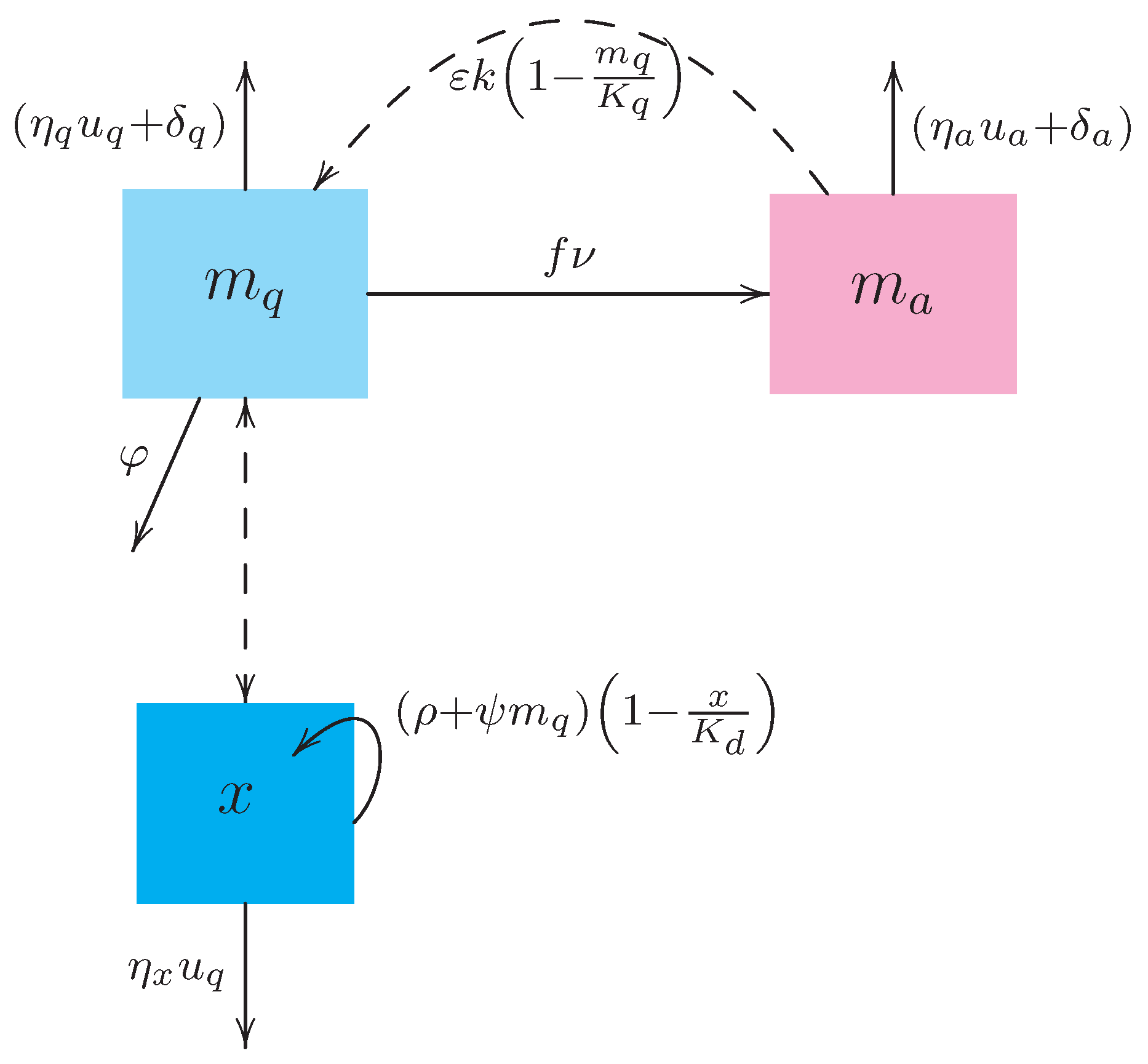
4.1. Preliminary Settings
In accordance with estimations performed in Reference [
32] for average daily temperature (23.9 °C) in the city of Cali, Colombia, and other data found in scientific literature,
Table 2 displays numerical values assigned to all constant parameters of the dynamical system (
1) which will be kept unaltered during the computational experiments involving numerical solutions of the optimal control problem (
10).
It is worth noting that, for parameter values from
Table 2, the basic offspring number defined by (
4a) is
meaning that one female mosquito produces on average 38 female mosquitoes in the absence of predacious species. This value matches the estimations of the basic offspring number obtained in other studies [
41].
A shallow basin we consider here has the volume of about 100 liters, and a maximal larval density is 8-10 individuals per liter [
34] what gives us the carrying capacity
(measured in thousands of individuals). We also assume the following stylized values related to the predacious species
where
(measured by the maximal number of individuals to be sustained by the environment) is defined to match the predator-prey ratio
in accordance with [
42]. To assume an abundance of immature and adult mosquitoes before the control intervention, we define
and
and also consider two options for
with
expressing the absence of predacious species during the control intervention, and
mimicking the introduction of three individuals (one male and two females) into the basin before the control intervention.
For all numerical experiments, we set meaning that the control intervention will remain in force during 30 days.
To perform numerical simulations we have employed the next-generation optimal control software package GPOPS-II
1 designed for MATLAB platform [
30] that implements an adaptive combination of direct and orthogonal collocation techniques known as Radau pseudospectral method [
43].
Our goal is to analyze different scenarios, whose determination results from the following considerations. First, there are scenarios where the biological control is either absent (
) or present (
). For the latter, we perform testings with two predacious species—one that is relatively cheap but does not fulfill the condition (
5) of predatory efficiency (to be referred further as inefficient or inert predator), and another that fulfills the condition (
5) but comes at a higher cost (to be called efficient or aggressive predator in the sequel). The unit cost (i.e., per individual) for both types of predacious species is denoted by
. As stated in Reference [
13], different larvivorous fish species constitute an example of inefficient or inert predators, while cyclopoid copepods are usually regarded as aggressive and efficient ones.
Second, there are scenarios either accounting or not for the chemical controls actions applied to adult insects only (insecticide spraying) or their immature stages only (larvicide spraying), or to both mosquito populations (combined use of insecticides and larvicides). Additionally, we consider two types of each chemical substance: one bearing high lethality and costs, and another bearing low lethality with moderate underlying costs.
In this context, it is worth recalling that the lethality of existent insecticides and larvicides admits variation between 20% and 80% [
44,
45,
46]; therefore, we suppose 30% efficiency for low-lethality substances and 70% efficiency for high-lethality substances. Such efficiencies are modeled by the parameters
(for insecticides) and
(for larvicides) in the dynamical system (
1), while the underlying (unit) costs of larvicide and insecticide are expressed, respectively, by
and
in the objective functional (
8). Furthermore, we assume that the upper bound of both control functions
are normalized to unity:
. The latter implies that the daily use of each chemical substance cannot exceed certain quantities
and
determined externally by the environmental regulations, that is,
Thus, and are the fractions of and to be used at each day of the chemical control intervention, and therefore . Note that the quantities and may have different values for larvicides and insecticides bearing lower or higher lethality; this is another reason why we have introduced normalization for and .
We also assume that the negative effect of larvicides on the predacious species (expressed in (
1) by the parameter
) increases with the larvicide’s lethality.
Finally, since our primary goal is to suppress the total mosquito population (and thus reduce the local incidence of dengue and other vector-borne diseases), in the integrand of the objective functional (
8), we assign the highest value to the weight coefficient
. Thus,
doubles the unit cost of the most expensive chemical control. As for the weight coefficient accompanying the terminal part of (
8), we set
to make it the largest and also offset the
T days of intervention.
4.2. Description of Considered Scenarios
In the previous subsection, we assigned numerical values to the majority of the constant parameters of the optimal control problem (
10) which will be kept unaltered during the computational experiments. However, numerical values for several parameters (namely, for
and
) have not been determined yet. These are exactly the parameters whose values will be varied in order to express different scenarios and to obtain the optimal strategies corresponding to each particular scenario.
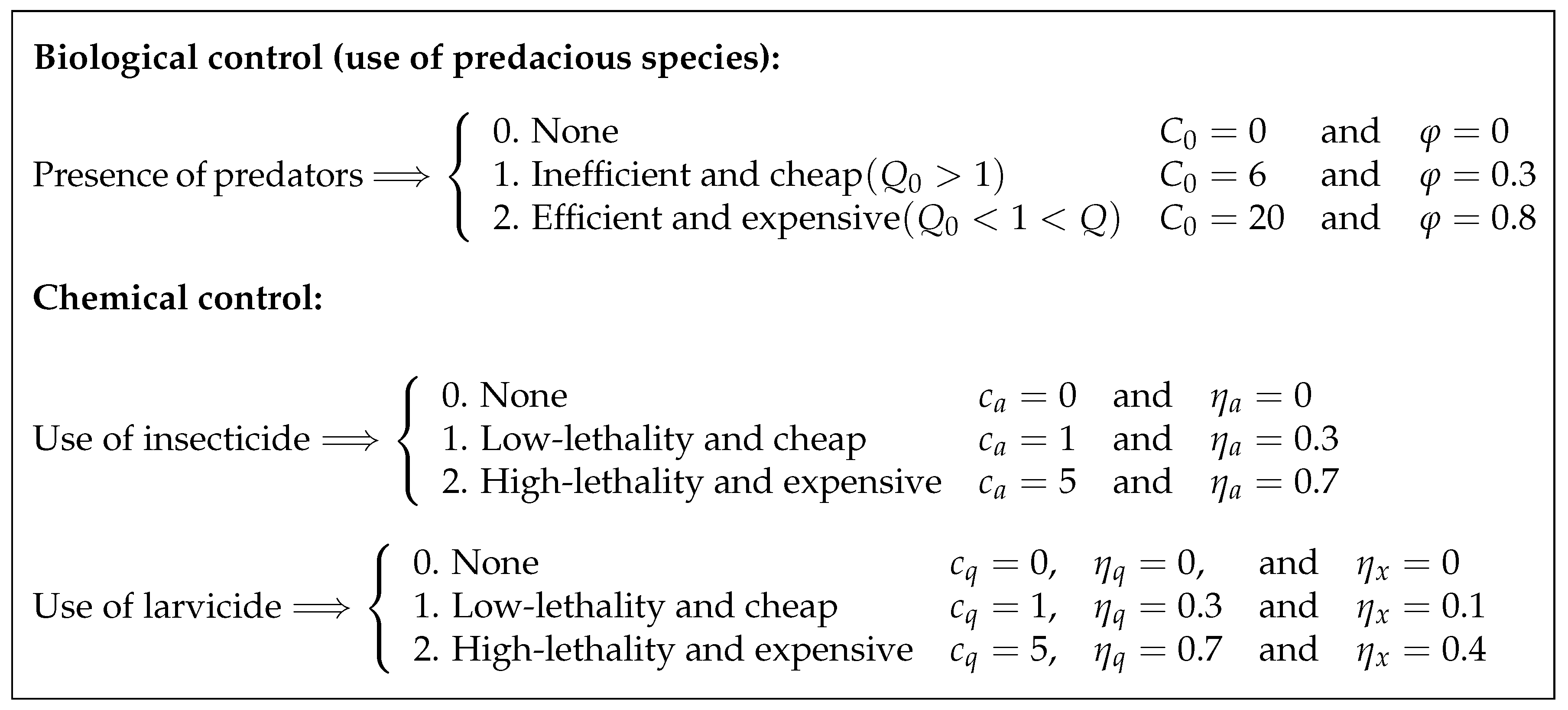
Altogether, we intend to explore 27 scenarios and to establish 27 corresponding strategies (schematically displayed in
Figure 2) which are further codified using three digits (
and 2) referring to the use of predacious species (first digit) for biological control, use of insecticide (second digit) on the adult population, and application of larvicide (third digit) on immature stages. Thus, the strategy denoted by 012 implies no use of predacious species and application of low-lethality insecticide in combination with high-lethality larvicide, while the strategy 000 corresponds to the absence of control intervention and will be referred to as “baseline case” in the sequel.
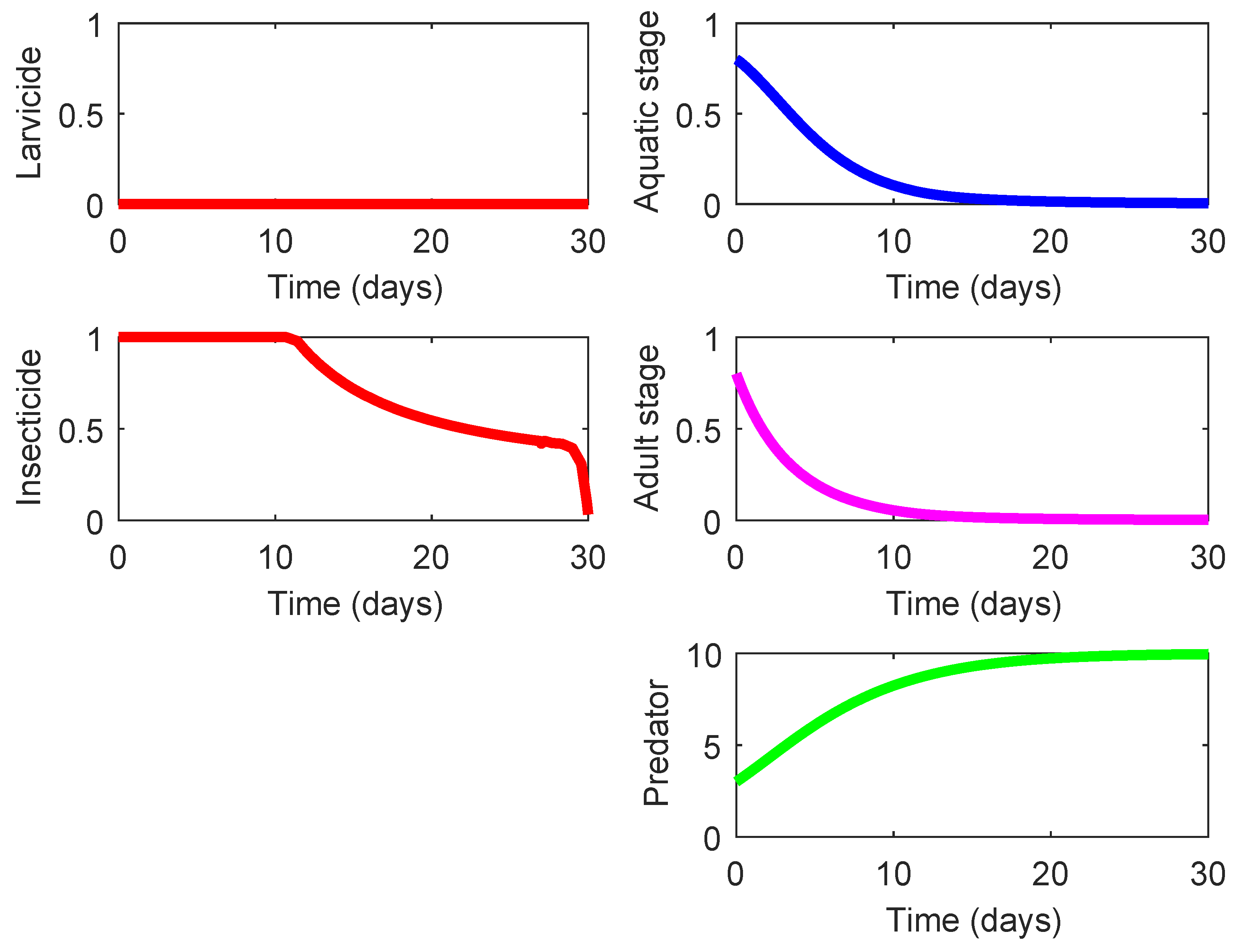
Figure 3 presents the profiles
and
corresponding to the baseline case (Strategy 000).
It is worthwhile to point out that we do not have any reliable information regarding the unit costs of insecticides and larvicides bearing different lethalities. Therefore, in view of the rationale given in References [
28,
29], the values assigned to the weight coefficients
and
(as displayed in the chart of
Figure 2) are taken only for the purpose of theoretical analysis; for practical purposes, these values should be adjusted to more realistic ones.
To characterize performance of each strategy, we evaluate the following important quantities for each run of GPOPS-II solver feeded diverse combinations of values
as indicated in
Figure 2:
The density of immature stages at the end of the intervention:
The cumulative density of adult females during the whole period of intervention: .
Cumulative fraction of larvicide used:
Cumulative fraction of insecticide used:
The total cost of the strategy that combines expenses related to the initial introduction of a predacious species and the costs of chemical control measures, that is,
In the above expressions,
denote the optimal states of the system (
1) corresponding to the optimal controls
delivered by the GPOPS-II solver under different scenarios described in
Figure 2. It is worthwhile to point out that the baseline case (Strategy 000) and two scenarios not relying upon the use of chemical substances (Strategies 100 and 200) do not require for numerical solutions of the optimal control problem (
10). Therefore, to obtain
and
corresponding to Strategies 100 and 200 it suffices to solve numerically the system (
1) with
and
.
The outcomes displayed in
Table 3 plainly indicate that Strategy 001 is the cheapest, while Strategies
guarantee the best results for suppression of the total mosquito population by the end of the control intervention. Among them, Strategy 220 does not require the use of larvicide and thus bears a lower total cost, while Strategy 221 appears a bit cheaper than Strategy 222 and requires a lesser amount of insecticide.
Note that for the baseline case (Strategy 000) we have
while its underlying cost is zero since no control action is applied.
Now, we are interested in determining which strategy or strategies may serve better the needs of the decision-makers. In order to proceed, we have to assess and quantify the benefits rendered by each strategy in order to employ the so-called cost-effectiveness approach to the result of numerical solutions of the optimal control problem in the sense of References [
28,
47]. In the following subsection, we perform the cost-effectiveness analysis of the strategies displayed in
Table 3.
4.3. Cost-Effectiveness Analysis
In economics, the cost-effectiveness analysis is the method that allows comparing the relative costs and outcomes (effects or benefits) of different courses of action. Using this approach, one can fairly assess an additional benefit that can be obtained by investing a unit of cost into a certain action in comparison to either no action taken or implementing a different action.
To employ this approach, it is necessary to quantify the relative costs and benefits of each strategy that models an external intervention. The relative costs of control strategies described by 27 scenarios given in
Figure 2 can be assessed by
(see Formula (
19) and the last column of
Table 3), while some additional considerations will be needed to quantify the benefit of each strategy.
First, let us recall that the primary goal of the control intervention consists in minimizing the density of immature stages by the end of the intervention (cf. terminal term in (
8)) and reducing the density of adult mosquitoes during the whole period of intervention (cf. first summand of the integrand in (
8)). Therefore, the effect of each strategy on the suppression of mosquito population can be quantified as
where
and
are defined by (
20) in regards to the baseline case (that is, without control intervention,
) while
and
correspond to the optimal states of the dynamical system (
1) for each optimal strategy (cf. first column of
Table 3).
The secondary goal of the decision-making (expressed by the last two summands of the integrand in (
8)) consists of minimizing the use of larvicide and insecticide since these chemical substances may have an adverse environmental effect besides generating additional costs. Therefore, we would also like to identify a strategy (or strategies) capable of reducing the mosquito population with a relatively low impact on the environment. To do so, it is plausible to assume that the negative environmental impact of these chemical substances is proportional to their quantities used for the strategy’s implementation. Let us recall that the maximal quantity of the larvicide (resp. insecticide) allowed to be used at each day
of control intervention is
(resp. to
). Therefore, the maximum amounts of both chemicals allowed to be used during the period
of control intervention cannot exceed
and
, respectively. By assuming that the quantities of larvicide and insecticide are directly related to their adverse effects on the environment, one may consider as an additional ecological benefit of each strategy the relative quantity of each chemical which is
not spent during the period of intervention. Under this setting, the eco-effect of each strategy can be quantified as
It is worth noting that, according to the above formula, the highest possible eco-benefit
is assigned to two strategies (coded as 100 and 200) that do not require the use of chemical substances. The combined effect
of each particular strategy can be then defined as a sum of
and
leading us to the following formula
Once the relative costs and effects are defined for each strategy, the cost-effectiveness analysis can be performed. One of the main indicators frequently used in this type of analysis is the so-called “Average Cost-Effectiveness Ratio” (or ACER) that expresses the average cost related to obtaining one unit of potential benefit by employing the underlying strategy [
28,
47]. In formal terms, ACER is the ratio of the cost to benefit of a particular control action in comparison to no action (i.e., baseline case), that is
where “Strategy X” can be replaced by one of the coded strategies given in the first column of
Table 3. Note that the costs corresponding to all considered strategies 001-222 are already quantified as
by means of the formula (
19) and their underlying values are given in the last column of
Table 3.
According to (
24), smaller values of ACER indicate higher efficiency of the control action in the sense that one unit of potential effect (or benefit) comes at a lower cost.
Table 4 displays the effects quantified by Formulas (
21) and (
23) together with underlying values of two ACER types evaluated for all strategies 001-222. In particular,
ACER (column 4 in
Table 4) gives the average cost-effectiveness ratio that only encompasses the effect of each strategy on the suppression of mosquito population, whereas
ACER (last column in
Table 4) also accounts for the eco-effect of each strategy.
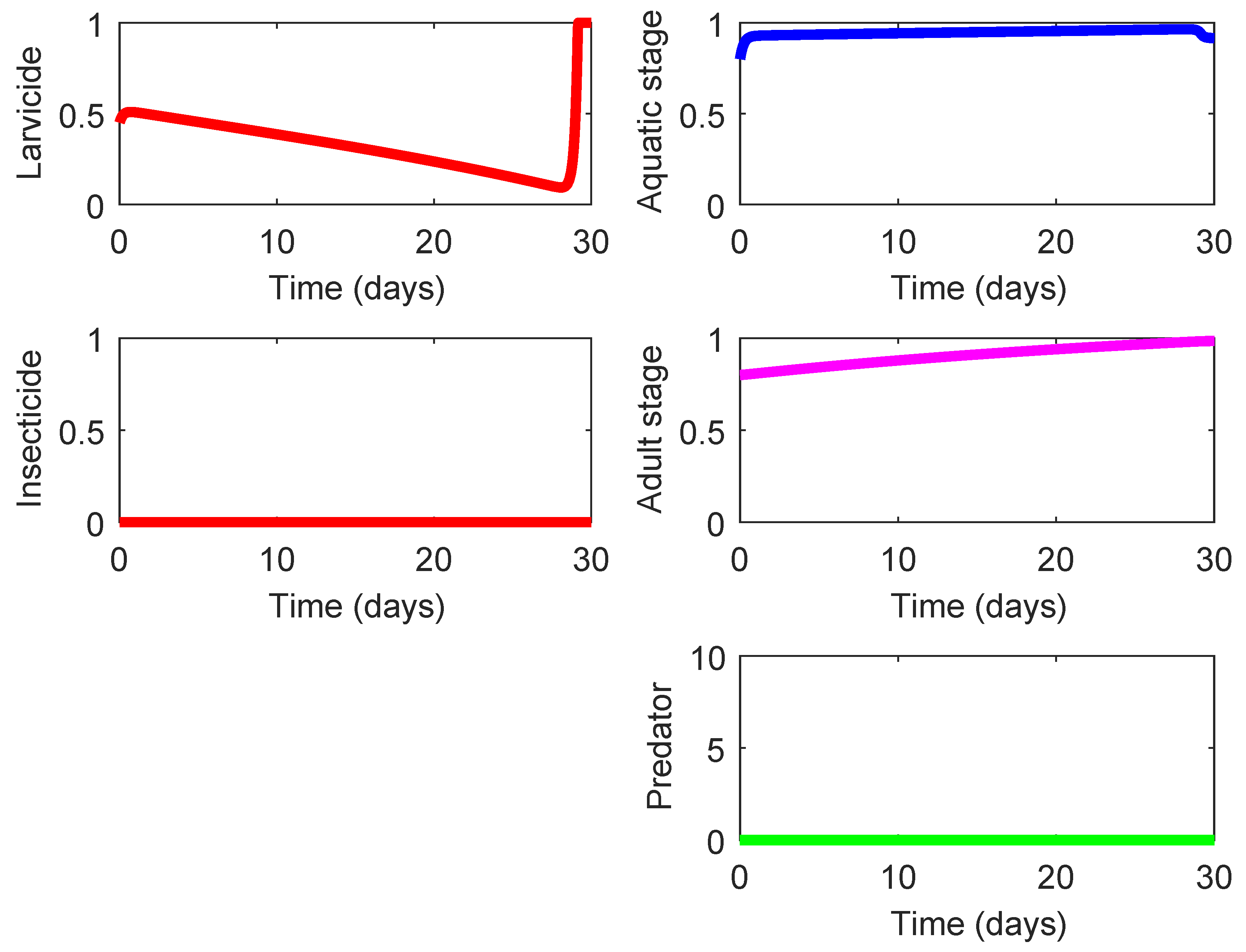
According to the total costs invested in the control intervention (see the first column of
Table 4), the cheapest strategy is the one coded as 001 (application of low-lethality larvicide in the absence of the predacious species, see
Figure 4); however, its effect on suppression of the mosquito population,
, is also the lowest comparing to other strategies. As shown on the left upper chart of
Figure 4, this strategy requires a moderate use of low-lethality larvicide (that is relatively cheap) during the major part of the control intervention (
and the application of the maximum quantity of the larvicide during the last two days (
).
From the practical standpoint, this
modus operandi evokes some doubts since the profiles of both mosquito populations
and
corresponding to Strategy 001 bear little difference from the uncontrolled case (baseline strategy, cf.
Figure 3), except for the last two days where a “strong action” is put in place. The latter is done as an attempt to minimize the terminal term of the functional (
8) to compensate for a rather weak effect of this strategy on the reduction of mosquito population. It is also worth noting that Strategy 001 requires for a relatively small amount of larvicide and does not require the use of insecticide. Due to this and the lowest cost, the average cost-effectiveness ratio of this strategy accounting for its eco-effect,
ACER, also exhibits the best result in comparison to other strategies.
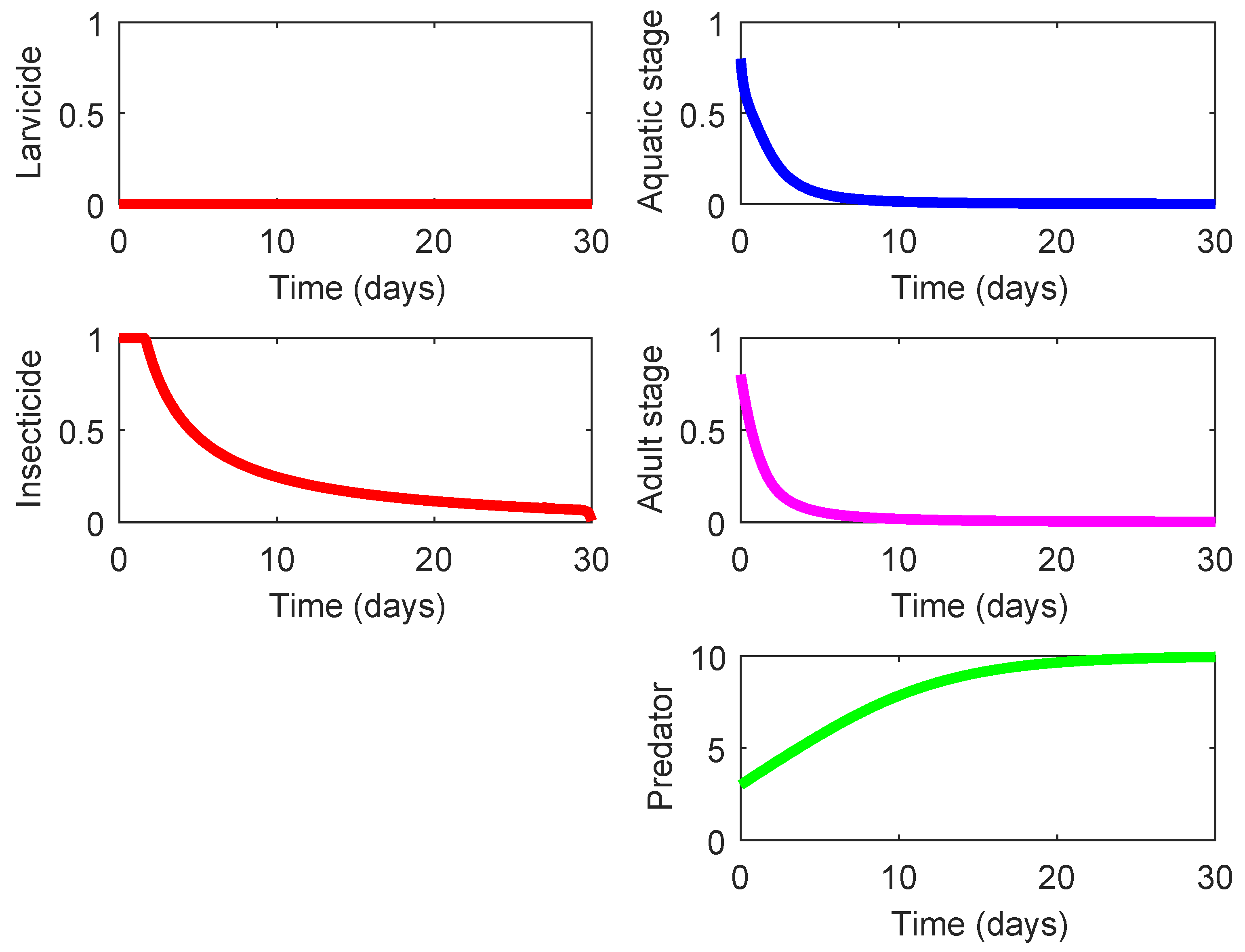
The last three rows in
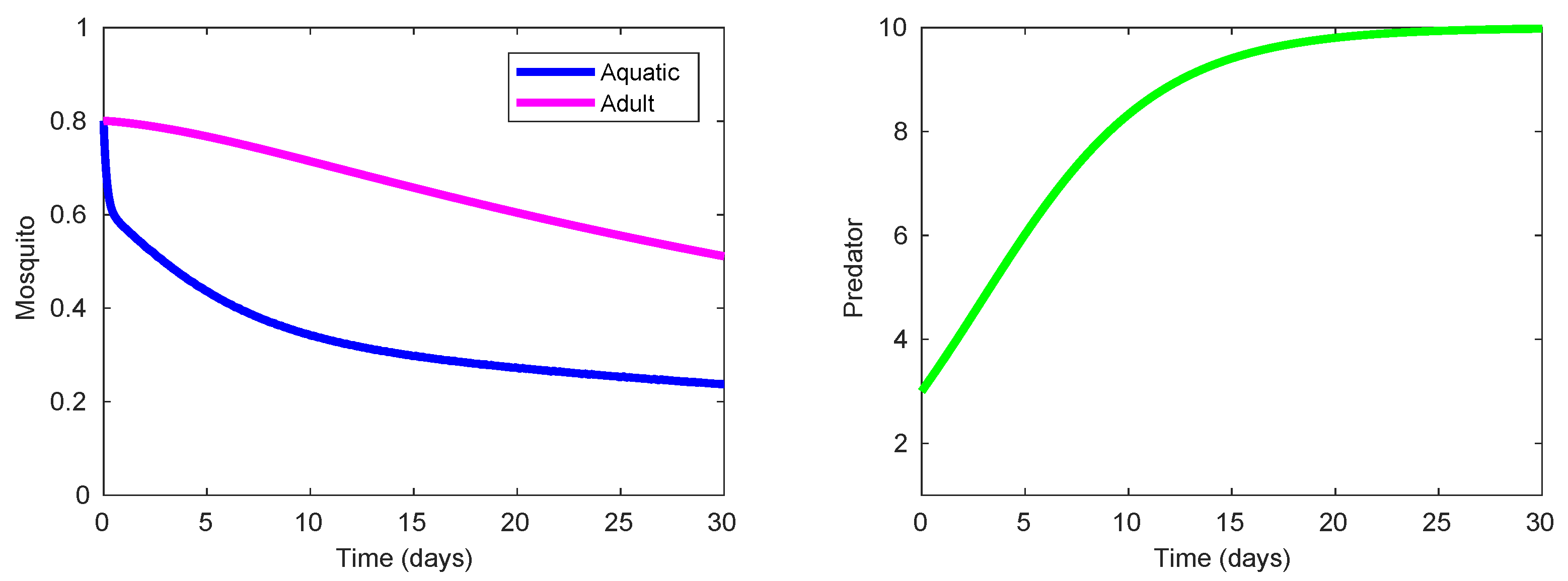
Table 4 display three strategies (Strategies 220, 221, and 222) that demonstrate the highest effect on suppression of the mosquito population and also exhibit a notable eco-effect. Among them, Strategy 220 (consisting in the use of high-lethality insecticide in the presence of efficient predacious species, see
Figure 5) possesses a slightly higher of
ACER since this strategy does not require the larvicide spraying unlike the other two strategies. As shown in
Figure 5, the presence of some efficient predacious specie combined with a moderate spraying of high-lethality insecticide guarantees steady decline of both mosquito population. The results for Strategies 221 and 222 are very similar to those presented in
Figure 5, and they are omitted here. The only difference is exhibited by the nonzero control profile
according to which the larvicide spraying should by employed (at a moderate mode) during the last couple of days in order to ensure the minimization of the terminal term in the objective functional (
8).
It should be pointed out that, besides rendering the best effects, Strategies 220, 221, and 222 also involve significant costs (10-times higher than the cost of Strategy 001), and for that reason, their ACER values are considerably larger than for other strategies.
In this context, one can easily detect in
Table 4 that Strategy 110 (consisting of spraying of low-lethality and cheap insecticide in the presence of inefficient or inert predacious species, see
Figure 6) exhibits the best value of
ACER related to the suppression of the mosquito population. As shown in
Figure 6, Strategy 110 requires to use a considerable amount of low-lethality insecticide that should be sprayed at the maximum daily rate during the first 12 days of control intervention, with a gradual reduction for the consequent 17 days, followed by total suspension at the end of intervention period. For that reason, the value of
ACER corresponding to the Strategy 110 is greater than for other strategies that require a smaller total amount of chemical substances.
We have already mentioned that the cheapest strategy (Strategy 001, see
Figure 4) is the one rendering the smallest benefit with regards to the suppression of the mosquito population. By revising the second column of
Table 4, we can identify that Strategy 100 (see
Figure 7 is also relatively cheap but its corresponding effect
is almost 10 times greater that that of Strategy 001. Moreover, Strategy 100 is eco-friendly because it does not require the use of chemical substances (
and
). For that reason, its average cost-effectiveness ratio
ACER that accounts for the eco-effect is the lowest besides the
ACER for Strategy 001.
Using the values of ACER (see the fourth and last columns in
Table 4) one can fairly assess the cost to be invested in each intervention strategy for obtaining one unit of the corresponding effect or benefit in comparison to the baseline case (i.e., no intervention and zero cost). On the other hand, it seems also useful to compare mutually exclusive strategies with each other.
For this purpose, there is another standard indicator which is known as
Incremental Cost-Effectiveness Ratio (or ICER). This indicator is defined by the difference in cost between two possible interventions, divided by the difference in their effect [
28,
47]. The value of ICER represents the average incremental cost associated with one additional unit of the measure of effect when two mutually exclusive strategies are compared. The value of ICER can be estimated as
and its calculation starts with the cheapest strategy. Thus, the value of ICER measures an additional cost per unit of additional outcome (effect or benefit) when the current Strategy X (bearing lower costs) is replaced by a new Strategy Y which is more expensive but renders a more notable effect.
In the first instance, let us calculate ICER for Strategies 001-222 taking into account only their effects on the suppression of mosquito population,
. The initial step is to arrange all strategies in ascending order regarding their costs and effects (given in the second and third columns in
Table 4). Strategies that do not comply with this order should be removed. The results are given in
Table 5 where Strategies 002, 011-022, 111-112, and 200-212 have been tossed out as non-abiding to the above-stated rule. Note that the cheapest strategy (in our case, Strategy 001) is compared with the baseline case (Strategy 000 without control intervention) and, therefore, its ICER coincides with its ACER.
The next step consists of eliminating all strategies whose
ICER-values do not comply with ascending order, as well as those with indefinite
ICER (i.e., strategies having
). For the set of remaining strategies, the values of
ICER are calculated again according to (
25) until there are no more strategies to be removed. Note that the lowest
ICER-value in the last column of
Table 5 corresponds to Strategy 110. Therefore, all strategies that standing above Strategy 110 in
Table 5 should be removed. As a result, we obtain the list of potentially cost-effective strategies (see
Table 6) where the last column indicates the estimated cost of one unit of outcome
when the current strategy is replaced by the consequent one. In particular, if Strategy 110 is replaced by Strategy 121, each unit of additional benefit will come at an additional (relative) cost of
units, which seems very high keeping in mind that the original (relative) cost per unit of benefit (that is, under Strategy 110) is just
units.
From the results displayed in
Table 6, we may conclude that Strategy 110 (use of the low-lethality insecticide in the presence of inefficient or inert predacious species, see its overall performance in
Figure 6) is indeed the most cost-effective one (according to both ACER and ICER) when the effects of all considered strategies are estimated only by their capacities for suppression of the mosquito population without accounting for their eco-effects.
A similar ICER-based analysis can be performed by encompassing the effects of Strategies 001-222 (quantified by
, see the fifth column of
Table 4) that also account for their eco-effects. For this purpose, all Strategies 001-222 should be arranged in ascending order with regards to their cost and effects (given in the second and fifth columns in
Table 4), and the strategies not complying with such an order must be removed. According to results presented in
Table 7, there is only one strategy (namely, Strategy 102) that does not comply with ascendant order for the
ICER-values (see the last column of
Table 7), and this strategy should be removed.
The final results, presented in
Table 8, indicate that five (out of six) potentially cost-effective strategies rely upon the use of only one chemical substance (either insecticide or larvicide), while the remaining one needs no chemical intervention (Strategy 100). This outcome plainly agrees with the way the strategies’ effects are quantified. Here, one should recall that
of each strategy accounts for its environmental impact, besides measuring its capacity for suppression of the mosquito population.
Let us recall that the best feature of Strategy 001 is its lowest cost
; however, this same strategy has the lowest effect
on the suppression of mosquito population (see
Figure 4). Therefore, there is little sense for employing this strategy despite the fact that it bears the lowest
ACERICER. As shown in
Table 8, if Strategy 001 is replaced by Strategy 100, each additional unit of
will require a moderate cost of
units. In particular, if Strategy 100 is applied instead of Strategy 001, it yields
additional units of
at the additional cost of
units.
Note that Strategy 100 is purely biological and only requires a one-time investment to acquire the predacious species (
) and does not involve operational expenses for spraying of chemical substances. Nonetheless, this environmentally friendly strategy exhibits a noticeable effect on the suppression of mosquito population (see
Figure 7). Thus, it seems reasonable to qualify Strategy 100 as more cogent for the practical use than Strategy 001 (the cheapest option).










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}