Tailoring Celluclast® Cocktail’s Performance towards the Production of Prebiotic Cello-Oligosaccharides from Waste Forest Biomass

 ,
,  ,
, 

Abstract
:1. Introduction
2. Results
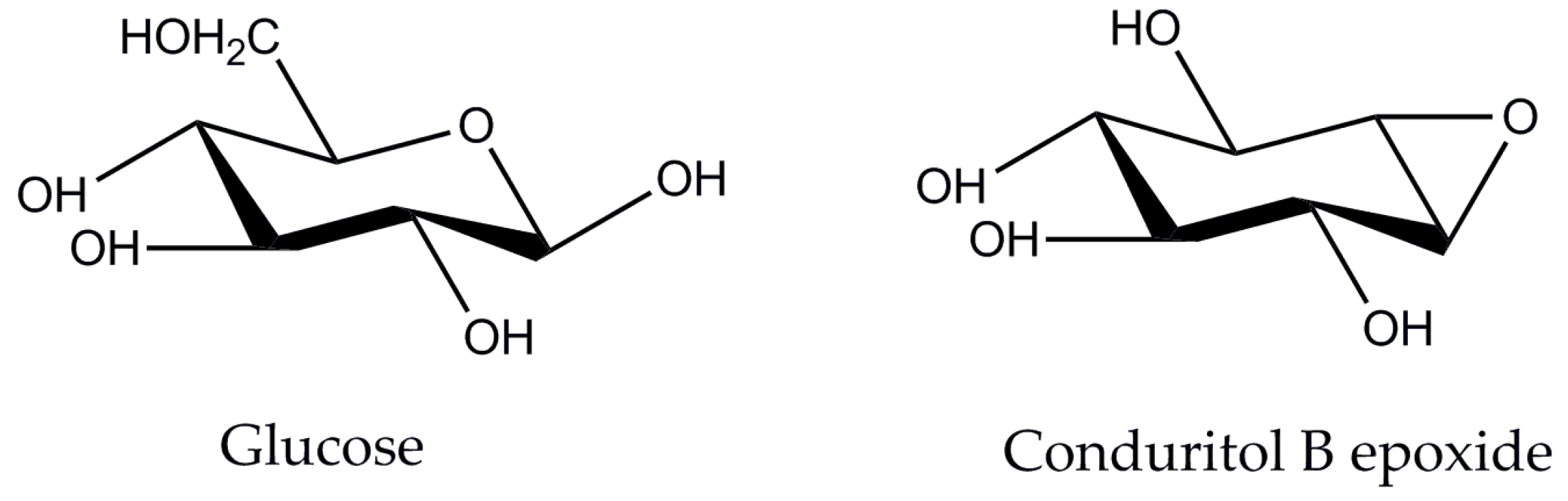
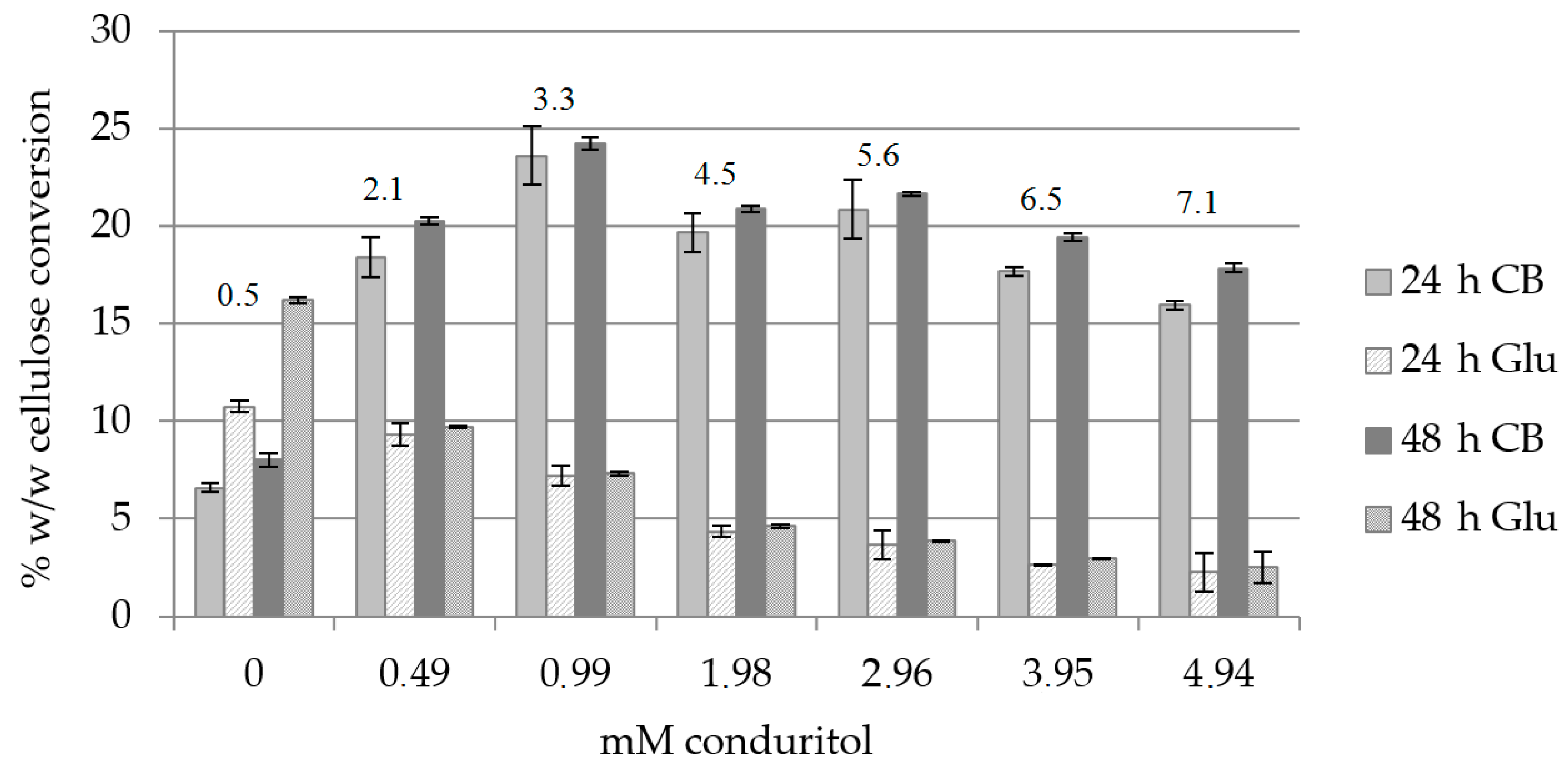
2.1. Effect of β-Glucosidase Inhibitor
2.2. Effect of Enzyme and Conduritol Mixture Preincubation
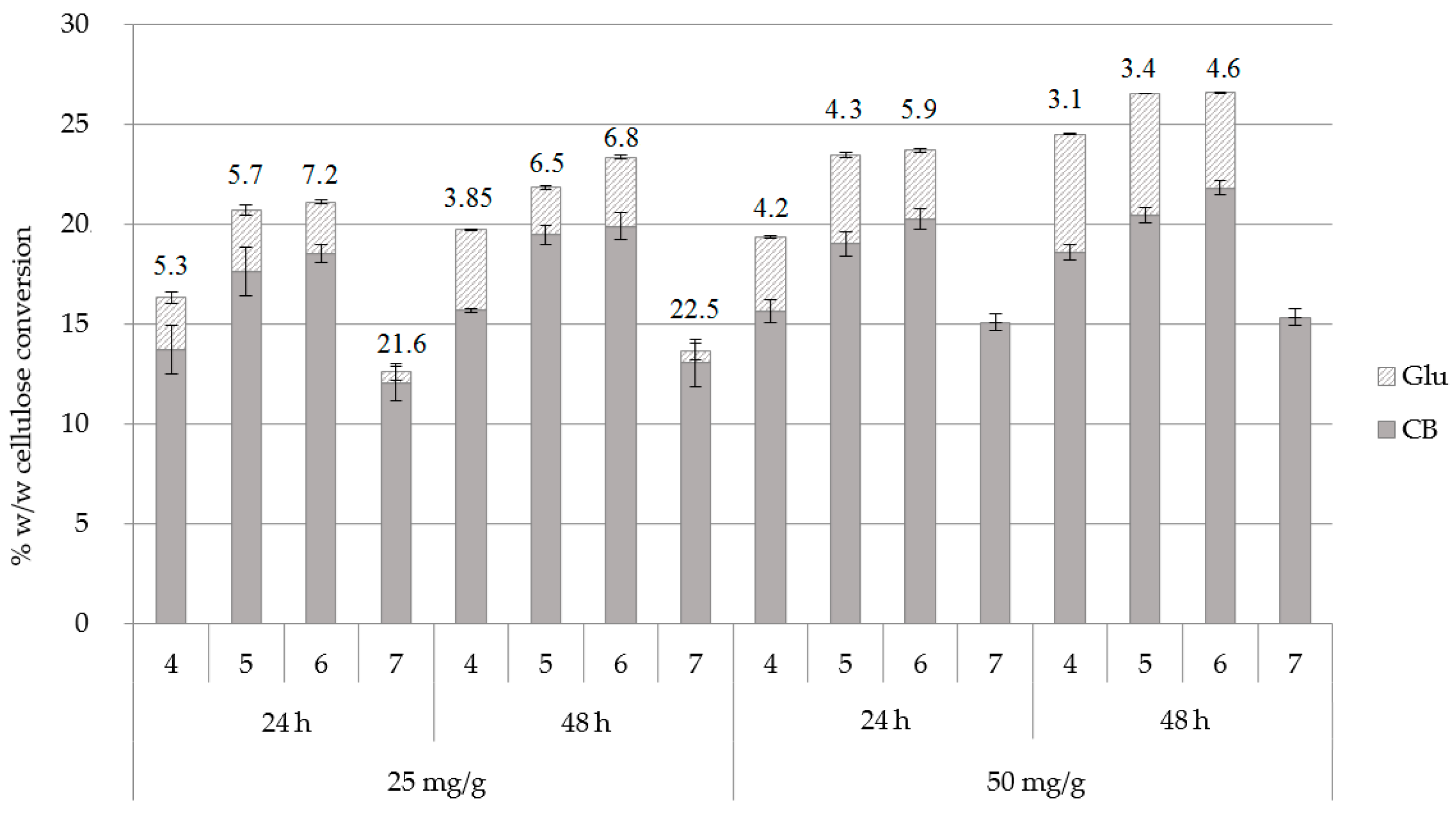
2.3. Effect of pH and Enzyme Loading
2.4. Effect of Hydrolysis Time, Inhibitor Concentration and Reaction Temperature at pH 7.0
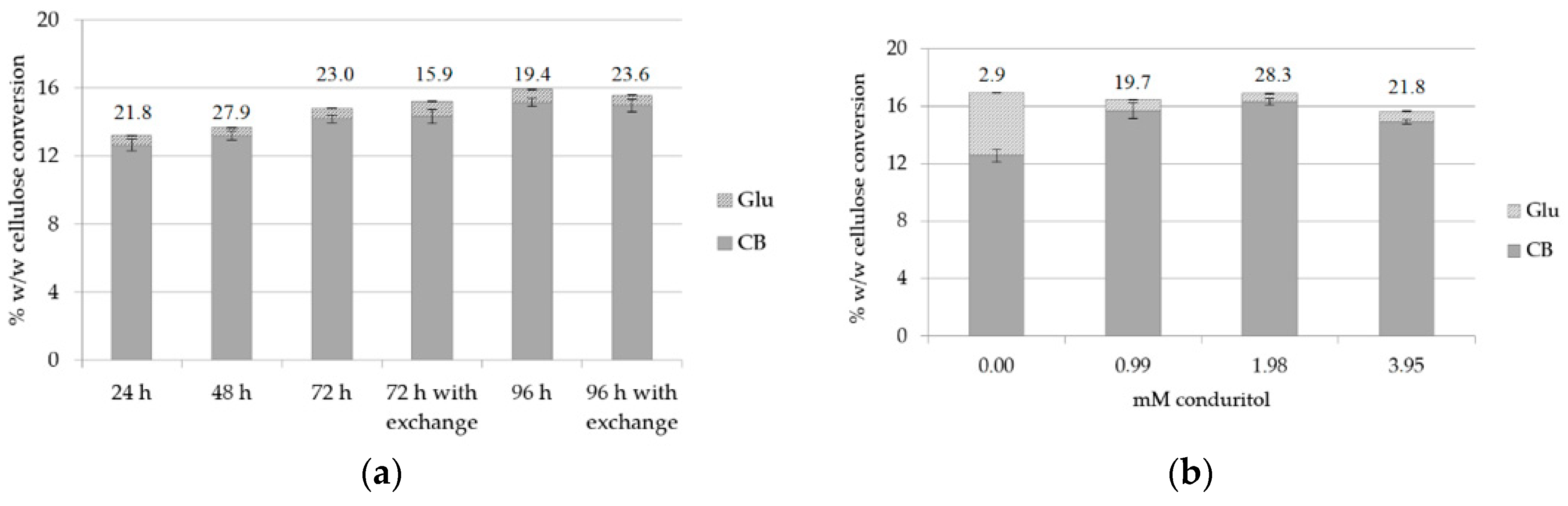
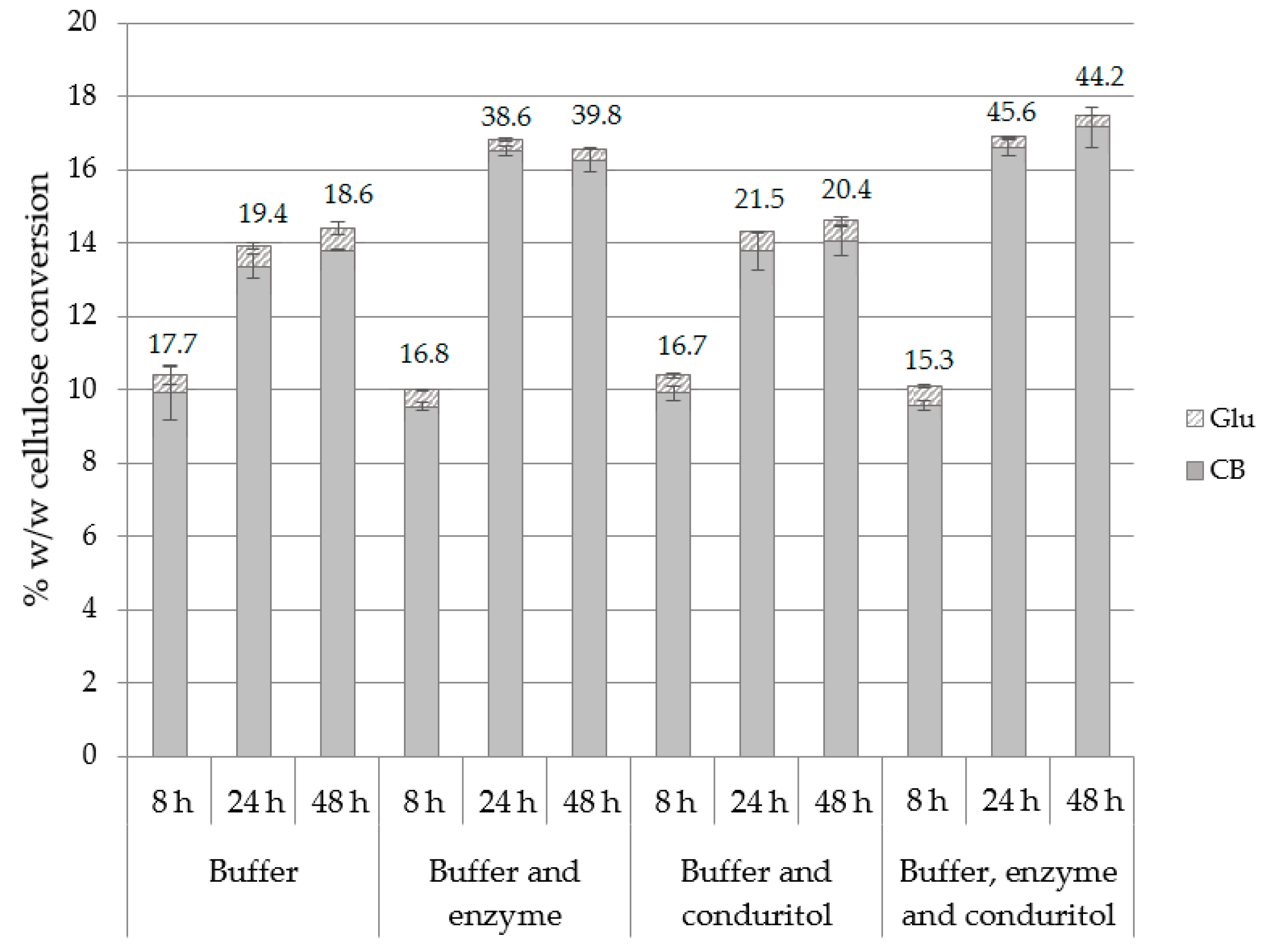
2.5. Effect of Buffer Exchange, Enzyme and Inhibitor Supplementation
2.6. Evaluation of LPMO on the Production of Cellobiose
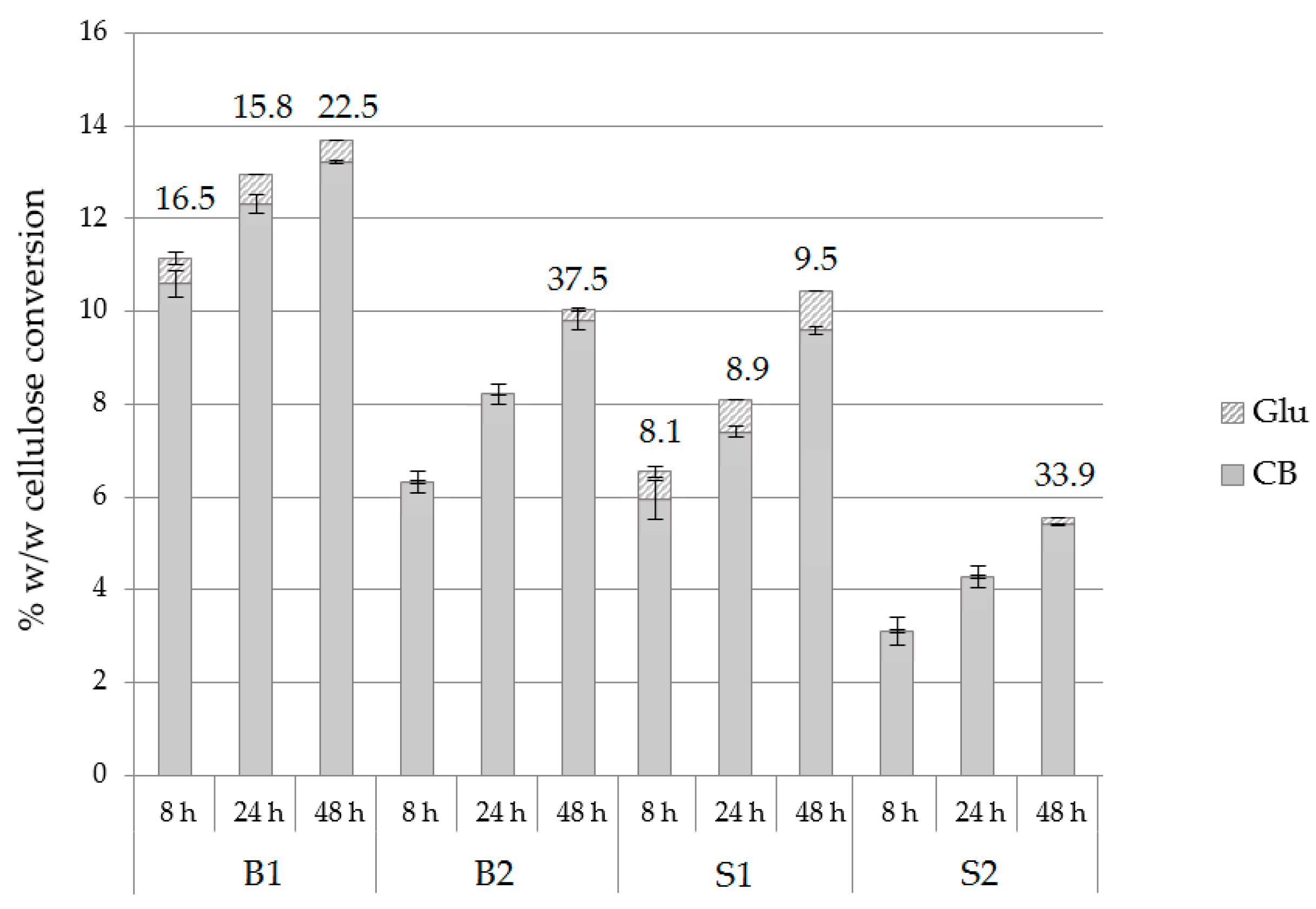
2.7. Evaluation of Different Substrates
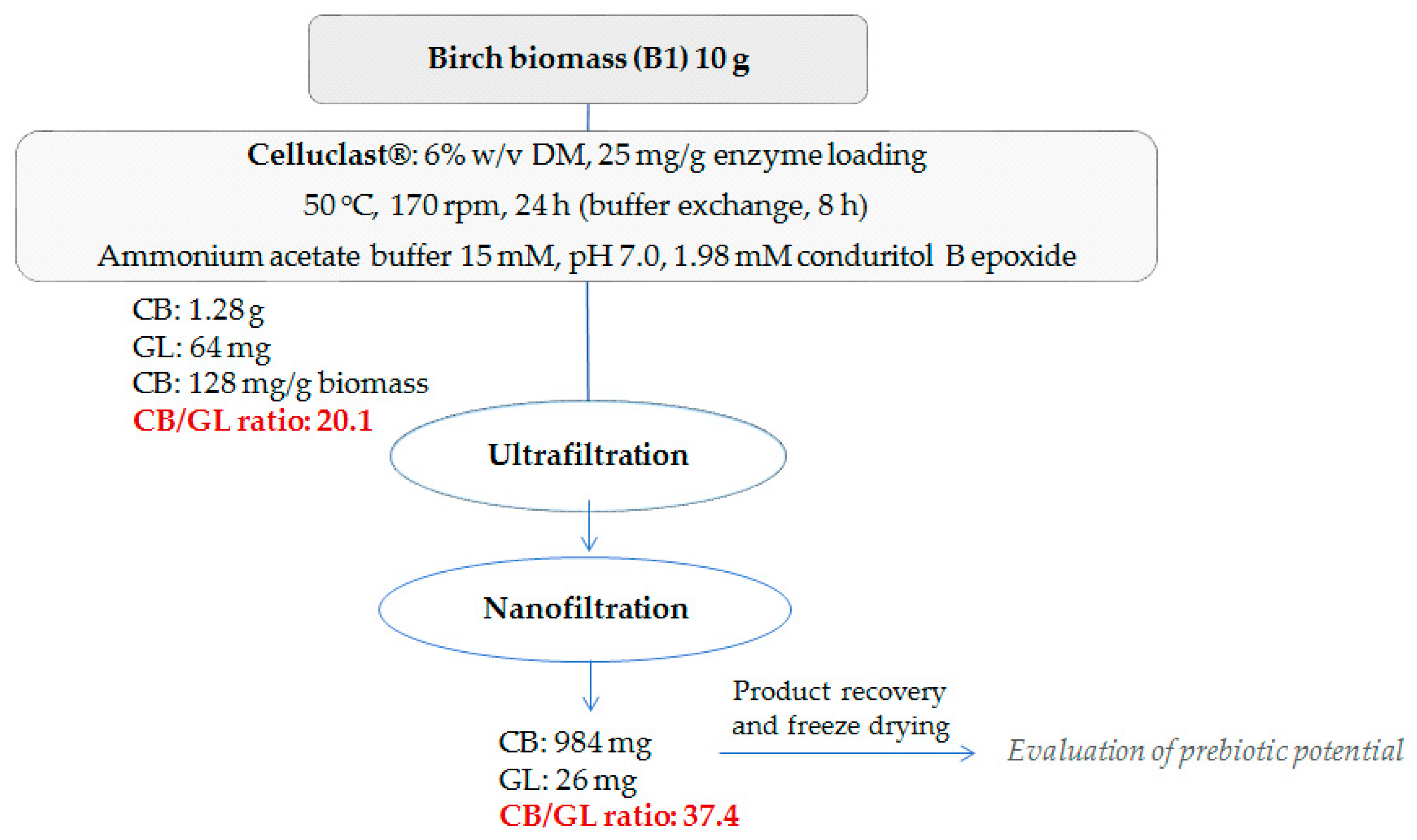
2.8. Scale-up Reaction and Downstream Processing for Product Recovery
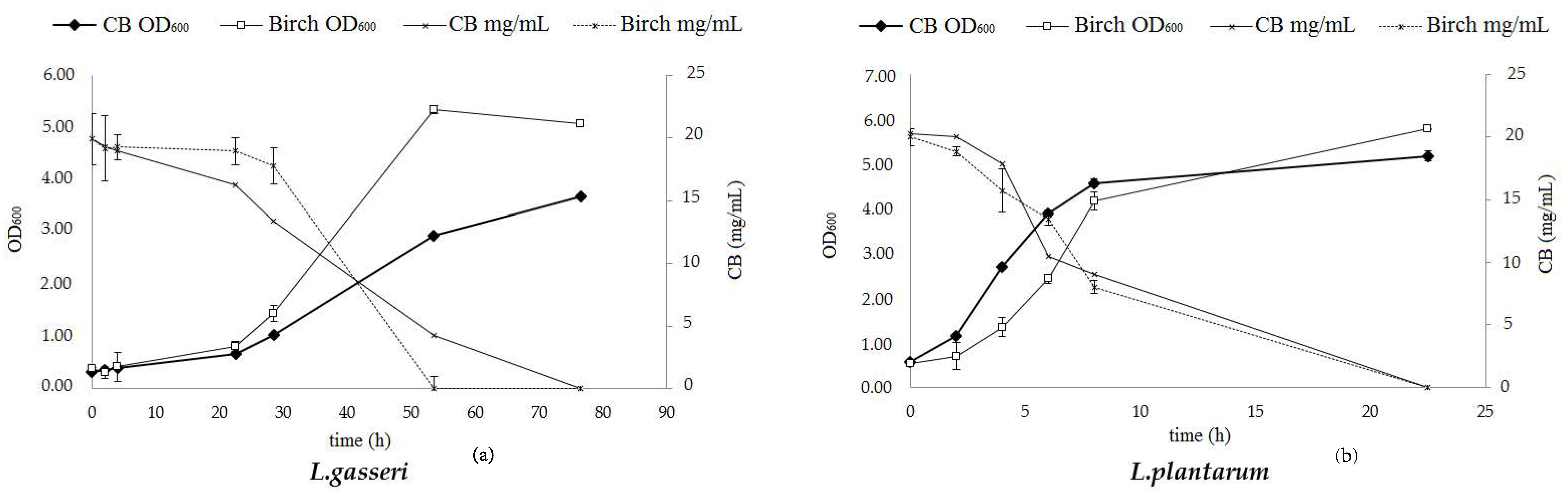
2.9. Evaluation of COS Prebiotic Activity
Growth Potential of Lactobacillus Strains on Pure Cellobiose and Birch-Derived Sugars
3. Discussion
4. Materials and Methods
4.1. Enzymes and Substrates
4.2. Hydrolysis of Lignocellulosic Materials
4.3. Evaluation of Synergistic Effect of PcLPMO9D with Cellulases towards Cellobiose Production
4.4. Scale-up Hydrolysis Reaction and Product Recovery
4.5. Determination of Prebiotic Potential of Birch Hydrolysate
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Gibson, G.R.; Roberfroid, M.B. Dietary modulation of the human colonic microbiota: Introducing the concept of prebiotics. J. Nutr. 1995, 125, 1401–1412. [Google Scholar] [CrossRef] [PubMed]
- Roberfroid, M. Prebiotics: The concept revisited. J. Nutr. 2007, 137, 830S–837S. [Google Scholar] [CrossRef] [PubMed]
- Swennen, K.; Courtin, C.M.; Delcour, J.A. Non-digestible oligosaccharides with prebiotic properties. Crit. Rev. Food Sci. Nutr. 2006, 46, 459–471. [Google Scholar] [CrossRef] [PubMed]
- Belorkar, S.A.; Gupta, A.K. Oligosaccharides: A boon from nature’s desk. AMB Express 2016, 6, 82. [Google Scholar] [CrossRef] [PubMed]
- Mussatto, S.I.; Mancilha, I.M. Non-digestible oligosaccharides: A review. Carbohydr. Polym. 2007, 68, 587–597. [Google Scholar] [CrossRef]
- Pokusaeva, K.; O’Connell-Motherway, M.; Zomer, A.; MacSharry, J.; Fitzgerald, G.F.; van Sinderen, D. Cellodextrin utilization by Bifidobacterium breve UCC2003. Appl. Environ. Microbiol. 2011, 77, 1681–1690. [Google Scholar] [CrossRef]
- Nakamura, S.; Oku, T.; Ichinose, M. Bioavailability of cellobiose by tolerance test and breath hydrogen excretion in humans. Nutrition 2004, 20, 979–983. [Google Scholar] [CrossRef]
- Basholli-Salihu, M.; Mueller, M.; Unger, F.M.; Viernstein, H. The use of cellobiose and fructooligosaccharide on growth and stability of Bifidobacterium infantis in fermented milk. Food Nutr. Sci. 2013, 4, 1301. [Google Scholar] [CrossRef]
- Satouchi, M.; Watanabe, T.; Wakabayashi, S.; Ohokuma, K.; Koshijima, T.; Kuwahara, M. Digestibility, absorptivity and physiological effects of cellooligosaccharides in human and rat. Nippon Eiyo Shokuryo Gakkaishi 1996, 49, 143–148. [Google Scholar] [CrossRef]
- Watanabe, T. Development of physiological functions of cellooligosaccharides. Cellul. Commun. 1998, 5, 91–97. [Google Scholar]
- Karnaouri, A.; Topakas, E.; Matsakas, L.; Rova, U.; Christakopoulos, P. Fine-tuned enzymatic hydrolysis of organosolv pretreated forest materials for the efficient production of cellobiose. Front. Chem 2018, 6, 128. [Google Scholar] [CrossRef] [PubMed]
- Vanderghem, C.; Boquel, P.; Blecker, C.; Paquot, M. A multistage process to enhance cellobiose production from cellulosic materials. Appl. Biochem. Biotechnol. 2010, 160, 2300–2307. [Google Scholar] [CrossRef] [PubMed]
- Tsuji, A.; Tominaga, K.; Nishiyama, N.; Yuasa, K. Comprehensive enzymatic analysis of the cellulolytic system in digestive fluid of the Sea Hare Aplysia kurodai. Efficient glucose release from sea lettuce by synergistic action of 45 kDa endoglucanase and 210 kDa ß-glucosidase. PLoS ONE 2013, 8, e65418. [Google Scholar] [CrossRef] [PubMed]
- Kuo, C.L.; Kallemeijn, W.W.; Lelieveld, L.T.; Mirzaian, M.; Zoutendijk, I.; Vardi, A.; Futerman, A.H.; Meijer, A.H.; Spaink, H.P.; Overkleeft, H.S.; et al. In vivo inactivation of glycosidases by conduritol B epoxide and cyclophellitol as revealed by activity-based protein profiling. FEBS J. 2019, 286, 3. [Google Scholar] [CrossRef] [PubMed]
- Witte, M.D.; van der Marel, G.A.; Aerts, J.M.; Overkleeft, H.S. Irreversible inhibitors and activity-based probes as research tools in chemical glycobiology. Org. Biomol. Chem. 2011, 9, 5908–5926. [Google Scholar] [CrossRef] [Green Version]
- Rodrigues, A.C.; Haven, M.Ø.; Lindedam, J.; Felby, C.; Gama, M. Celluclast and Cellic® CTec2: Saccharification/fermentation of wheat straw, solid-liquid partition and potential of enzyme recycling by alkaline washing. Enzym. Microb. Technol. 2015, 79–80, 70–77. [Google Scholar] [CrossRef]
- Hu, J.; Arantes, V.; Pribowo, A.; Gourlay, K.; Saddler, J. Substrate factors that influence the synergistic interaction of AA9 and cellulases during the enzymatic hydrolysis of biomass. Energy Environ. Sci. 2014, 7, 2308–2315. [Google Scholar] [CrossRef]
- Matsakas, L.; Nitsos, C.; Raghavendran, V.; Yakimenko, O.; Persson, G.; Olsson, E.; Rova, U.; Olsson, L.; Christakopoulos, P. A novel hybrid organosolv: Steam explosion method for the efficient fractionation and pretreatment of birch biomass. Biotechnol. Biofuels 2018, 11, 1–14. [Google Scholar] [CrossRef]
- Matsakas, L.; Raghavendran, V.; Yakimenko, O.; Persson, G.; Olsson, E.; Rova, U.; Olsson, L.; Christakopoulos, P. Lignin-first biomass fractionation using a hybrid organosolv—Steam explosion pretreatment technology improves the saccharification and fermentability of spruce biomass. Bioresour. Technol. 2019, 273, 521–528. [Google Scholar] [CrossRef]
- Umezurike, G.M. The mechanism of action of beta-glucosidase from Botryodiplodia theobromae Pat. Biochem. J. 1987, 241, 455–462. [Google Scholar] [CrossRef]
- Willför, S.; Sundberg, A.; Hemming, J.; Holmbom, B. Polysaccharides in some industrially important softwood species. Wood Sci. Technol. 2005, 39, 245–257. [Google Scholar] [CrossRef]
- Jiao, L.F.; Ke, Y.L.; Xiao, K.; Song, Z.H.; Hu, C.H.; Shi, B. Effects of cello-oligosaccharide on intestinal microbiota and epithelial barrier function of weanling pigs. J. Anim. Sci. 2015, 93, 1157–1164. [Google Scholar] [CrossRef] [PubMed]
- Mansfield, S.D.; Mooney, C.; Saddler, J.N. Substrate and enzyme characteristics that limit cellulose hydrolysis. Biotechnol. Prog. 1999, 15, 804–816. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Zhang, L.; Liu, D. Biomass recalcitrance. Part I: The chemical compositions and physical structures affecting the enzymatic hydrolysis of lignocellulose. Biofuels Bioprod. Biorefin. 2012, 6, 465–482. [Google Scholar] [CrossRef]
- Ajisaka, K.; Nishida, H.; Fujimoto, H. The synthesis of oligosaccharides by the reversed hydrolysis reaction of beta-glucosidase at high substrate concentration and at high temperature. Biotechnol. Lett 1987, 9, 243–248. [Google Scholar] [CrossRef]
- Bucke, C. Oligosaccharide synthesis using glycosidases. J. Chem. Technol. Biotechnol. 1996, 67, 217–220. [Google Scholar] [CrossRef]
- Murphy, L.; Bohlin, C.; Baumann, M.J.; Olsen, S.N.; Sørensen, T.H.; Anderson, L.; Borch, K.; Westh, P. Product inhibition of five Hypocrea jecorina cellulases. Enzym. Microb. Technol 2009, 52, 163–169. [Google Scholar] [CrossRef]
- Qing, Q.; Yang, B.; Wyman, C.E. Xylooligomers are strong inhibitors of cellulose hydrolysis by enzymes. Bioresour. Technol. 2010, 101, 9624. [Google Scholar] [CrossRef]
- Modenbach, A.A.; Nokes, S.E. Enzymatic hydrolysis of biomass at high-solids loadings—A review. Biosyst. Agric. Eng. Fac. Publ. 2013. [Google Scholar] [CrossRef]
- Herlet, J.; Kornberger, P.; Roessler, B.; Glanz, J.; Schwarz, W.H.; Liebl, W.; Zverlov, V.V. A new method to evaluate temperature vs. pH activity profiles for biotechnological relevant enzymes. Biotechnol. Biofuels 2017, 10, 234. [Google Scholar] [CrossRef]
- Qi, B.; Luo, J.; Chen, G.; Chen, X.; Wan, Y. Application of ultrafiltration and nanofiltration for recycling cellulase and concentrating glucose from enzymatic hydrolyzate of steam exploded wheat straw. Bioresour. Technol. 2012, 104, 466–472. [Google Scholar] [CrossRef] [PubMed]
- Villares, A.; Moreau, C.; Bennati-Granier, C.; Garajova, S.; Foucat, L.; Falourd, X.; Saake, B.; Berrin, J.G.; Cathala, B. Lytic polysaccharide monooxygenases disrupt the cellulose fibers structure. Sci. Rep. 2017, 7, 40262. [Google Scholar] [CrossRef] [PubMed]
- Nitsos, C.; Stoklosa, R.; Karnaouri, A.; Vörös, D.; Lange, H.; Hodge, D.; Crestini, C.; Rova, U.; Christakopoulos, P. Isolation and characterization of organosolv and alkaline lignins from hardwood and softwood biomass. ACS Sustain. Chem. Eng. 2016, 4, 5181–5193. [Google Scholar] [CrossRef]
- Raghavendran, V.; Nitsos, C.; Matsakas, L.; Rova, U.; Christakopoulos, P.; Olsson, L. A comparative study of the enzymatic hydrolysis of batch organosolv-pretreated birch and spruce biomass. AMB Express 2018, 8, 114. [Google Scholar] [CrossRef]
- Karnaouri, A.; Rova, U.; Christakopoulos, P. Effect of different pretreatment methods on birch outer bark: New biorefinery routes. Molecules 2016, 21, 427. [Google Scholar] [CrossRef] [PubMed]
- Karnaouri, A.; Muraleedharan, M.N.; Dimarogona, M.; Topakas, E.; Rova, U.; Sandgren, M.; Christakopoulos, P. Recombinant expression of thermostable processive MtEG5 endoglucanase and its synergism with MtLPMO from Myceliophthora thermophila during the hydrolysis of lignocellulosic substrates. Biotechnol. Biofuels 2017, 10, 126. [Google Scholar] [CrossRef] [PubMed]
- Westereng, B.; Ishida, T.; Vaaje-Kolstad, G.; Wu, M.; Eijsink, V.G.; Igarashi, K.; Samejima, M.; Ståhlberg, J.; Horn, S.J.; Sandgren, M. The putative endoglucanase PcGH61D from Phanerochaete chrysosporium is a metal-dependent oxidative enzyme that cleaves cellulose. PLoS ONE 2011, 6, e27807. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 24 h | 48 h | |||||
|---|---|---|---|---|---|---|
| CB (% w/w) | Glu (% w/w) | CB:Glu | CB (% w/w) | Glu (% w/w) | CB:Glu | |
| No preincubation | 17.7 ± 0.9 | 2.7 ± 0.1 | 6.7 | 19.5 ± 1.1 | 3.0 ± 1.0 | 6.5 |
| Preincubation | 17.0 ± 1.1 | 2.9 ± 0.0 | 5.8 | 19.9 ± 1.0 | 3.4 ± 1.1 | 5.8 |
| Temperature/Incubation Time | CB | Glu | CB:Glu | mg CB/g Substrate |
|---|---|---|---|---|
| 50 °C, 12 h | 13.0 ± 1.1 | 0.5 ± 0.1 | 27.1 | 111.97 |
| 50 °C, 24 h | 16.3 ± 0.5 | 0.6 ± 0.1 | 28.0 | 140.66 |
| 45 °C, 12 h | 16.7 ± 0.7 | 1.4 ± 0.1 | 12.1 | 143.97 |
| 45 °C, 24 h | 20.1 ± 0.4 | 1.2 ± 0.0 | 17.3 | 173.45 |
| 40 °C, 12 h | 12.8 ± 1.0 | 1.3 ± 0.2 | 9.6 | 110.64 |
| 40 °C, 24 h | 18.5 ± 0.2 | 1.7 ± 0.2 | 10.7 | 159.4 |
| Conditions | CB (% w/w) | Glu (% w/w) | CB:Glu | mg CB/g Substrate | % CB Increase | % Glu Increase |
|---|---|---|---|---|---|---|
| 25 mg enzyme/g sub | 16.0 ± 1.7 | 0.7 ± 0.1 | 21.8 | 145.7 | 0 | 0 |
| 25 mg enzyme/g sub + 2.5 mg PcLPMO9D/g sub | 24.3 ± 1.2 | 1.9 ± 0.1 | 13.1 | 220.9 | 51.5 | 151.6 |
| 27.5 mg enzyme/g sub | 16.0 ± 1.0 | 0.7 ± 1.0 | 21.8 | 145.4 | 0 | 0 |
| L. gasseri | L. plantarum | |||||||
|---|---|---|---|---|---|---|---|---|
| Cellobiose | Birch Hydrolysate | Cellobiose | Birch Hydrolysate | |||||
| 0 h | 77 h | 0 h | 77 h | 0 h | 23 h | 0 h | 23 h | |
| cellobiose | 19.7 ± 1.1 | 0.1 ± 0.0 | 19.6 ± 2.0 | 0.0 ± 0.00 | 20.1 ± 0.06 | 0.0 ± 0.0 | 20.0 ± 2.6 | 0.1 ± 0.1 |
| lactic acid | 0.1 ± 0.1 | 8.5 ± 1.4 | 0.3 ± 0.1 | 39.5 ± 3.9 | 0.4 ± 0.01 | 21.1 ± 2.9 | 1.2 ± 0.2 | 37.1 ± 2.9 |
| acetic acid | 4.0 ± 1.3 | 4.1 ± 0.0 | 4.2 ± 0.8 | 4.4 ± 1.0 | 4.3 ± 0.25 | 3.8 ± 0.6 | 4.5 ± 1.7 | 4.4 ± 1.9 |
| propionic acid | 0.9 ± 0.0 | 1.03 ± 0.7 | 1.1 ± 0.0 | 0.8 ± 0.2 | 0.9 ± 0.12 | 0.9 ± 0.0 | 1.1 ± 0.7 | 0.8 ± 0.2 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Karnaouri, A.; Matsakas, L.; Bühler, S.; Muraleedharan, M.N.; Christakopoulos, P.; Rova, U. Tailoring Celluclast® Cocktail’s Performance towards the Production of Prebiotic Cello-Oligosaccharides from Waste Forest Biomass. Catalysts 2019, 9, 897. https://doi.org/10.3390/catal9110897
Karnaouri A, Matsakas L, Bühler S, Muraleedharan MN, Christakopoulos P, Rova U. Tailoring Celluclast® Cocktail’s Performance towards the Production of Prebiotic Cello-Oligosaccharides from Waste Forest Biomass. Catalysts. 2019; 9(11):897. https://doi.org/10.3390/catal9110897
Chicago/Turabian StyleKarnaouri, Anthi, Leonidas Matsakas, Saskja Bühler, Madhu Nair Muraleedharan, Paul Christakopoulos, and Ulrika Rova. 2019. "Tailoring Celluclast® Cocktail’s Performance towards the Production of Prebiotic Cello-Oligosaccharides from Waste Forest Biomass" Catalysts 9, no. 11: 897. https://doi.org/10.3390/catal9110897
APA StyleKarnaouri, A., Matsakas, L., Bühler, S., Muraleedharan, M. N., Christakopoulos, P., & Rova, U. (2019). Tailoring Celluclast® Cocktail’s Performance towards the Production of Prebiotic Cello-Oligosaccharides from Waste Forest Biomass. Catalysts, 9(11), 897. https://doi.org/10.3390/catal9110897