

Recent Progress in the Mechanism and Engineering of α/β Hydrolases for Chiral Chemical Production


 and
and
Abstract
:1. Introduction
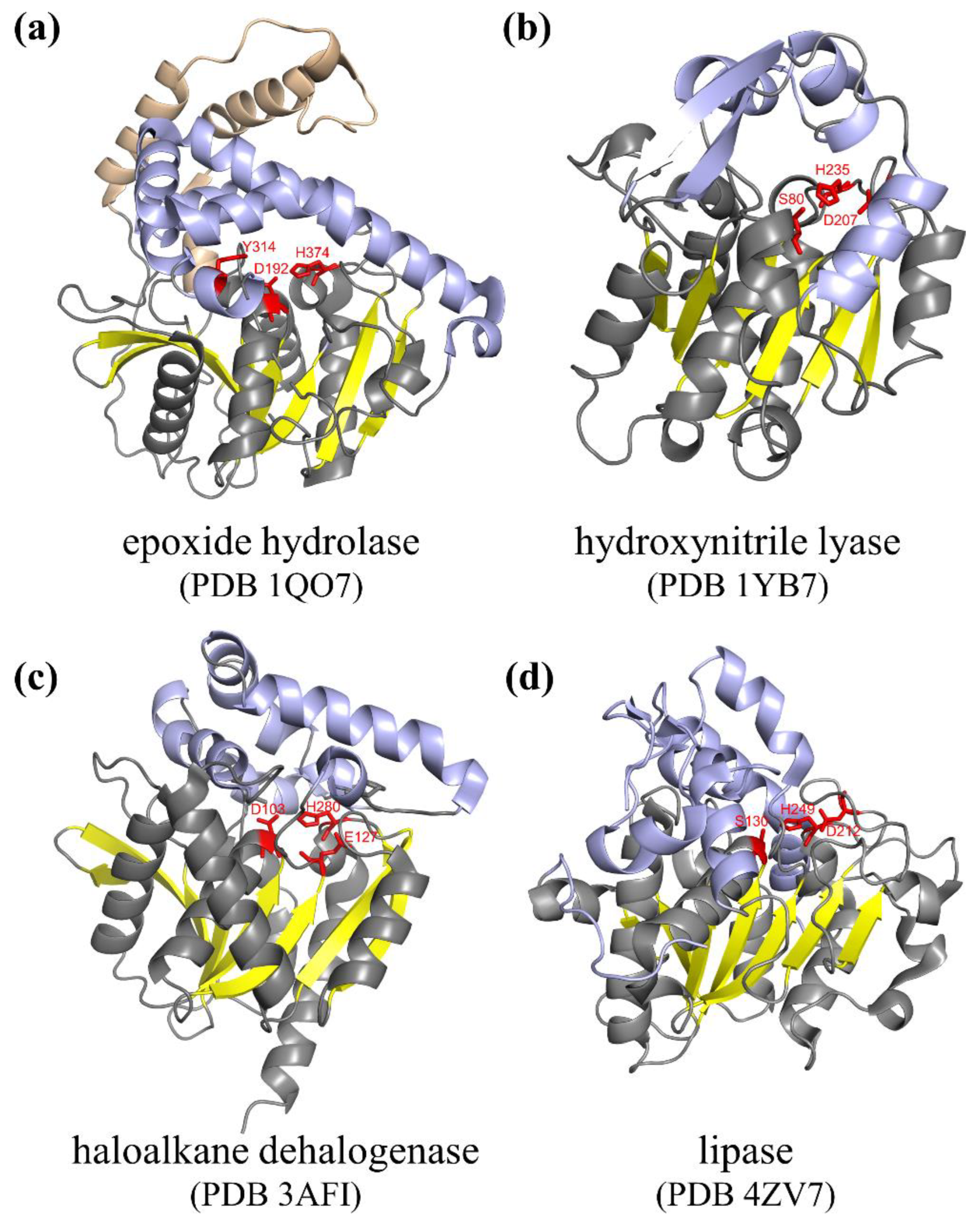
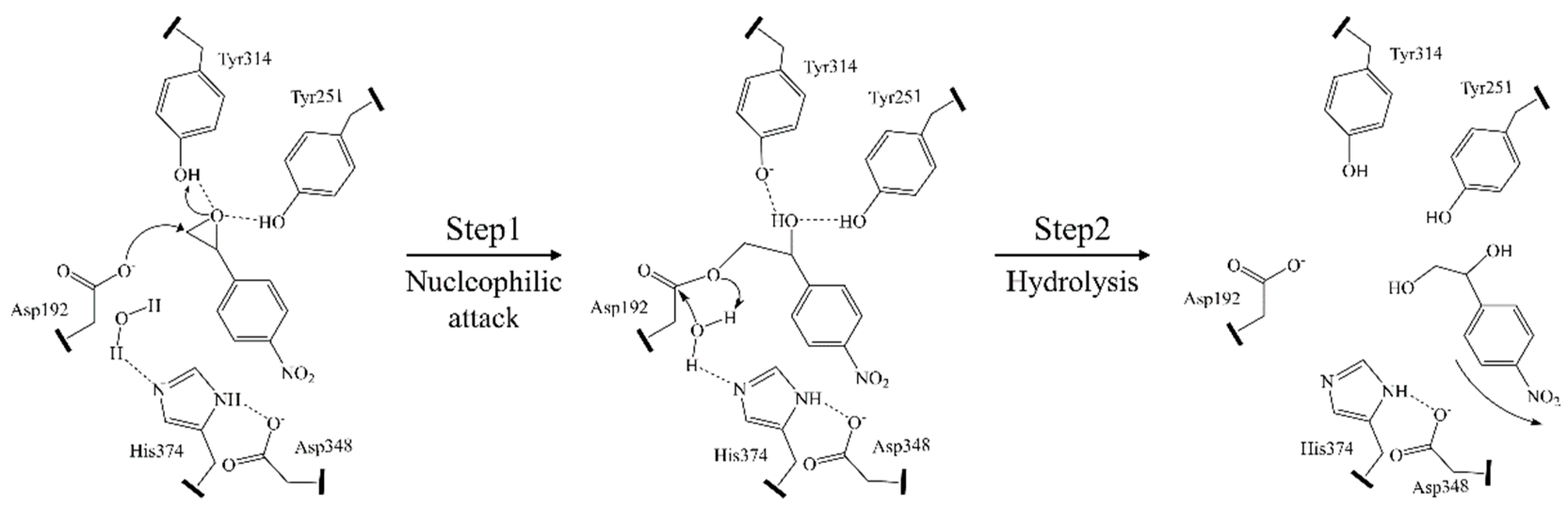
2. Structure and Catalytic Mechanism of α/β Hydrolase
3. Regio- and Stereo-Selectivity of α/β Hydrolases
4. Screening of New α/β Hydrolases from Nature
5. Engineering of Natural α/β Hydrolases
5.1. Engineering of α/β Hydrolases to Catalyze Different Reactions
5.2. Catalytic Activity Enhancement of α/β Hydrolases
5.3. Regio- and Stereo-Selectivity Engineering of α/β Hydrolases
5.4. Stability Enhancement of α/β Hydrolases
6. Other Progress in Biocatalyst Development Based on α/β Hydrolases
7. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Hilterhaus, L.; Liese, A. Building blocks. Adv. Biochem. Eng. Biotechnol. 2007, 105, 133–173. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Ryoo, D.H.; Lee, J.M.; Ryoo, J.J. Synthesis and application of C2 and C3 symmetric (R)-phenylglycinol-derived chiral stationary phases. Chirality 2016, 28, 186–191. [Google Scholar] [CrossRef]
- Hu, X.; Guo, X.; Sun, S.; Zhu, B.; Yu, J.; Guo, X. Enantioseparation of nine indanone and tetralone derivatives by HPLC using carboxymethyl-β-cyclodextrin as the mobile phase additive. Chirality 2017, 29, 38–47. [Google Scholar] [CrossRef]
- Xie, S.M.; Fu, N.; Li, L.; Yuan, B.Y.; Zhang, J.H.; Li, Y.X.; Yuan, L.M. Homochiral metal-organic cage for gas chromatographic separations. Anal. Chem. 2018, 90, 9182–9188. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.; Mu, X.; Qiao, J.; Su, Y.; Qi, L. New chiral ligand exchange capillary electrophoresis system with chiral amino amide ionic liquids as ligands. Talanta 2017, 175, 451–456. [Google Scholar] [CrossRef]
- Lynch, C.C.; De Los Santos, Z.A.; Wolf, C. Chiroptical sensing of unprotected amino acids, hydroxy acids, amino alcohols, amines and carboxylic acids with metal salts. Chem. Commun. 2019, 55, 6297–6300. [Google Scholar] [CrossRef]
- Wu, S.; Yin, Z.Z.; Wu, D.; Tao, Y.; Kong, Y. Chiral enantioselective assemblies induced from achiral porphyrin by L- and D-lysine. Langmuir 2019, 35, 16761–16769. [Google Scholar] [CrossRef]
- Wang, L.; Jin, Z.; Wang, X.; Zeng, S.; Sun, C.; Pan, Y. Pair of stereodynamic chiral benzylicaldehyde probes for determination of absolute configuration of amino acid residues in peptides by mass spectrometry. Anal. Chem. 2017, 89, 11902–11907. [Google Scholar] [CrossRef]
- Wu, D.; Yu, Y.; Zhang, J.; Guo, L.; Kong, Y. Chiral poly(ionic liquid) with nonconjugated backbone as a fluorescent enantioselective sensor for phenylalaninol and tryptophan. ACS Appl. Mater. Interfaces 2018, 10, 23362–23368. [Google Scholar] [CrossRef] [PubMed]
- Temel, F.; Erdemir, S.; Tabakci, B.; Akpinar, M.; Tabakci, M. Selective chiral recognition of alanine enantiomers by chiral calix [4]arene coated quartz crystal microbalance sensors. Anal. Bioanal. Chem. 2019, 411, 2675–2685. [Google Scholar] [CrossRef]
- Jiang, W.; Fang, B. Synthesizing chiral drug intermediates by biocatalysis. Appl. Biochem. Biotechnol. 2020, 192, 146–179. [Google Scholar] [CrossRef]
- Patel, R.N. Biocatalysis for synthesis of pharmaceuticals. Bioorg. Med. Chem. 2018, 26, 1252–1274. [Google Scholar] [CrossRef]
- Sun, H.; Zhang, H.; Ang, E.L.; Zhao, H. Biocatalysis for the synthesis of pharmaceuticals and pharmaceutical intermediates. Bioorg. Med. Chem. 2018, 26, 1275–1284. [Google Scholar] [CrossRef] [PubMed]
- Holmquist, M. Alpha/Beta-hydrolase fold enzymes: Structures, functions and mechanisms. Curr. Protein Pept. Sci. 2000, 1, 209–235. [Google Scholar] [CrossRef] [PubMed]
- Lee, E.Y. Epoxide hydrolase-mediated enantioconvergent bioconversions to prepare chiral epoxides and alcohols. Biotechnol. Lett. 2008, 30, 1509–1514. [Google Scholar] [CrossRef]
- Koudelakova, T.; Bidmanova, S.; Dvorak, P.; Pavelka, A.; Chaloupkova, R.; Prokop, Z.; Damborsky, J. Haloalkane dehalogenases: Biotechnological applications. Biotechnol. J. 2013, 8, 32–45. [Google Scholar] [CrossRef]
- Wu, S.; Snajdrova, R.; Moore, J.C.; Baldenius, K.; Bornscheuer, U.T. Biocatalysis: Enzymatic synthesis for industrial applications. Angew. Chem. Int. Ed. 2021, 60, 88–119. [Google Scholar] [CrossRef]
- Saini, P.; Sareen, D. An overview on the enhancement of enantioselectivity and stability of microbial epoxide hydrolases. Mol. Biotechnol. 2017, 59, 98–116. [Google Scholar] [CrossRef]
- Schanstra, J.P.; Janssen, D.B. Kinetics of halide release of haloalkane dehalogenase: Evidence for a slow conformational change. Biochemistry 1996, 35, 5624–5632. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rea, D.; Fülöp, V. Prolyl oligopeptidase structure and dynamics. CNS Neurol. Disord. Drug Targets 2011, 10, 306–310. [Google Scholar] [CrossRef]
- Pries, F.; van den Wijngaard, A.J.; Bos, R.; Pentenga, M.; Janssen, D.B. The role of spontaneous cap domain mutations in haloalkane dehalogenase specificity and evolution. J. Biol. Chem. 1994, 269, 17490–17494. [Google Scholar] [CrossRef]
- Zou, J.; Hallberg, B.M.; Bergfors, T.; Oesch, F.; Arand, M.; Mowbray, S.L.; Jones, T.A. Structure of Aspergillus niger epoxide hydrolase at 1.8 Å resolution: Implications for the structure and function of the mammalian microsomal class of epoxide hydrolases. Structure 2000, 15, 111–122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gartler, G.; Kratky, C.; Gruber, K. Structural determinants of the enantioselectivity of the hydroxynitrile lyase from Hevea brasiliensis. J. Biotechnol. 2007, 129, 87–97. [Google Scholar] [CrossRef]
- Prokop, Z.; Sato, Y.; Brezovsky, J.; Mozga, T.; Chaloupkova, R.; Koudelakova, T.; Jerabek, P.; Stepankova, V.; Natsume, R.; van Leeuwen, J.G.; et al. Enantioselectivity of haloalkane dehalogenases and its modulation by surface loop engineering. Angew. Chem. Int. Ed. 2010, 49, 6111–6115. [Google Scholar] [CrossRef] [PubMed]
- Strzelczyk, P.; Bujacz, G.D.; Kielbasinski, P.; Blaszczyk, J. Crystal and molecular structure of hexagonal form of lipase B from Candida antarctica. Acta Biochim. Pol. 2016, 63, 103–109. [Google Scholar] [CrossRef] [PubMed]
- Lau, E.Y.; Newby, Z.E.; Bruice, T.C. A theoretical examination of the acid-catalyzed and noncatalyzed ring-opening reaction of an oxirane by nucleophilic addition of acetate. Implications to epoxide hydrolases. J. Am. Chem. Soc. 2001, 123, 3350–3357. [Google Scholar] [CrossRef] [PubMed]
- Widersten, M.; Gurell, A.; Lindberg, D. Structure-function relationships of epoxide hydrolases and their potential use in biocatalysis. Biochim. Biophys. Acta 2010, 1800, 316–326. [Google Scholar] [CrossRef]
- Orru, R.V.; Faber, K. Stereoselectivities of microbial epoxide hydrolases. Curr. Opin. Chem. Biol. 1999, 3, 16–21. [Google Scholar] [CrossRef] [PubMed]
- Casas-Godoy, L.; Gasteazoro, F.; Duquesne, S.; Bordes, F.; Marty, A.; Sandoval, G. Lipases: An overview. Methods Mol. Biol. 2018, 1835, 3–38. [Google Scholar] [CrossRef]
- Jaeger, K.E.; Dijkstra, B.W.; Reetz, M.T. Bacterial biocatalysts: Molecular biology, three-dimensional structures, and biotechnological applications of lipases. Annu. Rev. Microbiol. 1999, 53, 315–351. [Google Scholar] [CrossRef] [Green Version]
- Lang, D.A.; Mannesse, M.L.; de Haas, G.H.; Verheij, H.M.; Dijkstra, B.W. Structural basis of the chiral selectivity of Pseudomonas cepacia lipase. Eur. J. Biochem. 1998, 254, 333–340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meng, X.; Guo, L.; Xu, G.; Wu, J.-P.; Yang, L.-R. A new mechanism of enantioselectivity toward chiral primary alcohol by lipase from Pseudomonas cepacia. J. Mol. Catal. B Enzym. 2014, 109, 109–115. [Google Scholar] [CrossRef]
- Bordes, F.; Cambon, E.; Dossat-Letisse, V.; Andre, I.; Croux, C.; Nicaud, J.M.; Marty, A. Improvement of Yarrowia lipolytica lipase enantioselectivity by using mutagenesis targeted to the substrate binding site. ChemBioChem 2009, 10, 1705–1713. [Google Scholar] [CrossRef]
- Kim, C.; Lee, J.; Cho, J.; Oh, Y.; Choi, Y.K.; Choi, E.; Park, J.; Kim, M.J. Kinetic and dynamic kinetic resolution of secondary alcohols with ionic-surfactant-coated Burkholderia cepacia lipase: Substrate scope and enantioselectivity. J. Org. Chem. 2013, 78, 2571–2578. [Google Scholar] [CrossRef] [PubMed]
- Kitamoto, Y.; Kuruma, Y.; Suzuki, K.; Hattori, T. Effect of solvent polarity on enantioselectivity in Candida antarctica lipase B catalyzed kinetic resolution of primary and secondary alcohols. J. Org. Chem. 2015, 80, 521–527. [Google Scholar] [CrossRef] [PubMed]
- Maldonado, M.R.; Alnoch, R.C.; de Almeida, J.M.; dos Santos, L.A.; Andretta, A.T.; Ropaín, R.d.P.C.; de Souza, E.M.; Mitchell, D.A.; Krieger, N. Key mutation sites for improvement of the enantioselectivity of lipases through protein engineering. Biochem. Eng. J. 2021, 172, 108047. [Google Scholar] [CrossRef]
- Adamu, A.; Wahab, R.A.; Aliyu, F.; Aminu, A.H.; Hamza, M.M.; Huyop, F. Haloacid dehalogenases of Rhizobium sp. and related enzymes: Catalytic properties and mechanistic analysis. Process. Biochem. 2020, 92, 437–446. [Google Scholar] [CrossRef]
- van Leeuwen, J.G.; Wijma, H.J.; Floor, R.J.; van der Laan, J.M.; Janssen, D.B. Directed evolution strategies for enantiocomplementary haloalkane dehalogenases: From chemical waste to enantiopure building blocks. ChemBioChem 2012, 13, 137–148. [Google Scholar] [CrossRef] [Green Version]
- Nuylert, A.; Nakabayashi, M.; Yamaguchi, T.; Asano, Y. Discovery and structural analysis to improve the enantioselectivity of hydroxynitrile lyase from Parafontaria laminata millipedes for (R)-2-chloromandelonitrile synthesis. ACS Omega 2020, 5, 27896–27908. [Google Scholar] [CrossRef]
- Vishnu Priya, B.; Sreenivasa Rao, D.H.; Gilani, R.; Lata, S.; Rai, N.; Akif, M.; Kumar Padhi, S. Enzyme engineering improves catalytic efficiency and enantioselectivity of hydroxynitrile lyase for promiscuous retro-nitroaldolase activity. Bioorg. Chem. 2022, 120, 105594. [Google Scholar] [CrossRef]
- Ding, Y.; Perez-Ortiz, G.; Peate, J.; Barry, S.M. Redesigning enzymes for biocatalysis: Exploiting structural understanding for improved selectivity. Front. Mol. Biosci. 2022, 9, 908285. [Google Scholar] [CrossRef] [PubMed]
- Rao, D.H.S.; Shivani, K.; Padhi, S.K. Immobilized Arabidopsis thaliana hydroxynitrile lyase-catalyzed retro-Henry reaction in the synthesis of (S)-β-nitroalcohols. Appl. Biochem. Biotechnol. 2021, 193, 560–576. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Yu, W.; Shen, L.; Zheng, H.; Guo, X.; Zhong, J.; Hu, T. Conjugation of haloalkane dehalogenase DhaA with arabinogalactan to increase its stability. J. Biotechnol. 2021, 335, 47–54. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Q.Q.; He, W.H.; Kong, X.D.; Fan, L.Q.; Zhao, J.; Li, S.X.; Xu, J.H. Heterologous overexpression of Vigna radiata epoxide hydrolase in Escherichia coli and its catalytic performance in enantioconvergent hydrolysis of p-nitrostyrene oxide into (R)-p-nitrophenyl glycol. Appl. Microbiol. Biotechnol. 2014, 98, 207–218. [Google Scholar] [CrossRef]
- Xu, W.; Xu, J.-H.; Pan, J.; Gu, Q.; Wu, X.-Y. Enantioconvergent hydrolysis of styrene epoxides by newly discovered epoxide hydrolases in mung bean. Org. Lett. 2006, 8, 1737–1740. [Google Scholar] [CrossRef]
- Hu, D.; Tang, C.; Li, C.; Kan, T.; Shi, X.; Feng, L.; Wu, M. Stereoselective hydrolysis of epoxides by reVrEH3, a novel Vigna radiata epoxide hydrolase with high enantioselectivity or high and complementary regioselectivity. J. Agric. Food Chem. 2017, 65, 9861–9870. [Google Scholar] [CrossRef]
- Hu, D.; Tang, C.-D.; Yang, B.; Liu, J.-C.; Yu, T.; Deng, C.; Wu, M.-C. Expression of a novel epoxide hydrolase of Aspergillus usamii E001 in Escherichia coli and its performance in resolution of racemic styrene oxide. J. Ind. Microbiol. Biotechnol. 2015, 42, 671–680. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Li, C.; Zhu, X.X.; Liu, Y.Y.; Zhao, J.; Wu, M.C. Highly regio- and enantio-selective hydrolysis of two racemic epoxides by GmEH3, a novel epoxide hydrolase from Glycine max. Int. J. Biol. Macromol. 2020, 164, 2795–2803. [Google Scholar] [CrossRef]
- Xue, F.; Liu, Z.-Q.; Zou, S.-P.; Wan, N.-W.; Zhu, W.-Y.; Zhu, Q.; Zheng, Y.-G. A novel enantioselective epoxide hydrolase from Agromyces mediolanus ZJB120203: Cloning, characterization and application. Process. Biochem. 2014, 49, 409–417. [Google Scholar] [CrossRef]
- Handelsmanl, J.; Rondon, M.R.; Brady, S.F.; Clardy, J.; Goodman, R.M. Molecular biological access to the chemistry of unknown soil microbes: A new frontier for natural products. Chem. Biol. 1998, 5, R245–R249. [Google Scholar] [CrossRef] [Green Version]
- Stojanovski, G.; Dobrijevic, D.; Hailes, H.C.; Ward, J.M. Identification and catalytic properties of new epoxide hydrolases from the genomic data of soil bacteria. Enzyme Microb. Technol. 2020, 139, 109592. [Google Scholar] [CrossRef]
- Ferrandi, E.E.; Sayer, C.; De Rose, S.A.; Guazzelli, E.; Marchesi, C.; Saneei, V.; Isupov, M.N.; Littlechild, J.A.; Monti, D. New thermophilic α/β class epoxide hydrolases found in metagenomes from hot environments. Front. Bioeng. Biotechnol. 2018, 6, 144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jimenez, D.J.; Dini-Andreote, F.; Ottoni, J.R.; de Oliveira, V.M.; van Elsas, J.D.; Andreote, F.D. Compositional profile of α/β-hydrolase fold proteins in mangrove soil metagenomes: Prevalence of epoxide hydrolases and haloalkane dehalogenases in oil-contaminated sites. Microb. Biotechnol. 2015, 8, 604–613. [Google Scholar] [CrossRef] [PubMed]
- Wu, K.; Wang, H.; Sun, H.; Wei, D. Efficient kinetic resolution of phenyl glycidyl ether by a novel epoxide hydrolase from Tsukamurella paurometabola. Appl. Microbiol. Biotechnol. 2015, 99, 9511–9521. [Google Scholar] [CrossRef] [PubMed]
- Ottoni, J.R.; Cabral, L.; de Sousa, S.T.P.; Junior, G.V.L.; Domingos, D.F.; Soares Junior, F.L.; da Silva, M.C.P.; Marcon, J.; Dias, A.C.F.; de Melo, I.S.; et al. Functional metagenomics of oil-impacted mangrove sediments reveals high abundance of hydrolases of biotechnological interest. World J. Microbiol. Biotechnol. 2017, 33, 141. [Google Scholar] [CrossRef]
- Zhang, L.; De, B.C.; Zhang, W.; Mandi, A.; Fang, Z.; Yang, C.; Zhu, Y.; Kurtan, T.; Zhang, C. Mutation of an atypical oxirane oxyanion hole improves regioselectivity of the α/β-fold epoxide hydrolase Alp1U. J. Biol. Chem. 2020, 295, 16987–16997. [Google Scholar] [CrossRef] [PubMed]
- Kotik, M.; Stepanek, V.; Kyslik, P.; Maresova, H. Cloning of an epoxide hydrolase-encoding gene from Aspergillus niger M200, overexpression in E. coli, and modification of activity and enantioselectivity of the enzyme by protein engineering. J. Biotechnol. 2007, 132, 8–15. [Google Scholar] [CrossRef]
- Zou, S.P.; Zheng, Y.G.; Wu, Q.; Wang, Z.C.; Xue, Y.P.; Liu, Z.Q. Enhanced catalytic efficiency and enantioselectivity of epoxide hydrolase from Agrobacterium radiobacter AD1 by iterative saturation mutagenesis for (R)-epichlorohydrin synthesis. Appl. Microbiol. Biotechnol. 2018, 102, 733–742. [Google Scholar] [CrossRef]
- Hu, D.; Hu, B.C.; Hou, X.D.; Zhang, D.; Lei, Y.Q.; Rao, Y.J.; Wu, M.C. Structure-guided regulation in the enantioselectivity of an epoxide hydrolase to produce enantiomeric monosubstituted epoxides and vicinal diols via kinetic resolution. Org. Lett. 2022, 24, 1757–1761. [Google Scholar] [CrossRef]
- Wen, Z.; Hu, D.; Hu, B.C.; Zhang, D.; Huang, J.F.; Wu, M.C. Structure-guided improvement in the enantioselectivity of an Aspergillus usamii epoxide hydrolase for the gram-scale kinetic resolution of ortho-trifluoromethyl styrene oxide. Enzym. Microb. Technol. 2021, 146, 109778. [Google Scholar] [CrossRef]
- Rui, L.; Cao, L.; Chen, W.; Reardon, K.F.; Wood, T.K. Protein engineering of epoxide hydrolase from Agrobacterium radiobacter AD1 for enhanced activity and enantioselective production of (R)-1-phenylethane-1,2-diol. Appl. Environ. Microbiol. 2005, 71, 3995–4003. [Google Scholar] [CrossRef] [Green Version]
- Horsman, G.P.; Lechner, A.; Ohnishi, Y.; Moore, B.S.; Shen, B. Predictive model for epoxide hydrolase-generated stereochemistry in the biosynthesis of nine-membered enediyne antitumor antibiotics. Biochemistry 2013, 52, 5217–5224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jochens, H.; Stiba, K.; Savile, C.; Fujii, R.; Yu, J.G.; Gerassenkov, T.; Kazlauskas, R.J.; Bornscheuer, U.T. Converting an esterase into an epoxide hydrolase. Angew. Chem. Int. Ed. 2009, 48, 3532–3535. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Luan, Z.; Chen, Q.; Xu, J.; Yu, H. Rational selection of circular permutation sites in characteristic regions of the α/β-hydrolase fold enzyme RhEst1. J. Mol. Catal. B: Enzym. 2016, 125, 75–80. [Google Scholar] [CrossRef]
- Jones, B.J.; Lim, H.Y.; Huang, J.; Kazlauskas, R.J. Comparison of five protein engineering strategies for stabilizing an α/β-hydrolase. Biochemistry 2017, 56, 6521–6532. [Google Scholar] [CrossRef]
- Padhi, S.K.; Fujii, R.; Legatt, G.A.; Fossum, S.L.; Berchtold, R.; Kazlauskas, R.J. Switching from an esterase to a hydroxynitrile lyase mechanism requires only two amino acid substitutions. Chem. Biol. 2010, 17, 863–871. [Google Scholar] [CrossRef] [Green Version]
- Beier, A.; Damborsky, J.; Prokop, Z. Transhalogenation catalysed by haloalkane dehalogenases engineered to stop natural pathway at intermediate. Adv. Synth. Catal. 2019, 361, 2438–2442. [Google Scholar] [CrossRef]
- Marek, M.; Chaloupkova, R.; Prudnikova, T.; Sato, Y.; Rezacova, P.; Nagata, Y.; Kuta Smatanova, I.; Damborsky, J. Structural and catalytic effects of surface loop-helix transplantation within haloalkane dehalogenase family. Comput. Struct. Biotechnol. J. 2020, 18, 1352–1362. [Google Scholar] [CrossRef]
- Bednar, D.; Beerens, K.; Sebestova, E.; Bendl, J.; Khare, S.; Chaloupkova, R.; Prokop, Z.; Brezovsky, J.; Baker, D.; Damborsky, J. FireProt: Energy- and evolution-based computational design of thermostable multiple-point mutants. PLoS Comput. Biol. 2015, 11, e1004556. [Google Scholar] [CrossRef]
- Floor, R.J.; Wijma, H.J.; Colpa, D.I.; Ramos-Silva, A.; Jekel, P.A.; Szymanski, W.; Feringa, B.L.; Marrink, S.J.; Janssen, D.B. Computational library design for increasing haloalkane dehalogenase stability. ChemBioChem 2014, 15, 1660–1672. [Google Scholar] [CrossRef] [Green Version]
- von Langermann, J.; Nedrud, D.M.; Kazlauskas, R.J. Increasing the reaction rate of hydroxynitrile lyase from Hevea brasiliensis toward mandelonitrile by copying active site residues from an esterase that accepts aromatic esters. ChemBioChem 2014, 15, 1931–1938. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nedrud, D.M.; Lin, H.; Lopez, G.; Padhi, S.K.; Legatt, G.A.; Kaz-Lauskas, R.J. Uncovering divergent evolution of α/β-hydrolases: A surprising residue substitution needed to convert Hevea brasiliensis hydroxynitrile lyase into an esterase. Chem. Sci. 2014, 5, 4265–4277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, Y.; Lutz, S. Improved triglyceride transesterification by circular permuted Candida antarctica lipase B. Biotechnol. Bioeng. 2010, 105, 44–50. [Google Scholar] [CrossRef] [PubMed]
- Paye, M.F.; Rose, H.B.; Robbins, J.M.; Yunda, D.A.; Cho, S.; Bommarius, A.S. A high-throughput pH-based colorimetric assay: Application focus on alpha/beta hydrolases. Anal. Biochem. 2018, 549, 80–90. [Google Scholar] [CrossRef]
- Zheng, Y.C.; Ding, L.Y.; Jia, Q.; Lin, Z.; Hong, R.; Yu, H.L.; Xu, J.H. A high-throughput screening method for the directed evolution of hydroxynitrile lyase towards cyanohydrin synthesis. ChemBioChem 2021, 22, 996–1000. [Google Scholar] [CrossRef]
- Glasner, M.E.; Truong, D.P.; Morse, B.C. How enzyme promiscuity and horizontal gene transfer contribute to metabolic innovation. FEBS J. 2020, 287, 1323–1342. [Google Scholar] [CrossRef] [Green Version]
- Kapoor, M.; Gupta, M.N. Lipase promiscuity and its biochemical applications. Process. Biochem. 2012, 47, 555–569. [Google Scholar] [CrossRef]
- Cadet, F.; Fontaine, N.; Li, G.; Sanchis, J.; Ng Fuk Chong, M.; Pandjaitan, R.; Vetrivel, I.; Offmann, B.; Reetz, M.T. A machine learning approach for reliable prediction of amino acid interactions and its application in the directed evolution of enantioselective enzymes. Sci. Rep. 2018, 8, 16757. [Google Scholar] [CrossRef] [Green Version]
- Bommarius, A.S.; Paye, M.F. Stabilizing biocatalysts. Chem. Soc. Rev. 2013, 42, 6534–6565. [Google Scholar] [CrossRef]
- Soderlund, I.; Tjarnhage, E.; Hamnevik, E.; Widersten, M. Facile synthesis of 2-hydroxyacetophenone from racemic styrene oxide catalyzed by engineered enzymes. Biotechnol. Lett. 2022, 44, 985–990. [Google Scholar] [CrossRef]
- Yu, D.; Wang, J.B.; Reetz, M.T. Exploiting designed oxidase-peroxygenase mutual benefit system for asymmetric cascade reactions. J. Am. Chem. Soc. 2019, 141, 5655–5658. [Google Scholar] [CrossRef] [Green Version]
- Yuan, J.; Lukito, B.R.; Li, Z. De novo biosynthesis of (S)- and (R)-phenylethanediol in yeast via artificial enzyme cascades. ACS Synth. Biol. 2019, 8, 1801–1808. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Hu, D.; Zong, X.; Li, J.; Ding, L.; Wu, M.; Li, J. Enantioconvergent hydrolysis of racemic styrene oxide at high concentration by a pair of novel epoxide hydrolases into (R)-phenyl-1,2-ethanediol. Biotechnol. Lett. 2017, 39, 1917–1923. [Google Scholar] [CrossRef]
- Xuan, J.S.; Feng, Y.G. Enantiomeric tartaric acid production using cis-epoxysuccinate hydrolase: History and perspectives. Molecules 2019, 24, 903. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, R.; Dong, S.; Feng, Y.; Cui, Q.; Xuan, J. Development of highly efficient whole-cell catalysts of cis-epoxysuccinic acid hydrolase by surface display. Bioresour. Bioprocess. 2022, 9, 92. [Google Scholar] [CrossRef]
- Bisogno, F.R.; Orden, A.A.; Pranzoni, C.A.; Cifuente, D.A.; Giordano, O.S.; Kurina Sanz, M. Atypical regioselective biohydrolysis on steroidal oxiranes by Aspergillus niger whole cells: Some stereochemical features. Steroids 2007, 72, 643–652. [Google Scholar] [CrossRef]
- Zheng, H.; Yu, W.L.; Guo, X.; Zhao, Y.Z.; Cui, Y.; Hu, T.; Zhong, J.Y. An effective immobilized haloalkane dehalogenase DhaA from Rhodococcus rhodochrous by adsorption, crosslink and PEGylation on meso-cellular foam. Int. J. Biol. Macromol. 2019, 125, 1016–1023. [Google Scholar] [CrossRef]
- Zhao, Y.Z.; Yu, W.L.; Zheng, H.; Guo, X.; Guo, N.; Hu, T.; Zhong, J.Y. PEGylation with the thiosuccinimido butylamine linker significantly increases the stability of haloalkane dehalogenase DhaA. J. Biotechnol. 2017, 254, 25–33. [Google Scholar] [CrossRef] [PubMed]
- Zou, S.P.; Xuan, X.L.; Wang, Z.J.; Zheng, Y.G. Conjugation of Agrobacterium radiobacter epoxide hydrolase with ficoll: Catalytic, kinetic and thermodynamic analysis. Int. J. Biol. Macromol. 2018, 119, 1098–1105. [Google Scholar] [CrossRef]
- Shan, Y.; Yu, W.; Shen, L.; Guo, X.; Zheng, H.; Zhong, J.; Hu, T.; Han, Y. Conjugation with inulin improves the environmental stability of haloalkane dehalogenase DhaA. Enzym. Microb. Technol. 2021, 149, 109832. [Google Scholar] [CrossRef] [PubMed]
- Huang, R.; Wu, M.; Goldman, M.J.; Li, Z. Encapsulation of enzyme via one-step template-free formation of stable organic-inorganic capsules: A simple and efficient method for immobilizing enzyme with high activity and recyclability. Biotechnol. Bioeng. 2015, 112, 1092–1101. [Google Scholar] [CrossRef] [PubMed]
- Salvi, H.M.; Yadav, G.D. Organic-inorganic epoxide hydrolase hybrid nanoflowers with enhanced catalytic activity: Hydrolysis of styrene oxide to 1-phenyl-1,2-ethanediol. J. Biotechnol. 2021, 341, 113–120. [Google Scholar] [CrossRef] [PubMed]
- Jangir, N.; Padhi, S.K. Immobilized Baliospermum montanum hydroxynitrile lyase catalyzed synthesis of chiral cyanohydrins. Bioorg. Chem. 2019, 84, 32–40. [Google Scholar] [CrossRef]
- Pei, X.; Wang, J.; Zheng, H.; Xiao, Q.; Wang, A.; Su, W. Catalytically active inclusion bodies (CatIBs) induced by terminally attached self-assembling coiled-coil domains: To enhance the stability of (R)-hydroxynitrile lyase. Enzym. Microb. Technol. 2022, 153, 109915. [Google Scholar] [CrossRef] [PubMed]
- Hu, D.; Wang, R.; Shi, X.L.; Ye, H.H.; Wu, Q.; Wu, M.C.; Chu, J.J. Kinetic resolution of racemic styrene oxide at a high concentration by recombinant Aspergillus usamii epoxide hydrolase in an n-hexanol/buffer biphasic system. J. Biotechnol. 2016, 236, 152–158. [Google Scholar] [CrossRef]
- Gao, P.; Wu, S.; Praveen, P.; Loh, K.C.; Li, Z. Enhancing productivity for cascade biotransformation of styrene to (S)-vicinal diol with biphasic system in hollow fiber membrane bioreactor. Appl. Microbiol. Biotechnol. 2017, 101, 1857–1868. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Type | Enzyme | Source | Mutation Site | Reaction/Effects | Reference |
|---|---|---|---|---|---|
| Epoxide hydrolase | Alp1U | Streptomyces ambofaciens | W187F/Y247F | Regioselective nucleophilic attack at C-2 of fluostatin C | [56] |
| Alp1U | Streptomyces ambofaciens | Y247F | Highly regioselective attack at C-3 of fluostatin C | [56] | |
| AmEH | Agromyces mediolanus ZJB120203 | - | Hydrolysis of (R)-ECH to enantiopure (S)-ECH | [49] | |
| AnEH | Aspergillus niger | - | Hydrolysis of epoxides to the more water-soluble and usually less toxic diols | [22] | |
| AnEH | Aspergillus niger | A217L | Improvement in enantioselectivity | [57] | |
| AnEH | Aspergillus niger | A217V | Increase of the activity to allyl glycidyl ether | [57] | |
| AnEH | Aspergillus niger | A217C | Increase of the activity towards allyl glycidyl ether and styrene oxide | [57] | |
| ArEH | Agrobacterium radiobacter AD1 | T247K/I108L/D131S | Improvement of activity, enantioselectivity, and thermostability | [58] | |
| AuEH2 | Aspergillus usamii E001 | - | Resolution of racemic styrene oxide | [47] | |
| AuEH2 | Aspergillus usamii | A214C/A250I | 12.6-fold enhanced enantiomeric ratio toward rac-styrene oxide | [59] | |
| AuEH2 | Aspergillus usamii | R322V/L344C | High enantioselectivity towards rac- ortho-trifluoromethyl styrene oxide | [60] | |
| EchA | Agrobacterium radiobacter AD1 | I219F | Enhanced enantioselectivity for styrene oxide | [61] | |
| EchA | Agrobacterium radiobacter AD1 | L190F | Enhanced activity for styrene oxide | [61] | |
| GmEH3 | Glycine max | - | Enantioconvergent hydrolysis of rac-epoxides with high enantiopurity and yield | [48] | |
| SgcF | Streptomyces griseus IFO 13 350 | W236Y/Q237M | 20-fold increased activity toward (S)-styrene oxide to yield an (S)-diol | [62] | |
| Sibe-EH | metagenomes | - | Desymmetrization of cis-2,3-epoxybutane producing the (2R,3R)-diol | [52] | |
| CH65-EH | metagenomes | - | EH activity toward a broad range of substrates and with high thermostability | [52] | |
| VrEH1 | Vigna radiata | - | Enantioconvergent hydrolysis of p-nitrostyrene oxide | [44,45] | |
| VrEH2 | Vigna radiata | - | Enantioconvergent hydrolysis of p-nitrostyrene oxide | [45] | |
| VrEH3 | Vigna radiata | - | High and complementary regioselectivity toward styrene oxides and high enantioselectivity toward o-cresyl glycidyl ether | [46] | |
| Esterase | PFE | Pseudomonas fluorescens | replacement of a loop (A120-V139) with the corresponding element (P132-Y152) of the epoxide hydrolase EchA | Conversion of an esterase into an epoxide hydrolase towards p-nitrostyrene oxide | [63] |
| RhEst1 | Rhodococcus sp. ECU1013 | circular permutation mutants with G20/T19, S22/N21, and G24&A23 as new termini, respectively | Improved thermostability | [64] | |
| SABP2 | Nicotiana tabacum | Q221M | Higher stability (6.6-fold half-life) | [65] | |
| SABP2 | Nicotiana tabacum | G12T/M239K | Switching from an esterase to a hydroxynitrile lyase | [66] | |
| Haloalkane dehalogenase | DbjA | Bradyrhizobium japonicum USDA110 | - | Excellent enantioselectivity for α-bromoesters and high enantioselectivity for two β-bromoalkanes | [24] |
| DbjA | Bradyrhizobium japonicum USDA110 | H280F | Realized the transhalogenation reaction | [67] | |
| DbeA | Bradyrhizobium elkanii USDA94 | surface loop-helix transplantation from haloalkane dehalogenase DbjA | Lower stability but increased activity with various halogenated substrates and altered its enantioselectivity | [68] | |
| DhaA | Rhodococcus rhodochrous | E20S/F80R/C128F/T148L/A155P/A172I/C176F/D198W/V219W/C262L/D266F | Increased thermostability (ΔTm 24.6 °C) | [69] | |
| DhaA | Rhodococcus rhodochrous | two mutants containing 13 and 17 mutation sites, respectively | Enantioselective production of (R)-and (S)-2,3-dichloropropan-1-ol, respectively | [38] | |
| LinB | Sphingomonas paucimobilis UT26 | E15T/A53L/A81K/F169V/A197P/D255A/A247F | Increased thermostability (ΔTm,app 23 °C) | [70] | |
| Hydroxynitrile lyase | HbHNL | Hevea brasiliensis | L121Y | Improved activity on an unnatural substrate mandelonitrile | [71] |
| HbHNL | Hevea brasiliensis | T11G/E79H/K236G | Lower hydroxynitrile lyase activity and higher esterase-specific activity | [72] | |
| Lipase | CALB | Candida antarctica | - | Kinetic resolution of racemic alcohols and amines or desymmetrization of diols and diacetates | [25] |
| CALB | Candida antarctica | a circular permutated variant of CALB with 283 /282 as the new termini | Higher catalytic activity (2.6- to 9-fold) for trans and interesterification of the different substrates | [73] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qiu, M.; Dong, S.; Cui, Q.; Feng, Y.; Xuan, J. Recent Progress in the Mechanism and Engineering of α/β Hydrolases for Chiral Chemical Production. Catalysts 2023, 13, 288. https://doi.org/10.3390/catal13020288
Qiu M, Dong S, Cui Q, Feng Y, Xuan J. Recent Progress in the Mechanism and Engineering of α/β Hydrolases for Chiral Chemical Production. Catalysts. 2023; 13(2):288. https://doi.org/10.3390/catal13020288
Chicago/Turabian StyleQiu, Mingzhe, Sheng Dong, Qiu Cui, Yingang Feng, and Jinsong Xuan. 2023. "Recent Progress in the Mechanism and Engineering of α/β Hydrolases for Chiral Chemical Production" Catalysts 13, no. 2: 288. https://doi.org/10.3390/catal13020288
APA StyleQiu, M., Dong, S., Cui, Q., Feng, Y., & Xuan, J. (2023). Recent Progress in the Mechanism and Engineering of α/β Hydrolases for Chiral Chemical Production. Catalysts, 13(2), 288. https://doi.org/10.3390/catal13020288