
Structural Characterization of β-Xylosidase XynB2 from Geobacillus stearothermophilus CECT43: A Member of the Glycoside Hydrolase Family GH52

 ,
,  ,
, 
 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cloning, Expression and Purification of Gst43XynB2
2.2. Activity Measurements
2.3. Thermal Shift Assays
2.4. Crystallization
2.5. Data Collection and Refinement
2.6. Sequence and Structural Analyses
3. Results and Discussion
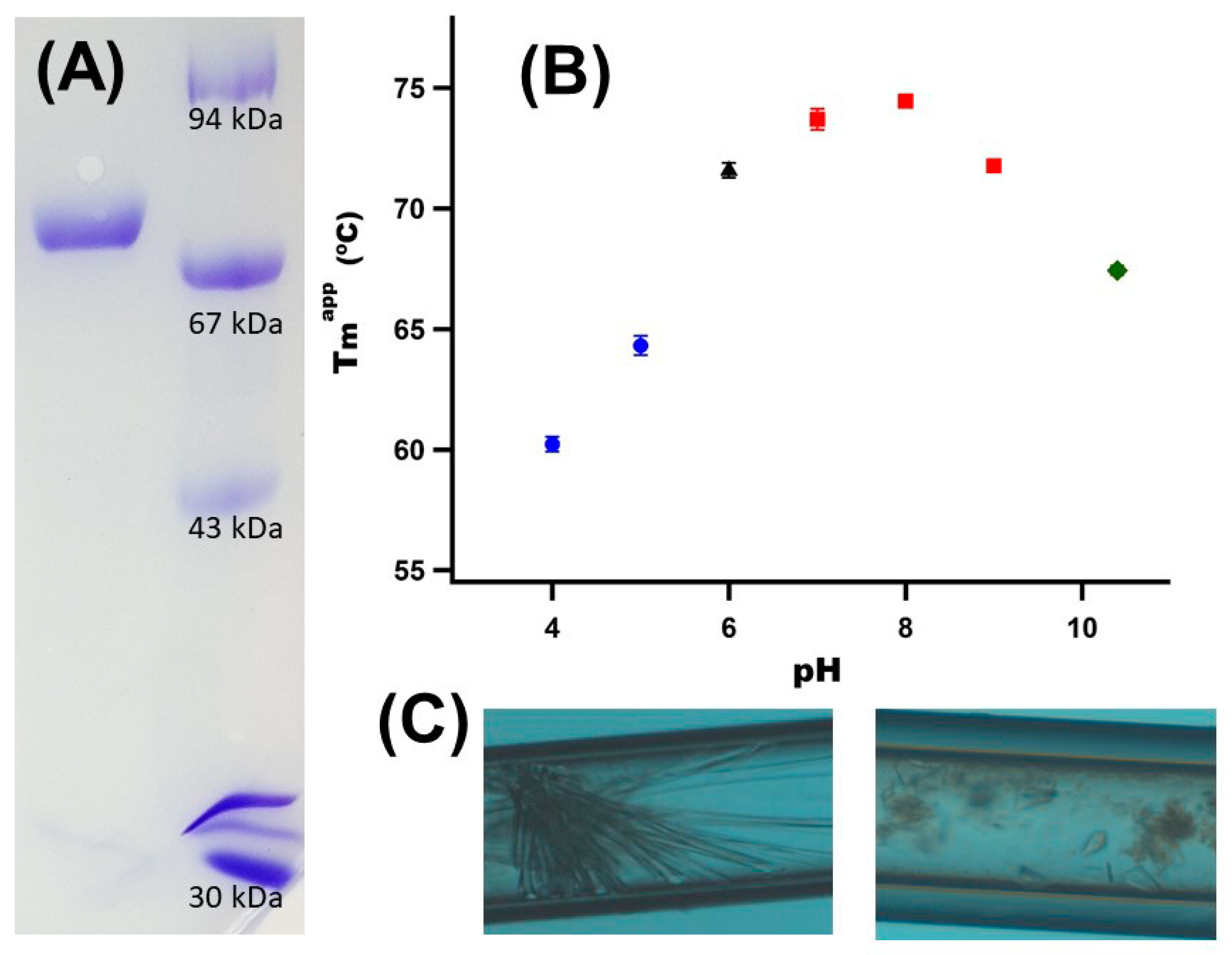
3.1. Gst43XynB2 Purification and Crystallization
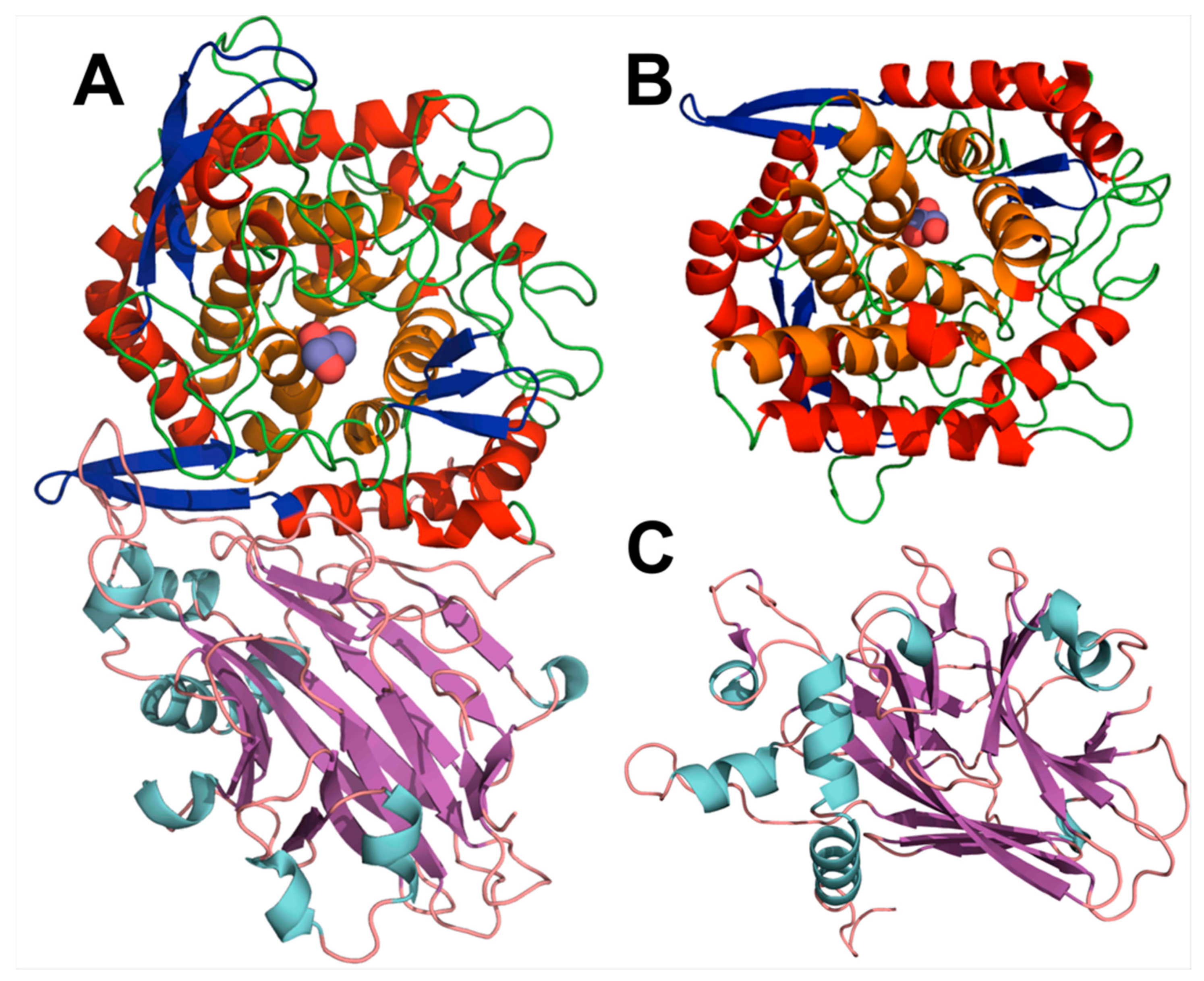
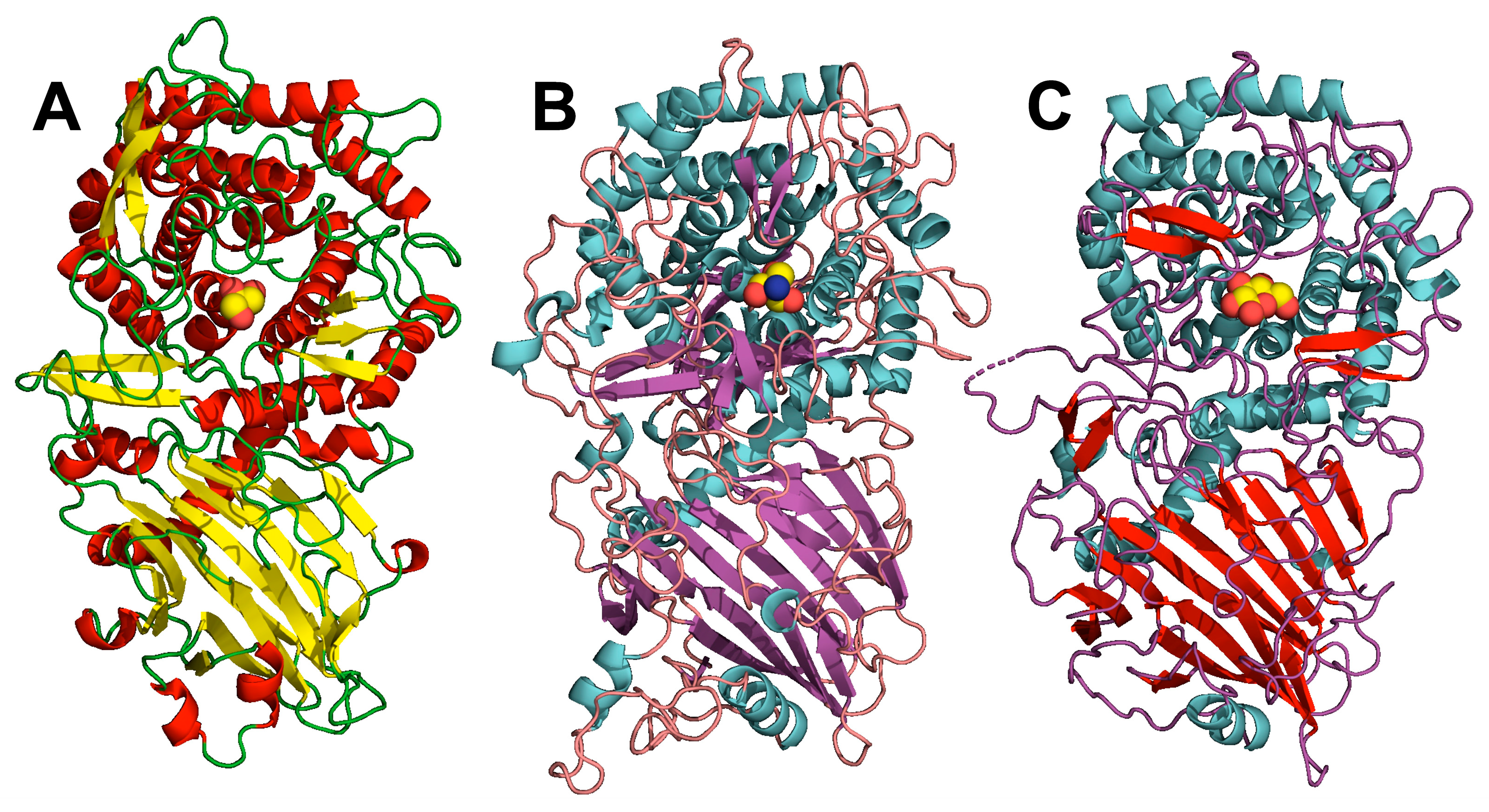
3.2. Gst43XynB2 Overall Fold
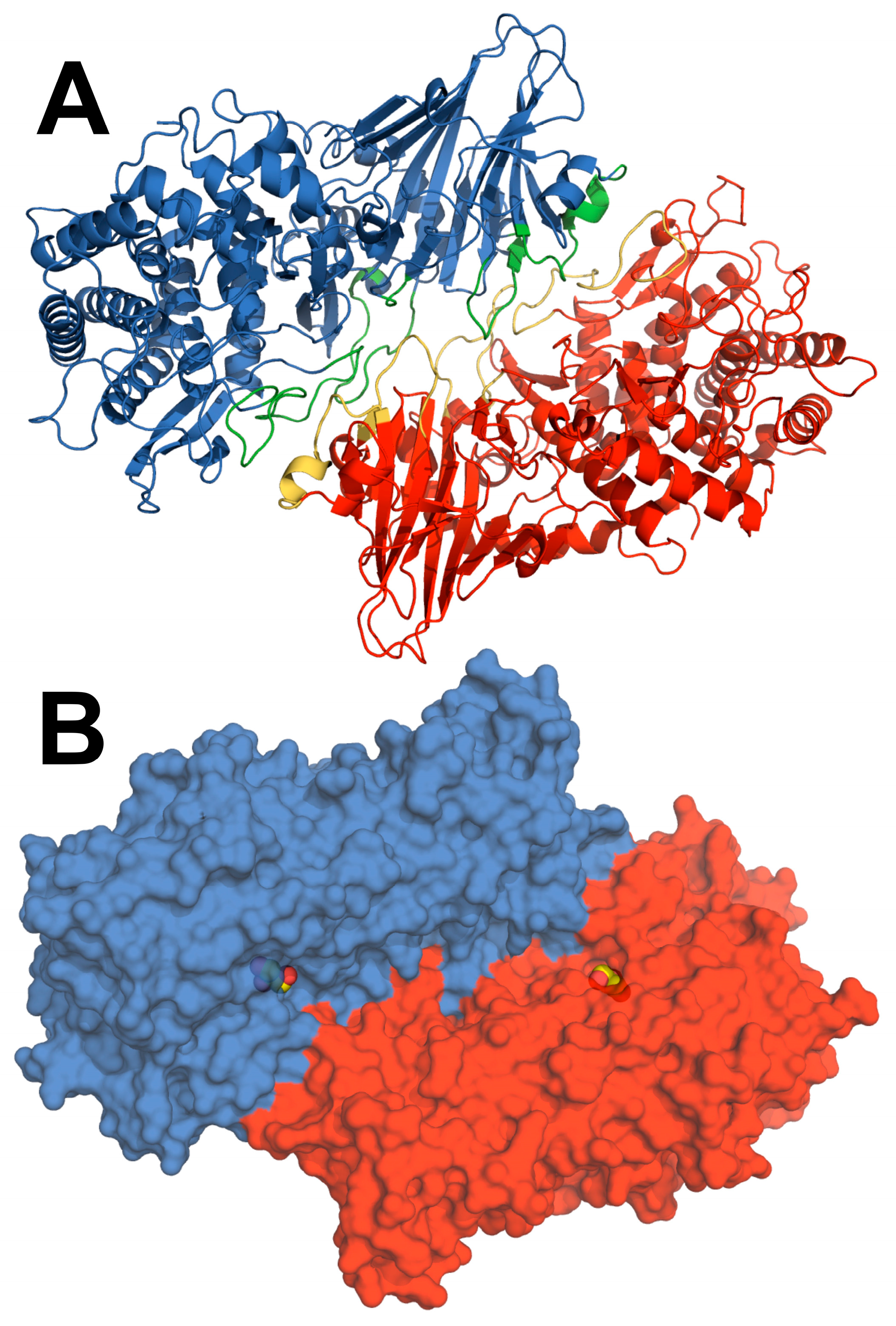
3.3. Insights into the Dimerization Interface of the GH52 Family
3.4. Comparison of GstXynB2 Structures Suggests the Involvement of a Dynamic Loop in Catalytic Cleft Closure
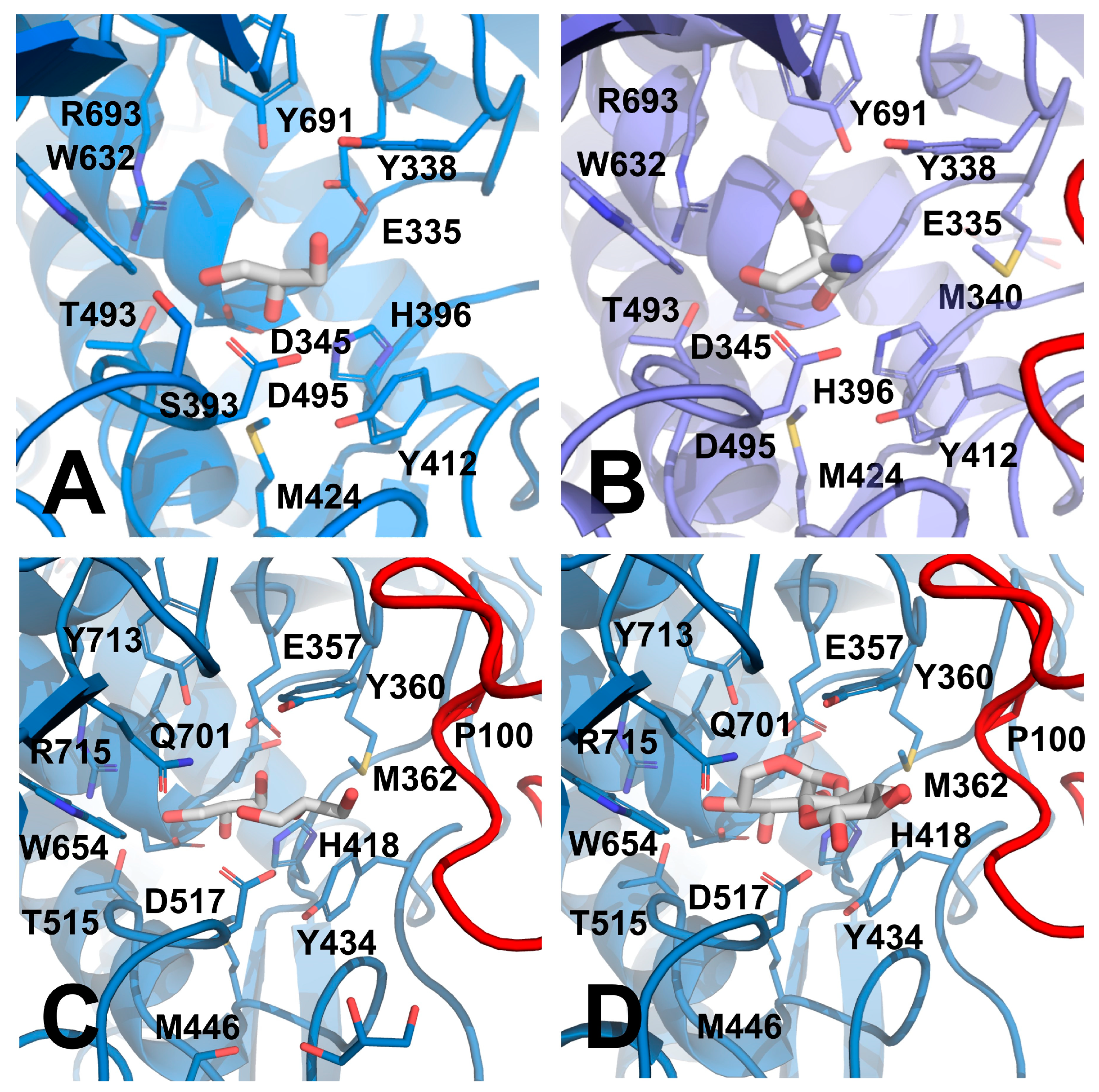
3.5. Structural Comparison of GH52 and GH116 Family
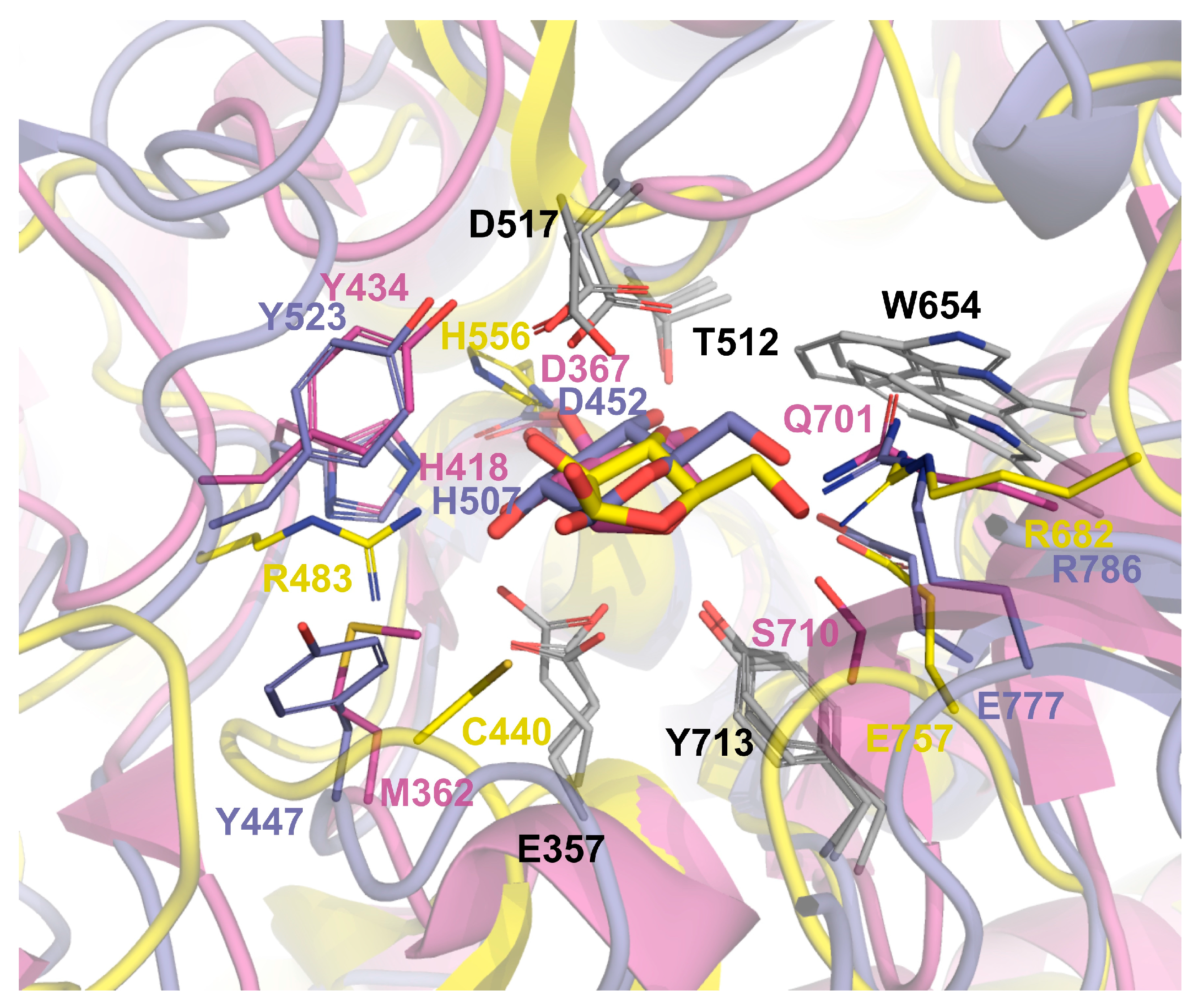
3.6. Insigths into the Glycone Specificity (−1 site) of GH52 and GH116 Families
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Knob, A.; Terrasan, C.R.F.; Carmona, E.C. β-Xylosidases from filamentous fungi: An overview. World J. Microbiol. Biotechnol. 2009, 26, 389–407. [Google Scholar] [CrossRef]
- Rohman, A.; Dijkstra, B.W.; Puspaningsih, N.N.T. β-Xylosidases: Structural Diversity, Catalytic Mechanism, and Inhibition by Monosaccharides. Int. J. Mol. Sci. 2019, 20, 5524. [Google Scholar] [CrossRef] [PubMed]
- Henrissat, B.; Davies, G. Structural and sequence-based classification of glycoside hydrolases. Curr. Opin. Struct. Biol. 1997, 7, 637–644. [Google Scholar] [CrossRef] [PubMed]
- Drula, E.; Garron, M.L.; Dogan, S.; Lombard, V.; Henrissat, B.; Terrapon, N. The carbohydrate-active enzyme database: Functions and literature. Nucleic Acids Res. 2022, 50, D571–D577. [Google Scholar] [CrossRef] [PubMed]
- Kaneko, S.; Fujimoto, Z. Chapter 8-β-D-Xylosidases: Structure-based substrate specificities and their applications. In Glycoside Hydrolases; Goyal, A., Kedar, S., Eds.; Academic Press: Cambridge, MA, USA, 2023; pp. 165–186. [Google Scholar]
- Nguyen, S.N.; Flores, A.; Talamantes, D.; Dar, F.; Valdez, A.; Schwans, J.; Berlemont, R. GeneHunt for rapid domain-specific annotation of glycoside hydrolases. Sci. Rep. 2019, 9, 10137. [Google Scholar] [CrossRef] [PubMed]
- Bravman, T.; Zolotnitsky, G.; Shulami, S.; Belakhov, V.; Solomon, D.; Baasov, T.; Shoham, G.; Shoham, Y. Stereochemistry of family 52 glycosyl hydrolases: A beta-xylosidase from Bacillus stearothermophilus T-6 is a retaining enzyme. FEBS Lett. 2001, 495, 39–43. [Google Scholar] [CrossRef] [PubMed]
- Bravman, T.; Zolotnitsky, G.; Belakhov, V.; Shoham, G.; Henrissat, B.; Baasov, T.; Shoham, Y. Detailed kinetic analysis of a family 52 glycoside hydrolase: A beta-xylosidase from Geobacillus stearothermophilus. Biochemistry 2003, 42, 10528–10536. [Google Scholar] [CrossRef] [PubMed]
- Bravman, T.; Belakhov, V.; Solomon, D.; Shoham, G.; Henrissat, B.; Baasov, T.; Shoham, Y. Identification of the catalytic residues in family 52 glycoside hydrolase, a beta-xylosidase from Geobacillus stearothermophilus T-6. J. Biol. Chem. 2003, 278, 26742–26749. [Google Scholar] [CrossRef]
- Contreras, L.M.; Gómez, J.; Prieto, J.; Clemente-Jiménez, J.M.; Las Heras-Vázquez, F.J.; Rodríguez-Vico, F.; Blanco, F.J.; Neira, J.L. The family 52 beta-xylosidase from Geobacillus stearothermophilus is a dimer: Structural and biophysical characterization of a glycoside hydrolase. Biochim. Biophys. Acta 2008, 1784, 1924–1934. [Google Scholar] [CrossRef]
- Quintero, D.; Velasco, Z.; Hurtado-Gómez, E.; Neira, J.L.; Contreras, L.M. Isolation and characterization of a thermostable beta-xylosidase in the thermophilic bacterium Geobacillus pallidus. Biochim. Biophys. Acta 2007, 1774, 510–518. [Google Scholar] [CrossRef]
- Suzuki, T.; Kitagawa, E.; Sakakibara, F.; Ibata, K.; Usui, K.; Kawai, K. Cloning, expression, and characterization of a family 52 beta-xylosidase gene (xysB) of a multiple-xylanase-producing bacterium, Aeromonas caviae ME-1. Biosci. Biotechnol. Biochem. 2001, 65, 487–494. [Google Scholar] [CrossRef] [PubMed]
- Teramoto, K.; Tsutsui, S.; Sato, T.; Fujimoto, Z.; Kaneko, S. Substrate Specificities of GH8, GH39, and GH52 β-xylosidases from Bacillus halodurans C-125 Toward Substituted Xylooligosaccharides. Appl. Biochem. Biotechnol. 2021, 193, 1042–1055. [Google Scholar] [CrossRef]
- Lee, T.-H.; Lee, Y.-E. Cloning, Sequencing and Expression of the Gene Encoding a Thermostable β -Xylosidase from Paenibacillus sp. DG-22. J. Life Sci. 2007, 17, 1197–1203. [Google Scholar] [CrossRef]
- Podkaminer, K.K.; Guss, A.M.; Trajano, H.L.; Hogsett, D.A.; Lynd, L.R. Characterization of xylan utilization and discovery of a new endoxylanase in Thermoanaerobacterium saccharolyticum through targeted gene deletions. Appl. Environ. Microbiol. 2012, 78, 8441–8447. [Google Scholar] [CrossRef] [PubMed]
- Espina, G.; Eley, K.; Pompidor, G.; Schneider, T.R.; Crennell, S.J.; Danson, M.J. A novel β-xylosidase structure from Geobacillus thermoglucosidasius: The first crystal structure of a glycoside hydrolase family GH52 enzyme reveals unpredicted similarity to other glycoside hydrolase folds. Acta Crystallogr. D Biol. Crystallogr. 2014, 70, 1366–1374. [Google Scholar] [CrossRef]
- Romero-Téllez, S.; Lluch, J.M.; González-Lafont, À.; Masgrau, L. Comparing Hydrolysis and Transglycosylation Reactions Catalyzed by. Front. Chem. 2019, 7, 200. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.; Liu, X.; Zhang, S.; Liu, Z. GH52 xylosidase from Geobacillus stearothermophilus: Characterization and introduction of xylanase activity by site-directed mutagenesis of Tyr509. J. Ind. Microbiol. Biotechnol. 2014, 41, 65–74. [Google Scholar] [CrossRef] [PubMed]
- Ben-David, A.; Shoham, G.; Shoham, Y. A universal screening assay for glycosynthases: Directed evolution of glycosynthase XynB2(E335G) suggests a general path to enhance activity. Chem. Biol. 2008, 15, 546–551. [Google Scholar] [CrossRef]
- Romero, G.; Contreras, L.M.; Aguirre, C.; Wilkesman, J.; Clemente-Jiménez, J.M.; Rodríguez-Vico, F.; Las Heras-Vázquez, F.J. Characterization of Cross-Linked Enzyme Aggregates of the Y509E Mutant of a Glycoside Hydrolase Family 52 β-xylosidase from. Molecules 2021, 26, 451. [Google Scholar] [CrossRef]
- Romero, G.; Contreras, L.M.; Aguirre Céspedes, C.; Wilkesman, J.; Clemente-Jiménez, J.M.; Rodríguez-Vico, F.; Las Heras-Vázquez, F.J. Efficiency Assessment between Entrapment and Covalent Bond Immobilization of Mutant β-Xylosidase onto Chitosan Support. Polymer 2023, 15, 3170. [Google Scholar] [CrossRef]
- Kubiak, M.; Kampen, I.; Schilde, C. Structure-Based Modeling of the Mechanical Behavior of Cross-Linked Enzyme Crystals. Crystals 2022, 12, 441. [Google Scholar] [CrossRef]
- Timofeev, V.; Samygina, V. Protein Crystallography: Achievements and Challenges. Crystals 2023, 13, 71. [Google Scholar] [CrossRef]
- Dann, R.; Lansky, S.; Lavid, N.; Zehavi, A.; Belakhov, V.; Baasov, T.; Dvir, H.; Manjasetty, B.; Belrhali, H.; Shoham, Y.; et al. Preliminary crystallographic analysis of Xyn52B2, a GH52 β-D-xylosidase from Geobacillus stearothermophilus T6. Acta Crystallogr. F Struct. Biol. Commun. 2014, 70, 1675–1682. [Google Scholar] [CrossRef] [PubMed]
- Charoenwattanasatien, R.; Pengthaisong, S.; Breen, I.; Mutoh, R.; Sansenya, S.; Hua, Y.; Tankrathok, A.; Wu, L.; Songsiriritthigul, C.; Tanaka, H.; et al. Bacterial β-Glucosidase Reveals the Structural and Functional Basis of Genetic Defects in Human Glucocerebrosidase 2 (GBA2). ACS Chem. Biol. 2016, 11, 1891–1900. [Google Scholar] [CrossRef] [PubMed]
- González-Ramírez, L.A.; Ruiz-Martínez, C.R.; Estremera-Andújar, R.A.; Nieves-Marrero, C.A.; García-Caballero, A.; Gavira, J.A.; López-Garriga, J.; García-Ruiz, J.M. Efficient Screening Methodology for Protein Crystallization Based on the Counter-Diffusion Technique. Cryst. Growth Des. 2017, 17, 6780–6786. [Google Scholar] [CrossRef]
- Kabsch, W. XDS. Acta Crystallogr. Sect. D Biol. Crystallogr. 2010, 66, 125–132. [Google Scholar] [CrossRef]
- Evans, P.R.; Murshudov, G.N. How good are my data and what is the resolution? Acta Crystallogr. Sect. D Biol. Crystallogr. 2013, 69, 1204–1214. [Google Scholar] [CrossRef]
- Vagin, A.; Teplyakov, A. Molecular replacement with MOLREP. Acta Crystallogr. Sect. D Biol. Crystallogr. 2010, 66, 22–25. [Google Scholar] [CrossRef]
- Winn, M.D.; Ballard, C.C.; Cowtan, K.D.; Dodson, E.J.; Emsley, P.; Evans, P.R.; Keegan, R.M.; Krissinel, E.B.; Leslie, A.G.; McCoy, A.; et al. Overview of the CCP4 suite and current developments. Acta Crystallogr. Sect. D Biol. Crystallogr. 2011, 67, 235–242. [Google Scholar] [CrossRef]
- Murshudov, G.N.; Skubák, P.; Lebedev, A.A.; Pannu, N.S.; Steiner, R.A.; Nicholls, R.A.; Winn, M.D.; Long, F.; Vagin, A.A. REFMAC5 for the refinement of macromolecular crystal structures. Acta Crystallogr. Sect. D Biol. Crystallogr. 2011, 67, 355–367. [Google Scholar] [CrossRef]
- Afonine, P.V.; Grosse-Kunstleve, R.W.; Echols, N.; Headd, J.J.; Moriarty, N.W.; Mustyakimov, M.; Terwilliger, T.C.; Urzhumtsev, A.; Zwart, P.H.; Adams, P.D. Towards automated crystallographic structure refinement with phenix.refine. Acta Crystallogr. Sect. D Biol. Crystallogr. 2012, 68, 352–367. [Google Scholar] [CrossRef] [PubMed]
- Liebschner, D.; Afonine, P.V.; Baker, M.L.; Bunkóczi, G.; Chen, V.B.; Croll, T.I.; Hintze, B.; Hung, L.W.; Jain, S.; McCoy, A.J.; et al. Macromolecular structure determination using X-rays, neutrons and electrons: Recent developments in Phenix. Acta Crystallogr. Sect. D Struct. Biol. 2019, 75, 861–877. [Google Scholar] [CrossRef] [PubMed]
- Emsley, P.; Lohkamp, B.; Scott, W.G.; Cowtan, K. Features and development of Coot. Acta Crystallogr. Sect. D Biol. Crystallogr. 2010, 66, 486–501. [Google Scholar] [CrossRef] [PubMed]
- Painter, J.; Merritt, E.A. Optimal description of a protein structure in terms of multiple groups undergoing TLS motion. Acta Crystallogr. Sect. D Biol. Crystallogr. 2006, 62, 439–450. [Google Scholar] [CrossRef] [PubMed]
- Chen, V.B.; Arendall, W.B.; Headd, J.J.; Keedy, D.A.; Immormino, R.M.; Kapral, G.J.; Murray, L.W.; Richardson, J.S.; Richardson, D.C. MolProbity: All-atom structure validation for macromolecular crystallography. Acta Crystallogr. Sect. D Biol. Crystallogr. 2010, 66, 12–21. [Google Scholar] [CrossRef] [PubMed]
- Madeira, F.; Park, Y.M.; Lee, J.; Buso, N.; Gur, T.; Madhusoodanan, N.; Basutkar, P.; Tivey, A.R.N.; Potter, S.C.; Finn, R.D.; et al. The EMBL-EBI search and sequence analysis tools APIs in 2019. Nucleic Acids Res. 2019, 47, W636–W641. [Google Scholar] [CrossRef]
- Robert, X.; Gouet, P. Deciphering key features in protein structures with the new ENDscript server. Nucleic Acids Res. 2014, 42, W320–W324. [Google Scholar] [CrossRef]
- Holm, L. DALI and the persistence of protein shape. Protein Sci. 2020, 29, 128–140. [Google Scholar] [CrossRef]
- Krissinel, E.; Henrick, K. Inference of macromolecular assemblies from crystalline state. J. Mol. Biol. 2007, 372, 774–797. [Google Scholar] [CrossRef]
- Laskowski, R.A.; Jabłońska, J.; Pravda, L.; Vařeková, R.S.; Thornton, J.M. PDBsum: Structural summaries of PDB entries. Protein Sci. 2018, 27, 129–134. [Google Scholar] [CrossRef]
- DeLano, W.L. PyMOL Molecular Graphics System. 2002. Available online: http://www.pymol.org. (accessed on 1 December 2023).
- Guex, N.; Peitsch, M.C. SWISS-MODEL and the Swiss-PdbViewer: An environment for comparative protein modeling. Electrophoresis 1997, 18, 2714–2723. [Google Scholar] [CrossRef] [PubMed]
- Kabsch, W. A solution for the best rotation to relate two sets of vectors. Acta Crystallogr. Sect. A Cryst. Phys. Diffr. Theor. Gen. Crystallogr. 1976, 32, 922–923. [Google Scholar] [CrossRef]
- Kuriata, A.; Gierut, A.M.; Oleniecki, T.; Ciemny, M.P.; Kolinski, A.; Kurcinski, M.; Kmiecik, S. CABS-flex 2.0: A web server for fast simulations of flexibility of protein structures. Nucleic Acids Res. 2018, 46, W338–W343. [Google Scholar] [CrossRef] [PubMed]
- Gotte, G.; Menegazzi, M. Protein Oligomerization. Int. J. Mol. Sci. 2023, 24, 10648. [Google Scholar] [CrossRef] [PubMed]
- Fraser, N.J.; Liu, J.W.; Mabbitt, P.D.; Correy, G.J.; Coppin, C.W.; Lethier, M.; Perugini, M.A.; Murphy, J.M.; Oakeshott, J.G.; Weik, M.; et al. Evolution of Protein Quaternary Structure in Response to Selective Pressure for Increased Thermostability. J. Mol. Biol. 2016, 428, 2359–2371. [Google Scholar] [CrossRef] [PubMed]
- Wilkens, C.; Vuillemin, M.; Pilgaard, B.; Polikarpov, I.; Morth, J.P. A GH115 α-glucuronidase structure reveals dimerization-mediated substrate binding and a proton wire potentially important for catalysis. Acta Crystallogr. Sect. D Struct. Biol. 2022, 78, 658–668. [Google Scholar] [CrossRef]
- McAndrew, R.P.; Park, J.I.; Heins, R.A.; Reindl, W.; Friedland, G.D.; D’haeseleer, P.; Northen, T.; Sale, K.L.; Simmons, B.A.; Adams, P.D. From soil to structure, a novel dimeric β-glucosidase belonging to glycoside hydrolase family 3 isolated from compost using metagenomic analysis. J. Biol. Chem. 2013, 288, 14985–14992. [Google Scholar] [CrossRef]
- Wu, X.; Zhao, S.; Tian, Z.; Han, C.; Jiang, X.; Wang, L. Dynamics of loops surrounding the active site architecture in GH5_2 subfamily TfCel5A for cellulose degradation. Biotechnol. Biofuels Bioprod. 2023, 16, 154. [Google Scholar] [CrossRef]
- Wu, M.; Bu, L.; Vuong, T.V.; Wilson, D.B.; Crowley, M.F.; Sandgren, M.; Ståhlberg, J.; Beckham, G.T.; Hansson, H. Loop motions important to product expulsion in the Thermobifida fusca glycoside hydrolase family 6 cellobiohydrolase from structural and computational studies. J. Biol. Chem. 2013, 288, 33107–33117. [Google Scholar] [CrossRef]
- Havukainen, R.; Törrönen, A.; Laitinen, T.; Rouvinen, J. Covalent binding of three epoxyalkyl xylosides to the active site of endo-1,4-xylanase II from Trichoderma reesei. Biochemistry 1996, 35, 9617–9624. [Google Scholar] [CrossRef]
- Hsieh, Y.C.; Wu, Y.J.; Chiang, T.Y.; Kuo, C.Y.; Shrestha, K.L.; Chao, C.F.; Huang, Y.C.; Chuankhayan, P.; Wu, W.G.; Li, Y.K.; et al. Crystal structures of Bacillus cereus NCTU2 chitinase complexes with chitooligomers reveal novel substrate binding for catalysis: A chitinase without chitin binding and insertion domains. J. Biol. Chem. 2010, 285, 31603–31615. [Google Scholar] [CrossRef] [PubMed]
- Hidaka, M.; Nishimoto, M.; Kitaoka, M.; Wakagi, T.; Shoun, H.; Fushinobu, S. The crystal structure of galacto-N-biose/lacto-N-biose I phosphorylase: A large deformation of a TIM barrel scaffold. J. Biol. Chem. 2009, 284, 7273–7283. [Google Scholar] [CrossRef] [PubMed]
- Meelua, W.; Thinkumrob, N.; Saparpakorn, P.; Pengthaisong, S.; Hannongbua, S.; Ketudat Cairns, J.R.; Jitonnom, J. Structural basis for inhibition of a GH116 β-glucosidase and its missense mutants by GBA2 inhibitors: Crystallographic and quantum chemical study. Chem. Biol. Interact. 2023, 384, 110717. [Google Scholar] [CrossRef] [PubMed]
- Pengthaisong, S.; Hua, Y.; Ketudat Cairns, J.R. Structural basis for transglycosylation in glycoside hydrolase family GH116 glycosynthases. Arch. Biochem. Biophys. 2021, 706, 108924. [Google Scholar] [CrossRef] [PubMed]
- Pengthaisong, S.; Piniello, B.; Davies, G.J.; Rovira, C.; Ketudat Cairns, J.R. Reaction Mechanism of Glycoside Hydrolase Family 116 Utilizes Perpendicular Protonation. ACS Catal. 2023, 13, 5850–5863. [Google Scholar] [CrossRef] [PubMed]
- Ferrara, M.C.; Cobucci-Ponzano, B.; Carpentieri, A.; Henrissat, B.; Rossi, M.; Amoresano, A.; Moracci, M. The identification and molecular characterization of the first archaeal bifunctional exo-β-glucosidase/N-acetyl-β-glucosaminidase demonstrate that family GH116 is made of three functionally distinct subfamilies. Biochim. Biophys. Acta 2014, 1840, 367–377. [Google Scholar] [CrossRef] [PubMed]
- Shimokawa, M.; Ishiwata, A.; Kashima, T.; Nakashima, C.; Li, J.; Fukushima, R.; Sawai, N.; Nakamori, M.; Tanaka, Y.; Kudo, A.; et al. Identification and characterization of endo-α-, exo-α-, and exo-β-D-arabinofuranosidases degrading lipoarabinomannan and arabinogalactan of mycobacteria. Nat. Commun. 2023, 14, 5803. [Google Scholar] [CrossRef]
- Cobucci-Ponzano, B.; Aurilia, V.; Riccio, G.; Henrissat, B.; Coutinho, P.M.; Strazzulli, A.; Padula, A.; Corsaro, M.M.; Pieretti, G.; Pocsfalvi, G.; et al. A new archaeal beta-glycosidase from Sulfolobus solfataricus: Seeding a novel retaining beta-glycan-specific glycoside hydrolase family along with the human non-lysosomal glucosylceramidase GBA2. J. Biol. Chem. 2010, 285, 20691–20703. [Google Scholar] [CrossRef]
- Huang, M.; Pengthaisong, S.; Charoenwattanasatien, R.; Thinkumrob, N.; Jitonnom, J.; Cairns, J.R.K. Systematic Functional and Computational Analysis of Glucose-Binding Residues in Glycoside Hydrolase Family GH116. Catalysts 2022, 12, 343. [Google Scholar] [CrossRef]
- Sansenya, S.; Mutoh, R.; Charoenwattanasatien, R.; Kurisu, G.; Ketudat Cairns, J.R. Expression and crystallization of a bacterial glycoside hydrolase family 116 β-glucosidase from Thermoanaerobacterium xylanolyticum. Acta Crystallogr. Sect. F Struct. Biol. Commun. 2015, 71, 41–44. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PDB ID. | 8QME |
| Beam Line | ID30B (ESRF) |
| Data collection | |
| Resolution range (Å) | 81.67–2.25 (2.33–2.25) |
| Space group | P 1 |
| Unit cell | |
| a, b, c (Å) | 81.12, 97.51, 107.40 |
| α, β, γ (°) | 107.46, 98.48, 106.55 |
| Unique reflections | 134,247 (13,293) |
| Multiplicity | 1.8 (1.7) |
| Completeness (%) | 97.17 (96.23) |
| Mean I/sigma (I) | 5.23 (1.31) |
| Wilson B-factor | 32.14 |
| R-merge | 0.09505 (0.5006) |
| CC1/2 | 0.985 (0.604) |
| Refinement | |
| R-work/R-free (%) | 17.79/21.29 |
| Number of atoms | |
| Protein | 22,179 |
| Ligands | 161 |
| Solvent | 1499 |
| Bond lengths (Å) | 0.003 |
| Bond angles (°) | 0.57 |
| Ramachandran (%) | |
| favoured | 97.46 |
| allowed | 2.54 |
| outliers | 0.00 |
| B-factor (Å2) | 35.56 |
| Gst43XynB2 | GthXynB2 | TxGH116 | ExoMA |
|---|---|---|---|
| PDB 8QME | PDB 4C1P | PDB 5BX5 | PDB 8IC7 |
| Glu335 | Glu357 | Glu441 | Glu431 |
| Tyr338 | Tyr360 | Tyr445 | Ser439/Cys440 |
| Met340 | Met362 | Tyr447 * | Cys440/Glu441/Cys758 * |
| Thr343 | Thr365 | Thr450 | Cys444 |
| Asp345 | Asp367 | Asp452 | His445 |
| Leu346 | Leu368 | Val453 | Val447 |
| His396 | His418 | His507 | Phe482/Arg483 * |
| Tyr412 | Tyr434 | Tyr523 | --- |
| Cys420/Phe421 | Cys442/Phe443 | Trp525 | Trp234 * |
| Met424 | Met446 | Trp531 | Ala498 |
| Thr493 | Thr515 | Thr591 | Thr555 |
| Tyr494 | Tyr516 | Tyr592 | Tyr556 |
| Asp495 | Asp 517 | Asp593 | Asp557 |
| Ser496 | Ser518 | Thr594 * | Tyr709 * |
| Trp632 | Trp654 | Trp732 | Trp714 |
| Gln679 | Gln701 | Arg786 * | Arg682 * |
| Ser688 | Ser710 | Ala787 * | Glu757/Cys758 * |
| Tyr691 | Tyr713 | Tyr790 | Tyr762 |
| Arg693 | Arg715 | Arg792 | Arg764 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gavira, J.A.; Contreras, L.M.; Alshamaa, H.M.; Clemente-Jiménez, J.M.; Rodríguez-Vico, F.; Las Heras-Vázquez, F.J.; Martínez-Rodríguez, S. Structural Characterization of β-Xylosidase XynB2 from Geobacillus stearothermophilus CECT43: A Member of the Glycoside Hydrolase Family GH52. Crystals 2024, 14, 18. https://doi.org/10.3390/cryst14010018
Gavira JA, Contreras LM, Alshamaa HM, Clemente-Jiménez JM, Rodríguez-Vico F, Las Heras-Vázquez FJ, Martínez-Rodríguez S. Structural Characterization of β-Xylosidase XynB2 from Geobacillus stearothermophilus CECT43: A Member of the Glycoside Hydrolase Family GH52. Crystals. 2024; 14(1):18. https://doi.org/10.3390/cryst14010018
Chicago/Turabian StyleGavira, Jose Antonio, Lellys M. Contreras, Hassan Mohamad Alshamaa, Josefa María Clemente-Jiménez, Felipe Rodríguez-Vico, Francisco Javier Las Heras-Vázquez, and Sergio Martínez-Rodríguez. 2024. "Structural Characterization of β-Xylosidase XynB2 from Geobacillus stearothermophilus CECT43: A Member of the Glycoside Hydrolase Family GH52" Crystals 14, no. 1: 18. https://doi.org/10.3390/cryst14010018
APA StyleGavira, J. A., Contreras, L. M., Alshamaa, H. M., Clemente-Jiménez, J. M., Rodríguez-Vico, F., Las Heras-Vázquez, F. J., & Martínez-Rodríguez, S. (2024). Structural Characterization of β-Xylosidase XynB2 from Geobacillus stearothermophilus CECT43: A Member of the Glycoside Hydrolase Family GH52. Crystals, 14(1), 18. https://doi.org/10.3390/cryst14010018