Immobilization of Old Yellow Enzymes via Covalent or Coordination Bonds


 , ,
, ,  ,
,
Abstract
:1. Introduction
2. Results and Discussion
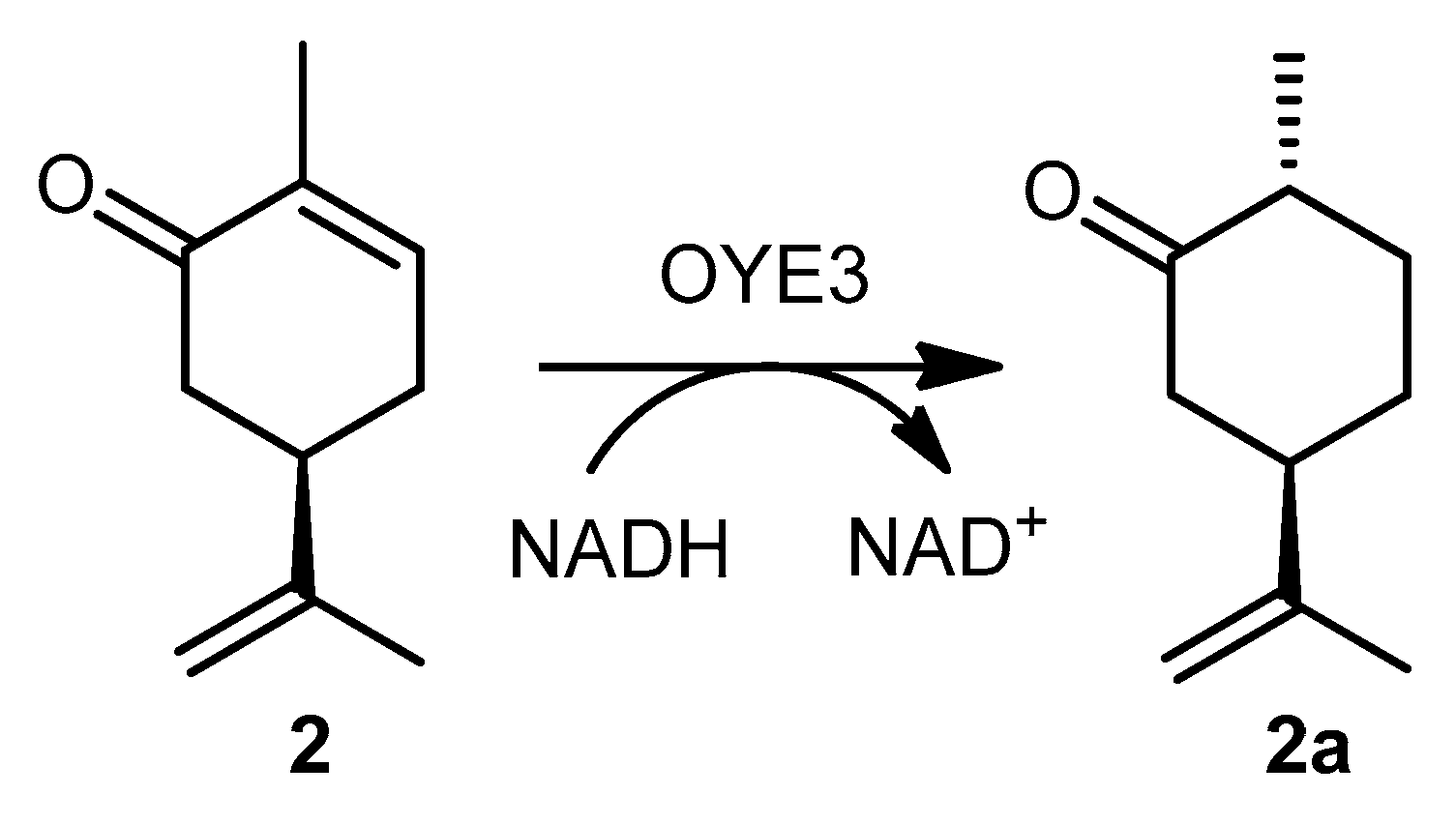
2.1. Enzyme Activity Assays
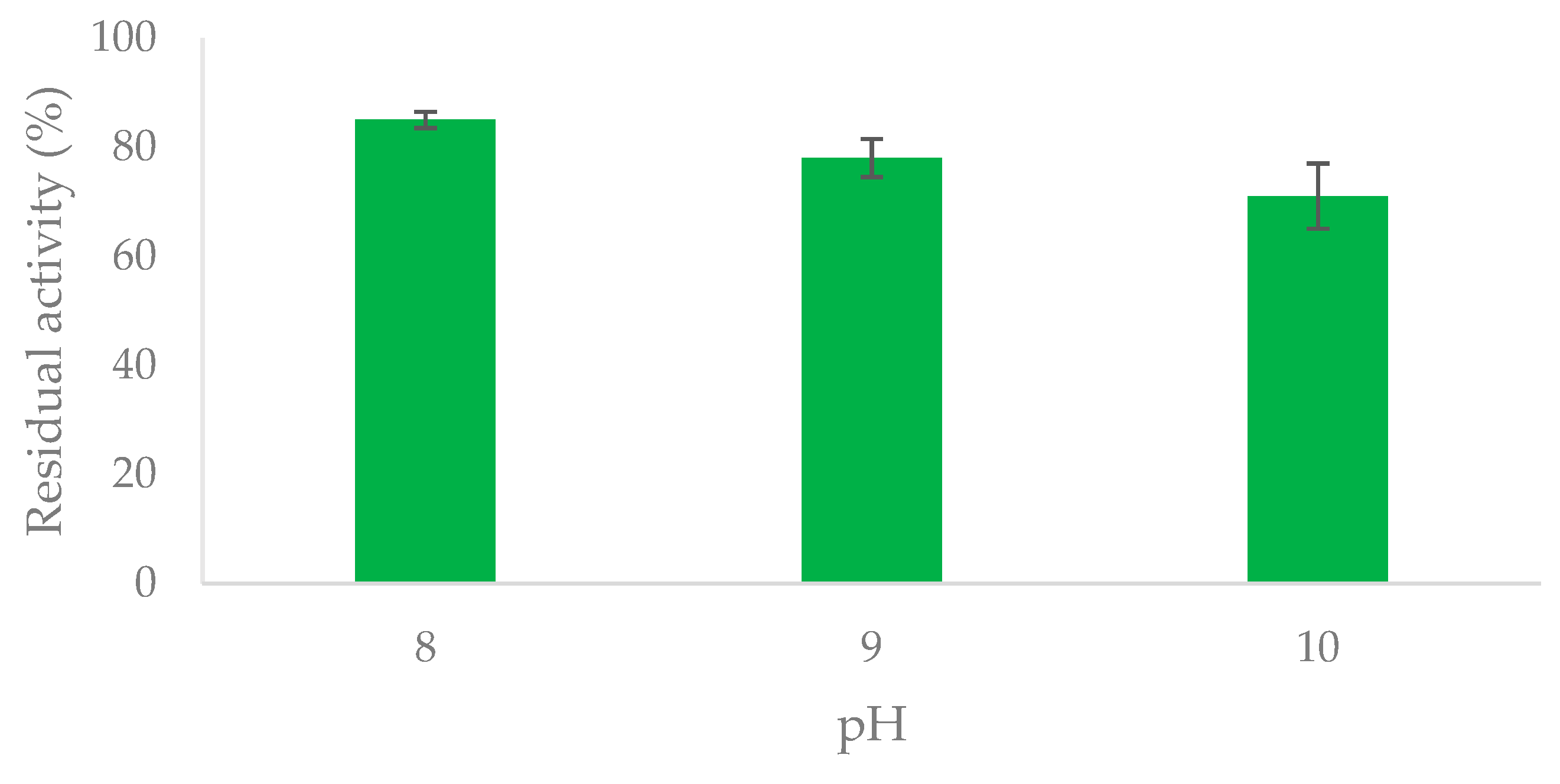
2.2. Preliminary Study of OYE3 Stability at Alkaline pH
2.3. Immobilization of OYE3
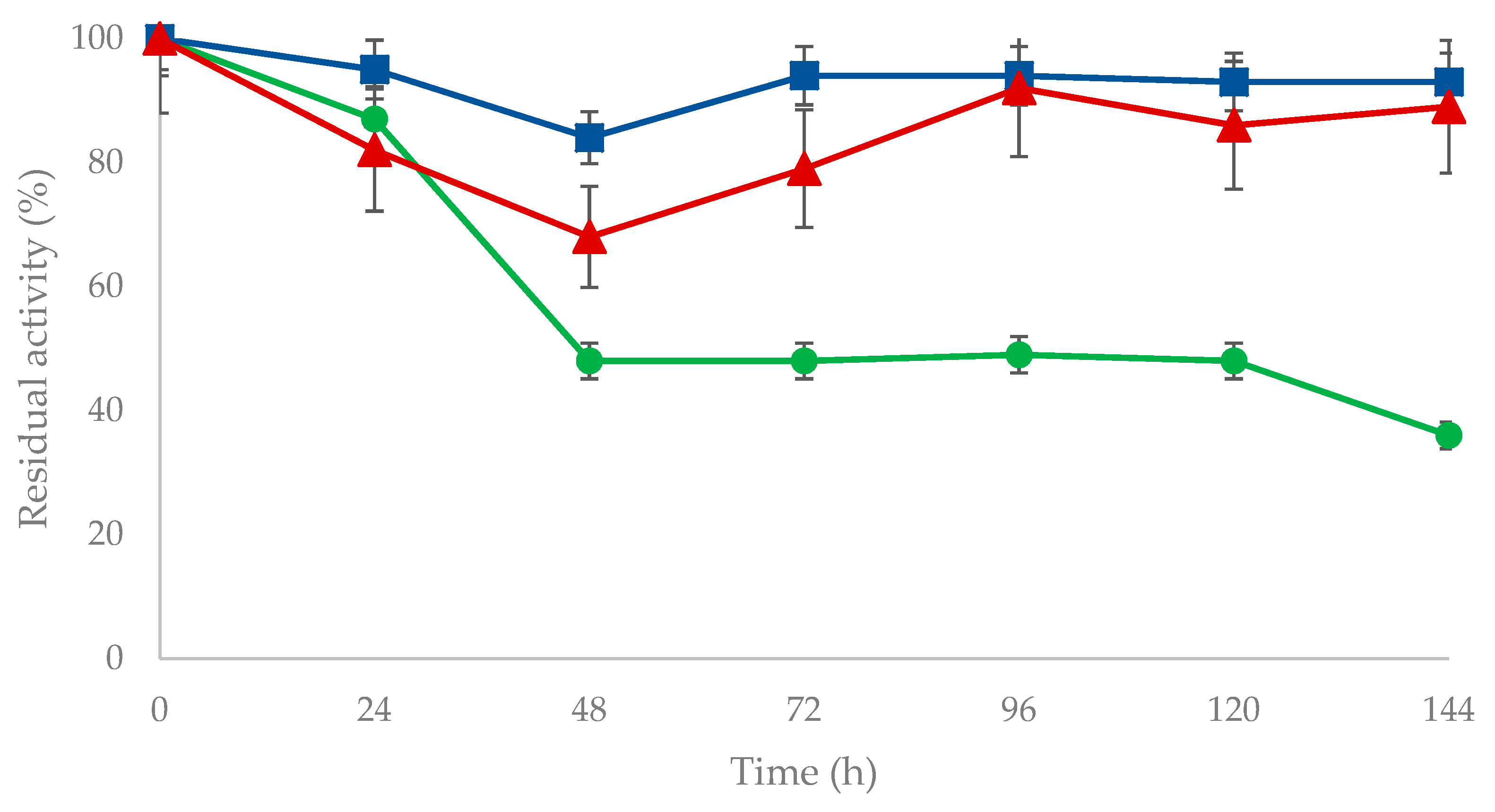
2.4. Stability of Immobilized OYE3
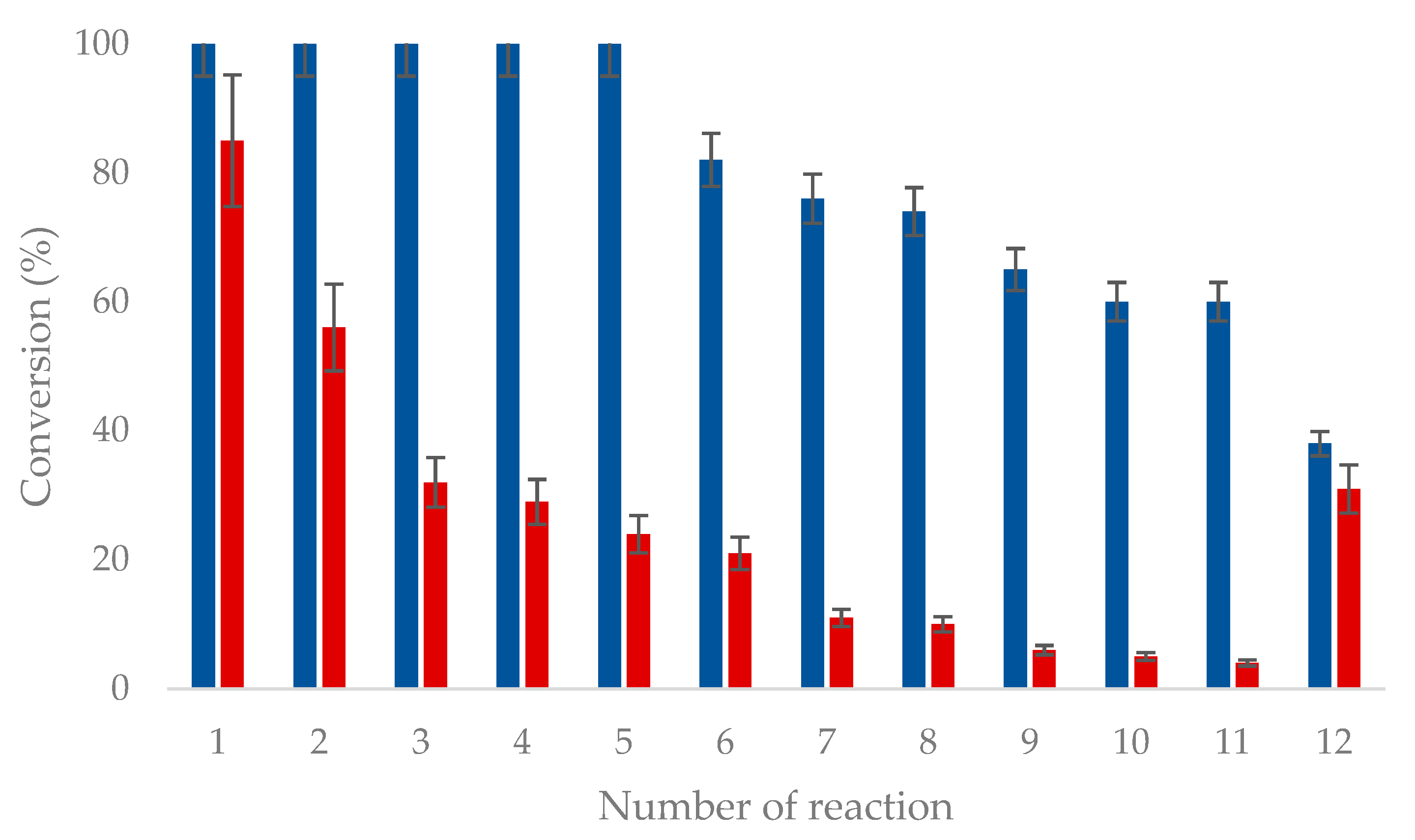
2.5. Recycling of Immobilized OYE3
2.6. Protein Leakage Assay of Immobilized OYE3
3. Materials and Methods
3.1. General
3.2. Enzyme Production
3.3. Activity Assays
3.3.1. Spectrophotometric Enzymatic Assay
3.3.2. GC-MS Enzymatic Assay
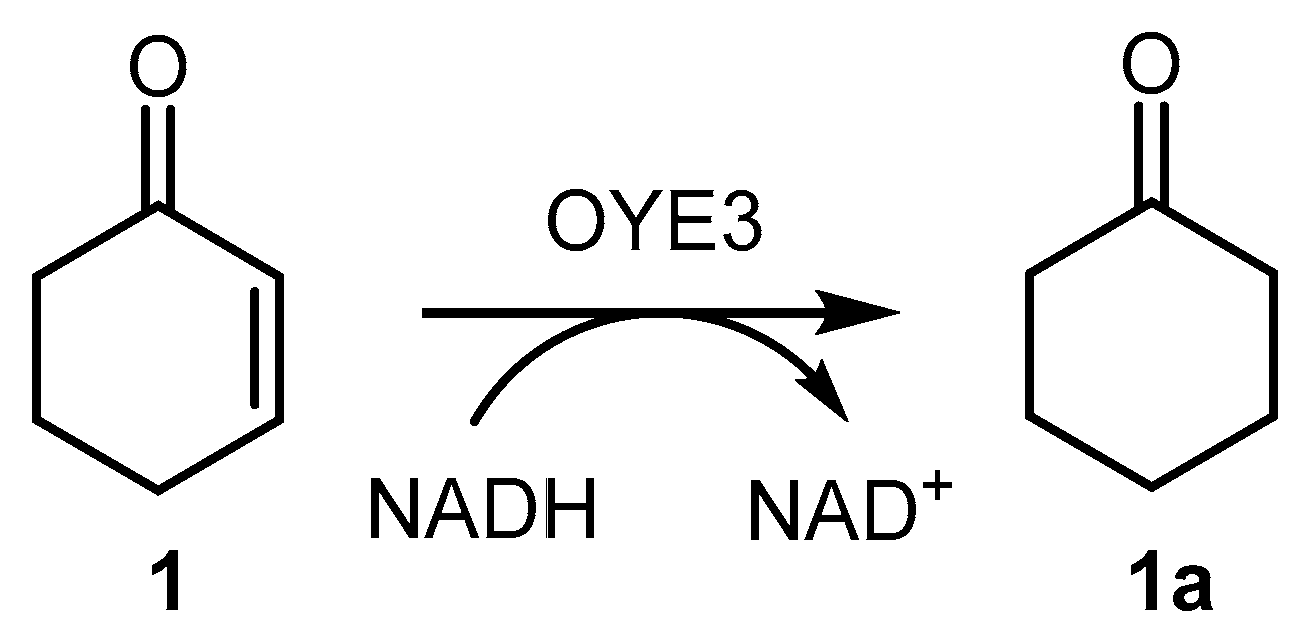
3.4. OYE3-Mediated Reduction of α-Methyl-Trans-Cinnamaldehyde (3)
3.5. Stability of OYE3 in the pH Range 8-10
3.6. Covalent Immobilization of OYE3 on Glyoxyl-Agarose (OYE3-GA)
3.7. Immobilization of OYE3 by Affinity-Based Adsorption (OYE3-EziG)
3.8. Stability of Non-Immobilized OYE3, OYE3-GA, and OYE3-EziG
3.9. Protein Leakage Assay
3.10. Recycling of Immobilized Enzymes
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Winkler, C.K.; Tasnádi, G.; Clay, D.; Hall, M.; Faber, K. Asymmetric bioreduction of activated alkenes to industrially relevant optically active compounds. J. Biotechnol. 2012, 162, 381–389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toogood, H.S.; Scrutton, N.S. New developments in ‘ene’-reductase catalysed biological hydrogenations. Curr. Opin. Chem. Biol. 2014, 19, 107–115. [Google Scholar] [CrossRef] [PubMed]
- Vaz, A.D.N.; Chakraborty, S.; Massey, V. Old yellow enzyme: Aromatization of cyclic enones and the mechanism of a novel dismutation reaction. Biochemistry 1995, 34, 4246–4256. [Google Scholar] [CrossRef] [PubMed]
- Gatti, F.G.; Parmeggiani, F.; Sacchetti, A. Synthetic strategies based on C=C bioreductions for the preparation of biologically active molecules. In Synthetic Methods for Biologically Active Molecules; Brenna, E., Ed.; WILEY-VCH Verlag GmbH & Co. KGaA: Weinheim, Germany, 2013; pp. 49–84. [Google Scholar]
- Toogood, H.S.; Scrutton, N.S. Discovery, characterization, engineering, and applications of ene-reductases for industrial biocatalysis. ACS Catal. 2018, 8, 3532–3549. [Google Scholar] [CrossRef] [PubMed]
- Winkler, C.K.; Faber, K.; Hall, M. Biocatalytic reduction of activated C=C-bonds and beyond: Emerging trends. Curr. Opin. Chem. Biol. 2018, 43, 97–105. [Google Scholar] [CrossRef] [PubMed]
- Classen, T.; Korpak, M.; Schölzel, M.; Pietruszka, J. Stereoselective enzyme cascades: An efficient synthesis of chiral γ-butyrolactones. ACS Catal. 2014, 4, 1321–1331. [Google Scholar] [CrossRef]
- Brenna, E.; Crotti, M.; Gatti, F.G.; Monti, D.; Parmeggiani, F.; Pugliese, A.; Tentori, F. Biocatalytic synthesis of chiral cyclic γ-oxoesters by sequential C–H hydroxylation, alcohol oxidation and alkene reduction. Green Chem. 2017, 19, 5122–5130. [Google Scholar] [CrossRef]
- Winkler, C.K.; Clay, D.; Davies, S.; O’Neill, P.; McDaid, P.; Debarge, S.; Steflik, J.; Karmilowicz, M.; Wong, J.W.; Faber, K. Chemoenzymatic asymmetric synthesis of pregabalin precursors via asymmetric bioreduction of β-cyanoacrylate esters using ene-reductases. J. Org. Chem. 2013, 78, 1525–1533. [Google Scholar] [CrossRef]
- Brenna, E.; Gatti, F.G.; Manfredi, A.; Monti, D.; Parmeggiani, F. Biocatalyzed enantioselective reduction of activated C=C bonds: Synthesis of enantiomerically enriched α-halo-β-arylpropionic acids. Eur. J. Org. Chem. 2011, 4015–4022. [Google Scholar] [CrossRef]
- Brenna, E.; Crotti, M.; Gatti, F.G.; Monti, D.; Parmeggiani, F.; Powell, R.W.; Santangelo, S.; Stewart, J.D. Opposite enantioselectivity in the bioreduction of (Z)-β-aryl-β-cyanoacrylates mediated by the tryptophan 116 mutants of old yellow enzyme 1: Synthetic approach to (R)- and (S)-β-aryl-γ-lactams. Adv. Synth. Catal. 2015, 357, 1849–1860. [Google Scholar] [CrossRef]
- Waller, J.; Toogood, H.S.; Karuppiah, V.; Rattray, N.J.W.; Mansell, D.J.; Leys, D.; Gardiner, J.M.; Fryszkowska, A.; Ahmed, S.T.; Bandichhor, R.; et al. Structural insights into the ene-reductase synthesis of profens. Org. Biomol. Chem. 2017, 15, 4440–4448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumru, C.; Classen, T.; Pietruszka, J. Enantioselective, catalytic one-pot synthesis of γ-butyrolactone-based fragrances. ChemCatChem 2018, 10, 4931–4940. [Google Scholar] [CrossRef]
- Guo, J.; Zhang, R.; Ouyang, J.; Zhang, F.; Qin, F.; Liu, G.; Zhang, W.; Li, H.; Ji, X.; Jia, X.; et al. Stereodivergent synthesis of carveol and dihydrocarveol through ketoreductasesene-reductases catalyzed asymmetric reduction. ChemCatChem 2018, 10, 5496–5504. [Google Scholar] [CrossRef]
- Stueckler, C.; Mueller, N.J.; Winkler, C.K.; Glueck, S.M.; Gruber, K.; Steinkellner, G.; Faber, K. Bioreduction of α-methylcinnamaldehyde derivatives: Chemo-enzymatic asymmetric synthesis of Lilial™ and Helional™. Dalton Trans. 2010, 39, 8472–8476. [Google Scholar] [CrossRef]
- Bryan, M.C.; Dunn, P.J.; Entwistle, D.; Gallou, F.; Koenig, S.G.; Hayler, J.D.; Hickey, M.R.; Hughes, S.; Kopach, M.E.; Moine, G.; et al. Key Green Chemistry research areas from a pharmaceutical manufacturers’ perspective revisited. Green Chem. 2018, 20, 5082–5103. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Xiao, W.; Xie, P.; Zheng, L. Co-immobilization of enoate reductase with a cofactor-recycling partner enzyme. Enzym. Microb. Technol. 2018, 109, 66–73. [Google Scholar] [CrossRef]
- Yoon, J.; Lee, S.H.; Tieves, F.; Rauch, M.; Hollmann, F.; Park, C.B. Light-harvesting dye−alginate hydrogel for solar-driven, sustainable biocatalysis of asymmetric hydrogenation. ACS Sustain. Chem. Eng. 2019, 7, 5632–5637. [Google Scholar] [CrossRef] [Green Version]
- Guisán, J.M. Aldehyde-agarose gels as activated supports for immobilization-stabilization of enzymes. Enzyme Microb. Technol. 1988, 10, 375–382. [Google Scholar] [CrossRef]
- Zucca, P.; Fernandez-Lafuente, R.; Sanjust, E. Agarose and its derivatives as supports for enzyme immobilization. Molecules 2016, 21, 1577. [Google Scholar] [CrossRef]
- Fernández-Lorente, G.; Lopez-Gallego, F.; Bolivar, J.M.; Rocha-Martin, J.; Moreno-Perez, S.; Guisán, J.M. Immobilization of proteins on highly activated glyoxyl supports: Dramatic increase of the enzyme stability via multipoint immobilization on pre-existing carriers. Curr. Org. Chem. 2015, 19, 1–13. [Google Scholar] [CrossRef]
- Dall’Oglio, F.; Contente, M.L.; Conti, P.; Molinari, F.; Monfredi, D.; Pinto, A.; Romano, D.; Ubiali, D.; Tamborini, L.; Serra, I. Flow-based stereoselective reduction of ketones using an immobilized ketoreductaseglucose dehydrogenase mixed bed system. Catal. Commun. 2017, 93, 29–32. [Google Scholar] [CrossRef]
- Cassimjee, K.E.; Kadow, M.; Wikmark, Y.; Svedendahl Humble, M.; Rothstein, M.L.; Rothstein, D.M.; Bäckvall, J.-E. A general protein purification and immobilization method on controlled porosity glass: Biocatalytic applications. Chem. Commun. 2014, 50, 9134–9137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thompson, M.P.; Derrington, S.R.; Heath, R.S.; Porter, J.L.; Mangas-Sanchez, J.; Devine, P.N.; Truppo, M.D.; Turner, N.J. A generic platform for the immobilisation of engineered biocatalysts. Tetrahedron 2019, 75, 327–334. [Google Scholar] [CrossRef]
- Chaparro-Riggers, J.F.; Rogers, T.A.; Vazquez-Figueroa, E.; Polizzi, K.M.; Bommarius, A.S. Comparison of three enoate reductases and their potential use for biotransformations. Adv. Synth. Catal. 2007, 309, 1521–1531. [Google Scholar] [CrossRef]
- Kohli, R.M.; Massey, V. The oxidative half-reaction of Old Yellow Enzyme. J. Biol. Chem. 1998, 273, 32763–32770. [Google Scholar] [CrossRef] [Green Version]
- Hulley, M.E.; Toogood, H.S.; Fryszkowska, A.; Mansell, D.; Stephens, G.M.; Gardiner, J.M.; Scrutton, N.S. Focused directed evolution of pentaerythritol tetranitrate reductase by using automated anaerobic kinetic screening of site-saturated libraries. ChemBioChem 2010, 11, 2433–2447. [Google Scholar] [CrossRef]
- Orrego, A.H.; Romero-Fernández, M.; Millán-Linares, M.C.; Yust, M.M.; Guisán, J.M.; Rocha-Martin, J. Stabilization of enzymes by multipoint covalent attachment on aldehyde-supports: 2-picoline borane as an alternative reducing agent. Catalysts 2018, 8, 333. [Google Scholar] [CrossRef] [Green Version]
- Sheldon, R.A.; van Pelt, S. Enzyme immobilisation in biocatalysis: Why, what and how. Chem. Soc. Rev. 2013, 42, 6223–6235. [Google Scholar] [CrossRef] [Green Version]
- EnginZyme: Enzyme Carrier, Biocatalysis Research. Available online: http://enginzyme.com/ (accessed on 30 January 2020).
- Mateo, C.; Palomo, J.M.; Fernandez-Lorente, G.; Guisán, J.M.; Fernandez-Lafuente, R. Improvement of enzyme activity, stability and selectivity via immobilization techniques. Enzym. Microb. Technol. 2007, 40, 1451–1463. [Google Scholar] [CrossRef]
- Foley, A.M.; Maguire, A.R. The impact of recent developments in technologies which enable the increased use of biocatalysts. Eur. J. Org. Chem. 2019, 3713–3734. [Google Scholar] [CrossRef]
- Sheldon, R.A.; Brady, D. The limits to biocatalysis: Pushing the envelope. Chem. Commun. 2018, 54, 6088–6104. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 27, 248–254. [Google Scholar] [CrossRef]
- Chevallet, M.; Luche, S.; Rabilloud, T. Silver staining of proteins in polyacrylamide gels. Nat. Protoc. 2006, 1, 1852–1858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brenna, E.; Gatti, F.G.; Monti, D.; Parmeggiani, F.; Sacchetti, A. Cascade coupling of ene reductases with alcohol dehydrogenases: Enantioselective reduction of prochiral unsaturated aldehydes. ChemCatChem 2012, 4, 653–659. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Carrier | Enzyme Loading (mg g−1) | Immobilization Yield 1 (%) | Activity 2 (U g−1) | Activity Recovery 3 (%) |
|---|---|---|---|---|
| Glyoxyl-agarose | 2.0 | 95 | 0.023 | 52 |
| EziGTM Opal | 4.2 | 100 | 0.037 | 54 |
| EziGTM Coral | 4.2 | 100 | 0.015 | 23 |
| EziGTM Amber | 4.2 | 100 | 0.014 | 19 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tentori, F.; Bavaro, T.; Brenna, E.; Colombo, D.; Monti, D.; Semproli, R.; Ubiali, D. Immobilization of Old Yellow Enzymes via Covalent or Coordination Bonds. Catalysts 2020, 10, 260. https://doi.org/10.3390/catal10020260
Tentori F, Bavaro T, Brenna E, Colombo D, Monti D, Semproli R, Ubiali D. Immobilization of Old Yellow Enzymes via Covalent or Coordination Bonds. Catalysts. 2020; 10(2):260. https://doi.org/10.3390/catal10020260
Chicago/Turabian StyleTentori, Francesca, Teodora Bavaro, Elisabetta Brenna, Danilo Colombo, Daniela Monti, Riccardo Semproli, and Daniela Ubiali. 2020. "Immobilization of Old Yellow Enzymes via Covalent or Coordination Bonds" Catalysts 10, no. 2: 260. https://doi.org/10.3390/catal10020260
APA StyleTentori, F., Bavaro, T., Brenna, E., Colombo, D., Monti, D., Semproli, R., & Ubiali, D. (2020). Immobilization of Old Yellow Enzymes via Covalent or Coordination Bonds. Catalysts, 10(2), 260. https://doi.org/10.3390/catal10020260