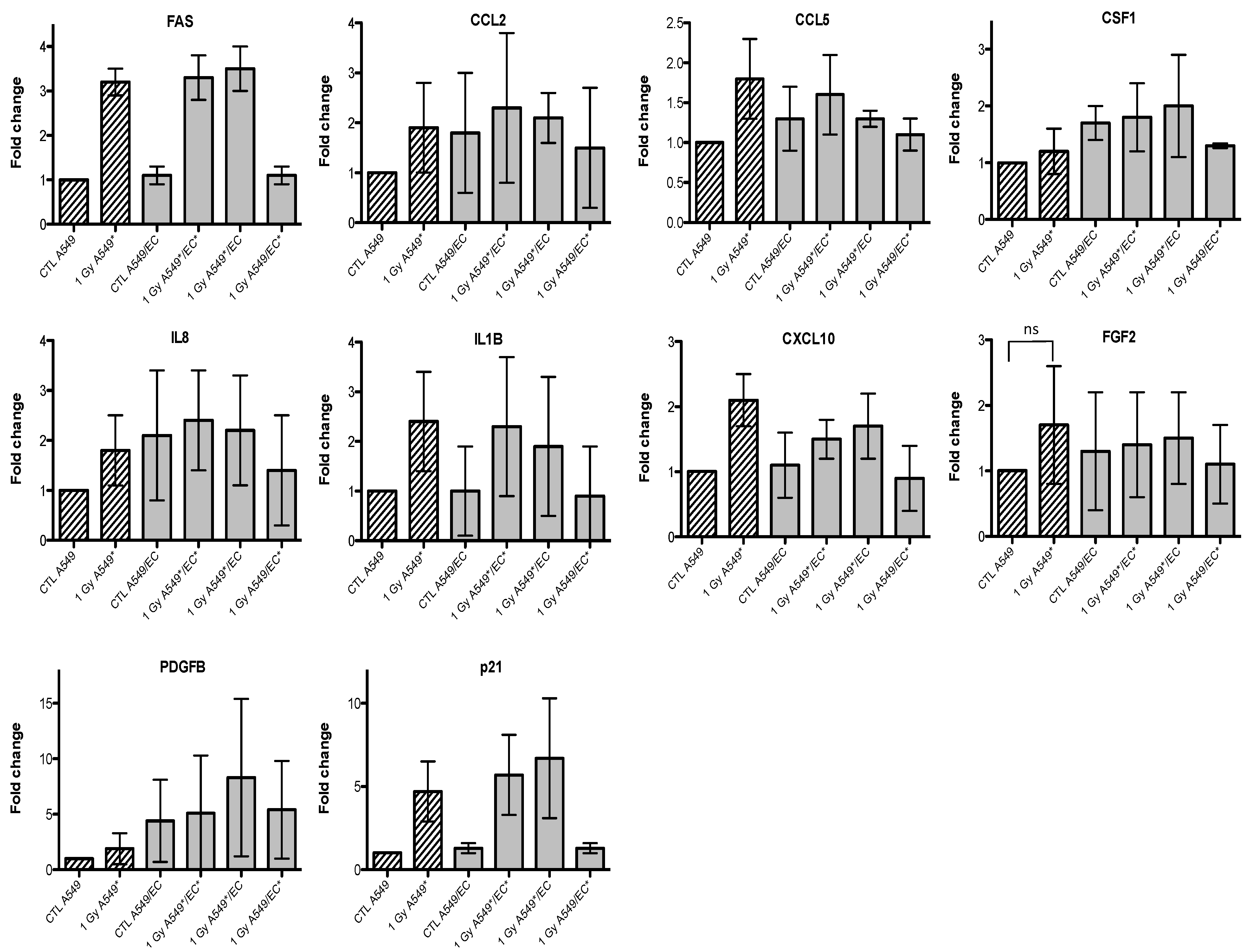
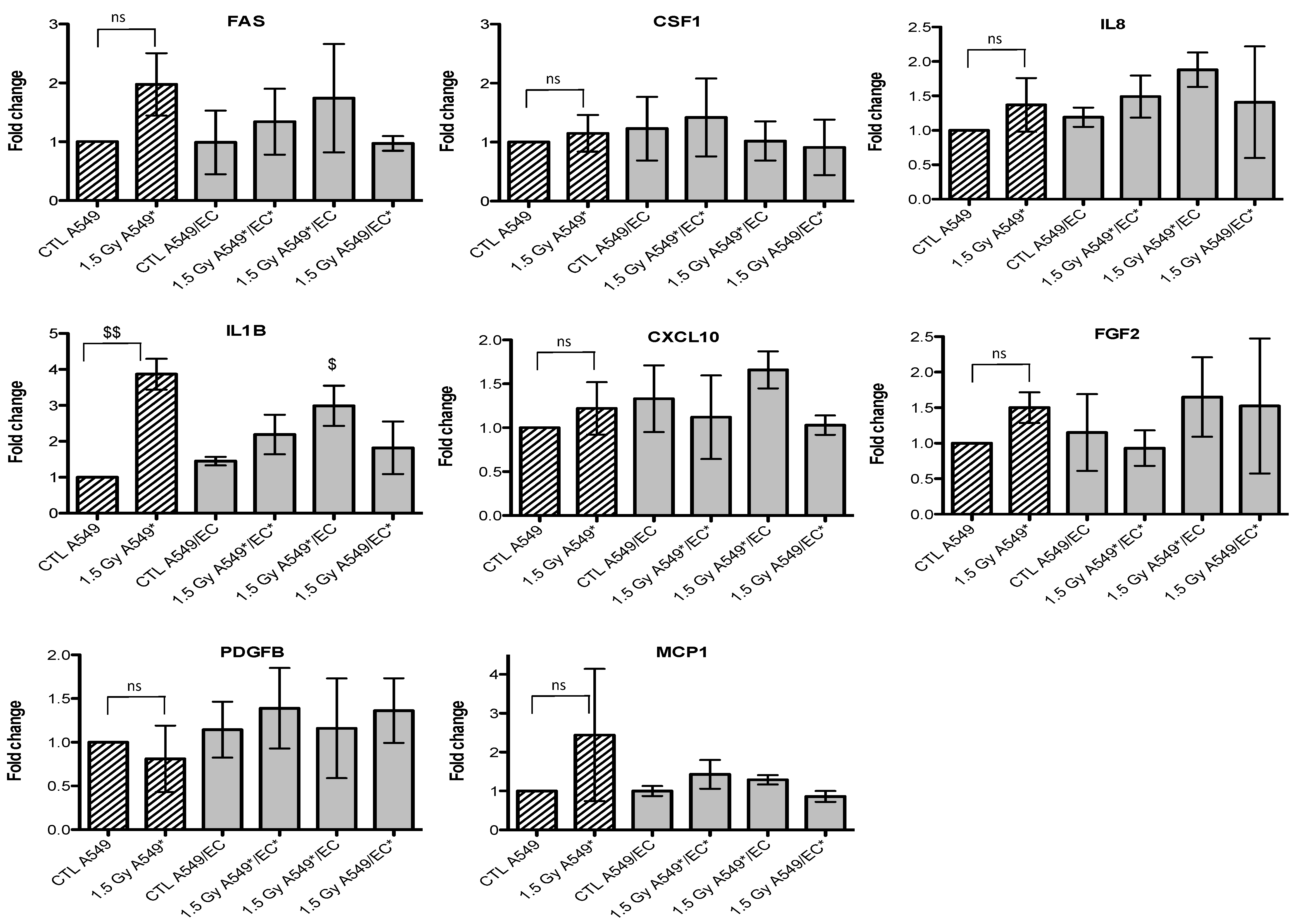
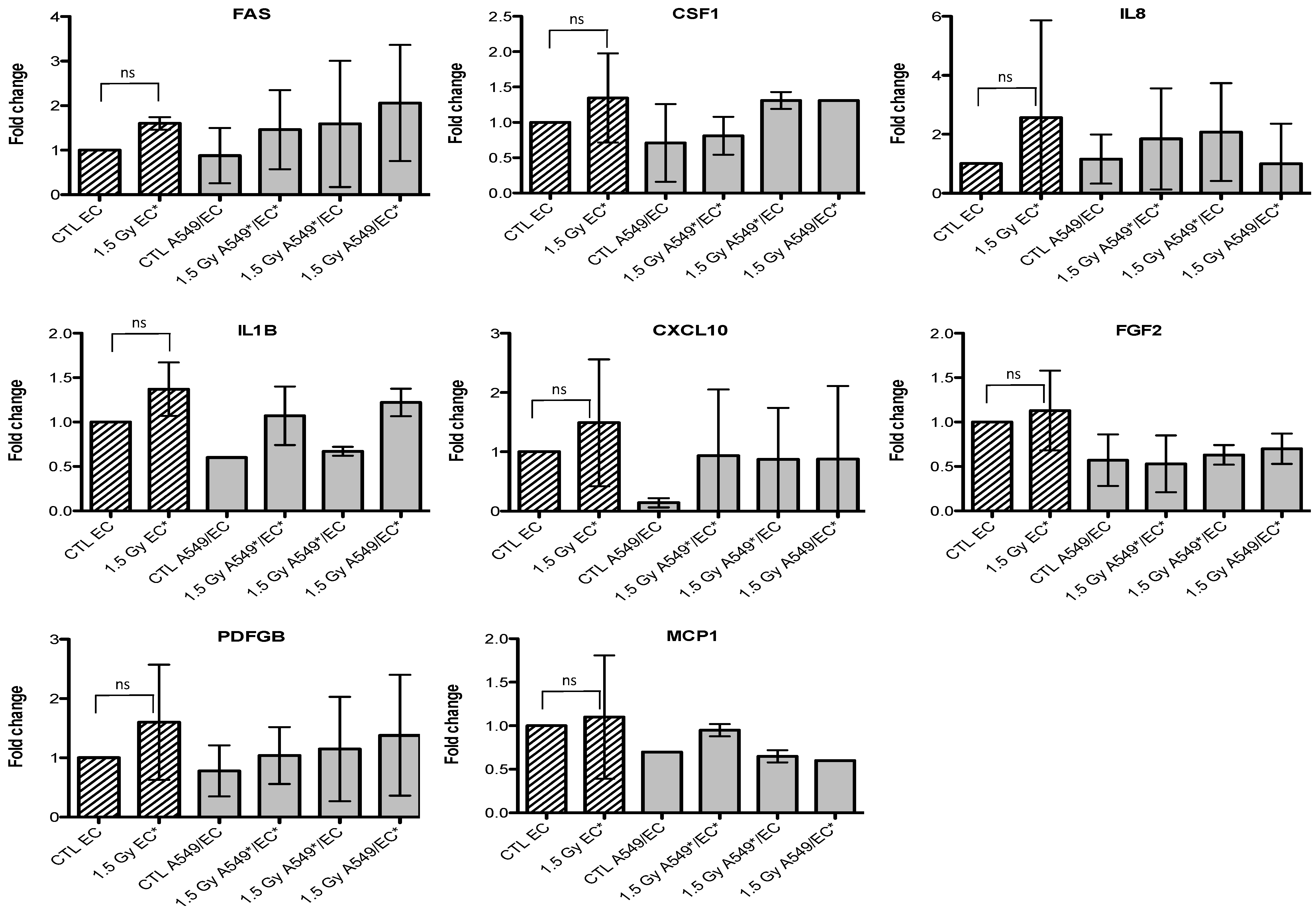
2.3. Effects of Particle Irradiation on Gene Expression in A549 Cells and EC in Mono- or Co-Culture Configurations




3. Discussion
4. Materials and Methods
4.1. Cell Culture
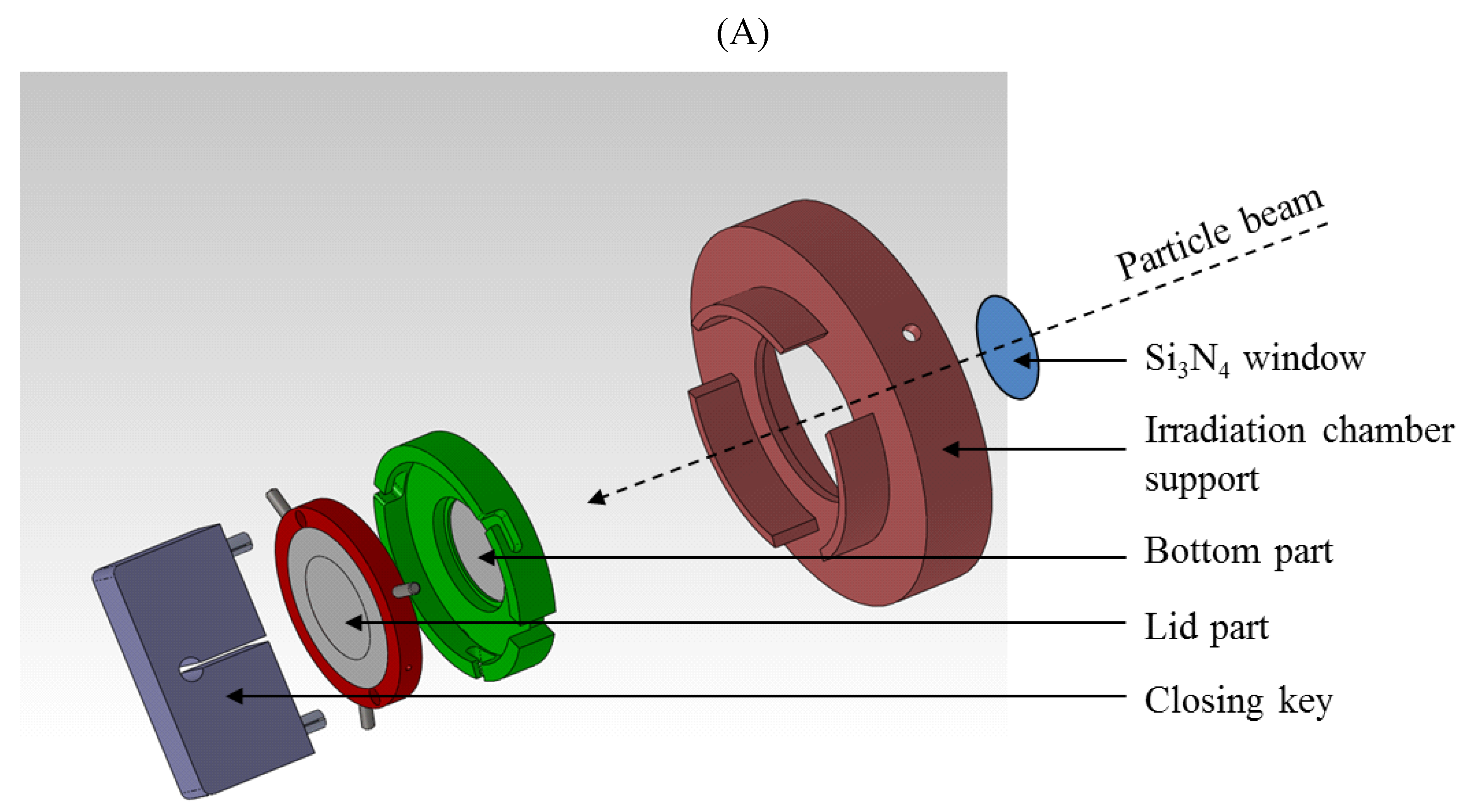
4.2. Irradiation and Co-Culture Irradiation Chambers
4.3. Clonogenic Assay
4.4. Cell Cycle Analysis
4.5. Gene Expression Analysis on TaqMan Low-Density Array
4.6. Quantitative Real-Time PCR
5. Conclusions
Acknowledgements
Author Contributions
Appendix



| Gene Symbol | A549 cell gene expression | EC gene expression | ||||
|---|---|---|---|---|---|---|
| 1 Gy A549*/EC* | 1 Gy A549*/EC | 1 Gy A549/EC* | 1 Gy A549*/EC* | 1 Gy A549*/EC | 1 Gy A549/EC* | |
| ACE | NE | 1.2 | 0.8 | NE | NE | NE |
| AGTR1 | NE | NE | NE | NE | NE | NE |
| AGTR2 | NE | NE | NE | NE | NE | NE |
| BAX | 0.8 | 2.1 | 0.9 | 1.1 | 1.0 | 1.6 |
| BCL2 | 1.0 | 1.1 | 2.0 | NE | 1.1 | NE |
| BCL2L1 | 2.5 | 2.7 | 0.6 | 1.5 | 1.2 | 2.1 |
| C3 | 2.1 | 2.2 | 0.7 | NE | NE | NE |
| CCL19 | NE | NE | NE | NE | NE | NE |
| CCL2 | 2.0 | 2.4 | 5.2 | 1.2 | 1.5 | 1.3 |
| CCL3 | NE | NE | NE | NE | NE | NE |
| CCL5 | 1.8 | 2.0 | 1.2 | 1.5 | 3.9 | 0.9 |
| CCR2 | NE | NE | NE | NE | NE | NE |
| CCR4 | NE | NE | NE | 0.5 | 0.7 | 0.8 |
| CCR5 | NE | NE | NE | NE | NE | NE |
| CCR7 | NE | NE | NE | NE | NE | NE |
| CD19 | NE | NE | NE | NE | NE | NE |
| CD28 | NE | NE | NE | NE | NE | NE |
| CD34 | NE | NE | NE | 0.8 | NE | 1.3 |
| CD38 | 1.9 | 2.1 | 0.9 | NE | NE | NE |
| CD3E | NE | NE | NE | NE | NE | NE |
| CD4 | NE | NE | NE | NE | NE | NE |
| CD40 | 1.4 | 2.4 | 1.8 | 1.2 | 0.9 | 1.4 |
| CD40LG | NE | NE | NE | NE | NE | NE |
| CD68 | 1.2 | 2.0 | 1.0 | 0.9 | 1.0 | 1.4 |
| CD80 | NE | NE | NE | NE | NE | NE |
| CD86 | NE | NE | NE | NE | NE | NE |
| CD8A | 0.5 | 0.5 | 0.5 | NE | NE | NE |
| COL4A5 | 2.0 | 2.3 | 0.6 | 1.0 | 0.9 | 1.3 |
| CSF1 | 2.0 | 2.1 | 1.0 | 1.3 | 1.6 | 1.3 |
| CSF2 | 9.5 | 3.4 | 2.7 | 0.9 | 0.9 | 1.1 |
| CSF3 | NE | NE | NE | 1.5 | 1.7 | 1.8 |
| CTLA4 | NE | NE | NE | NE | NE | NE |
| CXCL10 | 3.4 | 4.4 | 1.1 | 10.5 | 5.5 | 7.8 |
| CXCL11 | 1.2 | 2.1 | 0.9 | 7.3 | 4.7 | 3.2 |
| CXCR3 | NE | NE | NE | NE | NE | NE |
| CYP1A2 | NE | NE | NE | NE | NE | NE |
| CYP7A1 | NE | NE | NE | NE | NE | NE |
| ECE1 | 1.2 | 1.3 | 0.6 | 0.9 | 0.8 | 1.1 |
| EDN1 | 1.4 | 1.3 | 0.9 | 1.0 | 0.8 | 1.0 |
| FAS | 6.4 | 5.0 | 1.0 | 1.4 | 0.8 | 1.2 |
| FASLG | NE | NE | NE | NE | NE | NE |
| FN1 | 1.1 | 1.4 | 0.9 | 1.4 | 2.1 | 1.3 |
| GNLY | NE | NE | NE | NE | NE | NE |
| GZMB | NE | NE | NE | NE | NE | NE |
| HLA-DRA | NE | 1.1 | 1.0 | 0.9 | 0.8 | 1.0 |
| HLA-DRB1 | NE | NE | NE | NE | NE | NE |
| HMOX1 | 0.0 | 1.3 | 1.1 | 1.8 | 2.1 | 1.2 |
| ICAM1 | NE | 1.7 | 1.1 | 0.8 | 0.8 | 0.5 |
| ICOS | NE | NE | NE | NE | NE | NE |
| IFNG | NE | NE | NE | NE | NE | NE |
| IKBKB | 0.8 | 0.8 | 0.6 | 0.7 | 0.9 | 0.8 |
| IL10 | NE | NE | NE | NE | NE | NE |
| IL12A | 1.7 | 0.9 | 0.9 | 1.9 | 1.3 | 1.4 |
| IL12B | NE | NE | NE | NE | NE | NE |
| IL13 | NE | NE | NE | NE | NE | NE |
| IL15 | NE | NE | NE | 1.1 | 1.1 | NE |
| IL17 | NE | NE | NE | NE | NE | NE |
| IL18 | 1.3 | 2.1 | 2.1 | 0.7 | 0.8 | 1.3 |
| IL1A | 2.6 | 2.5 | 1.5 | 0.9 | 0.8 | 0.7 |
| IL1B | 3.0 | 3.1 | 0.7 | 0.8 | 1.0 | 0.6 |
| IL2 | NE | NE | NE | NE | NE | NE |
| IL2RA | NE | NE | NE | NE | NE | NE |
| IL3 | NE | NE | NE | NE | NE | NE |
| IL4 | NE | NE | NE | NE | NE | NE |
| IL5 | NE | NE | NE | NE | NE | NE |
| IL6 | 3.1 | 3.1 | 1.3 | 1.1 | 1.1 | 1.2 |
| IL7 | NE | NE | NE | 1.1 | 1.3 | NE |
| IL8 | 1.8 | 1.7 | 1.0 | 0.5 | 1.5 | 0.4 |
| IL9 | NE | NE | NE | NE | NE | NE |
| LRP2 | NE | NE | NE | NE | NE | NE |
| LTA | NE | NE | NE | 8.2 | 5.8 | 8.7 |
| MYH6 | NE | NE | NE | NE | NE | NE |
| NFKB2 | 1.4 | 1.4 | 0.9 | 0.8 | 1.1 | 0.6 |
| NOS2A | NE | NE | NE | NE | NE | NE |
| PRF1 | NE | NE | NE | NE | NE | NE |
| PTGS2 | NE | NE | NE | NE | NE | NE |
| PTPRC | NE | NE | NE | NE | NE | NE |
| REN | NE | NE | NE | NE | NE | NE |
| RPL3L | NE | NE | NE | 0.4 | 0.3 | 0.4 |
| SELE | NE | NE | NE | NE | NE | NE |
| SELP | NE | NE | NE | 0.9 | 0.8 | 0.9 |
| SKI | 0.8 | 1.0 | 0.9 | 0.7 | 0.9 | 1.2 |
| SMAD3 | 0.7 | 0.6 | 0.4 | 1.0 | 0.9 | 1.4 |
| SMAD7 | 0.9 | 1.1 | 1.2 | 0.5 | 0.9 | 0.4 |
| STAT3 | 1.3 | 1.6 | 0.8 | 1.0 | 0.9 | 1.5 |
| TBX21 | NE | NE | NE | NE | NE | NE |
| TGFB1 | 1.0 | 1.2 | 0.8 | NE | NE | NE |
| TNF | 1.6 | 1.1 | NE | NE | NE | NE |
| TNFRSF18 | NE | NE | NE | NE | NE | NE |
| VEGF | 1.2 | 1.4 | 1.0 | 1.2 | 1.0 | 1.0 |
Conflicts of Interest
References
- Delaney, G.; Jacob, S.; Featherstone, C.; Barton, M. The role of radiotherapy in cancer treatment: Estimating optimal utilization from a review of evidence-based clinical guidelines. Cancer 2005, 104, 1129–1137. [Google Scholar] [CrossRef] [PubMed]
- Halperin, E.C. Particle therapy and treatment of cancer. Lancet Oncol. 2006, 7, 676–685. [Google Scholar] [CrossRef] [PubMed]
- Van der Kogel, A.; Joiner, M.C. Basic Clinical Radiobiology; Hodder Education: London, UK, 2009. [Google Scholar]
- Hada, M.; Georgakilas, A.G. Formation of clustered DNA damage after high-let irradiation: A review. J. Radiat. Res. 2008, 49, 203–210. [Google Scholar] [CrossRef] [PubMed]
- Durante, M.; Loeffler, J.S. Charged particles in radiation oncology. Nat. Rev. Clin. Oncol. 2010, 7, 37–43. [Google Scholar] [CrossRef] [PubMed]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Barros, M.; Paris, F.; Cordon-Cardo, C.; Lyden, D.; Rafii, S.; Haimovitz-Friedman, A.; Fuks, Z.; Kolesnick, R. Tumor response to radiotherapy regulated by endothelial cell apoptosis. Science 2003, 300, 1155–1159. [Google Scholar] [CrossRef] [PubMed]
- Gorski, D.H.; Beckett, M.A.; Jaskowiak, N.T.; Calvin, D.P.; Mauceri, H.J.; Salloum, R.M.; Seetharam, S.; Koons, A.; Hari, D.M.; Kufe, D.W.; et al. Blockage of the vascular endothelial growth factor stress response increases the antitumor effects of ionizing radiation. Cancer Res. 1999, 59, 3374–3378. [Google Scholar] [PubMed]
- Moeller, B.J.; Cao, Y.; Li, C.Y.; Dewhirst, M.W. Radiation activates hif-1 to regulate vascular radiosensitivity in tumors: Role of reoxygenation, free radicals, and stress granules. Cancer Cell 2004, 5, 429–441. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.H.; Yoo, K.C.; Cui, Y.H.; Uddin, N.; Lim, E.J.; Kim, M.J.; Nam, S.Y.; Kim, I.G.; Suh, Y.; Lee, S.J. Radiation promotes malignant progression of glioma cells through hif-1 alpha stabilization. Cancer Lett. 2014, 354, 132–141. [Google Scholar] [CrossRef] [PubMed]
- Borovski, T.; Beke, P.; van Tellingen, O.; Rodermond, H.M.; Verhoeff, J.J.; Lascano, V.; Daalhuisen, J.B.; Medema, J.P.; Sprick, M.R. Therapy-resistant tumor microvascular endothelial cells contribute to treatment failure in glioblastoma multiforme. Oncogene 2013, 32, 1539–1548. [Google Scholar] [CrossRef] [PubMed]
- Kuwahara, Y.; Mori, M.; Kitahara, S.; Fukumoto, M.; Ezaki, T.; Mori, S.; Echigo, S.; Ohkubo, Y.; Fukumoto, M. Targeting of tumor endothelial cells combining 2 gy/day of x-ray with everolimus is the effective modality for overcoming clinically relevant radioresistant tumors. Cancer Med. 2014, 3, 310–321. [Google Scholar] [CrossRef] [PubMed]
- Tavora, B.; Reynolds, L.E.; Batista, S.; Demircioglu, F.; Fernandez, I.; Lechertier, T.; Lees, D.M.; Wong, P.P.; Alexopoulou, A.; Elia, G.; et al. Endothelial-cell fak targeting sensitizes tumours to DNA-damaging therapy. Nature 2014, 514, 112–116. [Google Scholar] [CrossRef] [PubMed]
- Camacho, M.; Pineiro, Z.; Alcolea, S.; Garcia, J.; Balart, J.; Terra, X.; Aviles-Jurado, F.X.; Soler, M.; Quer, M.; Leon, X.; et al. Prostacyclin-synthase expression in head and neck carcinoma patients and its prognostic value in the response to radiotherapy. J. Pathol. 2014, 235, 125–135. [Google Scholar] [CrossRef]
- Riquier, H.; Wera, A.C.; Heuskin, A.C.; Feron, O.; Lucas, S.; Michiels, C. Comparison of x-ray and alpha particle effects on a human cancer and endothelial cells: Survival curves and gene expression profiles. Radiother. Oncol. 2013, 106, 397–403. [Google Scholar] [CrossRef] [PubMed]
- Wera, A.-C.; Riquier, H.; Heuskin, A.-C.; Michiels, C.; Lucas, S. In vitro irradiation station for broad beam radiobiological experiments. Nucl. Instrum. Methods Phys. Res. B 2011, 269, 3120–3124. [Google Scholar] [CrossRef]
- Wera, A.C.; Heuskin, A.C.; Riquier, H.; Michiels, C.; Lucas, S. Low-let proton irradiation of a549 non-small cell lung adenocarcinoma cells: Dose response and rbe determination. Radiat. Res. 2013, 179, 273–281. [Google Scholar] [CrossRef] [PubMed]
- Heuskin, A.C.; Wera, A.C.; Riquier, H.; Michiels, C.; Lucas, S. Low-dose hypersensitivity and bystander effect are not mutually exclusive in a549 lung carcinoma cells after irradiation with charged particles. Radiat. Res. 2013, 180, 491–498. [Google Scholar] [CrossRef] [PubMed]
- Lobrich, M.; Jeggo, P.A. The impact of a negligent g2/m checkpoint on genomic instability and cancer induction. Nat. Rev. Cancer 2007, 7, 861–869. [Google Scholar] [CrossRef] [PubMed]
- Kargiotis, O.; Geka, A.; Rao, J.S.; Kyritsis, A.P. Effects of irradiation on tumor cell survival, invasion and angiogenesis. J. Neurooncol. 2010, 100, 323–338. [Google Scholar] [CrossRef] [PubMed]
- Brown, C.K.; Khodarev, N.N.; Yu, J.; Moo-Young, T.; Labay, E.; Darga, T.E.; Posner, M.C.; Weichselbaum, R.R.; Mauceri, H.J. Glioblastoma cells block radiation-induced programmed cell death of endothelial cells. FEBS Lett. 2004, 565, 167–170. [Google Scholar] [CrossRef] [PubMed]
- Moeller, B.J.; Dewhirst, M.W. Raising the bar: How hif-1 helps determine tumor radiosensitivity. Cell Cycle 2004, 3, 1107–1110. [Google Scholar] [CrossRef] [PubMed]
- Fokas, E.; Kraft, G.; An, H.; Engenhart-Cabillic, R. Ion beam radiobiology and cancer: Time to update ourselves. Biochim. Biophys. Acta 2009, 1796, 216–229. [Google Scholar] [PubMed]
- Habrand, J.L.; Schneider, R.; Alapetite, C.; Feuvret, L.; Petras, S.; Datchary, J.; Grill, J.; Noel, G.; Helfre, S.; Ferrand, R.; et al. Proton therapy in pediatric skull base and cervical canal low-grade bone malignancies. Int. J. Radiat. Oncol. Biol. Phys. 2008, 71, 672–675. [Google Scholar] [CrossRef] [PubMed]
- Olsen, D.R.; Overgaard, J. Leveraging clinical performance by technological excellence—The case of particle therapy. Radiother. Oncol. 2010, 95, 1–2. [Google Scholar] [CrossRef] [PubMed]
- De Ruysscher, D.; Mark Lodge, M.; Jones, B.; Brada, M.; Munro, A.; Jefferson, T.; Pijls-Johannesma, M. Charged particles in radiotherapy: A 5-year update of a systematic review. Radiother. Oncol. 2012, 103, 5–7. [Google Scholar] [CrossRef] [PubMed]
- Goitein, M. Trials and tribulations in charged particle radiotherapy. Radiother. Oncol. 2010, 95, 23–31. [Google Scholar] [CrossRef] [PubMed]
- Folkman, J. Looking for a good endothelial address. Cancer Cell 2002, 1, 113–115. [Google Scholar] [CrossRef] [PubMed]
- Begg, A.C.; Stewart, F.A.; Vens, C. Strategies to improve radiotherapy with targeted drugs. Nat. Rev. Cancer 2011, 11, 239–253. [Google Scholar] [CrossRef] [PubMed]
- Hamada, N.; Hara, T.; Omura-Minamisawa, M.; Funayama, T.; Sakashita, T.; Sora, S.; Yokota, Y.; Nakano, T.; Kobayashi, Y. Energetic heavy ions overcome tumor radioresistance caused by overexpression of bcl-2. Radiother. Oncol. 2008, 89, 231–236. [Google Scholar] [CrossRef] [PubMed]
- Gerashchenko, B.I.; Howell, R.W. Cell proximity is a prerequisite for the proliferative response of bystander cells co-cultured with cells irradiated with gamma-rays. Cytometry A 2003, 56, 71–80. [Google Scholar] [CrossRef] [PubMed]
- Heuskin, A.C.; Michiels, C.; Lucas, S. Toward computer simulation of high-let in vitro survival curves. Phys. Med. Biol. 2013, 58, 6495–6510. [Google Scholar] [CrossRef] [PubMed]
- Rashi-Elkeles, S.; Elkon, R.; Shavit, S.; Lerenthal, Y.; Linhart, C.; Kupershtein, A.; Amariglio, N.; Rechavi, G.; Shamir, R.; Shiloh, Y. Transcriptional modulation induced by ionizing radiation: P53 remains a central player. Mol. Oncol. 2011, 5, 336–348. [Google Scholar] [CrossRef] [PubMed]
- Girdhani, S.; Lamont, C.; Hahnfeldt, P.; Abdollahi, A.; Hlatky, L. Proton irradiation suppresses angiogenic genes and impairs cell invasion and tumor growth. Radiat. Res. 2012, 178, 33–45. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Liu, Y.; Zhang, H.; Sun, C.; Zhao, Q.; Di, C.; Li, H.; Gan, L.; Wang, Y. Effects of carbon-ion beam irradiation on the angiogenic response in lung adenocarcinoma a549 cells. Cell Biol. Int. 2014, 38, 1304–1310. [Google Scholar] [CrossRef] [PubMed]
- Franken, N.A.; Rodermond, H.M.; Stap, J.; Haveman, J.; van Bree, C. Clonogenic assay of cells in vitro. Nat. Protoc. 2006, 1, 2315–2319. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Riquier, H.; Abel, D.; Wera, A.-C.; Heuskin, A.-C.; Genard, G.; Lucas, S.; Michiels, C. Effects of Alpha Particle and Proton Beam Irradiation as Putative Cross-Talk between A549 Cancer Cells and the Endothelial Cells in a Co-Culture System. Cancers 2015, 7, 481-502. https://doi.org/10.3390/cancers7010481
Riquier H, Abel D, Wera A-C, Heuskin A-C, Genard G, Lucas S, Michiels C. Effects of Alpha Particle and Proton Beam Irradiation as Putative Cross-Talk between A549 Cancer Cells and the Endothelial Cells in a Co-Culture System. Cancers. 2015; 7(1):481-502. https://doi.org/10.3390/cancers7010481
Chicago/Turabian StyleRiquier, Hélène, Denis Abel, Anne-Catherine Wera, Anne-Catherine Heuskin, Géraldine Genard, Stéphane Lucas, and Carine Michiels. 2015. "Effects of Alpha Particle and Proton Beam Irradiation as Putative Cross-Talk between A549 Cancer Cells and the Endothelial Cells in a Co-Culture System" Cancers 7, no. 1: 481-502. https://doi.org/10.3390/cancers7010481
APA StyleRiquier, H., Abel, D., Wera, A. -C., Heuskin, A. -C., Genard, G., Lucas, S., & Michiels, C. (2015). Effects of Alpha Particle and Proton Beam Irradiation as Putative Cross-Talk between A549 Cancer Cells and the Endothelial Cells in a Co-Culture System. Cancers, 7(1), 481-502. https://doi.org/10.3390/cancers7010481