
Cimetidine and Clobenpropit Attenuate Inflammation-Associated Colorectal Carcinogenesis in Male ICR Mice

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals, Chemicals and Diet
2.2. Experiment 1 (the Eighteen-Week Experiment)
2.2.1. Counting the Number of Mucosal Ulcers in the Colorectum
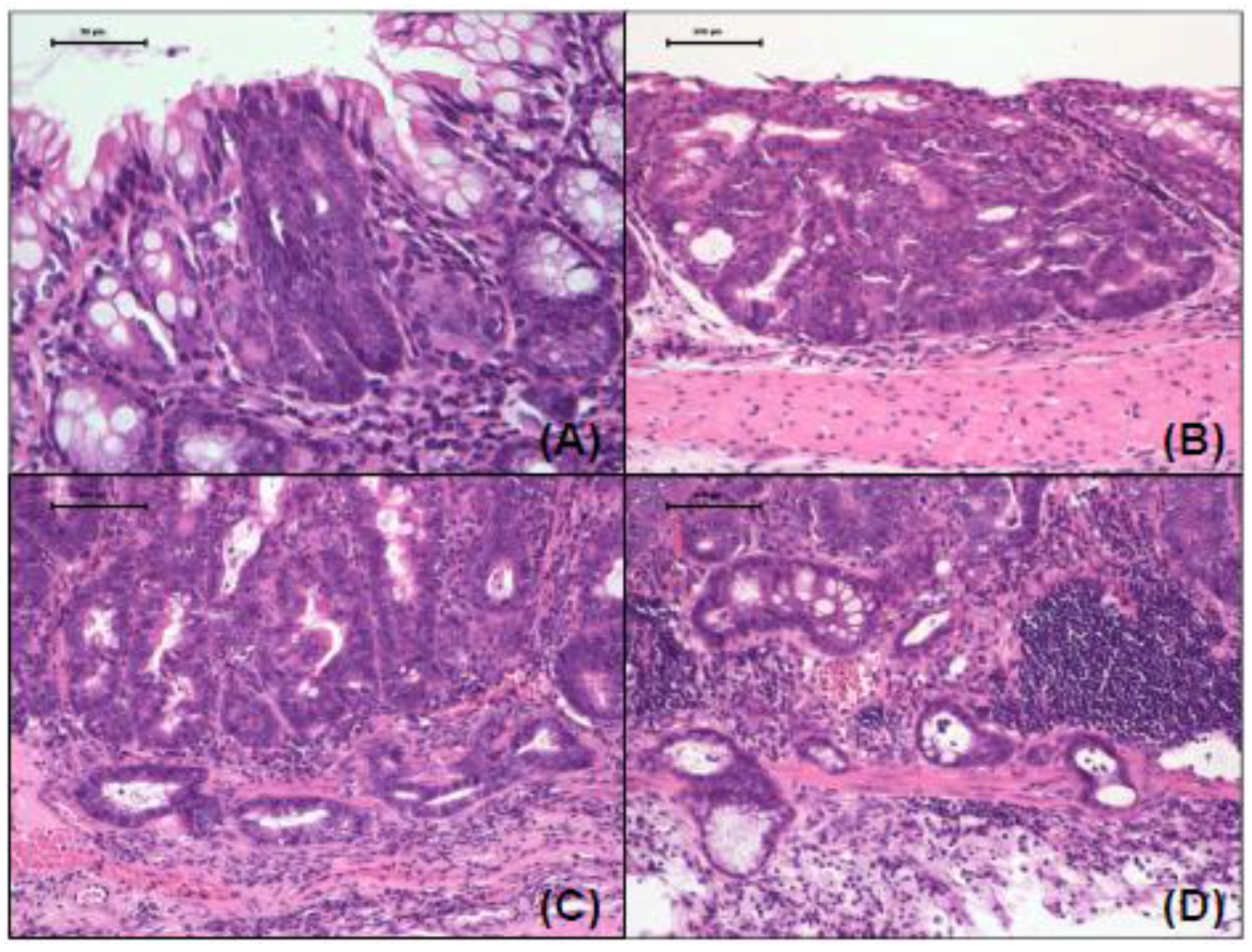
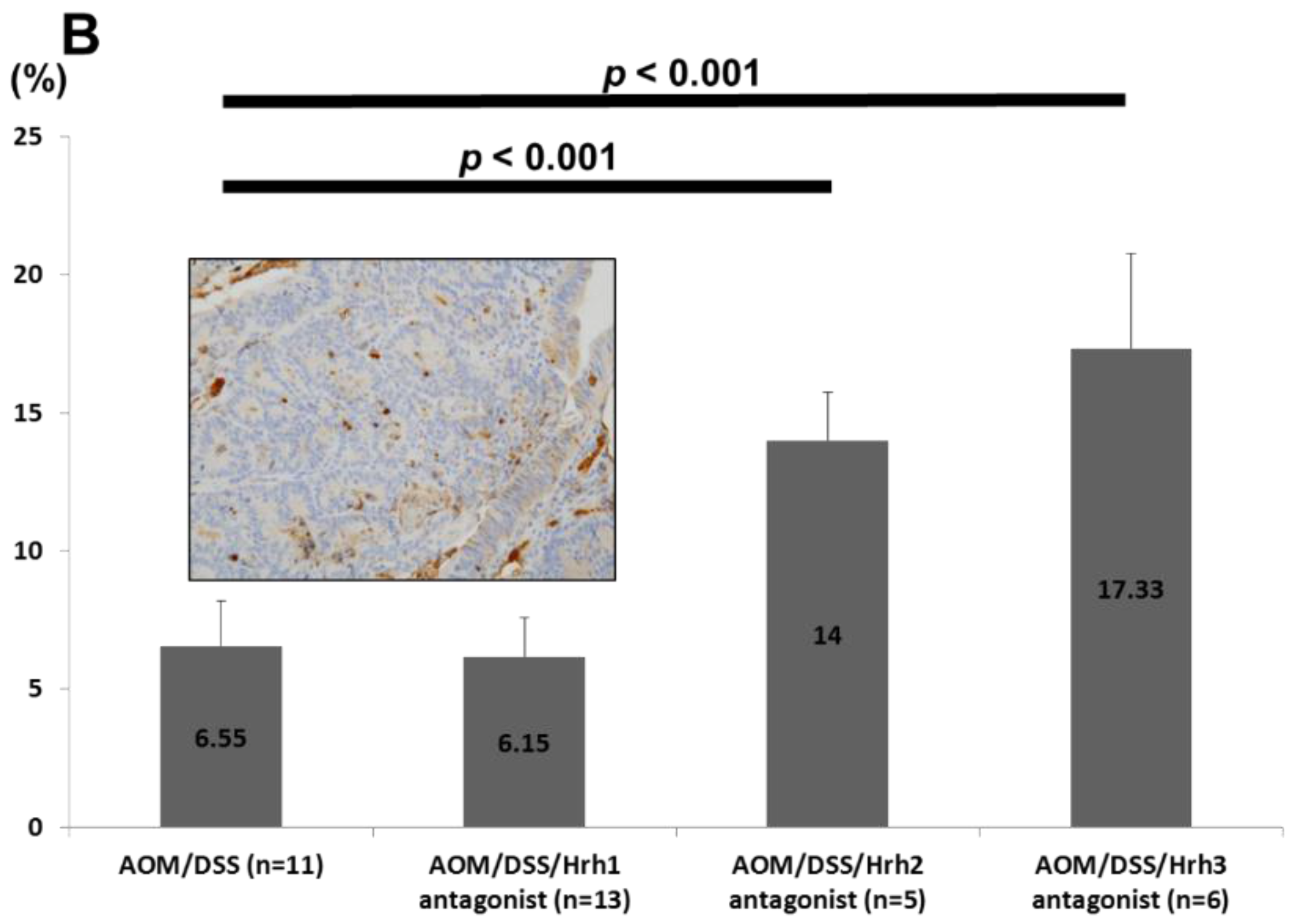
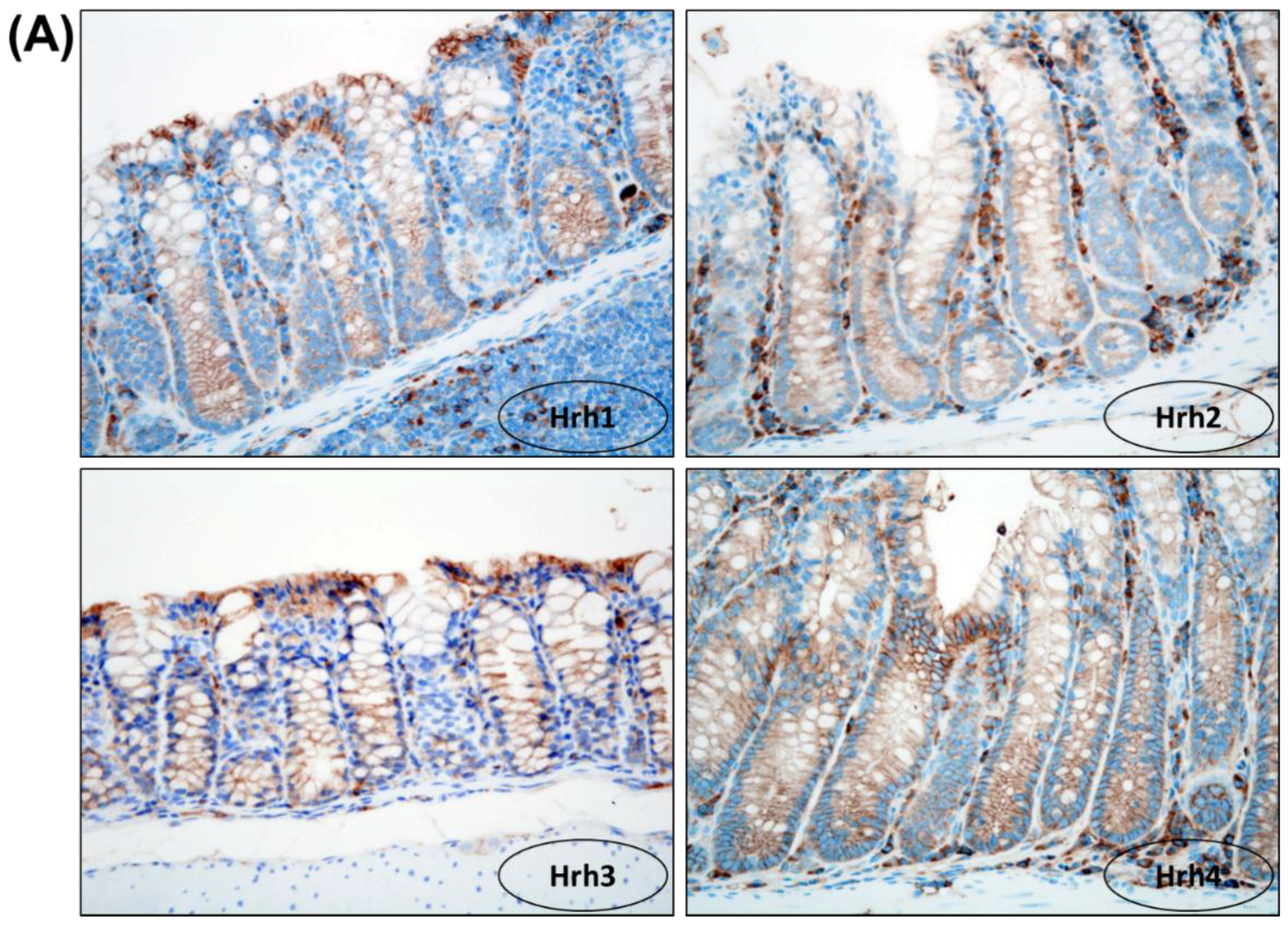
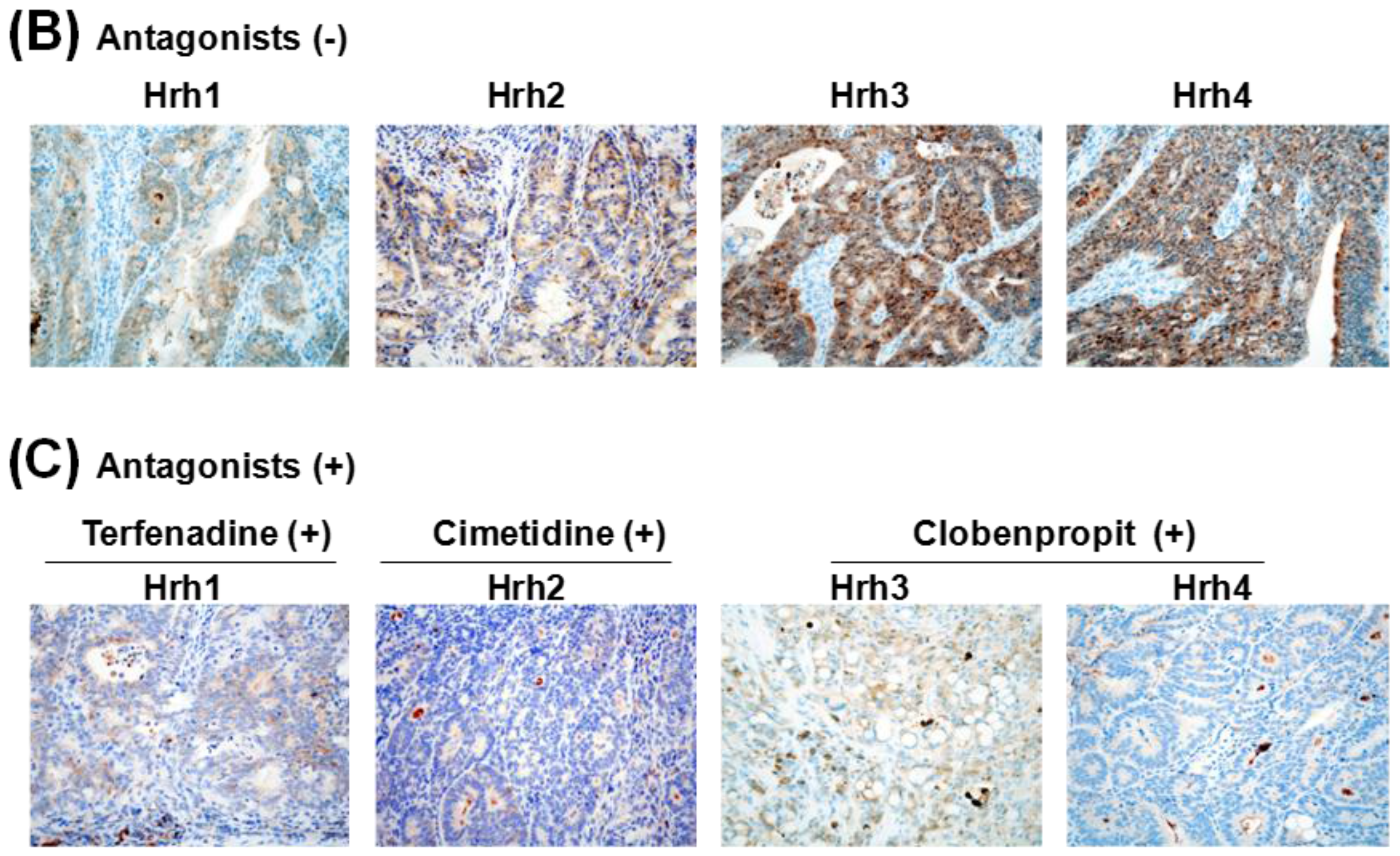
2.2.2. Immunohistochemical Analysis of Minichromosome Maintenance Protein 2 (MCM2), Cleaved Caspase-3, Hrh1, Hrh2, Hrh3 and Hrh4 in Colonic Adenocarcinoma
2.2.3. Oxidative Stress and Antioxidant Status Measurements
2.3. Experiment 2 (the Four-Week Experiment)
2.4. Total RNA Extraction and Quantitative Real-Time (RT)-PCR
2.5. Statistical Analysis
3. Results
3.1. Experiment 1 (the Eighteen-Week Experiment)
3.1.1. General Observations in Experiment 1 (18-Week Study)
3.1.2. Incidences and Multiplicities of Colorectal Mucosal Ulcers, High-Grade Dysplasia, Adenoma and Adenocarcinoma
3.1.3. Biomarkers of Oxidative Stress and Antioxidant Defense Status
3.1.4. Immunohistochemical Analysis of MCM2 (Cell Proliferation) and Caspase-3 (Apoptosis) in Colorectal Adenocarcinomas
3.1.5. Immunohistochemical Expression of Hrh1, Hrh2, Hrh3 and Hrh4 in Adenocarcinomas
3.2. Experiment 2 (the Four-Week Experiment)
3.2.1. General Observations
3.2.2. Biomarkers of Oxidative Stress and Antioxidant Defense Status
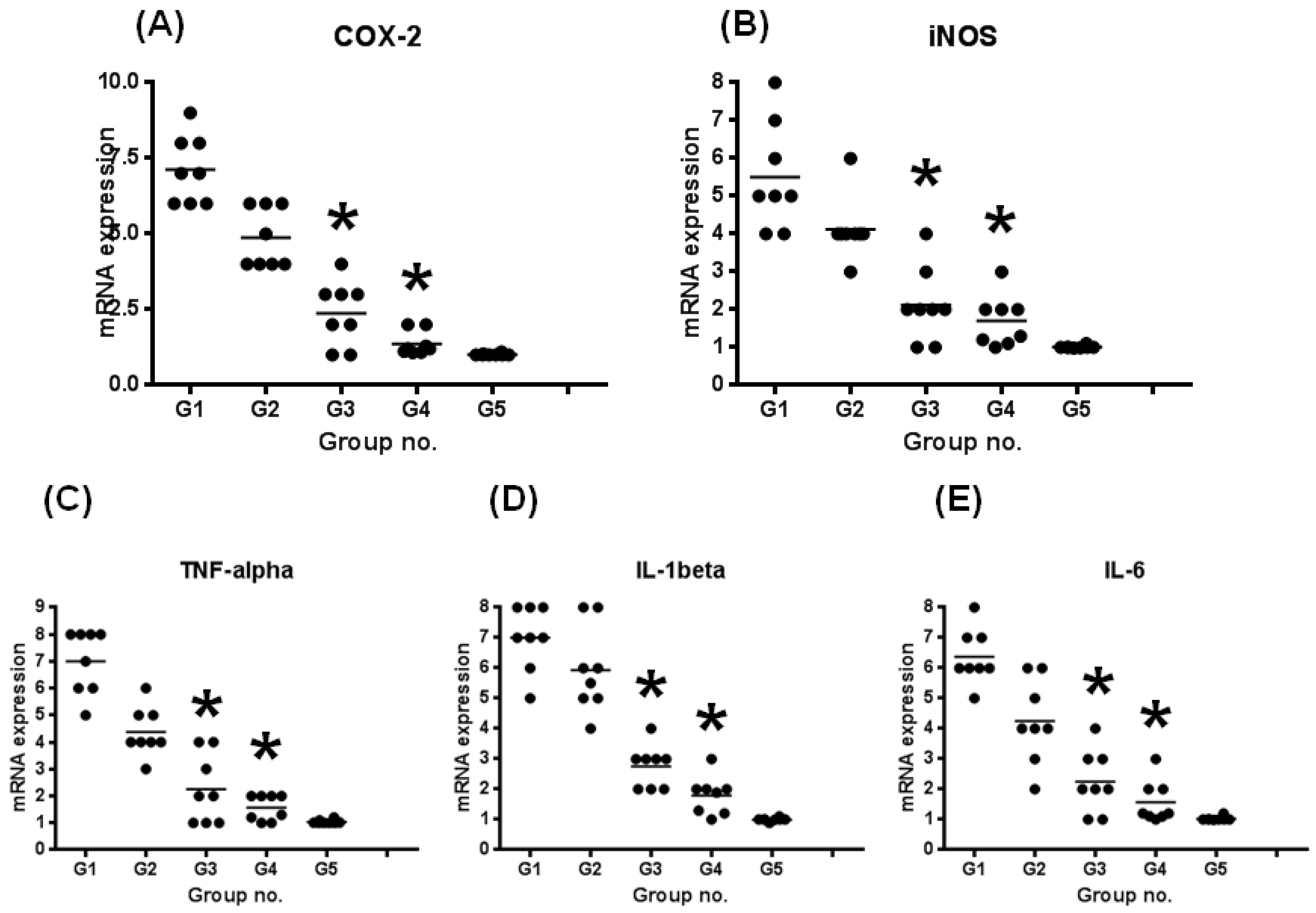
3.2.3. mRNA Expression Levels of Inducible Inflammatory Enzymes and Proinflammatory Cytokines
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Balkwill, F.; Mantovani, A. Inflammation and cancer: Back to Virchow? Lancet 2001, 357, 539–545. [Google Scholar] [CrossRef]
- Tanaka, T.; Suzuki, R. Inflammation and Cancer. In Cancer: Disease Progression and Chemoprevention; Tanaka, T., Ed.; Research Signpost: Kerala, India, 2007; pp. 27–44. [Google Scholar]
- Ullman, T.A.; Itzkowitz, S.H. Intestinal inflammation and cancer. Gastroenterology 2011, 140, 1807–1816. [Google Scholar] [CrossRef] [PubMed]
- Munkholm, P. Review article: The incidence and prevalence of colorectal cancer in inflammatory bowel disease. Aliment. Pharmacol. Ther. 2003, 18, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Eaden, J.A.; Abrams, K.R.; Mayberry, J.F. The risk of colorectal cancer in ulcerative colitis: A meta-analysis. Gut 2001, 48, 526–535. [Google Scholar] [CrossRef] [PubMed]
- Canavan, C.; Abrams, K.R.; Mayberry, J. Meta-analysis: Colorectal and small bowel cancer risk in patients with Crohn’s disease. Aliment. Pharmacol. Ther. 2006, 23, 1097–1104. [Google Scholar] [CrossRef] [PubMed]
- Sung, J.J.; Lau, J.Y.; Goh, K.L.; Leung, W.K. Increasing incidence of colorectal cancer in Asia: Implications for screening. Lancet Oncol. 2005, 6, 871–876. [Google Scholar] [CrossRef]
- Yasui, Y.; Kim, M.; Oyama, T.; Tanaka, T. Colorectal carcinogenesis and suppression of tumor development by inhibition of enzymes and molecular targets. Curr. Enzym. Inhib. 2009, 5, 1–26. [Google Scholar] [CrossRef]
- Tanaka, T.; Kohno, H.; Suzuki, R.; Yamada, Y.; Sugie, S.; Mori, H. A novel inflammation-related mouse colon carcinogenesis model induced by azoxymethane and dextran sodium sulfate. Cancer Sci. 2003, 94, 965–973. [Google Scholar] [CrossRef] [PubMed]
- Kohno, H.; Suzuki, R.; Yasui, Y.; Miyamoto, S.; Wakabayashi, K.; Tanaka, T. Ursodeoxycholic acid versus sulfasalazine in colitis-related colon carcinogenesis in mice. Clin. Cancer Res. 2007, 13, 2519–2525. [Google Scholar] [CrossRef] [PubMed]
- Walter, M.; Stark, H. Histamine receptor subtypes: A century of rational drug design. Front. Biosci. (Schol. Ed.) 2012, 4, 461–488. [Google Scholar] [CrossRef] [PubMed]
- Cataldi, M.; Borriello, F.; Granata, F.; Annunziato, L.; Marone, G. Histamine receptors and antihistamines: From discovery to clinical applications. Chem. Immunol. Allergy 2014, 100, 214–226. [Google Scholar] [PubMed]
- Xie, H.; He, S.H. Roles of histamine and its receptors in allergic and inflammatory bowel diseases. World J. Gastroenterol. 2005, 11, 2851–2857. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, T.; Ishikawa, H. Mast cells and inflammation-associated colorectal carcinogenesis. Springer Semin. Immunopathol. 2013, 35, 245–254. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, T. Development of an inflammation-associated colorectal cancer model and its application for research on carcinogenesis and chemoprevention. Int. J. Inflam. 2012. [Google Scholar] [CrossRef] [PubMed]
- Azer, S.A. Overview of molecular pathways in inflammatory bowel disease associated with colorectal cancer development. Eur. J. Gastroenterol. Hepatol. 2013, 25, 271–281. [Google Scholar] [CrossRef] [PubMed]
- Terhaar Sive Droste, J.S.; Tuynman, J.B.; van Dullemen, H.M.; Mulder, C.J. Chemoprevention for colon cancer: New opportunities, fact or fiction? Scand. J. Gastroenterol. Suppl. 2006, 243, 158–164. [Google Scholar] [CrossRef] [PubMed]
- Ohshima, H.; Tazawa, H.; Sylla, B.S.; Sawa, T. Prevention of human cancer by modulation of chronic inflammatory processes. Mutat. Res. 2005, 591, 110–122. [Google Scholar] [CrossRef] [PubMed]
- Coruzzi, G.; Adami, M.; Pozzoli, C. Role of histamine H4 receptors in the gastrointestinal tract. Front. Biosci. (Schol Ed.) 2012, 4, 226–239. [Google Scholar] [CrossRef] [PubMed]
- Masini, E.; Fabbroni, V.; Giannini, L.; Vannacci, A.; Messerini, L.; Perna, F.; Cortesini, C.; Cianchi, F. Histamine and histidine decarboxylase up-regulation in colorectal cancer: Correlation with tumor stage. Inflamm. Res. 2005, 54, S80–S81. [Google Scholar] [CrossRef] [PubMed]
- Medina, V.A.; Rivera, E.S. Histamine receptors and cancer pharmacology. Br. J. Pharmacol. 2010, 161, 755–767. [Google Scholar] [CrossRef] [PubMed]
- Cianchi, F.; Cortesini, C.; Schiavone, N.; Perna, F.; Magnelli, L.; Fanti, E.; Bani, D.; Messerini, L.; Fabbroni, V.; Perigli, G.; et al. The role of cyclooxygenase-2 in mediating the effects of histamine on cell proliferation and vascular endothelial growth factor production in colorectal cancer. Clin. Cancer Res. 2005, 11, 6807–6815. [Google Scholar] [CrossRef] [PubMed]
- Adams, W.J.; Lawson, J.A.; Morris, D.L. Cimetidine inhibits in vivo growth of human colon cancer and reverses histamine stimulated in vitro and in vivo growth. Gut 1994, 35, 1632–1636. [Google Scholar] [CrossRef] [PubMed]
- Boer, K.; Helinger, E.; Helinger, A.; Pocza, P.; Pos, Z.; Demeter, P.; Baranyai, Z.; Dede, K.; Darvas, Z.; Falus, A. Decreased expression of histamine H1 and H4 receptors suggests disturbance of local regulation in human colorectal tumours by histamine. Eur. J. Cell. Biol. 2008, 87, 227–236. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, K.; Tanaka, S.; Ichikawa, A. Effect of cimetidine on intratumoral cytokine expression in an experimental tumor. Biochem. Biophys. Res. Commun. 2001, 281, 1113–1119. [Google Scholar] [CrossRef] [PubMed]
- Tomita, K.; Izumi, K.; Okabe, S. Roxatidine- and cimetidine-induced angiogenesis inhibition suppresses growth of colon cancer implants in syngeneic mice. J. Pharmacol. Sci. 2003, 93, 321–330. [Google Scholar] [CrossRef] [PubMed]
- Tomita, K.; Okabe, S. Exogenous histamine stimulates colorectal cancer implant growth via immunosuppression in mice. J. Pharmacol. Sci. 2005, 97, 116–123. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.D.; Ai, W.; Asfaha, S.; Bhagat, G.; Friedman, R.A.; Jin, G.; Park, H.; Shykind, B.; Diacovo, T.G.; Falus, A.; et al. Histamine deficiency promotes inflammation-associated carcinogenesis through reduced myeloid maturation and accumulation of CD11b+Ly6G+ immature myeloid cells. Nat. Med. 2011, 17, 87–95. [Google Scholar] [CrossRef] [PubMed]
- Lampiasi, N.; Azzolina, A.; Montalto, G.; Cervello, M. Histamine and spontaneously released mast cell granules affect the cell growth of human hepatocellular carcinoma cells. Exp. Mol. Med. 2007, 39, 284–294. [Google Scholar] [CrossRef] [PubMed]
- Nicolau-Galmes, F.; Asumendi, A.; Alonso-Tejerina, E.; Perez-Yarza, G.; Jangi, S.M.; Gardeazabal, J.; Arroyo-Berdugo, Y.; Careaga, J.M.; Diaz-Ramon, J.L.; Apraiz, A.; et al. Terfenadine induces apoptosis and autophagy in melanoma cells through ROS-dependent and -independent mechanisms. Apoptosis 2011, 16, 1253–1267. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.T.; Chen, Y.H.; Hsu, J.L.; Leu, W.J.; Yu, C.C.; Chan, S.H.; Ho, Y.F.; Hsu, L.C.; Guh, J.H. Terfenadine induces anti-proliferative and apoptotic activities in human hormone-refractory prostate cancer through histamine receptor-independent Mcl-1 cleavage and Bak up-regulation. Naunyn Schmiedebergs Arch. Pharmacol. 2014, 387, 33–45. [Google Scholar] [CrossRef] [PubMed]
- Fang, Z.; Yao, W.; Xiong, Y.; Li, J.; Liu, L.; Shi, L.; Zhang, W.; Zhang, C.; Nie, L.; Wan, J. Attenuated expression of HRH4 in colorectal carcinomas: a potential influence on tumor growth and progression. BMC Cancer 2011. [Google Scholar] [CrossRef] [PubMed]
- Medina, V.; Croci, M.; Crescenti, E.; Mohamad, N.; Sanchez-Jimenez, F.; Massari, N.; Nunez, M.; Cricco, G.; Martin, G.; Bergoc, R.; et al. The role of histamine in human mammary carcinogenesis: H3 and H4 receptors as potential therapeutic targets for breast cancer treatment. Cancer Biol. Ther. 2008, 7, 28–35. [Google Scholar] [CrossRef] [PubMed]
- Meng, F.; Han, Y.; Staloch, D.; Francis, T.; Stokes, A.; Francis, H. The H4 histamine receptor agonist, clobenpropit, suppresses human cholangiocarcinoma progression by disruption of epithelial mesenchymal transition and tumor metastasis. Hepatology 2011, 54, 1718–1728. [Google Scholar] [CrossRef] [PubMed]
- Hirose, H.; Kawabe, H.; Komiya, N.; Saito, I. Relations between serum reactive oxygen metabolites (ROMs) and various inflammatory and metabolic parameters in a Japanese population. J. Atheroscler. Thromb. 2009, 16, 77–82. [Google Scholar] [CrossRef] [PubMed]
- Komatsu, F.; Kagawa, Y.; Ishiguro, K.; Kawabata, T.; Purvee, B.; Otgon, J.; Chimedregzen, U. The association of very high hair manganese accumulation and high oxidative stress in Mongolian people. Curr. Aging Sci. 2009, 2, 28–42. [Google Scholar] [CrossRef] [PubMed]
- Cornelli, U.; Terranova, R.; Luca, S.; Cornelli, M.; Alberti, A. Bioavailability and antioxidant activity of some food supplements in men and women using the D-Roms test as a marker of oxidative stress. J. Nutr. 2001, 131, 3208–3211. [Google Scholar] [PubMed]
- Yasui, Y.; Hosokawa, M.; Mikami, N.; Miyashita, K.; Tanaka, T. Dietary astaxanthin inhibits colitis and colitis-associated colon carcinogenesis in mice via modulation of the inflammatory cytokines. Chem. Biol. Interact. 2011, 193, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Ten Eick, A.P.; Blumer, J.L.; Reed, M.D. Safety of antihistamines in children. Drug Saf. 2001, 24, 119–147. [Google Scholar] [CrossRef] [PubMed]
- Lefranc, F.; Yeaton, P.; Brotchi, J.; Kiss, R. Cimetidine, an unexpected anti-tumor agent, and its potential for the treatment of glioblastoma (review). Int. J. Oncol. 2006, 28, 1021–1030. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.D.; Wang, Y.J.; Chen, C.H.; Yu, C.F.; Chen, L.C.; Lin, J.K.; Liang, Y.C.; Lin, S.Y.; Ho, Y.S. Molecular mechanisms of G0/G1 cell-cycle arrest and apoptosis induced by terfenadine in human cancer cells. Mol. Carcinog. 2003, 37, 39–50. [Google Scholar] [CrossRef] [PubMed]
- Lawson, J.A.; Adams, W.J.; Morris, D.L. Ranitidine and cimetidine differ in their in vitro and in vivo effects on human colonic cancer growth. Br. J. Cancer 1996, 73, 872–876. [Google Scholar] [CrossRef] [PubMed]
- Caignard, A.; Martin, M.; Reisser, D.; Thomas, B.; Martin, F. Effects of cimetidine and indomethacin on the growth of dimethylhydrazine-induced or transplanted intestinal cancers in the rat. Br. J. Cancer 1984, 50, 661–665. [Google Scholar] [CrossRef] [PubMed]
- Paik, W.H.; Ryu, J.K.; Jeong, K.S.; Park, J.M.; Song, B.J.; Lee, S.H.; Kim, Y.T.; Yoon, Y.B. Clobenpropit enhances anti-tumor effect of gemcitabine in pancreatic cancer. World J. Gastroenterol. 2014, 20, 8545–8557. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, T.; de Azevedo, M.B.; Duran, N.; Alderete, J.B.; Epifano, F.; Genovese, S.; Tanaka, M.; Curini, M. Colorectal cancer chemoprevention by 2 beta-cyclodextrin inclusion compounds of auraptene and 4'-geranyloxyferulic acid. Int. J. Cancer 2010, 126, 830–840. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group No. | Treatment | No. of Mice Examined | Mucosal Ulcer | High-Grade Dysplasia | Adenoma (AD) | Adeno-Carcinoma (ADC) | Total Tumor (AD+ADC) |
|---|---|---|---|---|---|---|---|
| 1 | AOM + DSS | 20 | 10/20 (50%) | 15/20 (75%) | 11/20 (55%) | 11/20 (55%) | 13/20 (65%) |
| 1.87 ± 1.81 a | 3.93 ± 2.43 | 2.40 ± 2.35 | 2.73 ± 3.45 | 5.13 ± 4.93 | |||
| 2 | AOM + DSS + 500 ppm terfenadine | 20 | 10/20 (50%) | 13/20 (65%) | 10/20 (50%) | 13/20 (65%) | 14/20 (70%) |
| 1.14 ± 0.95 | 3.29 ± 2.37 | 1.50 ± 1.70 | 2.50 ± 1.34 | 4.00 ± 2.11 | |||
| 3 | AOM + DSS + 500 ppm cimetidine | 20 | 6/20 (30%) | 11/20 (55%) | 6/20 (30%) | 5/20 (25%) | 8/20 (40%) |
| 0.53 ± 0.74 b | 1.13 ± 1.13 c | 0.53 ± 0.74 b | 0.60 ± 1.12 b | 1.13 ± 1.73 c | |||
| 4 | AOM + DSS + 10 ppm clobenpropit | 20 | 8/20 (40%) | 13/20 (65%) | 7/20 (35%) | 6/20 (30%) | 8/20 (40%) |
| 0.60 ± 0.63 b | 1.67 ± 1.29 b | 0.73 ± 0.96 b | 0.60 ± 0.91 b | 1.33 ± 1.72 c | |||
| 5 | 500 ppm terfenadine | 8 | 0/8 (0%) | 0/8 (0%) | 0/8 (0%) | 0/8 (0%) | 0/8 (0%) |
| 6 | 500 ppm cimetidine | 8 | 0/8 (0%) | 0/8 (0%) | 0/8 (0%) | 0/8 (0%) | 0/8 (0%) |
| 7 | 10 ppm clobenpropit | 8 | 0/8 (0%) | 0/8 (0%) | 0/8 (0%) | 0/8 (0%) | 0/8 (0%) |
| 8 | None | 8 | 0/8 (0%) | 0/8 (0%) | 0/8 (0%) | 0/8 (0%) | 0/8 (0%) |
| Group No. | Treatment | No. of Mice Examined | d-ROM a (CARR U b) | BAP a (mM/L) | d-ROM/BAP Ratio |
|---|---|---|---|---|---|
| 1 | AOM + DSS | 20 | 126 ± 11 c | 2931 ± 302 | 0.04361 ± 0.00823 |
| 2 | AOM + DSS + 500 ppm terfenadine | 20 | 133 ± 11 | 2958 ± 470 | 0.04795 ± 0.00591 |
| 3 | AOM + DSS + 500 ppm cimetidine | 20 | 123 ± 26 | 1630 ± 987 | 0.09594 ± 0.05643 |
| 4 | AOM + DSS + 10 ppm clobenpropit | 20 | 131 ± 15 | 2328 ± 1088 | 0.07353 ± 0.02484 |
| 5 | 500 ppm terfenadine | 8 | 138 ± 9 | 2658 ± 909 | 0.04968± 0.01197 |
| 6 | 500 ppm cimetidine | 8 | 130 ± 8 | 2753 ± 1802 | 0.09091 ± 0.08894 |
| 7 | 10 ppm clobenpropit | 8 | 147 ± 10 | 1405 ± 801 | 0.15870 ± 0.06784 |
| 8 | None | 8 | 142 ± 18 | 2477 ± 954 | 0.08182 ± 0.05208 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tanaka, T.; Kochi, T.; Shirakami, Y.; Mori, T.; Kurata, A.; Watanabe, N.; Moriwaki, H.; Shimizu, M. Cimetidine and Clobenpropit Attenuate Inflammation-Associated Colorectal Carcinogenesis in Male ICR Mice. Cancers 2016, 8, 25. https://doi.org/10.3390/cancers8020025
Tanaka T, Kochi T, Shirakami Y, Mori T, Kurata A, Watanabe N, Moriwaki H, Shimizu M. Cimetidine and Clobenpropit Attenuate Inflammation-Associated Colorectal Carcinogenesis in Male ICR Mice. Cancers. 2016; 8(2):25. https://doi.org/10.3390/cancers8020025
Chicago/Turabian StyleTanaka, Takuji, Takahiro Kochi, Yohei Shirakami, Takayuki Mori, Ayumi Kurata, Naoki Watanabe, Hisataka Moriwaki, and Masahito Shimizu. 2016. "Cimetidine and Clobenpropit Attenuate Inflammation-Associated Colorectal Carcinogenesis in Male ICR Mice" Cancers 8, no. 2: 25. https://doi.org/10.3390/cancers8020025
APA StyleTanaka, T., Kochi, T., Shirakami, Y., Mori, T., Kurata, A., Watanabe, N., Moriwaki, H., & Shimizu, M. (2016). Cimetidine and Clobenpropit Attenuate Inflammation-Associated Colorectal Carcinogenesis in Male ICR Mice. Cancers, 8(2), 25. https://doi.org/10.3390/cancers8020025