

Study of the Interaction of Sorption and Catalytic Centers in Carboxypeptidase T by X-ray Analysis


Abstract
:1. Introduction
2. Materials and Methods
2.1. Macromolecule Production
2.2. Crystallization
2.3. Data Collection and Processing
2.4. Structure Solution and Refinement
2.5. Kinetic Experiments
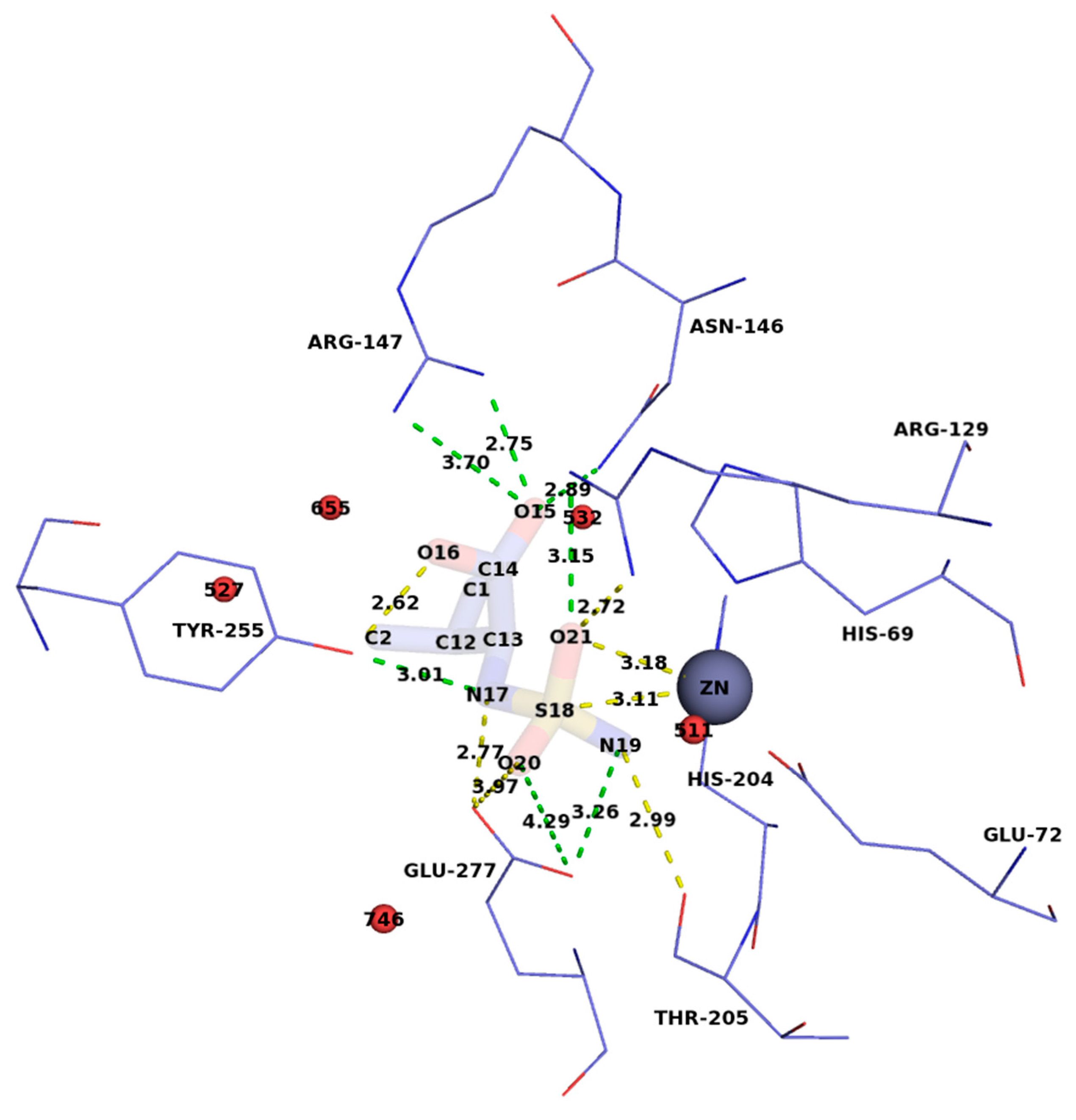
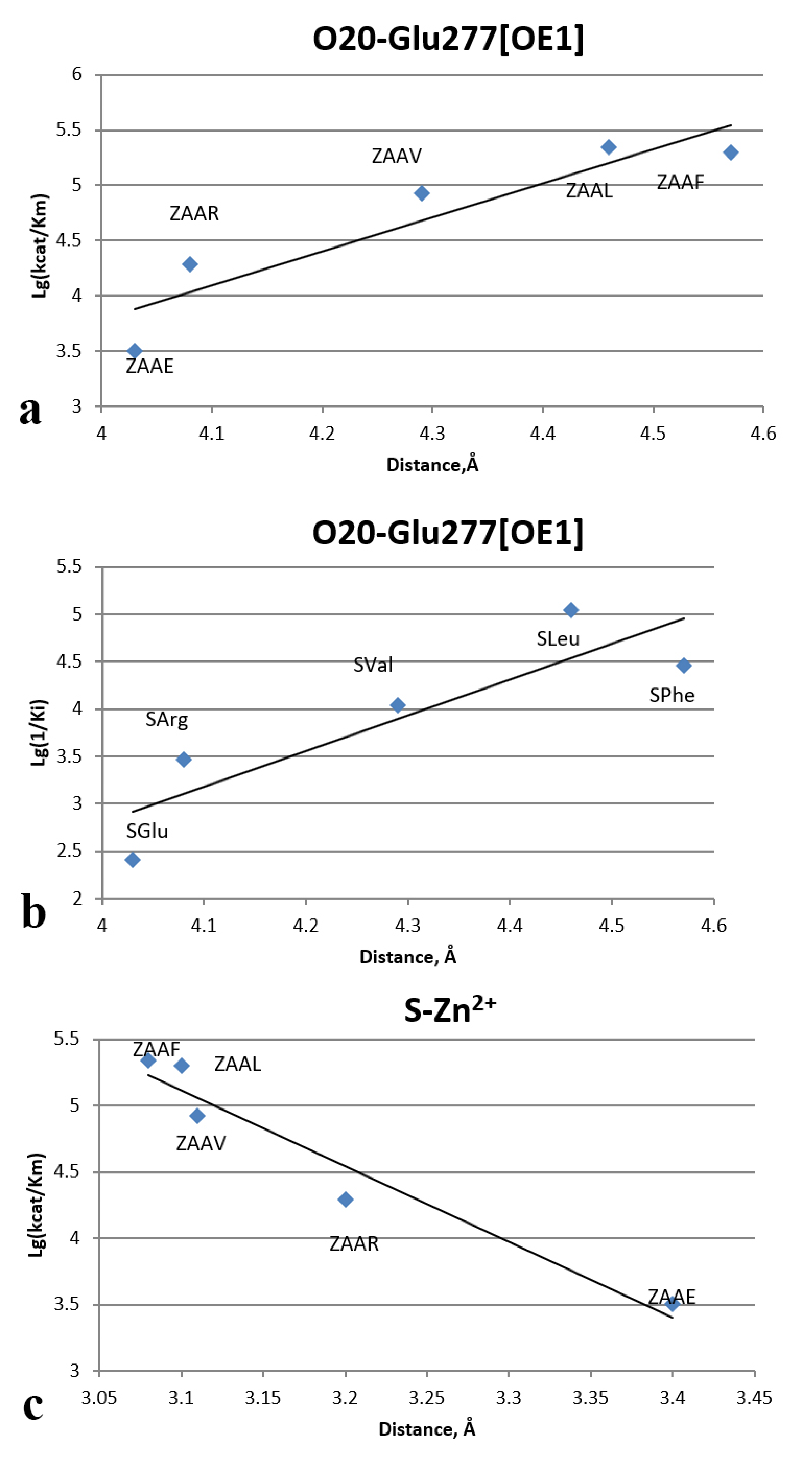
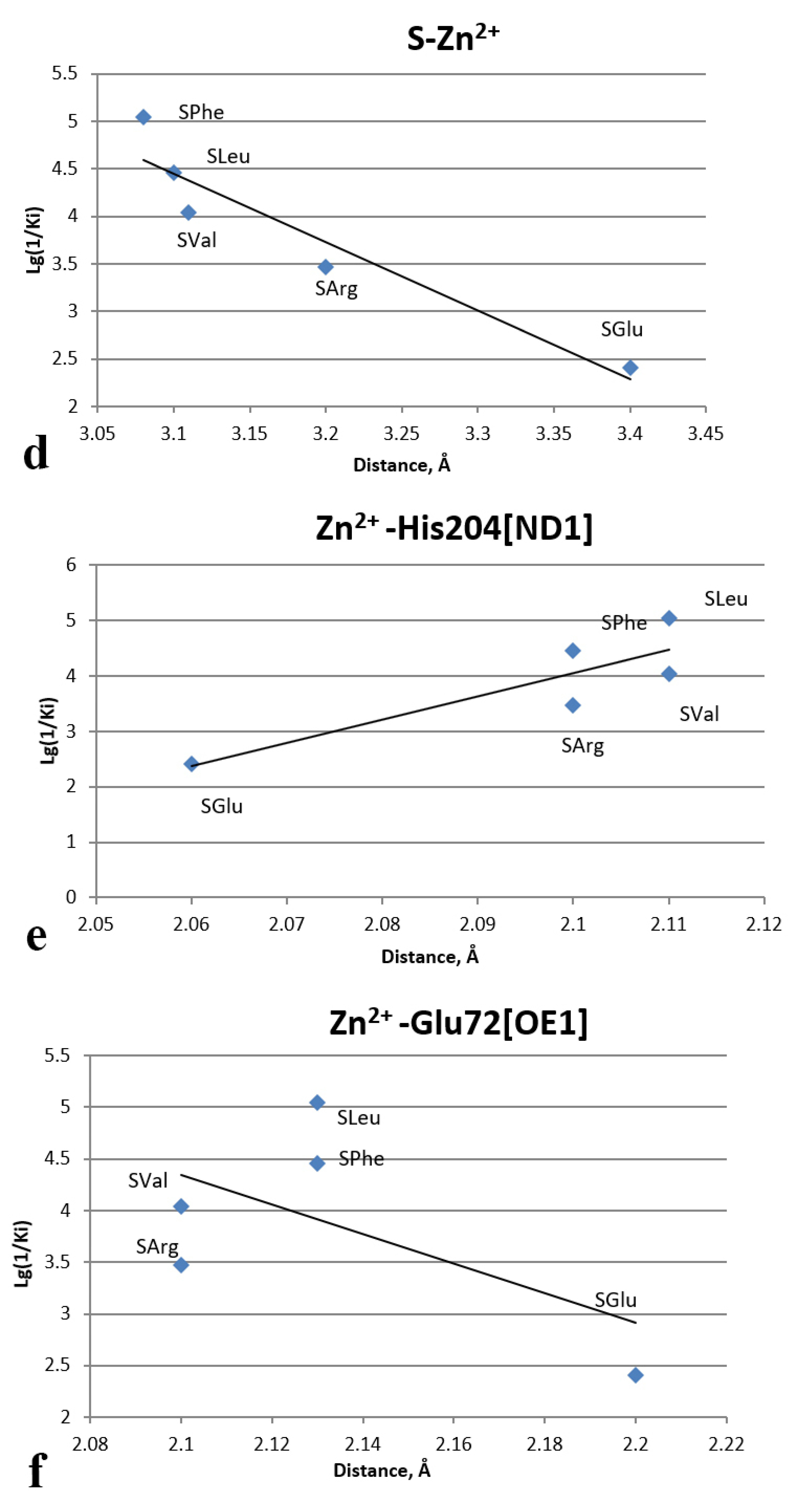
3. Results and Discussion
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Turk, B.; Turk, D.; Turk, V. Protease signalling: The cutting edge. EMBO J. 2012, 31, 1630–1643. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Silva, J.G.; Español, Y.; Velasco, G.; Quesada, V. The Degradome database: Expanding roles of mammalian proteases in life and disease. Nucleic Acids Res. 2016, 44, D351–D355. [Google Scholar] [CrossRef] [PubMed]
- Bunnage, M.E.; Blagg, J.; Steele, J.; Owen, D.R.; Allerton, C.; McElroy, A.B.; Miller, D.; Ringer, T.; Butcher, K.; Beaumont, K.; et al. Discovery of potent & selective inhibitors of activated thrombin-activatable fibrinolysis inhibitor for the treatment of thrombosis. J. Med. Chem. 2007, 50, 6095–6103. [Google Scholar]
- Vallee, B.L.; Neurath, H. Carboxypeptidase, a zinc metalloenzyme. J. Biol. Chem. 1955, 217, 253–262. [Google Scholar] [CrossRef]
- Christianson, D.W.; Lipscomb, W.N. Carboxypeptidase A. Acc. Chem. Res. 1989, 22, 62–69. [Google Scholar] [CrossRef]
- Lipscomb, W.M.; Strater, N. Recent Advances in Zinc Enzymology. Chem. Rev. 1996, 96, 2375–2433. [Google Scholar] [CrossRef]
- Schechter, I.; Berger, A. On the size of the active site in proteases. I. Papain. 1967. Biochem. Biophys Res. Commun. 2012, 425, 497–502. [Google Scholar] [CrossRef] [PubMed]
- Bown, D.P.; Gatehouse, J.A. Characterization of a digestive carboxypeptidase from the insect pest corn earworm (Helicoverpa armigera) with novel specificity towards C-terminal glutamate residues. Eur. J. Biochem. 2004, 271, 2000–2011. [Google Scholar] [CrossRef]
- Edge, M.; Forder, C.; Hennam, J.; Lee, I.; Tonge, D.; Hardern, I.; Fitton, J.; Eckersley, K.; East, S.; Blakey, D.; et al. Engineered human carboxypeptidase B enzymes that hydrolyse hippuryl-L-glutamic acid: Reversed-polarity mutants. Protein Eng. 1998, 11, 1229–1234. [Google Scholar] [CrossRef]
- Garcia-Guerrero, M.C.; Garcia-Pardo, J.; Berenguer, E.; Fernandez-Alvarez, R.; Barfi, G.B.; Lyons, P.J.; Aviles, F.X.; Huber, R.; Lorenzo, J.; Reverter, D. Crystal structure and mechanism of human carboxypeptidase O: Insights into its specific activity for acidic residues. Proc. Natl. Acad. Sci. USA 2018, 115, E3932–E3939. [Google Scholar] [CrossRef] [Green Version]
- Teplyakov, A.; Polyakov, K.; Obmolova, G.; Strokopytov, B.; Kuranova, I.; Osterman, A.L.; Grishin, N.; Smulevitch, S.; Zagnitko, O.; Galperina, O.; et al. Crystal structure of carboxypeptidase T from Thermoactinomyces vulgaris. Eur. J. Biochem. 1992, 208, 281–288. [Google Scholar] [CrossRef]
- Osterman, A.L.; Stepanov, V.M.; Rudenskaia, G.N.; Khodova, O.M.; Tsaplina, I.A. Carboxypeptidase T—intracellular carboxypeptidase of Thermoactinomycetes—A distant analog of animal carboxypeptidase. Biokhimiia 1984, 49, 292–301. [Google Scholar] [PubMed]
- Grishin, A.M.; Akparov, V.K.; Chestukhina, G.G. Structural principles of the broad substrate specificity of Thermoactinomyces vulgaris carboxypeptidase T—Role of amino acid residues at positions 260 and 262. Protein Eng. Des. Sel. Engl. 2008, 21, 545–551. [Google Scholar] [CrossRef] [Green Version]
- Akparov, V.K.; Grishin, A.M.; Yusupova, M.P.; Ivanova, N.M.; Chestukhina, G.G. Structural principles of the wide substrate specificity of Thermoactinomyces vulgaris carboxypeptidase T. Reconstruction of the carboxypeptidase B primary specificity pocket. Biochemistry 2007, 72, 416–423. [Google Scholar] [CrossRef]
- Akparov, V.; Timofeev, V.; Khaliullin, I.; Švedas, V.; Kuranova, I.; Švedas, V. Structure of the carboxypeptidase B complex with N-sulfamoyl-L-phenylalanine—a transition state analog of non-specific substrate. J. Biomol. Struct. Dyn. Engl. 2018, 36, 956–965. [Google Scholar] [CrossRef] [PubMed]
- Akparov, V.K.; Timofeev, V.I.; Konstantinova, G.E.; Khaliullin, I.G.; Kuranova, I.P.; Rakitina, T.V.; Švedas, V. The nature of the ligand’s side chain interacting with the S1’-subsite of metallocarboxypeptidase T (from Thermoactinomyces vulgaris) determines the geometry of the tetrahedral transition complex. PLoS ONE 2019, 14, e0226636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okai, M.; Yamamura, A.; Hayakawa, K.; Tsutsui, S.; Miyazono, K.I.; Lee, W.C.; Nagata, K.; Inoue, Y.; Tanokura, M. Insight into the transition between the open and closed conformations of Thermus thermophilus carboxypeptidase. Biochem. Biophys Res. Commun. 2017, 484, 787–793. [Google Scholar] [CrossRef]
- Akparov, V.K.; Belianova, L.P.; Baratova, L.A.; Stepanov, V.M. Subtilisin 72: A serine protease from Bac. subtilis strain 72—An enzyme similar to subtilisin Carlsberg. Biokhimiia 1979, 44, 886–891. [Google Scholar]
- Liublinskaia, L.A.; Iakushcheva, L.D.; Stepanov, V.M. The synthesis of the peptide substrates of the subtilisin and its analogs. Bioorg. Khim. 1977, 3, 273–279. [Google Scholar]
- Voiushina, T.L.; Liublinskaia, L.A.; Timokhina, E.A.; Stepanov, V.M. Synthesis of p-nitroanilides of acylated peptides catalyzed by thermolysin. Bioorg. Khim. 1987, 13, 615–622. [Google Scholar]
- Cueni, L.B.; Bazzone, T.J.; Riordan, J.F.; Vallee, B.L. Affinity chromatographic sorting of carboxypeptidase A and its chemically modified derivatives. Anal. Biochem. 1980, 107, 341–349. [Google Scholar] [CrossRef]
- Novagen pET System Manual TB055, 7th ed.; Novagen: Madison, WI, USA, 1997.
- Takahashi, S.; Tsurumura, T.; Aritake, K.; Furubayashi, N.; Sato, M.; Yamanaka, M.; Hirota, E.; Sano, S.; Kobayashi, T.; Tanaka, T.; et al. High-quality crystals of human haematopoietic prostaglandin D synthase with novel inhibitors. Acta Cryst. Sect. F Struct. Biol. Cryst. Commun. 2010, 66, 846–850. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Ruíz, J.M.; Gonzalez-Ramirez, A.L.; Gavira, J.A.; Otálora, F. Granada Crystallisation Box: A new device for protein crystallisation by counter-diffusion techniques. Acta Crystallogr. Sect. D Biol. Crystallogr. 2002, 58, 1638–1642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takahashi, S.; Ohta, K.; Furubayashi, N.; Yan, B.; Koga, M.; Wada, Y.; Yamada, M.; Inaka, K.; Tanaka, H.; Miyoshi, H.; et al. JAXA protein crystallization in space: Ongoing improvements for growing high-quality crystals. J. Synchrotron Radiat. Int. Union Crystallogr. 2013, 20, 968–973. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, S.; Koga, M.; Yan, B.; Furubayashi, N.; Kamo, M.; Inaka, K.; Tanaka, H. JCB-SGT crystallization devices applicable to PCG experiments and their crystallization conditions. Int. J. Microgravity Sci. Appl. 2019, 36, 1–9. [Google Scholar]
- McPherson, A. Macromolecular crystal growth in microgravity. Crystallogr. Rev. 1996, 6, 157–308. [Google Scholar] [CrossRef]
- Battye, T.; Kontogiannis, L.; Johnson, O.; Powell, H. iMOSFLM: A new graphical interface for diffraction-image processing with MOSFLM. Leslie, AG. Acta Crystallogr. D Biol. Crystallogr. 2011, 67 Pt 4, 271–281. [Google Scholar] [CrossRef] [Green Version]
- McCoy, A.J.; Grosse-Kunstleve, R.W.; Adams, P.D.; Winn, M.D.; Storoni, L.C.; Read, R.J. Phaser crystallographic software. J. Appl. Crystallogr. 2007, 40, 658–674. [Google Scholar] [CrossRef] [Green Version]
- Murshudov, G.N.; Skubák, P.; Lebedev, A.A.; Pannu, N.S.; Steiner, R.A.; Nicholls, R.A.; Winn, M.D.; Long, F.; Vagin, A.A. REFMAC5 for the refinement of macromolecular crystal structures. Acta Crystallogr. Sect. D Biol. Crystallogr. 2011, 67, 355–367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Emsley, P.; Cowtan, K. Coot: Model-building tools for molecular graphics. Acta Crystallogr. Sect. D Biol. Crystallogr. 2004, 60, 2126–2132. [Google Scholar] [CrossRef] [Green Version]
- Park, J.D.; Kim, D.H.; Kim, S.J.; Woo, J.R.; Ryu, S.E. Sulfamide-based inhibitors for carboxypeptidase A. Novel type transition state analogue inhibitors for zinc proteases. J. Med. Chem. 2002, 45, 5295–5302. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Method | Counter-Diffusion Technique (in Microgravity) |
|---|---|
| Capillary type | Glass capillary |
| Temperature (K) | 293 |
| Protein concentration (gL−1) | 10 |
| Protein-containing solution (capillary) | 10 mM MES buffer pH 6.0, 0.25 M NaCl, 0.1 mM ZnSO4 and 1 mM CaCl2 |
| Precipitating solution (cylinder) | 1.4 M (NH4)2SO4, 50 mM MES pH 6.0, 0.1 mM ZnSO4, 1 mM CaCl2, 5% MPD, and 100 mM N-sulfamoyl-L-Valine |
| Diffraction Source | Spring8 Beamline BL41XU |
|---|---|
| Wavelength (Å) | 0.8 |
| Temperature (K) | 100 |
| Detector | DECTRIS EIGER X 16M |
| Crystal-detector distance (mm) | 300 |
| Rotation range per image (°) | 0.1 |
| Total rotation range (°) | 360 |
| Exposure time per image (s) | 0.1 |
| Space group | P6322 |
| a, b, c (Å) | 157.531, 157.531, 104,523 |
| α, β, γ (°) | 90, 90, 120 |
| Mosaicity (°) | 0.11 |
| Resolution range (Å) | 30.00–1.9 (2.0–1.9) |
| No. of unique reflections | 60,407 |
| Completeness (%) | 99.97 (99.89) |
| Redundancy | 17.49 (18.25) |
| 〈I/σ(I)〉 | 7.7316 (2.16) |
| Rmerge | 0.144 (0.330) |
| Resolution Range (Å) | 20–1.9 (1.95–1.9) |
|---|---|
| Completeness (%) | 99.9 (99.8) |
| σ cutoff | F > 0.000σ(F) |
| No. of reflections, working set | 57327 (4173) |
| No. of reflections, test set | 3001 (193) |
| Final Rcryst | 0.142 (0.156) |
| Final Rfree | 0.151 (0.168) |
| No. of non-H atoms | |
| Protein | 2581 |
| Ion | 6 |
| Ligand | 12 |
| Water | 262 |
| R.m.s. deviations | |
| Bonds (Å) | 0.014 |
| Angles (°) | 1.845 |
| Average B factor (Å2) | 13.76 |
| Ramachandran plot | |
| Favoured regions (%) | 98.0 |
| Additionally allowed (%) | 2.0 |
| Outliers (%) | 0.0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Akparov, V.; Timofeev, V.; Kuranova, I.; Khaliullin, I. Study of the Interaction of Sorption and Catalytic Centers in Carboxypeptidase T by X-ray Analysis. Crystals 2021, 11, 1088. https://doi.org/10.3390/cryst11091088
Akparov V, Timofeev V, Kuranova I, Khaliullin I. Study of the Interaction of Sorption and Catalytic Centers in Carboxypeptidase T by X-ray Analysis. Crystals. 2021; 11(9):1088. https://doi.org/10.3390/cryst11091088
Chicago/Turabian StyleAkparov, Valerij, Vladimir Timofeev, Inna Kuranova, and Ilias Khaliullin. 2021. "Study of the Interaction of Sorption and Catalytic Centers in Carboxypeptidase T by X-ray Analysis" Crystals 11, no. 9: 1088. https://doi.org/10.3390/cryst11091088
APA StyleAkparov, V., Timofeev, V., Kuranova, I., & Khaliullin, I. (2021). Study of the Interaction of Sorption and Catalytic Centers in Carboxypeptidase T by X-ray Analysis. Crystals, 11(9), 1088. https://doi.org/10.3390/cryst11091088