NG2 Proteoglycan Enhances Brain Tumor Progression by Promoting Beta-1 Integrin Activation in both Cis and Trans Orientations

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Cis Interaction of NG2 with β1 Integrins
2.1. NG2 in Glioma Cells
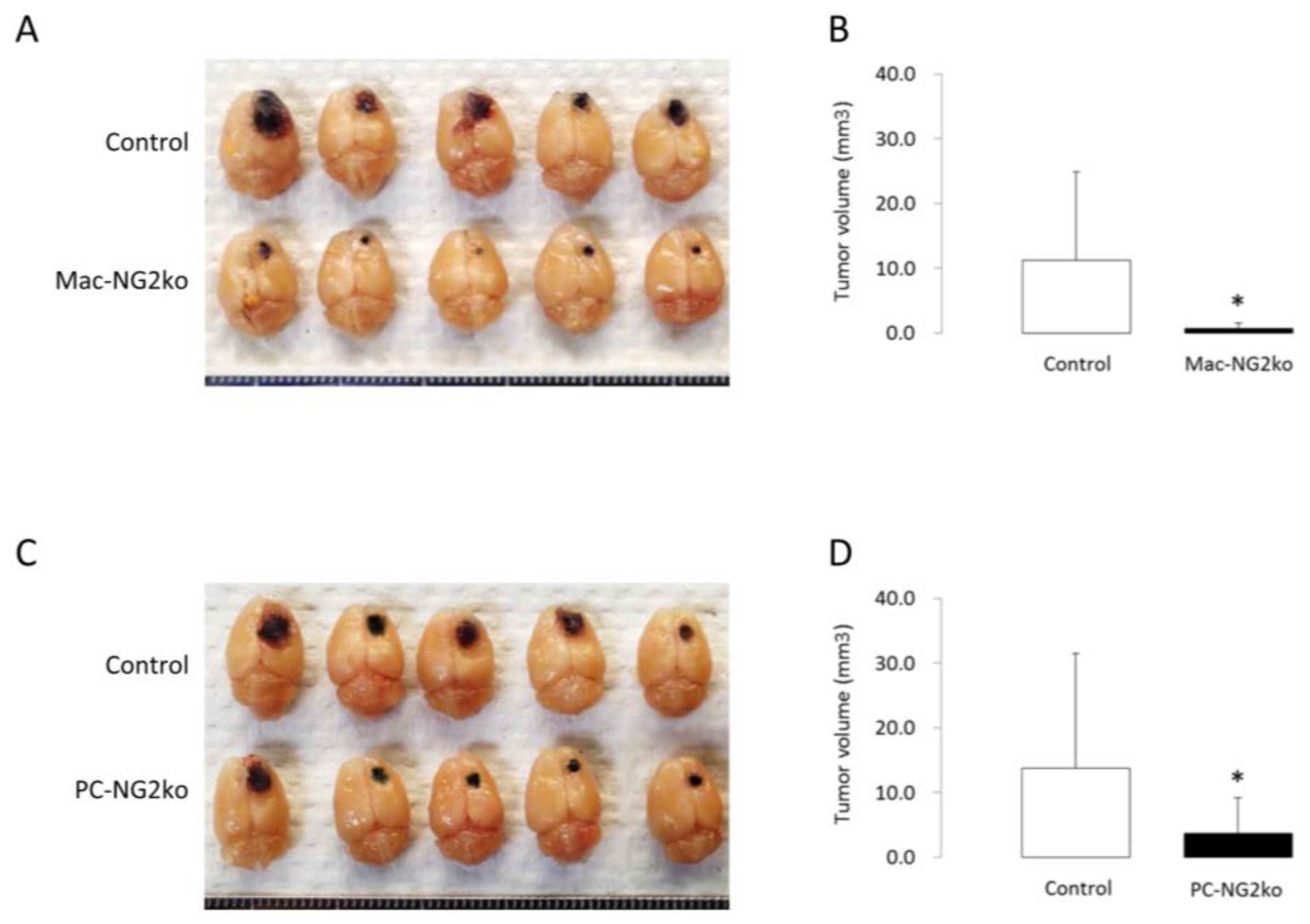
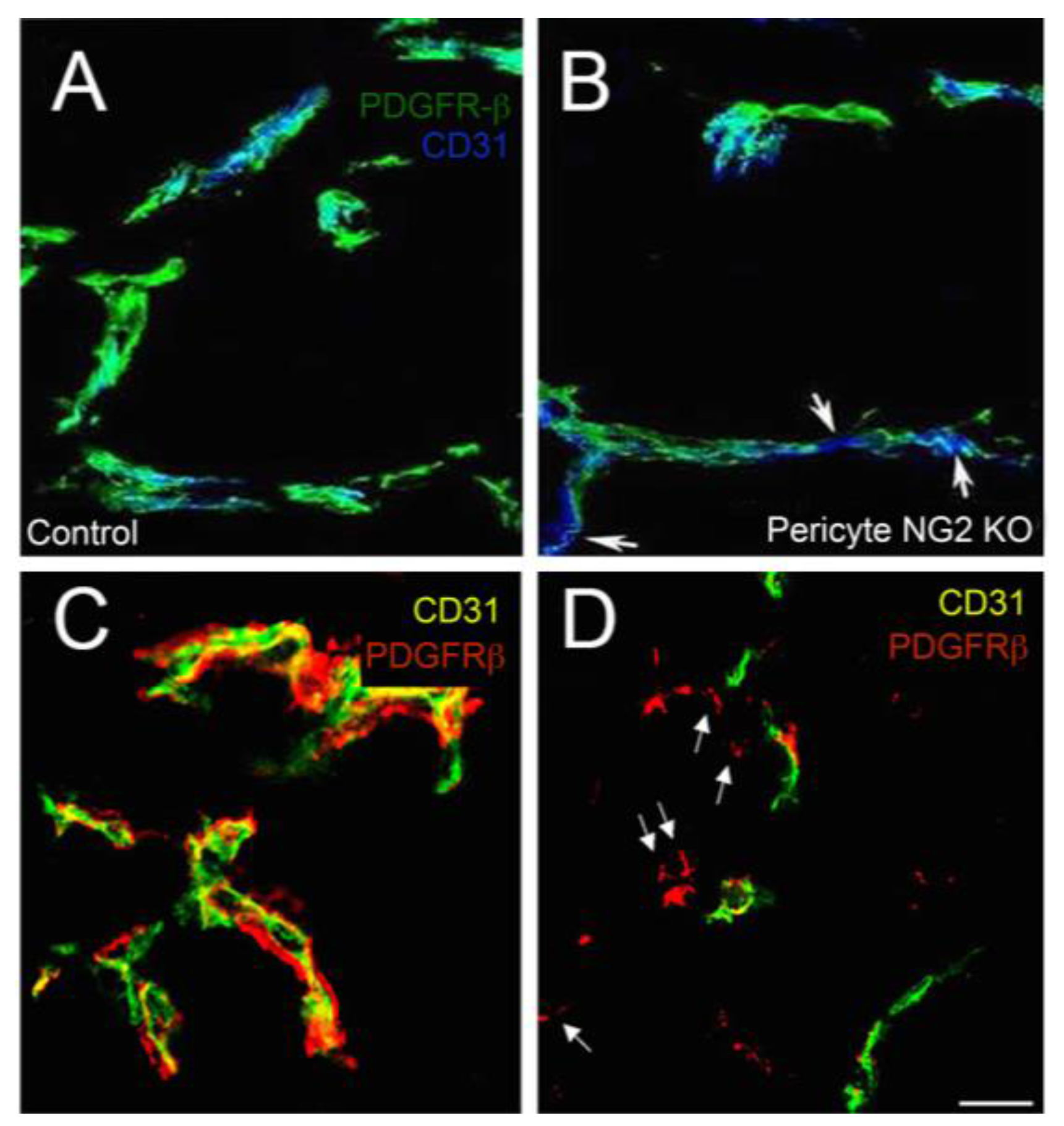
2.2. NG2 in Pericytes
2.3. NG2 in Macrophages
3. Trans Interactions of NG2 with β1 Integrins
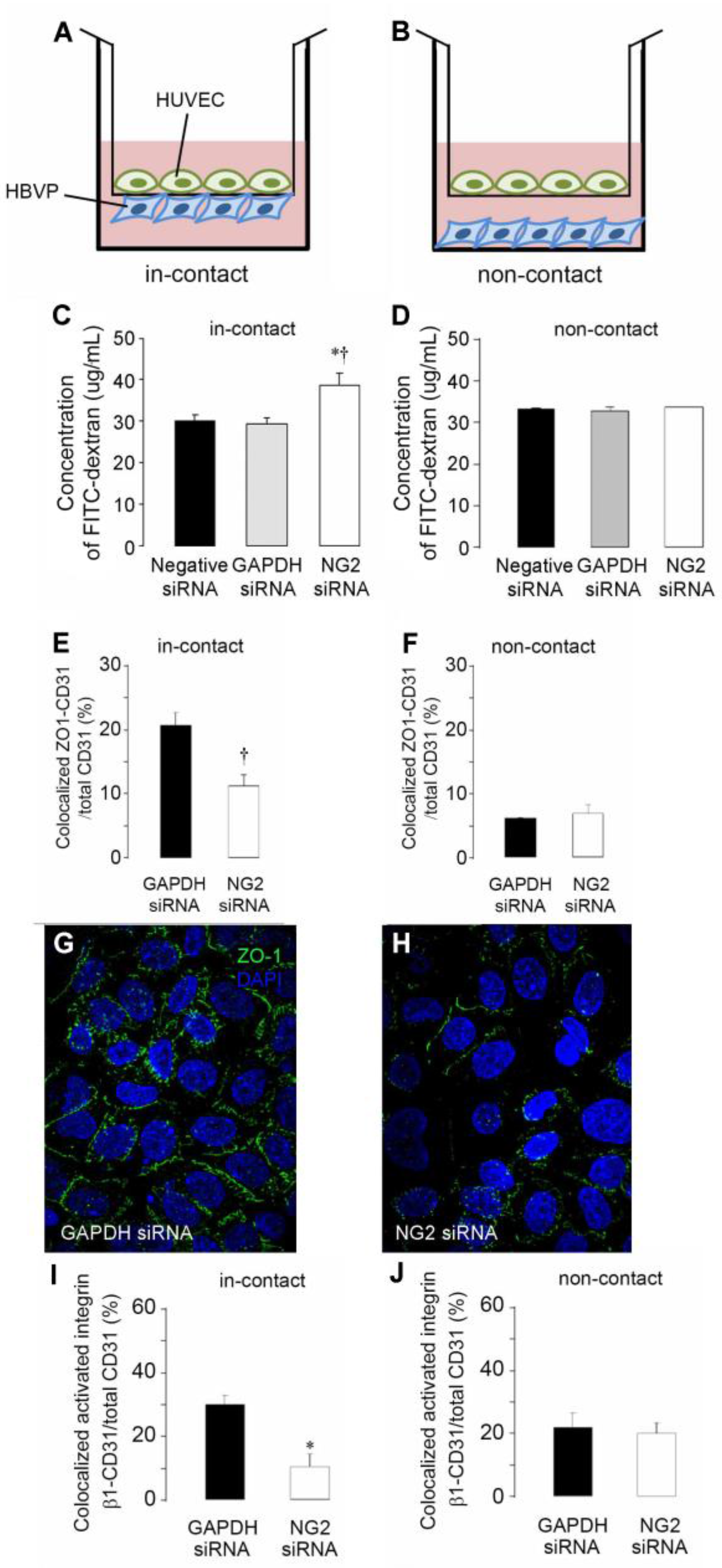
3.1. Effects of Soluble NG2 on Endothelial Cells
3.2. Effects of Pericyte NG2 on Endothelial Cells
4. Future Prospects
- What domains of NG2 and the integrin heterodimer are responsible for their interaction? Our results indicate that the NG2-integrin interaction is mediated by the extracellular domains of the two proteins, rather than by the cytoplasmic domains. However, we do not know what part of the NG2 ectodomain is involved in the interaction, or whether NG2 binds to the α or β integrin subunit (or to both). This also leaves open the questions of whether NG2 preferentially interacts with specific β1 heterodimers and how the spatial requirements for the interaction can be satisfied in both the cis and trans orientations.
- Does NG2 influence crosstalk between integrins and receptor tyrosine kinases? Crosstalk between integrins and receptor tyrosine kinases is proposed to be an important aspect of amplifying and/or modulating transmembrane signaling [74,75,76,77]. Since NG2 has the ability to activate both of these signaling entities, it is of interest to know how the proteoglycan may affect crosstalk that occurs between the two and what implications this may have for the properties (e.g., proliferation, motility, survival) of both tumor and stromal cells.
- How does NG2 interact with the cytoskeleton and how is this interaction affected by phosphorylation of the NG2 cytoplasmic domain? We propose that the phosphorylation state of NG2 determines its interaction with cytoplasmic scaffolding proteins and thus the localization of both NG2 and its β1 binding partner to specific membrane microdomains (lamellipodia versus apical microprotrusions). It remains for us to identify these cytoplasmic scaffolding proteins and to demonstrate how they anchor the NG2-integrin complex at sites that specifically influence proliferation versus motility.
- How does NG2-dependent cis activation of integrins influence the biology of macrophages? Although we can demonstrate enhanced activation of both β1 and β2 integrins in NG2-positive THP1 macrophages, we have not yet established whether this influences macrophage interaction with the vascular endothelium (e.g. via binding to VCAM1 or ICAM1) or the ability of macrophages to extravasate from the circulation into tumors and other sites of inflammation. This will be an important addition to the spectrum of cellular events that are impacted by NG2.
Acknowledgments
Conflicts of Interest
References
- Fang, X.; Burg, M.A.; Barritt, D.; Dahlin-Huppe, K.; Nishiyama, A.; Stallcup, W.B. Cytoskeletal reorganization induced by engagement of the NG2 proteoglycan leads to cell spreading and migration. Mol. Biol. Cell 1999, 10, 3373–3387. [Google Scholar] [CrossRef] [PubMed]
- Majumdar, M.; Vuori, K.; Stallcup, W.B. Engagement of the NG2 proteoglycan triggers cell spreading via rac and p130cas. Cell. Signal. 2003, 15, 79–84. [Google Scholar] [CrossRef]
- Chekenya, M.; Krakstad, C.; Svendsen, A.; Netland, I.A.; Staalesen, V.; Tysnes, B.B.; Selheim, F.; Wang, J.; Sakariassen, P.O.; Sandal, T.; et al. The progenitor cell marker NG2/MPG promotes chemoresistance by activation of integrin-dependent PI3K/Akt signaling. Oncogene 2008, 27, 5182–5194. [Google Scholar] [CrossRef] [PubMed]
- Fukushi, J.; Makagiansar, I.T.; Stallcup, W.B. NG2 proteoglycan promotes endothelial cell motility and angiogenesis via engagement of galectin-3 and alpha3beta1 integrin. Mol. Biol. Cell 2004, 15, 3580–3590. [Google Scholar] [CrossRef] [PubMed]
- Iida, J.; Meijne, A.M.; Spiro, R.C.; Roos, E.; Furcht, L.T.; McCarthy, J.B. Spreading and focal contact formation of human melanoma cells in response to the stimulation of both melanoma-associated proteoglycan (NG2) and alpha 4 beta 1 integrin. Cancer Res. 1995, 55, 2177–2185. [Google Scholar] [PubMed]
- Knutson, J.R.; Iida, J.; Fields, G.B.; McCarthy, J.B. CD44/chondroitin sulfate proteoglycan and alpha 2 beta 1 integrin mediate human melanoma cell migration on type IV collagen and invasion of basement membranes. Mol. Biol. Cell 1996, 7, 383–396. [Google Scholar] [CrossRef] [PubMed]
- Makagiansar, I.T.; Williams, S.; Dahlin-Huppe, K.; Fukushi, J.; Mustelin, T.; Stallcup, W.B. Phosphorylation of NG2 proteoglycan by protein kinase C-alpha regulates polarized membrane distribution and cell motility. J. Biol. Chem. 2004, 279, 55262–55270. [Google Scholar] [CrossRef] [PubMed]
- Makagiansar, I.T.; Williams, S.; Mustelin, T.; Stallcup, W.B. Differential phosphorylation of NG2 proteoglycan by ERK and PKCalpha helps balance cell proliferation and migration. J. Cell Biol. 2007, 178, 155–165. [Google Scholar] [CrossRef] [PubMed]
- You, W.K.; Yotsumoto, F.; Sakimura, K.; Adams, R.H.; Stallcup, W.B. NG2 proteoglycan promotes tumor vascularization via integrin-dependent effects on pericyte function. Angiogenesis 2014, 17, 61–76. [Google Scholar] [CrossRef] [PubMed]
- Cattaruzza, S.; Ozerdem, U.; Denzel, M.; Ranscht, B.; Bulian, P.; Cavallaro, U.; Zanocco, D.; Colombatti, A.; Stallcup, W.B.; Perris, R. Multivalent proteoglycan modulation of FGF mitogenic responses in perivascular cells. Angiogenesis 2013, 16, 309–327. [Google Scholar] [CrossRef] [PubMed]
- Goretzki, L.; Burg, M.A.; Grako, K.A.; Stallcup, W.B. High-affinity binding of basic fibroblast growth factor and platelet-derived growth factor-AA to the core protein of the NG2 proteoglycan. J. Biol. Chem. 1999, 274, 16831–16837. [Google Scholar] [CrossRef] [PubMed]
- Grako, K.A.; Ochiya, T.; Barritt, D.; Nishiyama, A.; Stallcup, W.B. PDGF (alpha)-receptor is unresponsive to PDGF-AA in aortic smooth muscle cells from the NG2 knockout mouse. J. Cell Sci. 1999, 112, 905–915. [Google Scholar] [PubMed]
- Grako, K.A.; Stallcup, W.B. Participation of the NG2 proteoglycan in rat aortic smooth muscle cell responses to platelet-derived growth factor. Exp. Cell Res. 1995, 221, 231–240. [Google Scholar] [CrossRef] [PubMed]
- Sugiarto, S.; Persson, A.I.; Munoz, E.G.; Waldhuber, M.; Lamagna, C.; Andor, N.; Hanecker, P.; Ayers-Ringler, J.; Phillips, J.; Siu, J.; et al. Asymmetry-defective oligodendrocyte progenitors are glioma precursors. Cancer Cell 2011, 20, 328–340. [Google Scholar] [CrossRef] [PubMed]
- Nishiyama, A.; Lin, X.H.; Giese, N.; Heldin, C.H.; Stallcup, W.B. Interaction between NG2 proteoglycan and PDGF alpha-receptor on O2A progenitor cells is required for optimal response to PDGF. J. Neurosci. Res. 1996, 43, 315–330. [Google Scholar] [CrossRef]
- Stallcup, W.B. The NG2 proteoglycan: past insights and future prospects. J. Neurocytol. 2002, 31, 423–435. [Google Scholar] [CrossRef] [PubMed]
- Stallcup, W.B.; Huang, F.J. A role for the NG2 proteoglycan in glioma progression. Cell Adh. Migr. 2008, 2, 192–201. [Google Scholar] [CrossRef] [PubMed]
- Cattaruzza, S.; Nicolosi, P.A.; Braghetta, P.; Pazzaglia, L.; Benassi, M.S.; Picci, P.; Lacrima, K.; Zanocco, D.; Rizzo, E.; Stallcup, W.B.; et al. NG2/CSPG4-collagen type VI interplays putatively involved in the microenvironmental control of tumour engraftment and local expansion. J. Mol. Cell Biol. 2013, 5, 176–193. [Google Scholar] [CrossRef] [PubMed]
- Smith, F.O.; Rauch, C.; Williams, D.E.; March, C.J.; Arthur, D.; Hilden, J.; Lampkin, B.C.; Buckley, J.D.; Buckley, C.V.; Woods, W.G.; et al. The human homologue of rat NG2, a chondroitin sulfate proteoglycan, is not expressed on the cell surface of normal hematopoietic cells but is expressed by acute myeloid leukemia blasts from poor-prognosis patients with abnormalities of chromosome band 11q23. Blood 1996, 87, 1123–1133. [Google Scholar] [PubMed]
- Behm, F.G.; Smith, F.O.; Raimondi, S.C.; Pui, C.H.; Bernstein, I.D. Human homologue of the rat chondroitin sulfate proteoglycan, NG2, detected by monoclonal antibody 7.1, identifies childhood acute lymphoblastic leukemias with t(4;11)(q21;q23) or t(11;19)(q23;p13) and MLL gene rearrangements. Blood 1996, 87, 1134–1139. [Google Scholar] [PubMed]
- Shoshan, Y.; Nishiyama, A.; Chang, A.; Mork, S.; Barnett, G.H.; Cowell, J.K.; Trapp, B.D.; Staugaitis, S.M. Expression of oligodendrocyte progenitor cell antigens by gliomas: implications for the histogenesis of brain tumors. Proc. Natl. Acad. Sci. USA 1999, 96, 10361–10366. [Google Scholar] [CrossRef] [PubMed]
- Al-Mayhani, M.T.; Grenfell, R.; Narita, M.; Piccirillo, S.; Kenney-Herbert, E.; Fawcett, J.W.; Collins, V.P.; Ichimura, K.; Watts, C. NG2 expression in glioblastoma identifies an actively proliferating population with an aggressive molecular signature. NeuroOncology 2011, 13, 830–845. [Google Scholar] [CrossRef] [PubMed]
- Persson, A.I.; Petritsch, C.; Swartling, F.J.; Itsara, M.; Sim, F.J.; Auvergne, R.; Goldenberg, D.D.; Vandenberg, S.R.; Nguyen, K.N.; Yakovenko, S.; et al. Non-stem cell origin for oligodendroglioma. Cancer Cell 2010, 18, 669–682. [Google Scholar] [CrossRef] [PubMed]
- Price, M.A.; Colvin Wanshura, L.E.; Yang, J.; Carlson, J.; Xiang, B.; Li, G.; Ferrone, S.; Dudek, A.Z.; Turley, E.A.; McCarthy, J.B. CSPG4, a potential therapeutic target, facilitates malignant progression of melanoma. Pigment Cell Melanoma. Res. 2011, 24, 1148–1157. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Svendsen, A.; Kmiecik, J.; Immervoll, H.; Skaftnesmo, K.O.; Planaguma, J.; Reed, R.K.; Bjerkvig, R.; Miletic, H.; Enger, P.O.; et al. Targeting the NG2/CSPG4 proteoglycan retards tumour growth and angiogenesis in preclinical models of GBM and melanoma. PLoS ONE 2011, 6, e23062. [Google Scholar] [CrossRef] [PubMed]
- Yadavilli, S.; Hwang, E.I.; Packer, R.J.; Nazarian, J. The Role of NG2 Proteoglycan in Glioma. Transl. Oncol. 2016, 9, 57–63. [Google Scholar] [CrossRef] [PubMed]
- Burg, M.A.; Grako, K.A.; Stallcup, W.B. Expression of the NG2 proteoglycan enhances the growth and metastatic properties of melanoma cells. J. Cell Physiol. 1998, 177, 299–312. [Google Scholar] [CrossRef]
- Stallcup, W.B.; You, W.K.; Kucharova, K.; Cejudo-Martin, P.; Yotsumoto, F. NG2 Proteoglycan-Dependent Contributions of Pericytes and Macrophages to Brain Tumor Vascularization and Progression. Microcirculation 2016, 23, 122–133. [Google Scholar] [CrossRef] [PubMed]
- Clausen, T.M.; Pereira, M.A.; Al Nakouzi, N.; Oo, H.Z.; Agerbaek, M.O.; Lee, S.; Orum-Madsen, M.S.; Kristensen, A.R.; El-Naggar, A.; Grandgenett, P.M.; et al. Oncofetal Chondroitin Sulfate Glycosaminoglycans Are Key Players in Integrin Signaling and Tumor Cell Motility. Mol. Cancer Res. 2016, 14, 1288–1299. [Google Scholar] [CrossRef] [PubMed]
- Viola, M.; Bruggemann, K.; Karousou, E.; Caon, I.; Carava, E.; Vigetti, D.; Greve, B.; Stock, C.; De Luca, G.; Passi, A.; et al. MDA-MB-231 breast cancer cell viability, motility and matrix adhesion are regulated by a complex interplay of heparan sulfate, chondroitin-/dermatan sulfate and hyaluronan biosynthesis. Glycoconj. J. 2016. [Google Scholar] [CrossRef] [PubMed]
- Garrigues, H.J.; Lark, M.W.; Lara, S.; Hellstrom, I.; Hellstrom, K.E.; Wight, T.N. The melanoma proteoglycan: restricted expression on microspikes, a specific microdomain of the cell surface. J. Cell Biol. 1986, 103, 1699–1710. [Google Scholar] [CrossRef] [PubMed]
- Barritt, D.S.; Pearn, M.T.; Zisch, A.H.; Lee, S.S.; Javier, R.T.; Pasquale, E.B.; Stallcup, W.B. The multi-PDZ domain protein MUPP1 is a cytoplasmic ligand for the membrane-spanning proteoglycan NG2. J. Cell Biochem. 2000, 79, 213–224. [Google Scholar] [CrossRef]
- Stegmuller, J.; Werner, H.; Nave, K.A.; Trotter, J. The proteoglycan NG2 is complexed with alpha-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid (AMPA) receptors by the PDZ glutamate receptor interaction protein (GRIP) in glial progenitor cells. Implications for glial-neuronal signaling. J. Biol. Chem. 2003, 278, 3590–3598. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, N.; Stegmuller, J.; Schatzle, P.; Karram, K.; Koroll, M.; Werner, H.B.; Nave, K.A.; Trotter, J. Interaction of syntenin-1 and the NG2 proteoglycan in migratory oligodendrocyte precursor cells. J. Biol. Chem. 2008, 283, 8310–8317. [Google Scholar] [CrossRef] [PubMed]
- Luque, A.; Gomez, M.; Puzon, W.; Takada, Y.; Sanchez-Madrid, F.; Cabanas, C. Activated conformations of very late activation integrins detected by a group of antibodies (HUTS) specific for a novel regulatory region (355-425) of the common beta 1 chain. J. Biol. Chem. 1996, 271, 11067–11075. [Google Scholar] [PubMed]
- Lathia, J.D.; Gallagher, J.; Heddleston, J.M.; Wang, J.; Eyler, C.E.; Macswords, J.; Wu, Q.; Vasanji, A.; McLendon, R.E.; Hjelmeland, A.B.; et al. Integrin alpha 6 regulates glioblastoma stem cells. Cell Stem Cell 2010, 6, 421–432. [Google Scholar] [CrossRef] [PubMed]
- Hanahan, D.; Folkman, J. Patterns and emerging mechanisms of the angiogenic switch during tumorigenesis. Cell 1996, 86, 353–364. [Google Scholar] [CrossRef]
- Bergers, G.; Benjamin, L.E. Tumorigenesis and the angiogenic switch. Nat. Rev. Cancer 2003, 3, 410. [Google Scholar] [CrossRef] [PubMed]
- Armulik, A.; Abramsson, A.; Betsholtz, C. Endothelial/pericyte interactions. Circ. Res. 2005, 97, 512–523. [Google Scholar] [CrossRef] [PubMed]
- Bergers, G.; Song, S. The role of pericytes in blood-vessel formation and maintenance. NeuroOncology 2005, 7, 452–464. [Google Scholar] [CrossRef] [PubMed]
- Ozerdem, U.; Grako, K.A.; Dahlin-Huppe, K.; Monosov, E.; Stallcup, W.B. NG2 proteoglycan is expressed exclusively by mural cells during vascular morphogenesis. Dev. Dyn. 2001, 222, 218–227. [Google Scholar] [CrossRef] [PubMed]
- Ozerdem, U.; Monosov, E.; Stallcup, W.B. NG2 proteoglycan expression by pericytes in pathological microvasculature. Microvasc. Res. 2002, 63, 129–134. [Google Scholar] [CrossRef] [PubMed]
- Ozerdem, U.; Stallcup, W.B. Early contribution of pericytes to angiogenic sprouting and tube formation. Angiogenesis 2003, 6, 241–249. [Google Scholar] [CrossRef] [PubMed]
- Ozerdem, U.; Stallcup, W.B. Pathological angiogenesis is reduced by targeting pericytes via the NG2 proteoglycan. Angiogenesis 2004, 7, 269–276. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.; She, Z.G.; Sakimura, K.; Roberts, A.; Kucharova, K.; Rowitch, D.H.; Stallcup, W.B. Ablation of NG2 proteoglycan leads to deficits in brown fat function and to adult onset obesity. PLoS ONE 2012, 7, e30637. [Google Scholar] [CrossRef] [PubMed]
- Clausen, B.E.; Burkhardt, C.; Reith, W.; Renkawitz, R.; Forster, I. Conditional gene targeting in macrophages and granulocytes using LysMcre mice. Transgenic. Res. 1999, 8, 265–277. [Google Scholar] [CrossRef] [PubMed]
- Stockmann, C.; Doedens, A.; Weidemann, A.; Zhang, N.; Takeda, N.; Greenberg, J.I.; Cheresh, D.A.; Johnson, R.S. Deletion of vascular endothelial growth factor in myeloid cells accelerates tumorigenesis. Nature 2008, 456, 814–818. [Google Scholar] [CrossRef] [PubMed]
- Yotsumoto, F.; You, W.K.; Cejudo-Martin, P.; Kucharova, K.; Sakimura, K.; Stallcup, W.B. NG2 proteoglycan-dependent recruitment of tumor macrophages promotes pericyte-endothelial cell interactions required for brain tumor vascularization. Oncoimmunology 2015, 4, e1001204. [Google Scholar] [CrossRef] [PubMed]
- Coffelt, S.B.; Lewis, C.E.; Naldini, L.; Brown, J.M.; Ferrara, N.; De Palma, M. Elusive identities and overlapping phenotypes of proangiogenic myeloid cells in tumors. Am. J. Pathol. 2010, 176, 1564–1576. [Google Scholar] [CrossRef] [PubMed]
- De Palma, M.; Venneri, M.A.; Galli, R.; Sergi Sergi, L.; Politi, L.S.; Sampaolesi, M.; Naldini, L. Tie2 identifies a hematopoietic lineage of proangiogenic monocytes required for tumor vessel formation and a mesenchymal population of pericyte progenitors. Cancer Cell 2005, 8, 211–226. [Google Scholar] [CrossRef] [PubMed]
- Dransfield, I.; Cabanas, C.; Barrett, J.; Hogg, N. Interaction of leukocyte integrins with ligand is necessary but not sufficient for function. J. Cell Biol. 1992, 116, 1527–1535. [Google Scholar] [CrossRef] [PubMed]
- Forlow, S.B.; White, E.J.; Barlow, S.C.; Feldman, S.H.; Lu, H.; Bagby, G.J.; Beaudet, A.L.; Bullard, D.C.; Ley, K. Severe inflammatory defect and reduced viability in CD18 and E-selectin double-mutant mice. J. Clin. Investig. 2000, 106, 1457–1466. [Google Scholar] [CrossRef] [PubMed]
- Jin, H.; Su, J.; Garmy-Susini, B.; Kleeman, J.; Varner, J. Integrin alpha4beta1 promotes monocyte trafficking and angiogenesis in tumors. Cancer Res. 2006, 66, 2146–2152. [Google Scholar] [CrossRef] [PubMed]
- Stark, K.; Eckart, A.; Haidari, S.; Tirniceriu, A.; Lorenz, M.; von Bruhl, M.L.; Gartner, F.; Khandoga, A.G.; Legate, K.R.; Pless, R.; et al. Capillary and arteriolar pericytes attract innate leukocytes exiting through venules and ‘instruct’ them with pattern-recognition and motility programs. Nat. Immunol. 2013, 14, 41–51. [Google Scholar] [CrossRef] [PubMed]
- Ulyanova, T.; Priestley, G.V.; Banerjee, E.R.; Papayannopoulou, T. Unique and redundant roles of alpha4 and beta2 integrins in kinetics of recruitment of lymphoid vs. myeloid cell subsets to the inflamed peritoneum revealed by studies of genetically deficient mice. Exp. Hematol. 2007, 35, 1256–1265. [Google Scholar] [CrossRef] [PubMed]
- Cohen, J.V.; Tawbi, H.; Margolin, K.A.; Amravadi, R.; Bosenberg, M.; Brastianos, P.K.; Chiang, V.L.; de Groot, J.; Glitza, I.C.; Herlyn, M.; et al. Melanoma central nervous system metastases: current approaches, challenges, and opportunities. Pigment Cell Melanoma. Res. 2016, 29, 627–642. [Google Scholar] [CrossRef] [PubMed]
- Bunse, S.; Garg, S.; Junek, S.; Vogel, D.; Ansari, N.; Stelzer, E.H.; Schuman, E. Role of N-cadherin cis and trans interfaces in the dynamics of adherens junctions in living cells. PLoS ONE 2013, 8, e81517. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Jin, X.; Harrison, O.; Shapiro, L.; Honig, B.H.; Ben-Shaul, A. Cooperativity between trans and cis interactions in cadherin-mediated junction formation. Proc. Natl. Acad. Sci. USA 2010, 107, 17592–17597. [Google Scholar] [CrossRef] [PubMed]
- Arvanitis, D.; Davy, A. Eph/ephrin signaling: networks. Genes Dev. 2008, 22, 416–429. [Google Scholar] [CrossRef] [PubMed]
- Nishiyama, A.; Lin, X.H.; Stallcup, W.B. Generation of truncated forms of the NG2 proteoglycan by cell surface proteolysis. Mol. Biol. Cell 1995, 6, 1819–1832. [Google Scholar] [CrossRef] [PubMed]
- Martin, S.; Levine, A.K.; Chen, Z.J.; Ughrin, Y.; Levine, J.M. Deposition of the NG2 proteoglycan at nodes of Ranvier in the peripheral nervous system. J. Neurosci. 2001, 21, 8119–8128. [Google Scholar] [PubMed]
- Jones, L.L.; Yamaguchi, Y.; Stallcup, W.B.; Tuszynski, M.H. NG2 is a major chondroitin sulfate proteoglycan produced after spinal cord injury and is expressed by macrophages and oligodendrocyte progenitors. J. Neurosci. 2002, 22, 2792–2803. [Google Scholar] [PubMed]
- Larsen, P.H.; Wells, J.E.; Stallcup, W.B.; Opdenakker, G.; Yong, V.W. Matrix metalloproteinase-9 facilitates remyelination in part by processing the inhibitory NG2 proteoglycan. J. Neurosci. 2003, 23, 11127–11135. [Google Scholar] [PubMed]
- Asher, R.A.; Morgenstern, D.A.; Properzi, F.; Nishiyama, A.; Levine, J.M.; Fawcett, J.W. Two separate metalloproteinase activities are responsible for the shedding and processing of the NG2 proteoglycan in vitro. Mol. Cell Neurosci. 2005, 29, 82–96. [Google Scholar] [CrossRef] [PubMed]
- Sakry, D.; Neitz, A.; Singh, J.; Frischknecht, R.; Marongiu, D.; Biname, F.; Perera, S.S.; Endres, K.; Lutz, B.; Radyushkin, K.; et al. Oligodendrocyte precursor cells modulate the neuronal network by activity-dependent ectodomain cleavage of glial NG2. PLoS Biol. 2014, 12, e1001993. [Google Scholar] [CrossRef] [PubMed]
- Joo, N.E.; Miao, D.; Bermudez, M.; Stallcup, W.B.; Kapila, Y.L. Shedding of NG2 by MMP-13 attenuates anoikis. DNA Cell Biol. 2014, 33, 854–862. [Google Scholar] [CrossRef] [PubMed]
- Tillet, E.; Ruggiero, F.; Nishiyama, A.; Stallcup, W.B. The membrane-spanning proteoglycan NG2 binds to collagens V and VI through the central nonglobular domain of its core protein. J. Biol. Chem. 1997, 272, 10769–10776. [Google Scholar] [PubMed]
- Lenter, M.; Uhlig, H.; Hamann, A.; Jeno, P.; Imhof, B.; Vestweber, D. A monoclonal antibody against an activation epitope on mouse integrin chain beta 1 blocks adhesion of lymphocytes to the endothelial integrin alpha 6 beta 1. Proc. Natl. Acad. Sci. USA 1993, 90, 9051–9055. [Google Scholar] [CrossRef] [PubMed]
- Kaya, M.; Ahishali, B. Assessment of permeability in barrier type of endothelium in brain using tracers: Evans blue, sodium fluorescein, and horseradish peroxidase. Methods Mol. Biol. 2011, 763, 369–382. [Google Scholar] [CrossRef] [PubMed]
- Foo, S.S.; Turner, C.J.; Adams, S.; Compagni, A.; Aubyn, D.; Kogata, N.; Lindblom, P.; Shani, M.; Zicha, D.; Adams, R.H. Ephrin-B2 controls cell motility and adhesion during blood-vessel-wall assembly. Cell 2006, 124, 161–173. [Google Scholar] [CrossRef] [PubMed]
- Stenzel, D.; Nye, E.; Nisancioglu, M.; Adams, R.H.; Yamaguchi, Y.; Gerhardt, H. Peripheral mural cell recruitment requires cell-autonomous heparan sulfate. Blood 2009, 114, 915–924. [Google Scholar] [CrossRef] [PubMed]
- Gibby, K.; You, W.K.; Kadoya, K.; Helgadottir, H.; Young, L.J.; Ellies, L.G.; Chang, Y.; Cardiff, R.D.; Stallcup, W.B. Early vascular deficits are correlated with delayed mammary tumorigenesis in the MMTV-PyMT transgenic mouse following genetic ablation of the NG2 proteoglycan. Breast Cancer Res. 2012, 14, R67. [Google Scholar] [CrossRef] [PubMed]
- Huang, F.J.; You, W.K.; Bonaldo, P.; Seyfried, T.N.; Pasquale, E.B.; Stallcup, W.B. Pericyte deficiencies lead to aberrant tumor vascularizaton in the brain of the NG2 null mouse. Dev. Biol. 2010, 344, 1035–1046. [Google Scholar] [CrossRef] [PubMed]
- Borges, E.; Jan, Y.; Ruoslahti, E. Platelet-derived growth factor receptor beta and vascular endothelial growth factor receptor 2 bind to the beta 3 integrin through its extracellular domain. J. Biol. Chem. 2000, 275, 39867–39873. [Google Scholar] [CrossRef] [PubMed]
- West, X.Z.; Meller, N.; Malinin, N.L.; Deshmukh, L.; Meller, J.; Mahabeleshwar, G.H.; Weber, M.E.; Kerr, B.A.; Vinogradova, O.; Byzova, T.V. Integrin beta3 crosstalk with VEGFR accommodating tyrosine phosphorylation as a regulatory switch. PLoS ONE 2012, 7, e31071. [Google Scholar] [CrossRef] [PubMed]
- Dan, L.; Jian, D.; Na, L.; Xiaozhong, W. Crosstalk between EGFR and integrin affects invasion and proliferation of gastric cancer cell line, SGC7901. Onco. Targets Ther. 2012, 5, 271–277. [Google Scholar] [CrossRef] [PubMed]
- Takada, Y.; Takada, Y.K.; Fujita, M. Crosstalk between insulin-like growth factor (IGF) receptor and integrins through direct integrin binding to IGF1. Cytokine Growth Factor Rev. 2017. [Google Scholar] [CrossRef] [PubMed]







© 2017 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stallcup, W.B. NG2 Proteoglycan Enhances Brain Tumor Progression by Promoting Beta-1 Integrin Activation in both Cis and Trans Orientations. Cancers 2017, 9, 31. https://doi.org/10.3390/cancers9040031
Stallcup WB. NG2 Proteoglycan Enhances Brain Tumor Progression by Promoting Beta-1 Integrin Activation in both Cis and Trans Orientations. Cancers. 2017; 9(4):31. https://doi.org/10.3390/cancers9040031
Chicago/Turabian StyleStallcup, William B. 2017. "NG2 Proteoglycan Enhances Brain Tumor Progression by Promoting Beta-1 Integrin Activation in both Cis and Trans Orientations" Cancers 9, no. 4: 31. https://doi.org/10.3390/cancers9040031
APA StyleStallcup, W. B. (2017). NG2 Proteoglycan Enhances Brain Tumor Progression by Promoting Beta-1 Integrin Activation in both Cis and Trans Orientations. Cancers, 9(4), 31. https://doi.org/10.3390/cancers9040031