
Targeting Intracellular Calcium Signaling ([Ca2+]i) to Overcome Acquired Multidrug Resistance of Cancer Cells: A Mini-Overview



{kind=link}
{kind=link}
Abstract
:1. Cancer Is a Global Public Health Problem
2. Need of New Strategies to Overcome Drug Resistance
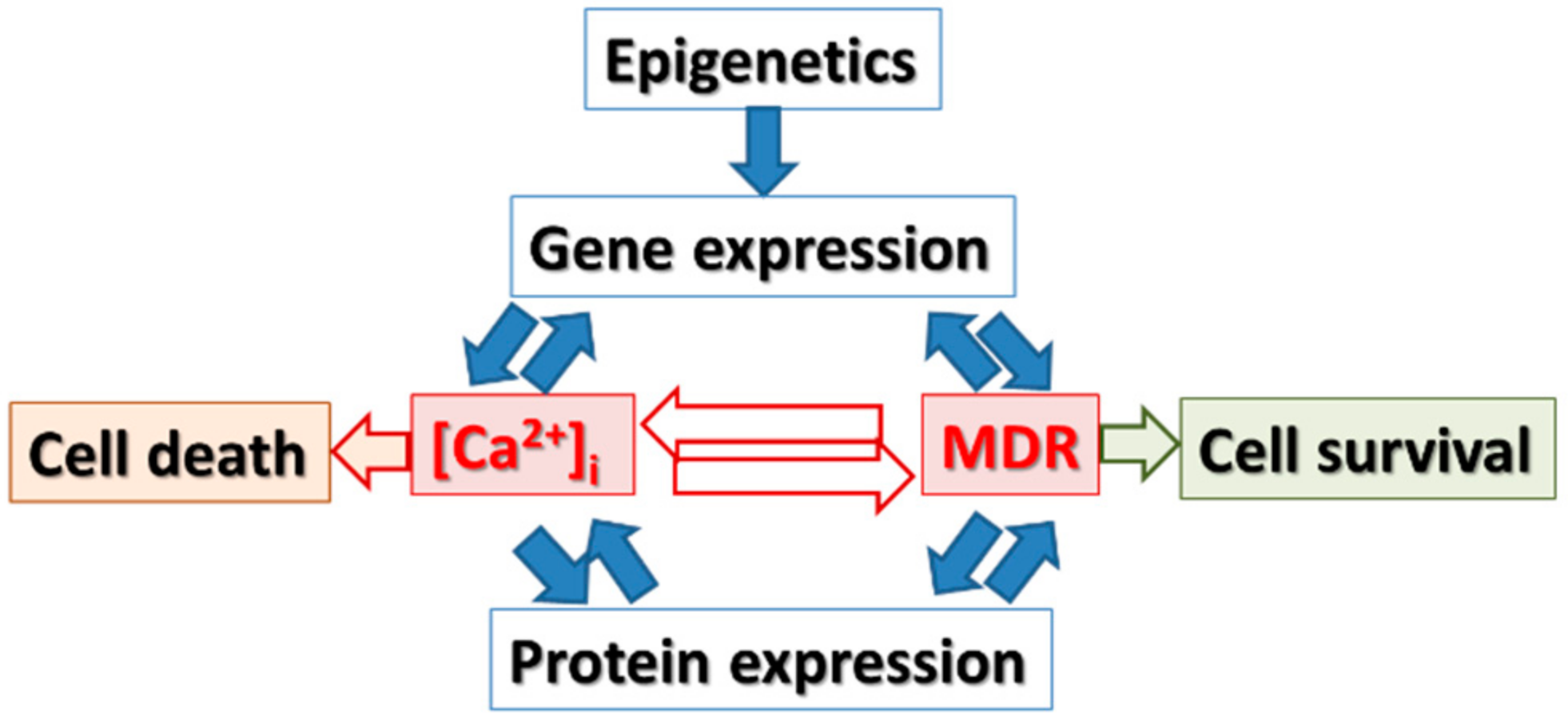
3. Hypothesis
4. Gene Expression and Epigenetic Changes Are Important for Cancer Pathogenesis: Are They Targets to Overcome Drug Resistance of Anticancer Drugs?
- (i)
- a decreased intracellular drug concentration, e.g., due to the expression and activity of drug transporters (i.e., ATP-binding cassette (ABC) transporters) or metabolic enzymes (i.e., glutathione S-transferase, cytochrome P450 enzymes);
- (ii)
- disturbances affecting the cell cycle arrest, apoptosis, and DNA repair (i.e., p53);
- (iii)
- activation of signaling pathways related to progression of cancer;
- (iv)
- epigenetic modifications (i.e., DNA methylation, miRNA, histone modification); and
- (v)
- an increase or alteration in the availability of drug targets;
- (vi)
- inactivation or compartmentalization of the agents;
- (vii)
- inhibition of apoptosis and aberrant bioactive sphingolipid metabolism;
- (viii)
- modification of gene expression
5. The Role of [Ca2+]i Signaling in the Regulation of Expressing Genes which Are Relevant to Drug Resistance
- the cell motility, tumor invasion, and metastasis;
- angiogenesis;
- genotoxicity due to [Ca2+]i modulation of the DNA damage response pathways thus controlling genomic stability and cell survival;
- transcription that takes place through [Ca2+]i oscillation frequency, or transcription factor NFAT (nuclear factor of activated T cells);
- telomerase activity (e.g., S100A8 can inhibit the activity of telomerase contributing to cell immortalization);
- differentiation due to the capability of [Ca2+]i signals to control the differentiation process through the extracellular Ca2+ sensing receptor and/or alterations in intracellular Ca2+;
- the cell cycle because [Ca2+]i is a key regulator of the cell cycle and proliferation, through regulation of Ras activity or the subcellular localization of key proteins associated with tumorigenesis (e.g., Ca2+ controls PTEN nuclear localization);
- apoptosis due to:
- (a)
- Ca2+ accumulation in the mitochondria and activation of mitochondrial membrane permeabilization; while
- (b)
- the reduction in the Ca2+ content in ER is associated with resistance to apoptosis; or
- (c)
6. The Role of Epigenetic Changes in Gene Expression, Regulation of [Ca2+]i Signaling and Drug Resistance Relevant Genes
7. Outlook
- ix.
- Identification and validation of genes that are deregulated upon chemotherapy: ((multi-) drug resistance and calcium modulating genes associated with cancer pathology (e.g., ABCB1, ABCC1, ABCG1, ITPR1, CACNG1, CACNA1D, CAMLG, CALB2, S100A, and TRPV family) (see recent work of [64]).
- x.
- The establishment and characterization of the relationship between deregulation of calcium signaling and MDR expression upon chemotherapy in cancer cells.
- xi.
- The identification and validation of selected MDR and calcium signaling related genes associated with acquired drug resistance of cancer cells.
- xii.
- The verification on how and why specific manipulations of calcium signaling in drug resistant cancer cells will overcome the drug resistance.
- xiii.
- To determine of the role of epigenetic manipulation of MDR and calcium signaling related genes will influence the acquired drug resistance of drug resistant cancer cells.
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Bener, A.; Ayub, H.; Kakil, R.; Ibrahim, W. Patterns of cancer incidence among the population of Qatar: A worldwide comparative study. Asian Pac. J. Cancer Prev. 2008, 9, 19–24. [Google Scholar]
- Al-Hamdan, N.; Ravichandran, K.; Al-Sayyad, J.; Al-Lawati, J.; Khazal, Z.; Al-Khateeb, F.; Abdulwahab, A.; Al-Asfour, A. Incidence of cancer in Gulf Cooperation Council countries, 1998–2001. East Mediterr. Health J. 2009, 15, 600–611. [Google Scholar]
- Brown, R.; Kerr, K.; Haoudi, A.; Darzi, A. Tackling cancer burden in the Middle East: Qatar as an example. Lancet Oncol. 2012, 13, e501–e508. [Google Scholar] [CrossRef]
- National Cancer Institute. Available online: https://www.cancer.gov/research/areas/global-health (accessed on 7 November 2015).
- World Health Organization. Available online: http://www.who.int/mediacentre/news/releases/2003/pr27/en/ (accessed on 7 November 2015).
- Kartal-Yandim, M.; Adan-Gokbulut, A.; Baran, Y. Molecular mechanisms of drug resistance and its reversal in cancer. Crit. Rev. Biotechnol. 2016, 36, 716–726. [Google Scholar] [CrossRef]
- Zahreddine, H.; Borden, K.L. Mechanisms and insights into drug resistance in cancer. Front Pharmacol. 2013, 4, 28. [Google Scholar] [CrossRef]
- Smith, C.I. Enigmas in tumor resistance to kinase inhibitors and calculation of the drug resistance index for cancer (DRIC). Semin. Cancer Biol. 2016. [Google Scholar] [CrossRef]
- Normanno, N.; Morabito, A.; De Luca, A.; Piccirillo, M.C.; Gallo, M.; Maiello, M.R.; Perrone, F. Target-based therapies in breast cancer: Current status and future perspectives. Endocr. Relat. Cancer 2009, 16, 675–702. [Google Scholar] [CrossRef]
- Colombo, P.E.; Milanezi, F.; Weigelt, B.; Reis-Filho, J.S. Microarrays in the 2010s: The contribution of microarray-based gene expression profiling to breast cancer classification, prognostication and prediction. Breast Cancer Res. 2011, 13, 212. [Google Scholar] [CrossRef]
- Zadran, S.; Remacle, F.; Levine, R.D. miRNA and mRNA cancer signatures determined by analysis of expression levels in large cohorts of patients. Proc. Natl. Acad. Sci. USA 2013, 110, 19160–19165. [Google Scholar] [CrossRef]
- Ciriello, G.; Miller, M.L.; Aksoy, B.A.; Senbabaoglu, Y.; Schultz, N.; Sander, C. Emerging landscape of oncogenic signatures across human cancers. Nat. Genet. 2013, 45, 1127–1133. [Google Scholar] [CrossRef]
- Iorio, M.V.; Casalini, P.; Piovan, C.; Braccioli, L.; Tagliabue, E. Breast cancer and microRNAs: Therapeutic impact. Breast 2011, 20, S63–S70. [Google Scholar] [CrossRef]
- Zwiefel, K.; Janni, W. Current standards in the treatment of breast cancer. Med. Monatsschrift Pharm. 2011, 34, 280–288, quiz 289–290. [Google Scholar]
- Florea, A.M.; Büsselberg, D. Breast cancer and possible mechanisms of therapy resistance. J. Glob. Local Health Sci. 2013, 2. [Google Scholar] [CrossRef]
- Goldhirsch, A.; Ingle, J.N.; Gelber, R.D.; Coates, A.S.; Thurlimann, B.; Senn, H.J. Thresholds for therapies: Highlights of the St. Gallen International Expert Consensus on the primary therapy of early breast cancer 2009. Ann. Oncol. 2009, 20, 1319–1329. [Google Scholar] [CrossRef]
- Schmidt, M.; Victor, A.; Bratzel, D.; Boehm, D.; Cotarelo, C.; Lebrecht, A.; Siggelkow, W.; Hengstler, J.G.; Elsasser, A.; Gehrmann, M.; et al. Long-termoutcome prediction by clinicopathological risk classification algorithms in node-negative breast cancer—Comparison between Adjuvant!, St. Gallen, and a novel risk algorithm used in the prospective randomized Node-Negative-Breast Cancer-3 (NNBC-3) trial. Ann. Oncol. 2009, 20, 258–264. [Google Scholar]
- Kim, H.K.; Choi, I.J.; Kim, C.G.; Kim, H.S.; Oshima, A.; Michalowski, A.; Green, J.E. A gene expression signature of acquired chemoresistance to cisplatin and fluorouracil combination chemotherapy in gastric cancer patients. PLoS ONE 2011, 6, e16694. [Google Scholar] [CrossRef]
- Galluzzi, L.; Senovilla, L.; Vitale, I.; Michels, J.; Martins, I.; Kepp, O.; Castedo, M.; Kroemer, G. Molecular mechanisms of cisplatin resistance. Oncogene 2012, 31, 1869–1883. [Google Scholar] [CrossRef]
- Florea, A.M.; Büsselberg, D. Cisplatin as an Anti-Tumor Drug: Cellular Mechanisms of Activity, Drug Resistance and Induced Side Effects. Cancers 2011, 3, 1351–1371. [Google Scholar] [CrossRef]
- Florea, A.M.; Büsselberg, D. Metals and breast cancer: Risk factors or healing agents? J. Toxicol. 2011, 2011, 159619. [Google Scholar] [CrossRef]
- Florea, A.M.; Büsselberg, D. Anti-cancer drugs interfere with intracellular calcium signaling. Neurotoxicology 2009, 30, 803–810. [Google Scholar] [CrossRef]
- Kutanzi, K.R.; Yurchenko, O.V.; Beland, F.A.; Checkhun, V.F.; Pogribny, I.P. MicroRNA-mediated drug resistance in breast cancer. Clin. Epigenetics 2011, 2, 171–185. [Google Scholar] [CrossRef]
- Saxena, M.; Stephens, M.A.; Pathak, H.; Rangarajan, A. Transcription factors that mediate epithelial-mesenchymal transition lead to multidrug resistance by upregulating ABC transporters. Cell Death Dis. 2011, 2, e179. [Google Scholar] [CrossRef]
- Karthikeyan, S.; Hoti, S.L. Development of Fourth Generation ABC Inhibitors from Natural Products: A Novel Approach to Overcome Cancer Multidrug Resistance. Anticancer Agents Med. Chem. 2015, 15, 605–615. [Google Scholar] [CrossRef]
- Choi, Y.H.; Yu, A.M. ABC transporters in multidrug resistance and pharmacokinetics, and strategies for drug development. Curr. Pharm. Des. 2014, 20, 793–807. [Google Scholar] [CrossRef]
- Liu, Y.; Peng, H.; Zhang, J.T. Expression profiling of ABC transporters in a drug-resistant breast cancer cell line using AmpArray. Mol. Pharmacol. 2005, 68, 430–438. [Google Scholar] [CrossRef]
- Gillet, J.P.; Efferth, T.; Steinbach, D.; Hamels, J.; de Longueville, F.; Bertholet, V.; Remacle, J. Microarray-based detection of multidrug resistance in human tumor cells by expression profiling of ATP-binding cassette transporter genes. Cancer Res. 2004, 64, 8987–8993. [Google Scholar] [CrossRef]
- Perou, C.M.; Sorlie, T.; Eisen, M.B.; van de Rijn, M.; Jeffrey, S.S.; Rees, C.A.; Pollack, J.R.; Ross, D.T.; Johnsen, H.; Akslen, L.A.; et al. Molecular portraits of human breast tumours. Nature 2000, 406, 747–752. [Google Scholar] [CrossRef]
- Sorlie, T.; Perou, C.M.; Tibshirani, R.; Aas, T.; Geisler, S.; Johnsen, H.; Hastie, T.; Eisen, M.B.; van de Rijn, M.; Jeffrey, S.S.; et al. Gene expression patterns of breast carcinomas distinguish tumor subclasses with clinical implications. Proc. Natl. Acad. Sci. USA 2001, 98, 10869–10874. [Google Scholar] [CrossRef]
- Monteith, G.R.; McAndrew, D.; Faddy, H.M.; Roberts-Thomson, S.J. Calcium and cancer: Targeting Ca2+ transport. Nat. Rev. Cancer 2007, 7, 519–530. [Google Scholar] [CrossRef]
- Monteith, G.R.; Davis, F.M.; Roberts-Thomson, S.J. Calcium channels and pumps in cancer: Changes and consequences. J. Biol. Chem. 2012, 287, 31666–31673. [Google Scholar] [CrossRef]
- Cmoch, A.; Groves, P.; Palczewska, M.; Pikuła, S. S100A proteins in propagation of a calcium signal in norm and pathology. Postepy Biochem. 2012, 58, 429–436. [Google Scholar]
- Capiod, T. The need for calcium channels in cell proliferation. Recent Pat. Anticancer Drug Discov. 2013, 8, 4–17. [Google Scholar] [CrossRef]
- Chen, Y.F.; Chen, Y.T.; Chiu, W.T.; Shen, M.R. Remodeling of calcium signaling in tumor progression. J. Biomed. Sci. 2013, 20, 23. [Google Scholar] [CrossRef]
- Zhou, Y.; Xue, S.; Yang, J.J. Calciomics: Integrative studies of Ca2+-binding proteins and their interactomes in biological systems. Metallomics 2013, 5, 29–42. [Google Scholar] [CrossRef]
- Lee, J.M.; Davis, F.M.; Roberts-Thomson, S.J.; Monteith, G.R. Ion channels and transporters in cancer. 4. Remodeling of Ca(2+) signaling in tumorigenesis: Role of Ca2+ transport. Am. J. Physiol. Cell Physiol. 2011, 301, C969–C976. [Google Scholar] [CrossRef]
- Naz, S.; Bashir, M.; Ranganathan, P.; Bodapati, P.; Santosh, V.; Kondaiah, P. Pro tumorigenic actions of S100A2 involves regulation of PI3/Akt signaling and functional interaction with Smad3. Carcinogenesis 2013, 35, 14–23. [Google Scholar] [CrossRef]
- Singh, N.; Promkan, M.; Liu, G.; Varani, J.; Chakrabarty, S. Role of calcium sensing receptor (CaSR) in tumorigenesis. Best Pract. Res. Clin. Endocrinol. Metab. 2013, 27, 455–463. [Google Scholar] [CrossRef]
- Diez-Fraile, A.; Lammens, T.; Benoit, Y.; D’Herde, K.G. The calcium-sensing receptor as a regulator of cellular fate in normal and pathological conditions. Curr. Mol. Med. 2013, 13, 282–295. [Google Scholar] [CrossRef]
- Racioppi, L.; Means, A.R. Calcium/calmodulin-dependent protein kinase kinase 2: Roles in signaling and pathophysiology. J. Biol. Chem. 2012, 287, 31658–31665. [Google Scholar] [CrossRef]
- Berchtold, M.W.; Villalobo, A. The many faces of calmodulin in cell proliferation, programmed cell death, autophagy, and cancer. Biochim. Biophys. Acta 2014, 1843, 398–435. [Google Scholar] [CrossRef]
- Frede, J.; Fraser, S.P.; Oskay-Özcelik, G.; Hong, Y.; Ioana Braicu, E.; Sehouli, J.; Gabra, H.; Djamgoz, M.B. Ovarian cancer: Ion channel and aquaporin expression as novel targets of clinical potential. Eur. J. Cancer 2013, 49, 2331–2344. [Google Scholar] [CrossRef]
- Akl, H.; Bultynck, G. Altered Ca2+ signaling in cancer cells: Proto-oncogenes and tumor suppressors targeting IP3 receptors. Biochim. Biophys. Acta 2013, 1835, 180–193. [Google Scholar] [CrossRef]
- Marchi, S.; Pinton, P. Alterations of calcium homeostasis in cancer cells. Curr. Opin. Pharmacol. 2016, 29, 1–6. [Google Scholar] [CrossRef]
- Stathopulos, P.B.; Ikura, M. Store operated calcium entry: From concept to structural mechanisms. Cell Calcium 2016. [Google Scholar] [CrossRef]
- Jardin, I.; Rosado, J.A. STIM and calcium channel complexes in cancer. Biochim. Biophys. Acta 2016, 1863, 1418–1426. [Google Scholar] [CrossRef]
- Wen, J.; Huang, Y.C.; Xiu, H.H.; Shan, Z.M.; Xu, K.Q. Altered expression of stromal interaction molecule (STIM)-calcium release-activated calcium channel protein (ORAI) and inositol 1,4,5-trisphosphate receptors (IP3Rs) in cancer: Will they become a new battlefield for oncotherapy? Chin. J. Cancer 2016, 35. [Google Scholar] [CrossRef]
- Déliot, N.; Constantin, B. Plasma membrane calcium channels in cancer: Alterations and consequences for cell proliferation and migration. Biochim. Biophys. Acta 2015, 1848, 2512–2522. [Google Scholar] [CrossRef]
- Lastraioli, E.; Iorio, J.; Arcangeli, A. Ion channel expression as promising cancer biomarker. Biochim. Biophys. Acta 2015, 1848, 2685–2702. [Google Scholar] [CrossRef]
- Fiorio Pla, A.; Kondratska, K.; Prevarskaya, N. STIM and ORAI proteins: Crucial roles in hallmarks of cancer. Am. J. Physiol. Cell Physiol. 2016, 310, C509–C519. [Google Scholar] [CrossRef]
- Chen, Y.F.; Hsu, K.F.; Shen, M.R. The store-operated Ca2+ entry-mediated signaling is important for cancer spread. Biochim. Biophys. Acta 2016, 1863, 1427–1435. [Google Scholar] [CrossRef]
- Anguita, E.; Villalobo, A. Src-family tyrosine kinases and the Ca2+ signal. Biochim. Biophys. Acta 2016. [Google Scholar] [CrossRef]
- Nguyen, O.N.; Grimm, C.; Schneider, L.S.; Chao, Y.K.; Atzberger, C.; Bartel, K.; Watermann, A.; Ulrich, M.; Mayr, D.; Wahl-Schott, C.; et al. Two-Pore Channel Function Is Crucial for the Migration of Invasive Cancer Cells. Cancer Res. 2017, 77, 1427–1438. [Google Scholar] [CrossRef]
- Day, T.K.; Bianco-Miotto, T. Common gene pathways and families altered by DNA methylation in breast and prostate cancers. Endocr. Relat. Cancer 2013, 20, R215–R232. [Google Scholar] [CrossRef]
- Ouadid-Ahidouch, H.; Rodat-Despoix, L.; Matifat, F.; Morin, G.; Ahidouch, A. DNA methylation of channel-related genes in cancers. Biochim. Biophys. Acta 2015, 1848, 2621–2628. [Google Scholar] [CrossRef]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef]
- Trimarchi, M.P.; Mouangsavanh, M.; Huang, T.H. Cancer epigenetics: A perspective on the role of DNA methylation in acquired endocrine resistance. Chin. J. Cancer 2011, 30, 749–756. [Google Scholar] [CrossRef]
- Hill, V.K.; Ricketts, C.; Bieche, I.; Vacher, S.; Gentle, D.; Lewis, C.; Maher, E.R.; Latif, F. Genome-wide DNA methylation profiling of CpG islands in breast cancer identifies novel genes associated with tumorigenicity. Cancer Res. 2011, 71, 2988–2999. [Google Scholar] [CrossRef]
- Cai, F.F.; Kohler, C.; Zhang, B.; Wang, M.H.; Chen, W.J.; Zhong, X.Y. Epigenetic therapy for breast cancer. Int. J. Mol. Sci. 2011, 12, 4465–4487. [Google Scholar] [CrossRef]
- Dedeurwaerder, S.; Fumagalli, D.; Fuks, F. Unravelling the epigenomic dimension of breast cancers. Curr. Opin. Oncol. 2011, 23, 559–565. [Google Scholar] [CrossRef]
- Spitzwieser, M.; Pirker, C.; Koblmüller, B.; Pfeiler, G.; Hacker, S.; Berger, W.; Heffeter, P.; Cichna-Markl, M. Promoter methylation patterns of ABCB1, ABCC1 and ABCG2 in human cancer cell lines, multidrug-resistant cell models and tumor, tumor-adjacent and tumor-distant tissues from breast cancer patients. Oncotarget 2016, 7, 73347–73369. [Google Scholar] [CrossRef]
- Wang, X.X.; Xiao, F.H.; Li, Q.G.; Liu, J.; He, Y.H.; Kong, Q.P. Large-scale DNA methylation expression analysis across 12 solid cancers reveals hypermethylation in the calcium-signaling pathway. Oncotarget 2017, 8. [Google Scholar] [CrossRef]
- Florea, A.-M.; Varghese, E.; McCallum, J.E.; Mahgoub, S.; Helmy, I.; Varghese, S.; Gopinath, N.; Sass, S.; Theis, F.J.; Reifenberger, G.; et al. Calcium-regulatory proteins as modulators of chemotherapy in human neuroblastoma. Oncotarget 2017, 8, 22876–22893. [Google Scholar] [CrossRef]


© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Büsselberg, D.; Florea, A.-M. Targeting Intracellular Calcium Signaling ([Ca2+]i) to Overcome Acquired Multidrug Resistance of Cancer Cells: A Mini-Overview. Cancers 2017, 9, 48. https://doi.org/10.3390/cancers9050048
Büsselberg D, Florea A-M. Targeting Intracellular Calcium Signaling ([Ca2+]i) to Overcome Acquired Multidrug Resistance of Cancer Cells: A Mini-Overview. Cancers. 2017; 9(5):48. https://doi.org/10.3390/cancers9050048
Chicago/Turabian StyleBüsselberg, Dietrich, and Ana-Maria Florea. 2017. "Targeting Intracellular Calcium Signaling ([Ca2+]i) to Overcome Acquired Multidrug Resistance of Cancer Cells: A Mini-Overview" Cancers 9, no. 5: 48. https://doi.org/10.3390/cancers9050048
APA StyleBüsselberg, D., & Florea, A. -M. (2017). Targeting Intracellular Calcium Signaling ([Ca2+]i) to Overcome Acquired Multidrug Resistance of Cancer Cells: A Mini-Overview. Cancers, 9(5), 48. https://doi.org/10.3390/cancers9050048