
Enhancing Soluble Expression of Phospholipase B for Efficient Catalytic Synthesis of L-Alpha-Glycerylphosphorylcholine



Abstract
:1. Introduction
2. Results and Discussion
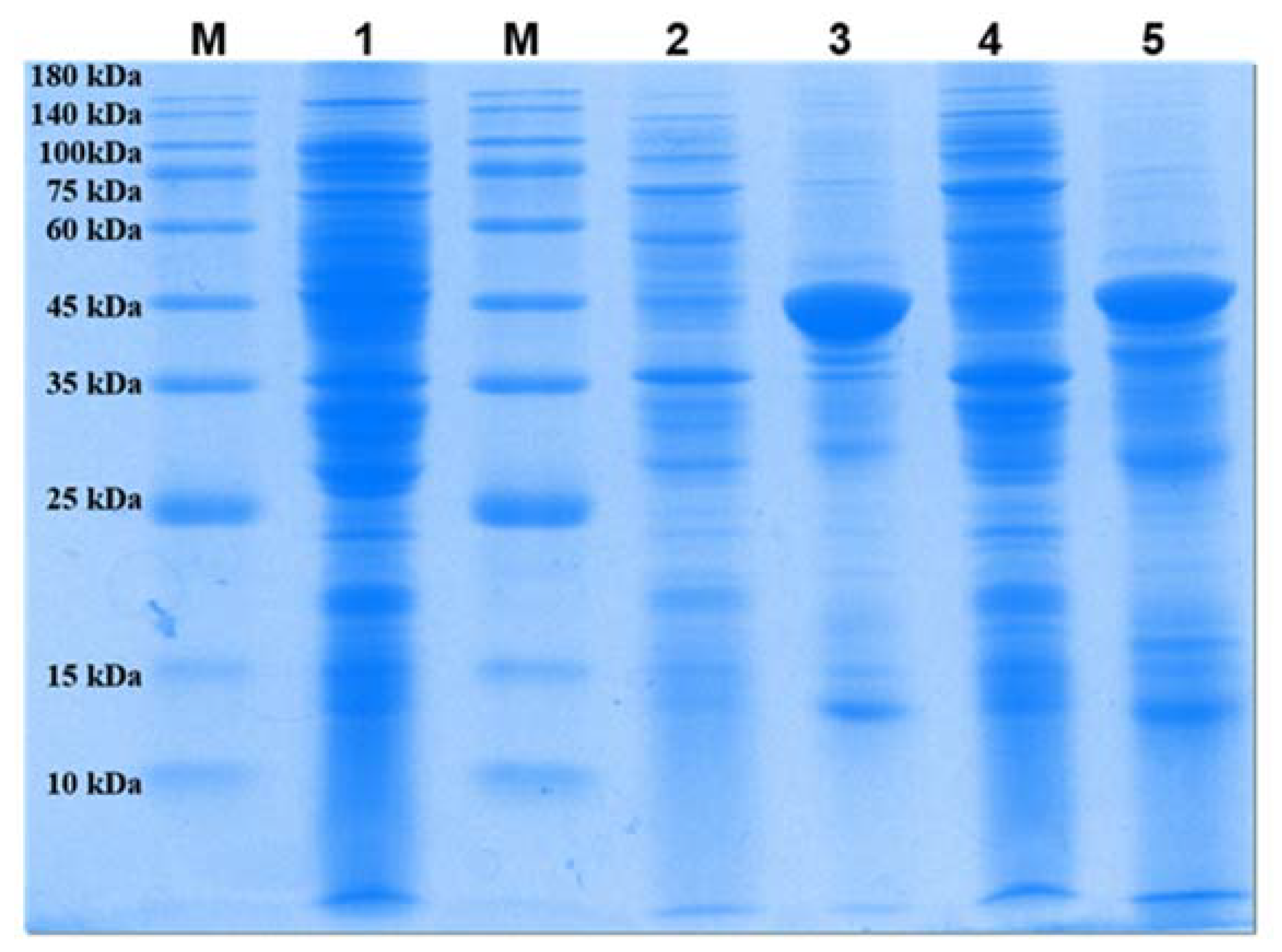
2.1. Expression of Recombinant PLB in E. coli BL21(DE3)
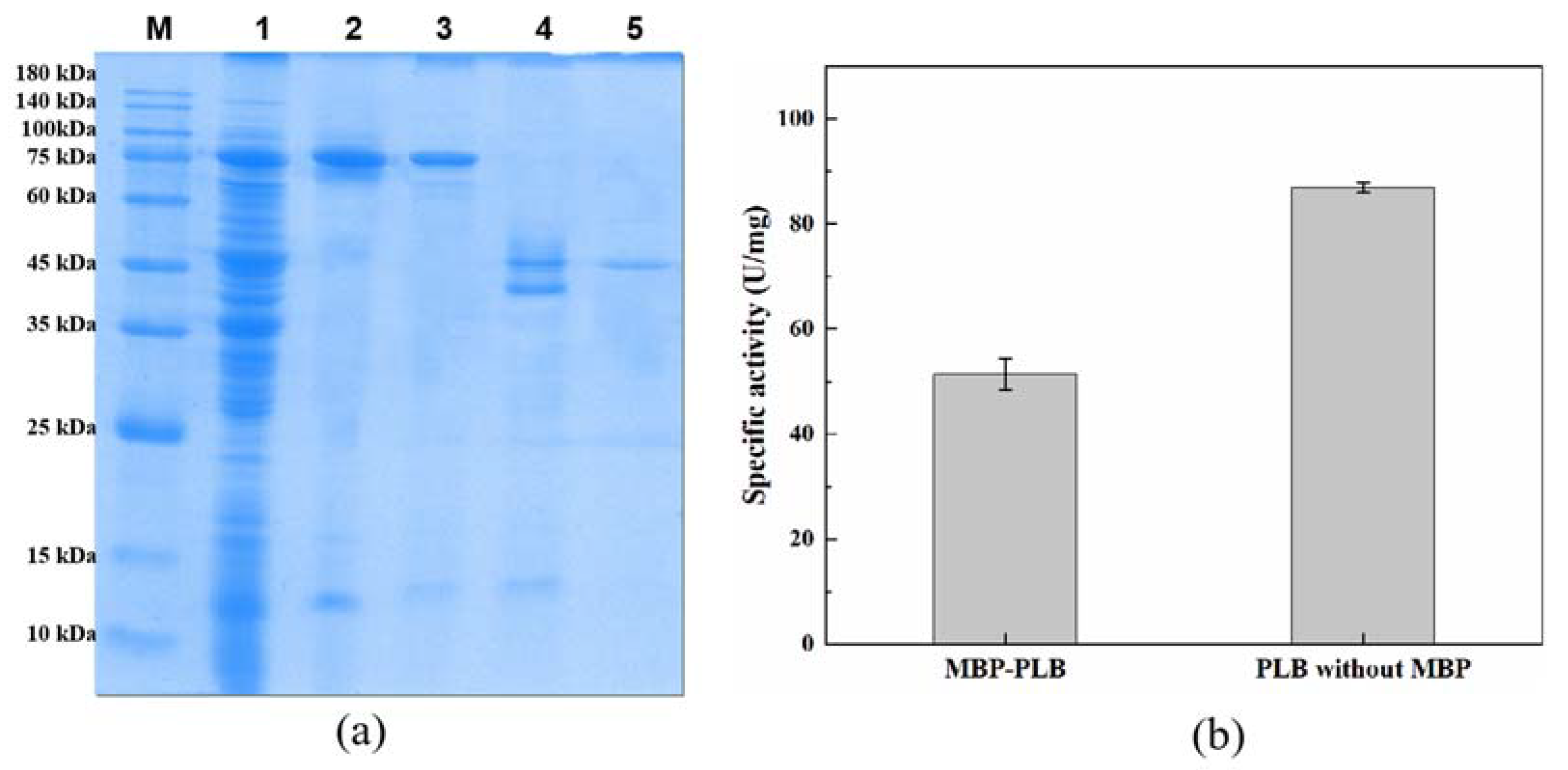
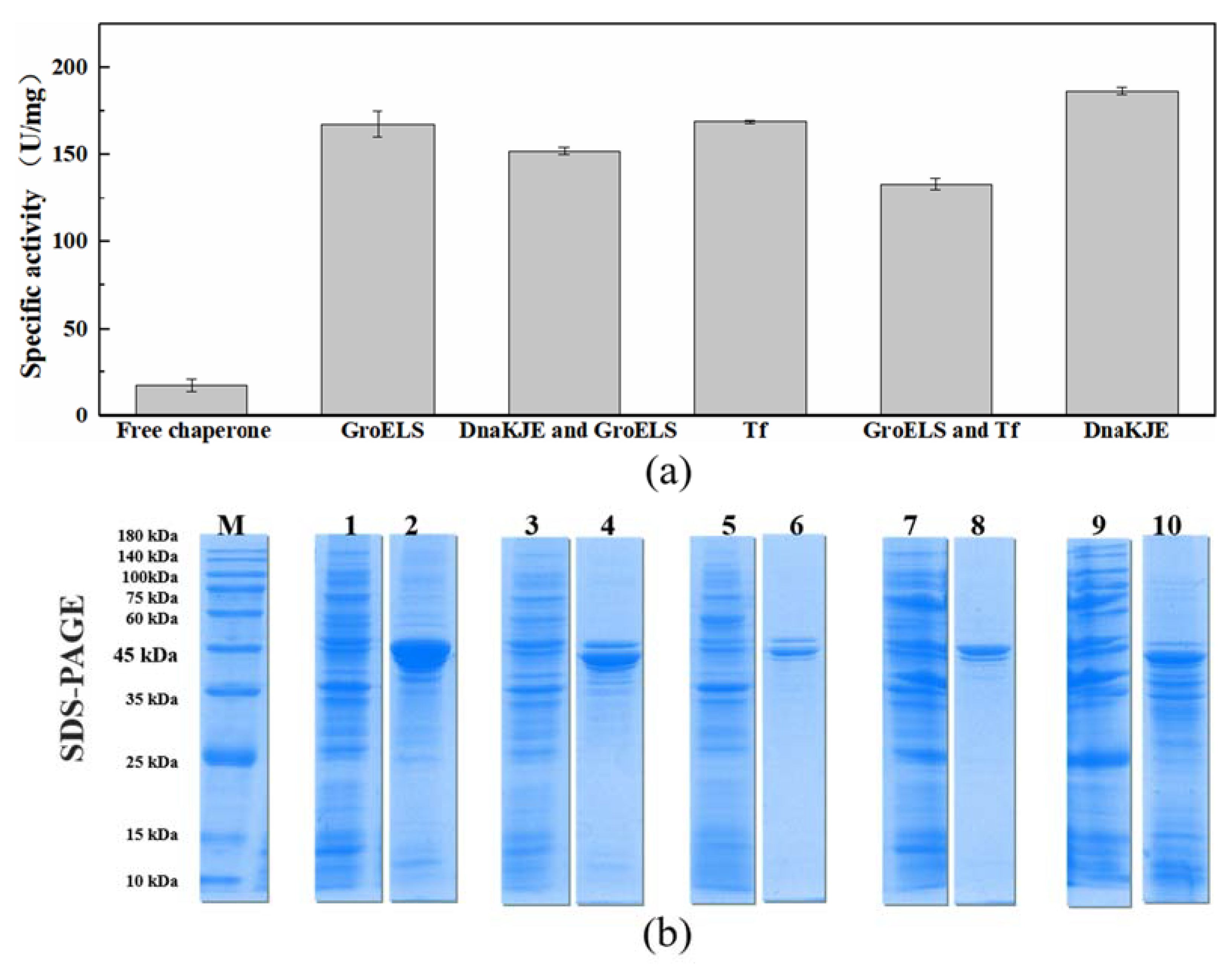
2.2. Promoting the Soluble Expression of PLB with the Assistance of Molecular Chaperones
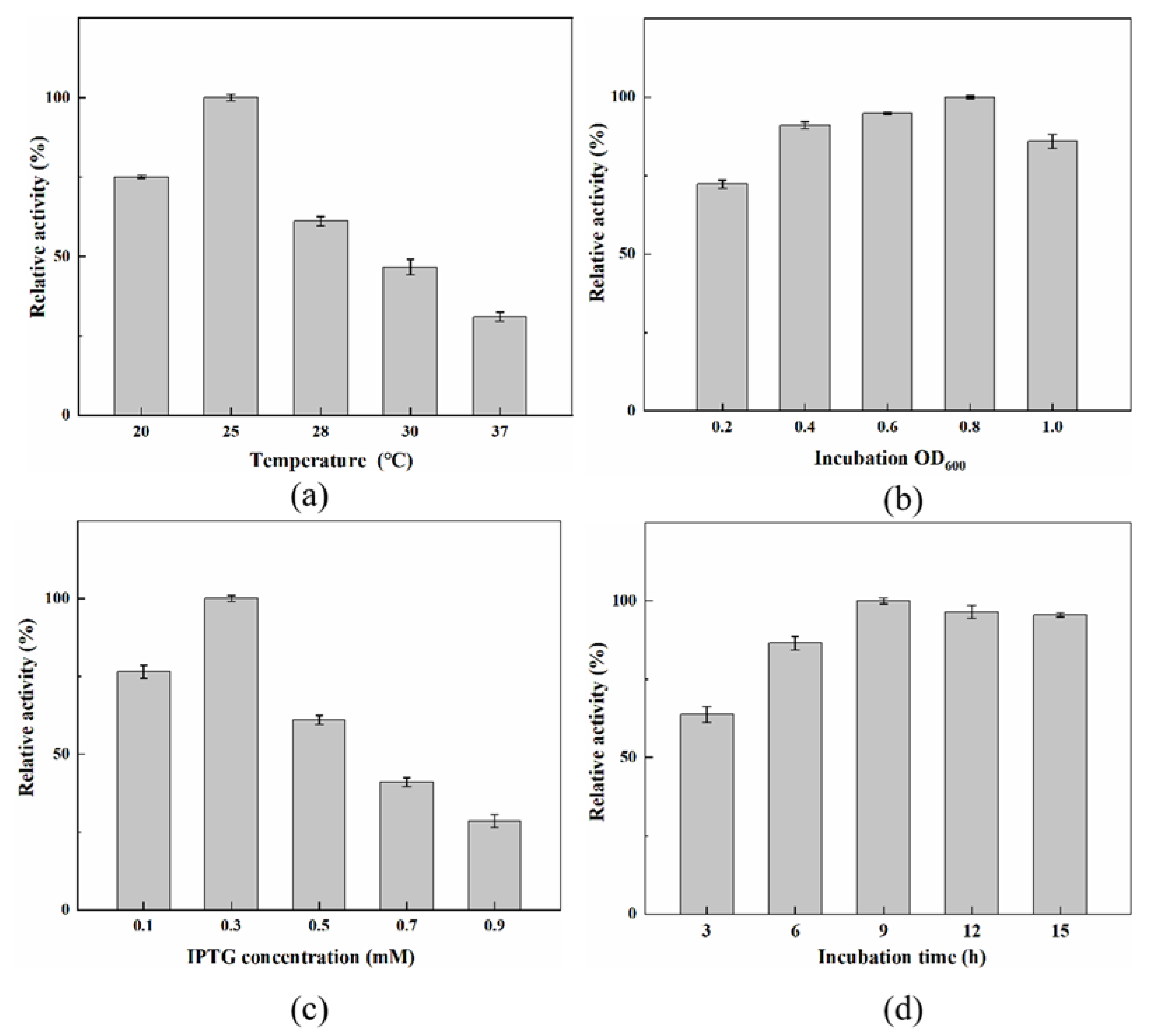
2.3. Optimizing the Expression Conditions for High-Level Soluble Production of PLB
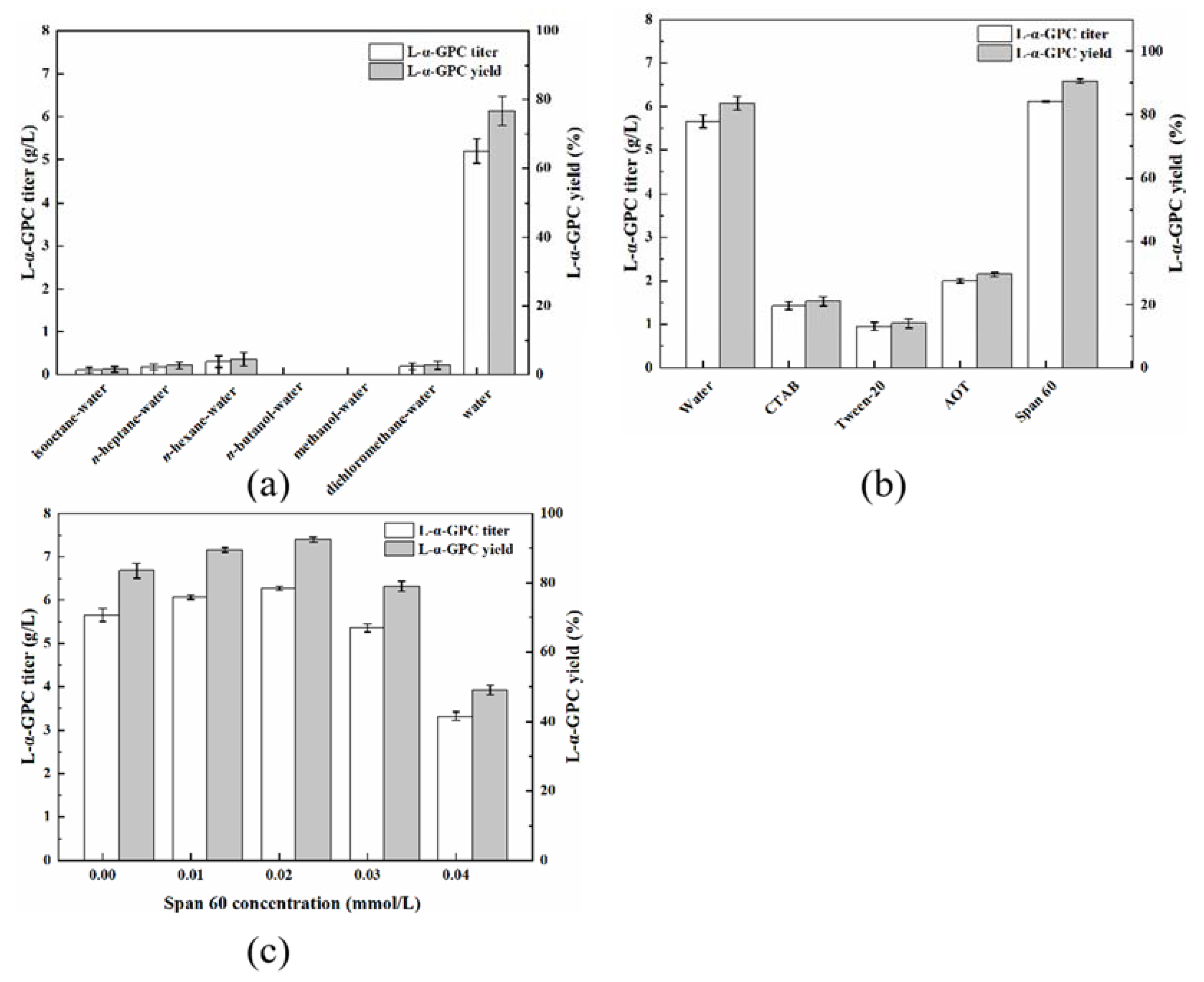
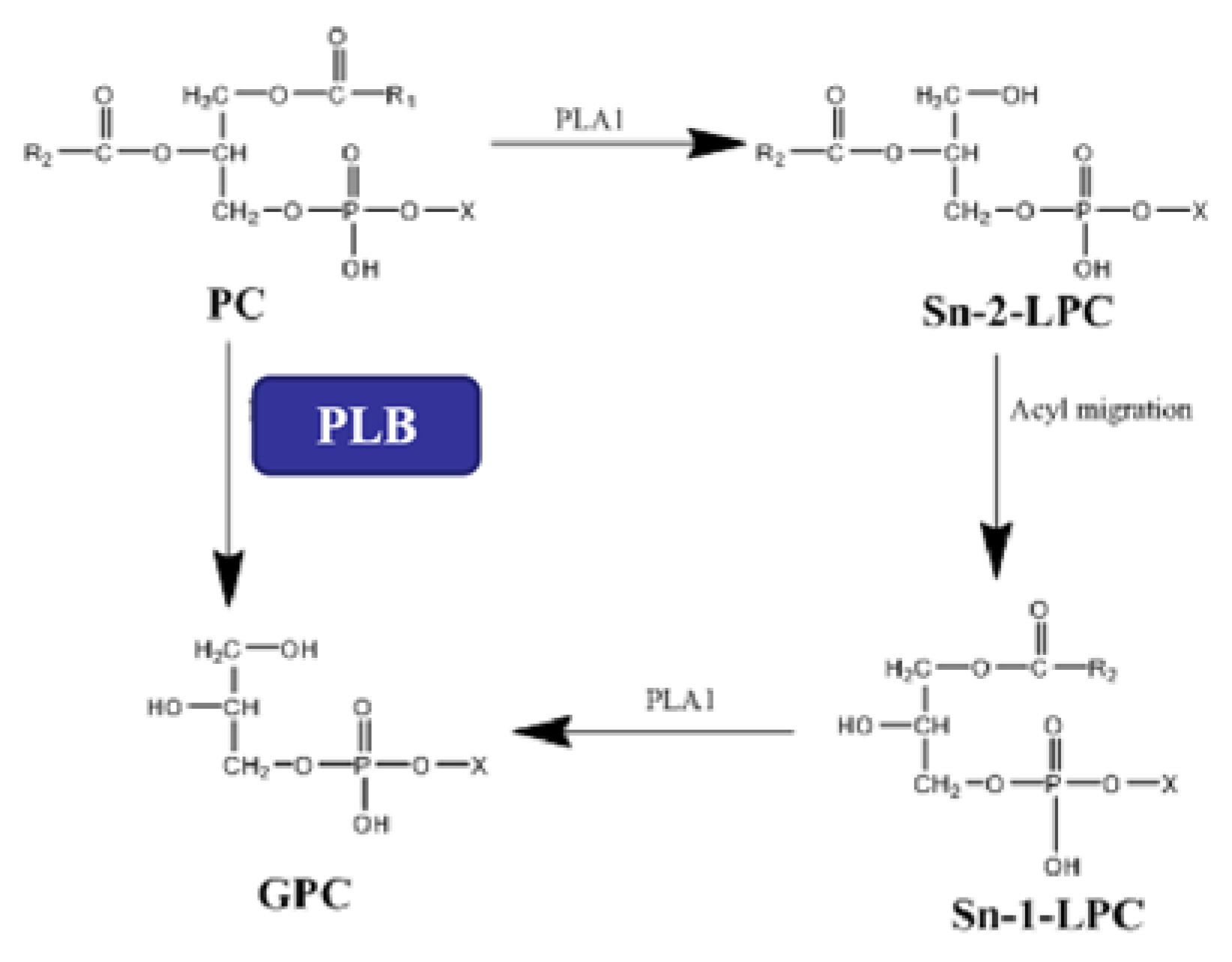
2.4. Construction of a PLB-Catalyzed Hydrolysis System for L-α-GPC Synthesis
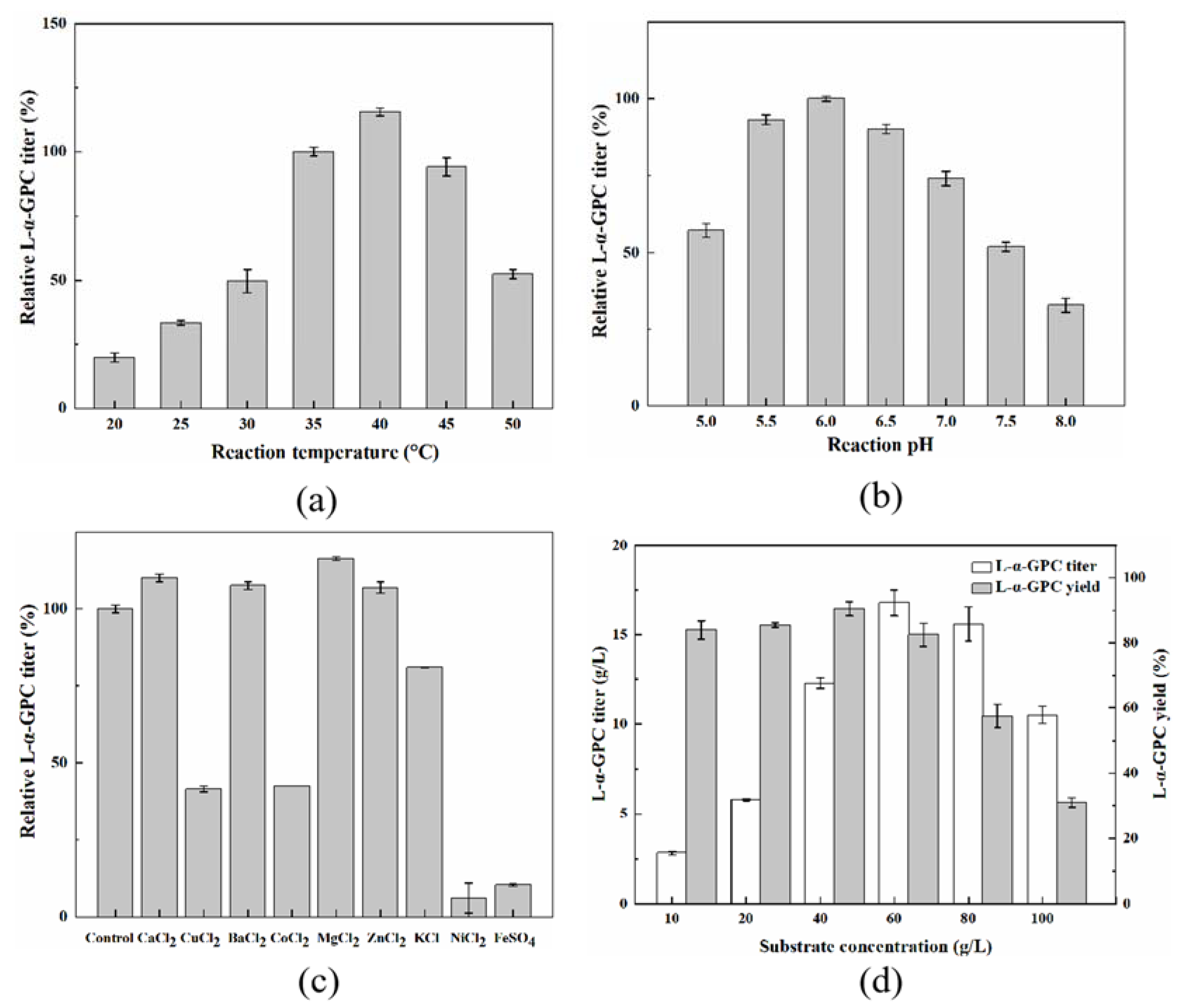
2.5. Effects of Reaction Conditions on L-α-GPC Synthesis in the PLB-Catalyzed Hydrolysis System
3. Materials and Methods
3.1. Bacterial Strains and Culture Conditions
3.2. Sodium Dodecyl Sulfate-Polyacrylamide Gel Electrophoresis (SDS-PAGE) Analysis and Purification of PLB
3.3. Enzymatic Assays
3.4. Production of L-α-GPC from PC Using PLB as the Catalyst
3.5. HPLC Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Du, Y.W.; He, W.; Zhou, W.Y.; Li, X.S. Disulfide phosphatidylcholines: Alternative phospholipids for the preparation of functional liposomes. Chem. Commun. 2019, 55, 8434–8437. [Google Scholar] [CrossRef] [PubMed]
- Jia, L.G.; Wang, C.; Wu, Y.F.; Ma, X.Y.; Zhang, X.; Chu, X.X.; Lu, F.P.; Liu, Y.H. Production of L-alpha-glycerylphosphorylcholine from oil refining waste using a novel cold-active phospholipase B from Bacillus velezensis. ACS Sustain. Chem. Eng. 2021, 9, 13337–13346. [Google Scholar] [CrossRef]
- Lee, S.H.; Choi, B.Y.; Kim, J.H.; Kho, A.R.; Sohn, M.; Song, H.K.; Choi, H.C.; Suh, S.W. Late treatment with choline alfoscerate (L-alpha glycerylphosphorylcholine, alpha-GPC) increases hippocampal neurogenesis and provides protection against seizure-induced neuronal death and cognitive impairment. Brain. Res. 2017, 1654, 66–76. [Google Scholar] [CrossRef] [PubMed]
- Tayebati, S.K. Phospholipid and lipid derivatives as potential neuroprotective compounds. Molecules 2018, 23, 2257. [Google Scholar] [CrossRef] [Green Version]
- Park, J.M.; De Castro, K.A.; Ahn, H.; Rhee, H. Facile syntheses of L-alpha-glycerophosphorylcholine. B. Korean. Chem. Soc. 2010, 31, 2689–2691. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.H.; Li, M.J.; Huang, L.; Gui, S.; Jia, L.B.; Zheng, D.; Fu, Y.; Zhang, Y.T.; Rui, J.Q.; Lu, F.P. Cloning, expression and characterisation of phospholipase B from Saccharomyces cerevisiae and its application in the synthesis of L-alpha-glycerylphosphorylcholine and peanut oil degumming. Biotechnol. Biotec. Equip. 2018, 32, 968–973. [Google Scholar] [CrossRef] [Green Version]
- Lu, Y.Y.; Zhang, A.L.; Wang, X.; Hao, N.; Chen, K.Q.; Ouyang, P.K. Surfactant enhanced l-alpha-glycerylphosphorylcholine production from phosphatidylcholine using phospholipase A(1) in the aqueous phase. Biocatal. Biotransfor. 2019, 37, 361–366. [Google Scholar] [CrossRef]
- Másson, M.; Loftsson, T.; Haraldsson, G. Marine lipids for prodrugs, soft compounds and other pharmaceutical applications. Pharmazie 2000, 3, 172–177. [Google Scholar]
- Zhang, K.Y.; Liu, Y.F.; Wang, X.G. Enzymatic preparation of L-alpha-glycerylphosphorylcholine in an aqueous medium. Eur. J. Lipid. Sci. Tech. 2012, 114, 1254–1260. [Google Scholar] [CrossRef]
- Lim, C.W.; Kim, B.H.; Kim, I.H.; Lee, M.W. Modeling and optimization of phospholipase A(1)-catalyzed hydrolysis of phosphatidylcholine using response surface methodology for lysophosphatidylcholine production. Biotechnol. Prog. 2015, 31, 35–41. [Google Scholar] [CrossRef]
- Li, H.; Cao, X.; Lu, Y.Y.; Ni, Y.; Wang, X.; Lu, Q.H.; Li, G.L.; Chen, K.Q.; Ouyang, P.K.; Tan, W.M. Alkaline modification of a metal-enzyme-surfactant nanocomposite to enhance the production of L-alpha-glycerylphosphorylcholine. Catalysts 2019, 9, 237. [Google Scholar] [CrossRef] [Green Version]
- Jiang, F.Y.; Huang, S.; Imadad, K.; Li, C. Cloning and expression of a gene with phospholipase B activity from Pseudomonas fluorescens in Escherichia coli. Bioresour. Technol. 2012, 104, 518–522. [Google Scholar] [CrossRef] [PubMed]
- Jiang, F.Y.; Wang, J.M.; Ju, L.C.; Kaleem, I.; Dai, D.Z.; Li, C. Optimization of degumming process for soybean oil by phospholipase B. J. Chem. Technol. Biot. 2011, 86, 1081–1087. [Google Scholar] [CrossRef]
- Huang, S.; Liang, M.L.; Xu, Y.H.; Rasool, A.; Li, C. Characteristics and vegetable oils degumming of recombinant phospholipase B. Chem. Eng. J. 2014, 237, 23–28. [Google Scholar] [CrossRef]
- Ullah, A.; Masood, R. The sequence and three-dimensional structure characterization of snake venom phospholipases B. Front. Mol. Biosci. 2020, 7, 175. [Google Scholar] [CrossRef]
- Kohler, G.A.; Brenot, A.; Haas-Stapleton, E.; Agabian, N.; Deva, R.; Nigam, S. Phospholipase A(2) and phospholipase B activities in fungi. BBA-Mol. Cell Biol. Lipids 2006, 1761, 1391–1399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, S.Y.; Zhao, L.S.; Larsson, A.; Venge, P. The identification of a phospholipase B precursor in human neutrophils. FEBS J. 2009, 276, 175–186. [Google Scholar] [CrossRef]
- Cai, Z.Z.; Wang, H.F.; Li, W.Z.; Lee, W.J.; Li, W.; Wang, Y.; Wang, Y. Preparation of L-alpha-glyceryl phosphorylcholine by hydrolysis of soy lecithin using phospholipase A1 in a novel solvent-free water in oil system. LWT-Food. Sci. Technol. 2020, 129, 109562. [Google Scholar] [CrossRef]
- Bang, H.J.; Kim, I.H.; Kim, B.H. Phospholipase A(1)-catalyzed hydrolysis of soy phosphatidylcholine to prepare L-alpha-glycerylphosphorylcholine in organic-aqueous media. Food. Chem. 2016, 190, 201–206. [Google Scholar] [CrossRef]
- San-Miguel, T.; Perez-Bermudez, P.; Gavidia, I. Production of soluble eukaryotic recombinant proteins in E-coli is favoured in early log-phase cultures induced at low temperature. Springerplus 2013, 2, 89. [Google Scholar] [CrossRef] [Green Version]
- García-Fruitós, E. Insoluble Proteins; Humana Press: New York, NY, USA, 2015; pp. 1–24. [Google Scholar]
- Fox, J.D.; Kapust, R.B.; Waugh, D.S. Single amino acid substitutions on the surface of Escherichia coli maltose-binding protein can have a profound impact on the solubility of fusion proteins. Protein Sci. 2001, 10, 622–630. [Google Scholar] [CrossRef] [Green Version]
- Raran-Kurussi, S.; Keefe, K.; Waugh, D.S. Positional effects of fusion partners on the yield and solubility of MBP fusion proteins. Protein Expres. Purif. 2015, 110, 159–164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raran-Kurussi, S.; Waugh, D.S. The ability to enhance the solubility of its fusion partners is an intrinsic property of maltose-binding protein but their folding is either spontaneous or chaperone-mediated. PLoS ONE 2012, 7, e49589. [Google Scholar] [CrossRef] [Green Version]
- Yoshimune, K.; Ninomiya, Y.; Wakayama, M.; Moriguchi, M. Molecular chaperones facilitate the soluble expression of N-acyl-D-amino acid amidohydrolases in Escherichia coli. J. Ind. Microbiol. Biotechnol. 2004, 31, 421–426. [Google Scholar] [CrossRef] [PubMed]
- Cui, S.S.; Lin, X.Z.; Shen, J.H. Effects of co-expression of molecular chaperones on heterologous soluble expression of the cold-active lipase Lip-948. Protein Expres. Purif. 2011, 77, 166–172. [Google Scholar]
- Farajnia, S.; Ghorbanzadeh, V.; Dariushnejad, H. Effect of molecular chaperone on the soluble expression of recombinant fab fragment in E. coli. Int. J. Pept. Res. Ther. 2020, 26, 251–258. [Google Scholar] [CrossRef]
- Falak, S.; Sajed, M.; Rashid, N. Strategies to enhance soluble production of heterologous proteins in Escherichia coli. Biologia 2022, 77, 893–905. [Google Scholar] [CrossRef]
- Vera, A.; Gonzalez-Montalban, N.; Aris, A.; Villaverde, A. The conformational quality of insoluble recombinant proteins is enhanced at low growth temperatures. Biotechnol. Bioeng. 2007, 96, 1101–1106. [Google Scholar] [CrossRef]
- Durrani, R.; Khan, F.I.; Ali, S.; Wang, Y.H.; Yang, B. A thermolabile phospholipase B from talaromyces marneffei GD-0079: Biochemical characterization and structure dynamics study. Biomolecules 2020, 10, 231. [Google Scholar] [CrossRef] [Green Version]
- Wei, T.; Xu, C.P.; Yu, X.; Jia, W.W.; Yang, K.P.; Jia, C.X.; Mao, D.B. Characterization of a novel thermophilic phospholipase B from Thermotoga lettingae TMO: Applicability in enzymatic degumming of vegetable oils. J. Ind. Microbiol. Biotechnol. 2015, 42, 515–522. [Google Scholar] [CrossRef]
- Uttatree, S.; Winayanuwattikun, P.; Charoenpanich, J. Isolation and Characterization of a Novel Thermophilic-Organic Solvent Stable Lipase from Acinetobacter baylyi. Appl. Biochem. Biotechnol. 2010, 162, 1362–1376. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Jiang, F.; Li, C.; Dai, D. Interfacial catalytic mechanism of Pseudomonas fluorescens phospholipase B. CIESC J. 2020, 71, 4255–4259. [Google Scholar]
- Wang, Z.L. Bioavailability of organic compounds solubilized in nonionic surfactant micelles. Appl. Microbiol. Biotechnol. 2011, 89, 523–534. [Google Scholar] [CrossRef] [PubMed]
- Hegde, R.R.; Kumar, S.; Aswal, V.K.; Verma, A.; Bhattacharya, S.S.; Ghosh, A. Small-angle neutron scattering study of nonionic surfactant micelles and phase transitions in w/o microemulsion. J. Dispers. Sci. Technol. 2014, 35, 783–788. [Google Scholar] [CrossRef]
- Azam, S.; Kiana, P.; Abolfazl, S. How the CMC adjust the liquid mixture density and viscosity of non-ionic surfactants at various temperatures? J. Mol. Liq. 2022, 347, 117971. [Google Scholar]
- Gacek, M.M.; Berg, J.C. Effect of surfactant hydrophile-lipophile balance (HLB) value on mineral oxide charging in apolar media. J. Colloid. Interf. Sci. 2015, 449, 192–197. [Google Scholar] [CrossRef]
- Chin, M.; Somasundaran, P. Enzyme activity and structural dynamics linked to micelle fromation: A fluorescence anisotropy and ESR study. Photochem. Photobiol. 2014, 90, 455–462. [Google Scholar] [CrossRef]
- Magalhaes, S.S.; Alves, L.; Sebastiao, M.; Medronho, B.; Almeida, Z.L.; Faria, T.Q.; Brito, R.M.M.; Moreno, M.J.; Antunes, F.E. Effect of ethyleneoxide groups of anionic surfactants on lipase activity. Biotechnol. Progr. 2016, 32, 1276–1282. [Google Scholar] [CrossRef]
- Fujino, S.; Akiyama, D.; Akaboshi, S.; Fujita, T.; Watanabe, Y.; Tamai, Y. Purification and characterization of phospholipase B from Candida utilis. Biosci. Biotech. Bioch. 2006, 70, 377–386. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.C.A.; Wright, L.C.; Golding, J.C.; Sorrell, T.C. Purification and characterization of secretory phospholipase B, lysophospholipase and lysophospholipase/transacylase from a virulent strain of the pathogenic fungus Cryptococcus neoformans. Biochem. J. 2000, 347, 431–439. [Google Scholar] [CrossRef]
- Mohamed, A.H.K.A.; Ayobe, M.H. Studies of phospholipase A and B activities of egyptian snake venoms and a scorpion toxin. Toxicon 1969, 6, 293–298. [Google Scholar] [CrossRef]
- Matsumoto, Y.; Mineta, S.; Murayama, K.; Sugimori, D. A novel phospholipase B from Streptomyces sp. NA684—Purifification, characterization, gene cloning, extracellular production and prediction of the catalytic residues. FEBS J. 2013, 280, 3780–3796. [Google Scholar] [CrossRef] [PubMed]
- Altenbach, C.; Seelig, J. Calcium binding to phosphatidylcholine bilayers as studied by deuterium magnetic resonance. Evidence for the formation of a calcium complex with two phospholipid molecules. Biochemistry 1984, 23, 3913–3920. [Google Scholar] [CrossRef]
- Binder, H.; Arnold, K.; Ulrich, A.S.; Zschornig, O. Interaction of Zn2+ with phospholipid membranes. Biophys. Chem. 2001, 90, 57–74. [Google Scholar] [CrossRef]
- Jiang, F.Y.; Wang, J.M.; Kaleem, I.; Dai, D.Z.; Zhou, X.H.; Li, C. Degumming of vegetable oils by a novel phospholipase B from Pseudomonas fluorescens BIT-18. Bioresour. Technol. 2011, 102, 8052–8056. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains or Plasmids | Description | Source |
|---|---|---|
| Strains | ||
| E. coli BL21(DE3) | Used as host strain | Invitrogen |
| E. coli BL21(DE3)/pET28a-PLB | E. coli BL21(DE3) harboring plasmid pET 28a-PLB | This work |
| E. coli BL21(DE3)/pMal-c5x-PLB | E. coli BL21(DE3) harboring plasmid pMal-c5x-PLB | This work |
| E. coli BL21(DE3)/pET28a-PLB-pGro7 | E. coli BL21(DE3) harboring plasmids pET 28a-PLB and pGro7 | This work |
| E. coli BL21(DE3)/pET28a-PLB-pG-KJE8 | E. coli BL21(DE3) harboring plasmids pET 28a-PLB and pG-KJE8 | This work |
| E. coli BL21(DE3)/pET28a-PLB-pTf16 | E. coli BL21(DE3) harboring plasmids pET 28a-PLB and pTf16 | This work |
| E. coli BL21(DE3)/pET28a-PLB-pG-Tf2 | E. coli BL21(DE3) harboring plasmids pET 28a-PLB and pG-Tf2 | This work |
| E. coli BL21(DE3)/pET28a-PLB-pKJE7 | E. coli BL21(DE3) harboring plasmids pET 28a-PLB and pKJE7 | This work |
| Plasmids | ||
| pET 28a | Expression vectors, KmR, PT7, f1 ori | Invitrogen |
| pET 28a-PLB | The synthetic codon-optimized coding sequence of PLB from Pseudomonas fluorescens inserted between Nco I and Xho I sites of pET28 | This work |
| pMal-c5x | Expression vector, AmpR, lacIq, ORF MBP, Fx, ori | NEB |
| pMal-c5x-PLB | The synthetic codon-optimized coding sequence of PLB inserted between Nco I and Bam HI sites of pMal-c5x | This work |
| pTf16 | tig, CmR, araB, pACYC ori | TaKaRa |
| pKJE7 | dnaK-dnaJ-grpE, CmR, araB, pACYC ori | TaKaRa |
| pGro7 | groES-groEL, CmR, araB, pACYC ori | TaKaRa |
| pG-Tf2 | groES-groEL-tig, CmR, Pzt-1, pACYC ori | TaKaRa |
| pG-KJE8 | dnaK-dnaJ-grpE, groES-groEL, CmR, araB, Pzt-1, pACYC ori | TaKaRa |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Feng, J.; Yang, W.; Lu, Y.; Li, H.; Xu, S.; Wang, X.; Chen, K. Enhancing Soluble Expression of Phospholipase B for Efficient Catalytic Synthesis of L-Alpha-Glycerylphosphorylcholine. Catalysts 2022, 12, 650. https://doi.org/10.3390/catal12060650
Feng J, Yang W, Lu Y, Li H, Xu S, Wang X, Chen K. Enhancing Soluble Expression of Phospholipase B for Efficient Catalytic Synthesis of L-Alpha-Glycerylphosphorylcholine. Catalysts. 2022; 12(6):650. https://doi.org/10.3390/catal12060650
Chicago/Turabian StyleFeng, Jiao, Wenjing Yang, Yuanyuan Lu, Hui Li, Sheng Xu, Xin Wang, and Kequan Chen. 2022. "Enhancing Soluble Expression of Phospholipase B for Efficient Catalytic Synthesis of L-Alpha-Glycerylphosphorylcholine" Catalysts 12, no. 6: 650. https://doi.org/10.3390/catal12060650
APA StyleFeng, J., Yang, W., Lu, Y., Li, H., Xu, S., Wang, X., & Chen, K. (2022). Enhancing Soluble Expression of Phospholipase B for Efficient Catalytic Synthesis of L-Alpha-Glycerylphosphorylcholine. Catalysts, 12(6), 650. https://doi.org/10.3390/catal12060650