
A Short and Practical Overview on Light-Sensing Proteins, Optogenetics, and Fluorescent Biomolecules inside Biomorphs Used as Optical Sensors

 , , , , ,
, , , , ,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Protein Isolation and Purification
2.1.1. Phototropins
2.1.2. Green-Light and Red-Light Signaling Protein from Condylactis gigantea
2.2. Synthesis of Silica–Carbonate of Alkaline Metals (Biomorphs)
2.3. Characterization of Biomorphs
2.3.1. Scanning Electron Microscopy
2.3.2. Raman Spectroscopy
2.3.3. Confocal Microscopy
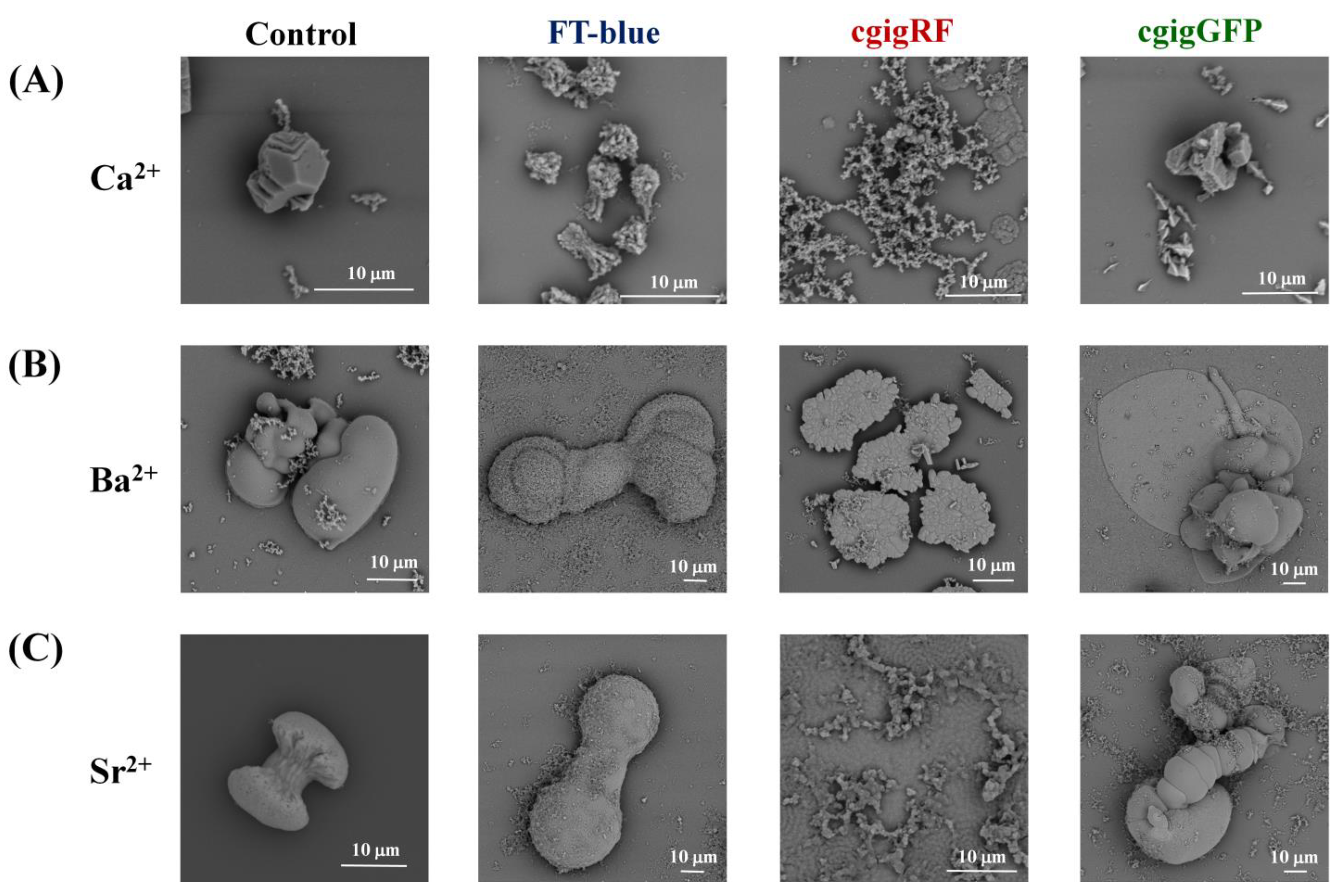
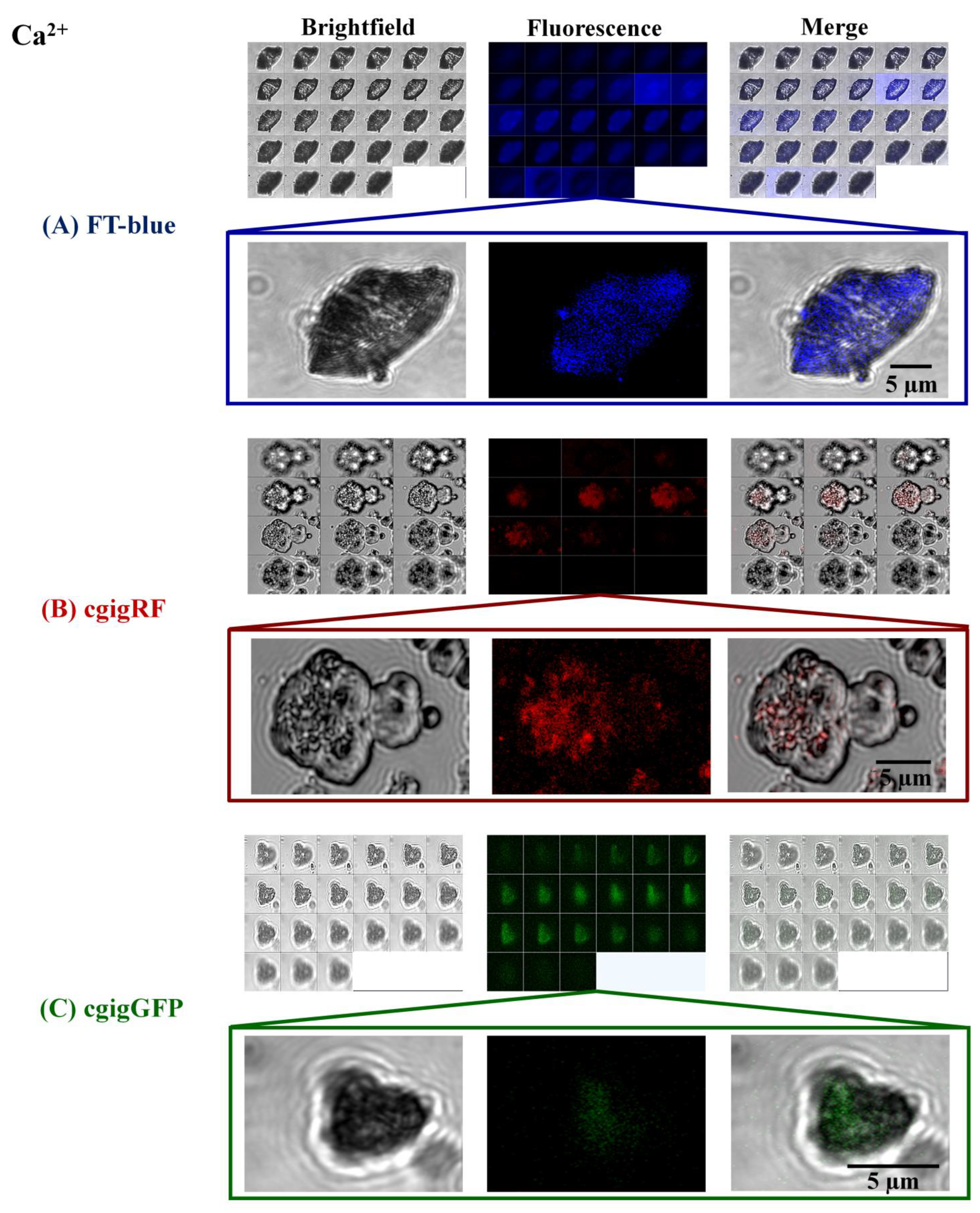
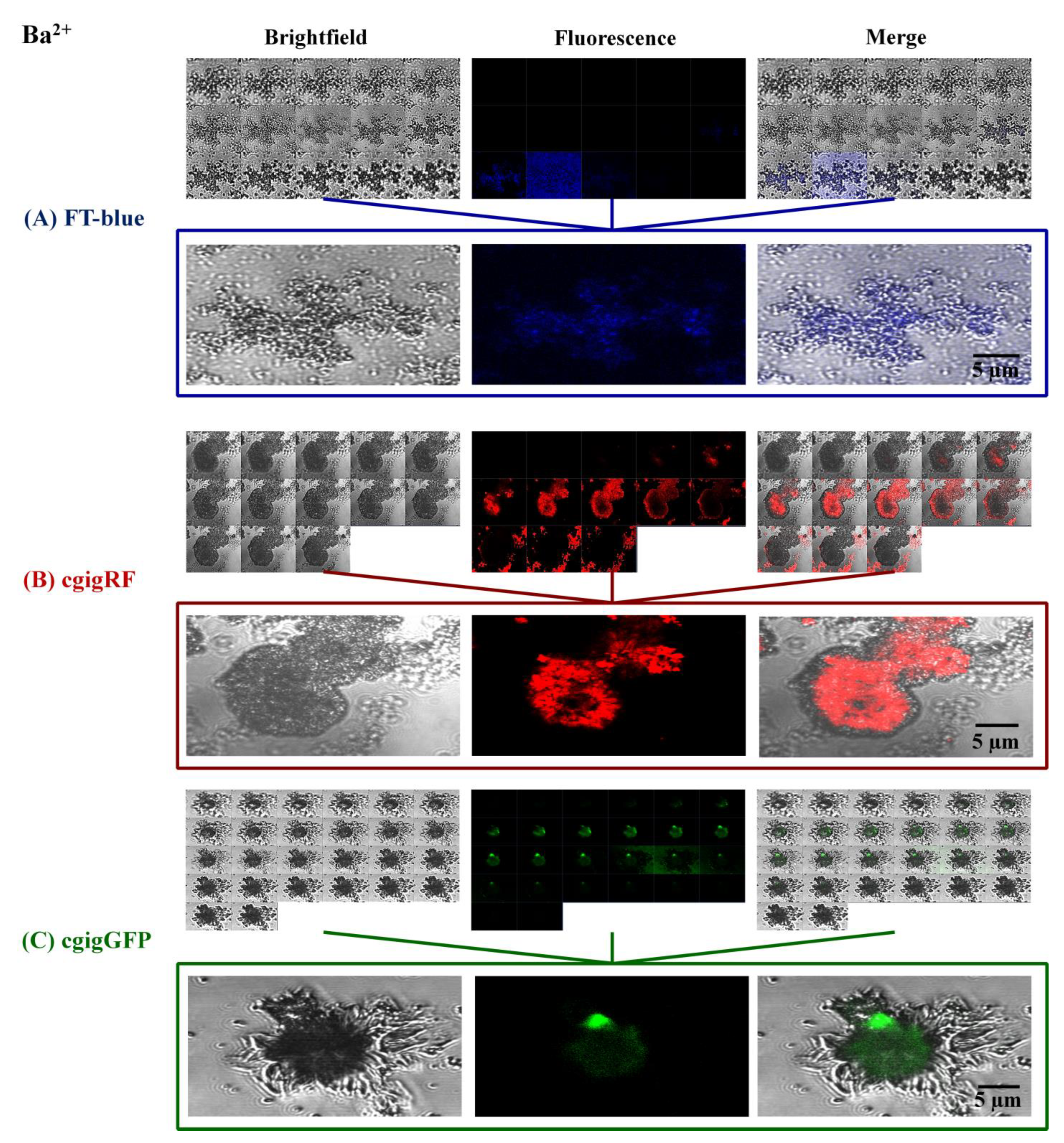
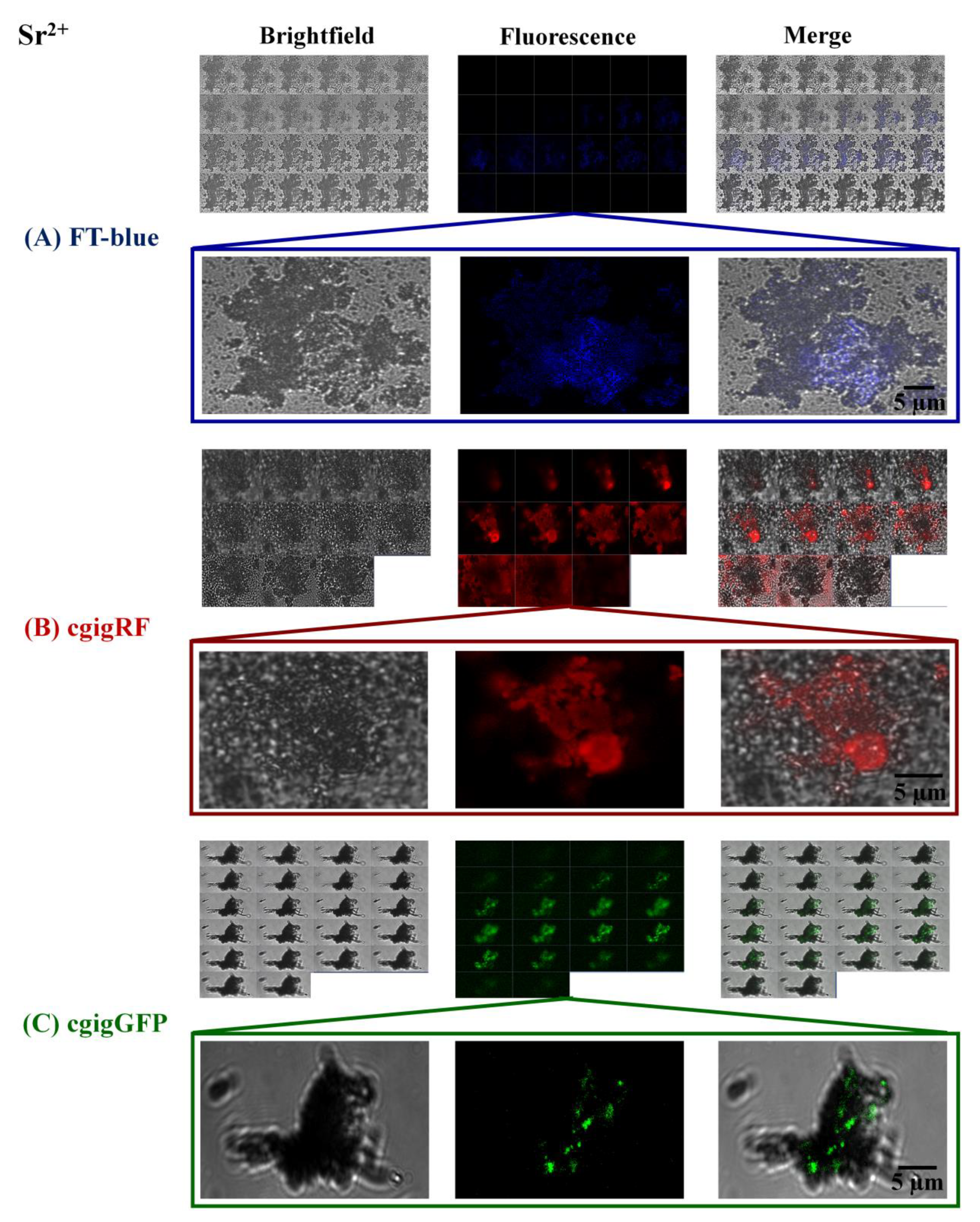
3. Results and Discussion
Proteins in Action, Working as Optical Sensors Inside Biomorphs
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Griffin, J.H.C.; Toledo-Ortiz, G. Plant photoreceptors and their signalling components in chloroplastic anterograde and retrograde communication. J. Exp. Bot. 2022, 73, 7126–7138. [Google Scholar] [CrossRef]
- Haga, K.; Sakai, T. Photosensory adaptation mechanisms in hypocotyl phototropism: How plants recognize the direction of a light source. J. Exp. Bot. 2023, 74, 1758–1769. [Google Scholar] [CrossRef]
- Kong, S.-G.; Okajima, K. Diverse photoreceptors and light responses in plants. J. Plant Res. 2016, 129, 111–114. [Google Scholar] [CrossRef]
- Chudakov, D.M.; Matz, M.V.; Lukyanov, S.; Lukyanov, K.A. Fluorescent Proteins and their applications in imaging living cells and tissues. Physiol. Rev. 2010, 90, 1103–1163. [Google Scholar] [CrossRef]
- Lambert, T.J. FPbase: A community-editable fluorescent protein database. Nat. Methods 2019, 16, 277–278. [Google Scholar] [CrossRef]
- Deo, S.K.; Daunert, S. Luminescent proteins from Aequorea victoria: Applications in drug discovery and in high throughput analysis. Anal. Bioanal. Chem. 2001, 369, 258–266. [Google Scholar] [CrossRef]
- Aihara, Y.; Maruyama, S.; Baird, A.H.; Iguchi, A.; Takahashi, S.; Minagawa, J. Green fluorescence from cnidarian hosts attracts symbiotic algae. Proc. Natl. Acad. Sci. USA 2019, 116, 2118–2123. [Google Scholar] [CrossRef]
- Kusmita, L.; Edi, A.N.P.; Franyoto, Y.D.; Mutmainah; Haryanti, S.; Nurcahyanti, A.D.R. Sun protection and antibacterial activities of carotenoids from the soft coral Sinularia sp. symbiotic bacteria from Panjang Island, North Java Sea. Saudi Pharm. J. 2023, 31, 101680. [Google Scholar] [CrossRef]
- Casal, J.J. Phytochromes, cryptochromes, phototropin: Photoreceptor interactions in plants. Photochem. Photobiol. 2000, 71, 1–11. [Google Scholar] [CrossRef]
- Cheng, M.-C.; Kathare, P.K.; Paik, I.; Huq, E. Phytochrome signaling networks. Annu. Rev. Plant Biol. 2021, 72, 217–244. [Google Scholar] [CrossRef]
- Briggs, W.R.; Huala, E. Blue-Light Photoreceptors in Higher Plants. Annu. Rev. Cell Dev. Biol. 1999, 15, 33–62. [Google Scholar] [CrossRef]
- Cordeiro, A.M.; Andrade, L.; Monteiro, C.C.; Leitão, G.; Wigge, P.A.; Saibo, N.J.M. Phytochrome-interacting factors: A promising tool to improve crop productivity. J. Exp. Bot. 2022, 73, 3881–3897. [Google Scholar] [CrossRef]
- Li, F.-W.; Mathews, S. Evolutionary aspects of plant photoreceptors. J. Plant Res. 2016, 129, 115–122. [Google Scholar] [CrossRef]
- Villafani, Y.; Yang, H.W.; Park, Y.-I. Color sensing and signal transmission diversity of cyanobacterial phytochromes and cyanobacteriochromes. Mol. Cells 2020, 43, 509–516. [Google Scholar]
- Kami, C.; Lorrain, S.; Hornitschek, P.; Fankhauser, C. Light-regulated plant growth and development. Curr. Top. Dev. Biol. 2010, 91, 29–66. [Google Scholar]
- Masamitsu, W. Chimera photoreceptor, neochrome, has arisen twice during plant evolution. Tanpakushitsu Kakusan Koso. Protein Nucleic Acid Enzym. 2006, 51, 1580–1589. [Google Scholar]
- Smith, H.L.; McAusland, L.; Murchie, E.H. Don’t ignore the green light: Exploring diverse roles in plant processes. J. Exp. Bot. 2017, 68, 2099–2110. [Google Scholar] [CrossRef]
- Wang, W.; Yu, L.-J.; Xu, C.; Tomizaki, T.; Zhao, S.; Umena, Y.; Chen, X.; Qin, X.; Xin, Y.; Suga, M.; et al. Structural basis for blue-green light harvesting and energy dissipation in diatoms. Science 2019, 363, eaav0365. [Google Scholar] [CrossRef]
- Willmer, C.M.; Johnston, W.R. Carbon dioxide assimilation in some aerial plant organs and tissues. Planta 1976, 130, 33–37. [Google Scholar] [CrossRef]
- Paik, I.; Huq, E. Plant photoreceptors: Multi-functional sensory proteins and their signaling networks. Semin. Cell Dev. Biol. 2019, 92, 114–121. [Google Scholar] [CrossRef]
- Neghab, H.K.; Soheilifar, M.H.; Grusch, M.; Ortega, M.M.; Djavid, G.E.; Saboury, A.A.; Goliaei, B. The state of the art of biomedical applications of optogenetics. Lasers Surg. Med. 2022, 54, 202–216. [Google Scholar] [CrossRef] [PubMed]
- Oesterhelt, D.; Stoeckenius, W. Rhodopsin-like Protein from the Purple Membrane of Halobacterium halobium. Nat. New Biol. 1971, 233, 149–152. [Google Scholar] [CrossRef]
- Boyden, E.S.; Zhang, F.; Bamberg, E.; Nagel, G.; Deisseroth, K. Millisecond-timescale, genetically targeted optical control of neural activity. Nat. Neurosci. 2005, 8, 1263–1268. [Google Scholar] [CrossRef]
- Katz, Y.; Sokoletsky, M.; Lampl, I. In-vivo optogenetics and pharmacology in deep intracellular recordings. J. Neurosci. Methods 2019, 325, 108324. [Google Scholar] [CrossRef]
- Sureda-Vives, M.; Sarkisyan, K.S. Bioluminescence-Driven Optogenetics. Life 2020, 10, 318. [Google Scholar] [CrossRef] [PubMed]
- Konrad, K.R.; Gao, S.; Zurbriggen, M.D.; Nagel, G. Optogenetic Methods in Plant Biology. Annu. Rev. Plant Biol. 2023, 74, 313–339. [Google Scholar] [CrossRef] [PubMed]
- Espinosa-Juárez, J.V.; Chiquete, E.; Estañol, B.; Aceves, J.d.J. Optogenetic and Chemogenic Control of Pain Signaling: Molecular Markers. Int. J. Mol. Sci. 2023, 24, 10220. [Google Scholar] [CrossRef] [PubMed]
- Zimmerman, S.; Kuhlman, B.; Yumerefendi, H. Engineering and Application of LOV2-Based Photoswitches. Methods Enzymol. 2016, 580, 169–190. [Google Scholar] [CrossRef] [PubMed]
- Hongdusit, A.; Zwart, P.H.; Sankaran, B.; Fox, J.M. Minimally disruptive optical control of protein tyrosine phosphatase 1B. Nat. Commun. 2020, 11, 788. [Google Scholar] [CrossRef]
- Carrasco-López, C.; Zhao, E.M.; Gil, A.A.; Alam, N.; Toettcher, J.E.; Avalos, J.L. Development of light-responsive protein binding in the monobody non-immunoglobulin scaffold. Nat. Commun. 2020, 11, 4045. [Google Scholar] [CrossRef]
- Chen, H.; Wu, F.; Xie, X.; Wang, W.; Li, Q.; Tu, L.; Li, B.; Kong, X.; Chang, Y. Hybrid Nanoplatform: Enabling a Precise Antitumor Strategy via Dual-Modal Imaging-Guided Photodynamic/Chemo-/Immunosynergistic Therapy. ACS Nano 2021, 15, 20643–20655. [Google Scholar] [CrossRef] [PubMed]
- Chmelyuk, N.S.; Oda, V.V.; Gabashvili, A.N.; Abakumov, M.A. Encapsulins: Structure, Properties, and Biotechnological Applications. Biochemistry 2023, 88, 35–49. [Google Scholar] [CrossRef] [PubMed]
- Kałafut, J.; Czapiński, J.; Przybyszewska-Podstawka, A.; Czerwonka, A.; Odrzywolski, A.; Sahlgren, C.; Rivero-Müller, A. Optogenetic control of NOTCH1 signaling. Cell Commun. Signal. 2022, 20, 67. [Google Scholar] [CrossRef] [PubMed]
- Adams, D.S.; Tseng, A.-S.; Levin, M. Light-activation of the Archaerhodopsin H+-pump reverses age-dependent loss of vertebrate regeneration: Sparking system-level controls in vivo. Biol. Open 2013, 2, 306–313. [Google Scholar] [CrossRef] [PubMed]
- Toh, P.J.Y.; Lai, J.K.H.; Hermann, A.; Destaing, O.; Sheetz, M.P.; Sudol, M.; Saunders, T.E. Optogenetic control of YAP cellular localisation and function. EMBO Rep. 2022, 23, e54401. [Google Scholar] [CrossRef]
- Toh, P.J.Y.; Sudol, M.; Saunders, T.E. Optogenetic control of YAP can enhance the rate of wound healing. Cell. Mol. Biol. Lett. 2023, 28, 39. [Google Scholar] [CrossRef]
- Conrad, K.S.; Bilwes, A.M.; Crane, B.R. Light-Induced Subunit Dissociation by a Light–Oxygen–Voltage Domain Photoreceptor from Rhodobacter sphaeroides. Biochemistry 2013, 52, 378–391. [Google Scholar] [CrossRef]
- Camsund, D.; Lindblad, P.; Jaramillo, A. Genetically engineered light sensors for control of bacterial gene expression. Biotechnol. J. 2011, 6, 826–836. [Google Scholar] [CrossRef]
- Zhou, Y.; Kong, D.; Wang, X.; Yu, G.; Wu, X.; Guan, N.; Weber, W.; Ye, H. A small and highly sensitive red/far-red optogenetic switch for applications in mammals. Nat. Biotechnol. 2022, 40, 262–272. [Google Scholar] [CrossRef]
- Reshetnikov, V.V.; Smolskaya, S.V.; Feoktistova, S.G.; Verkhusha, V.V. Optogenetic approaches in biotechnology and biomaterials. Trends Biotechnol. 2022, 40, 858–874. [Google Scholar] [CrossRef]
- McIsaac, R.S.; Bedbrook, C.N.; Arnold, F.H. Recent advances in engineering microbial rhodopsins for optogenetics. Curr. Opin. Struct. Biol. 2015, 33, 8–15. [Google Scholar] [CrossRef] [PubMed]
- Brinks, D.; Adam, Y.; Kheifets, S.; Cohen, A.E. Painting with Rainbows: Patterning light space, time, and wavelength for multiphoton optogenetic sensing control. Acc. Chem. Res. 2016, 49, 2518–2526. [Google Scholar] [CrossRef] [PubMed]
- Kandori, H. Retinal Proteins: Photochemistry and Optogenetics. Bull. Chem. Soc. Jpn. 2020, 93, 76–85. [Google Scholar] [CrossRef]
- Brechun, K.E.; Zhen, D.; Jaikaran, A.S.I.; Borisenko, V.; Kumauchi, M.; Hoff, W.D.; Arndt, K.M.; Woolley, G.A. Detection of Incorporation of p-Coumaric Acid into Photoactive Yellow Protein Variants in Vivo. Biochemistry 2019, 58, 2682–2694. [Google Scholar] [CrossRef]
- Romei, M.G.; Lin, C.-Y.; Boxer, S.G. Structural and spectroscopic characterization of photoactive yellow protein and photoswitchable fluorescent protein constructs containing heavy atoms. J. Photochem. Photobiol. A Chem. 2020, 401, 112738. [Google Scholar] [CrossRef] [PubMed]
- Woloschuk, R.M.; Reed, P.M.; Jaikaran, A.S.; Demmans, K.Z.; Youn, J.; Kanelis, V.; Uppalapati, M.; Woolley, G.A. Structure-based design of photoswitchable affibody scaffold. Protein Sci. 2021, 30, 2359. [Google Scholar] [CrossRef]
- Chernov, K.G.; Redchuk, T.A.; Omelina, E.S.; Verkhusha, V.V. Near-infrared flourescent proteins, biosensors, and optogenic tools engineered from phytochromes. Chem. Rev. 2017, 117, 6423. [Google Scholar] [CrossRef]
- Seong, J.; Lin, M.Z. Optobiochemistry: Genetically encoded control of protein activity by light. Annu. Rev. Biochem. 2021, 90, 475–501. [Google Scholar] [CrossRef]
- García-Ruiz, J.M.; van Zuilen, M.A.; Bach, W. Mineral self-organization on a lifeless planet. Phys. Life Rev. 2020, 34-35, 62–82. [Google Scholar] [CrossRef]
- Noorduin, W.L.; Grinthal, A.; Mahadevan, L.; Aizenberg, J. rationally designed complex, hierarchical microarchitectures. Science 2013, 340, 832–837. [Google Scholar] [CrossRef]
- Cuéllar-Cruz, M.; Islas, S.R.; Ramírez-Ramírez, N.; Pedraza-Reyes, M.; Moreno, A. Protection of the DNA from selected species of five kingdoms in nature by Ba(II), Sr(II), and Ca(II) Silica-Carbonates: Implications about biogenicity and evolving from prebiotic chemistry to biological chemistry. ACS Omega 2022, 7, 37410–37426. [Google Scholar] [CrossRef]
- Labas, Y.A.; Gurskaya, N.G.; Yanushevich, Y.G.; Fradkov, A.F.; Lukyanov, K.A.; Lukyanov, S.A.; Matz, M.V. Diversity and evolution of the green fluorescent protein family. Proc. Natl. Acad. Sci. USA 2002, 99, 4256–4261. [Google Scholar] [CrossRef]
- Cuéllar-Cruz, M. Influence of abiotic factors in the chemical origin of life: Biomorphs as a study model. ACS Omega 2021, 6, 8754–8763. [Google Scholar] [CrossRef]
- Vagenas, N.V.; Gatsouli, A.; Kontoyannis, C.G. Quantitative analysis of synthetic calcium carbonate polymorphs using FT-IR spectroscopy. Talanta 2003, 59, 831–836. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, P.; Berglund, K.A. In Situ Monitoring of Calcium Carbonate Polymorphs during batch crystallization in the presence of polymeric additives Using Raman Spectroscopy. Cryst. Growth Des. 2003, 3, 941–946. [Google Scholar] [CrossRef]
- Lin, C.-C.; Liu, L.-G. High-pressure Raman spectroscopic study of post-aragonite phase transition in witherite (BaCO3). Eur. J. Miner. 1997, 9, 785–792. [Google Scholar] [CrossRef]
- Buzgar, N.; Apopei, A.I. The Raman study of certain carbonates. Geologie 2009, 2, 97–112. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Type of Synthesis | Protein | Raman (cm−1) | Composition |
|---|---|---|---|
| Ca2+ | Control | 156, 282, 711, 1085 | Calcite |
| FT-blue | 155, 281, 750, 1084 | Calcite | |
| cgigRF | 156, 280, 711, 1084 | Calcite | |
| cgigGFP | 113, 302, 754, 1089 | Aragonite | |
| Ba2+ | Control | 91, 137, 153, 224, 689, 1057 | Witherite |
| FT-blue | 99, 140, 151, 233, 695, 1059, 1455, 2852 | Witherite | |
| cgigRF | 93, 140, 154, 222, 690, 1058, 2939 | Witherite | |
| cgigGFP | 95, 132, 223, 690, 1058, 1503, 2860 | Witherite | |
| Sr2+ | Control | 151, 184, 700, 1072 | Strontianite |
| FT-blue | 112, 150, 183, 252, 700, 1072, 1443, 1616, 2936 | Strontianite | |
| cgigRF | 109, 148, 180, 700, 1072, 2948 | Strontianite | |
| cgigGFP | 112, 150, 181, 699, 1071, 2929 | Strontianite |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Galindo-García, U.; Vanegas-Reza, M.; Arreguín-Espinosa, R.; Pérez, K.S.; Pérez-Solis, R.; Mendoza, M.E.; Cervantes-Quintero, K.Y.; Islas, S.R.; Cuéllar-Cruz, M.; Moreno, A. A Short and Practical Overview on Light-Sensing Proteins, Optogenetics, and Fluorescent Biomolecules inside Biomorphs Used as Optical Sensors. Crystals 2023, 13, 1343. https://doi.org/10.3390/cryst13091343
Galindo-García U, Vanegas-Reza M, Arreguín-Espinosa R, Pérez KS, Pérez-Solis R, Mendoza ME, Cervantes-Quintero KY, Islas SR, Cuéllar-Cruz M, Moreno A. A Short and Practical Overview on Light-Sensing Proteins, Optogenetics, and Fluorescent Biomolecules inside Biomorphs Used as Optical Sensors. Crystals. 2023; 13(9):1343. https://doi.org/10.3390/cryst13091343
Chicago/Turabian StyleGalindo-García, Ulises, María Vanegas-Reza, Roberto Arreguín-Espinosa, Karina Sandra Pérez, Ricardo Pérez-Solis, María Eugenia Mendoza, Karla Yadira Cervantes-Quintero, Selene R. Islas, Mayra Cuéllar-Cruz, and Abel Moreno. 2023. "A Short and Practical Overview on Light-Sensing Proteins, Optogenetics, and Fluorescent Biomolecules inside Biomorphs Used as Optical Sensors" Crystals 13, no. 9: 1343. https://doi.org/10.3390/cryst13091343
APA StyleGalindo-García, U., Vanegas-Reza, M., Arreguín-Espinosa, R., Pérez, K. S., Pérez-Solis, R., Mendoza, M. E., Cervantes-Quintero, K. Y., Islas, S. R., Cuéllar-Cruz, M., & Moreno, A. (2023). A Short and Practical Overview on Light-Sensing Proteins, Optogenetics, and Fluorescent Biomolecules inside Biomorphs Used as Optical Sensors. Crystals, 13(9), 1343. https://doi.org/10.3390/cryst13091343