
Size and Shape Controlled Crystallization of Hemoglobin for Advanced Crystallography

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction

2. Results and Discussion
2.1. High-Quality, Thin Hb Crystal Growth for XRC Combined with Photoexcitation and Spectrophotometry
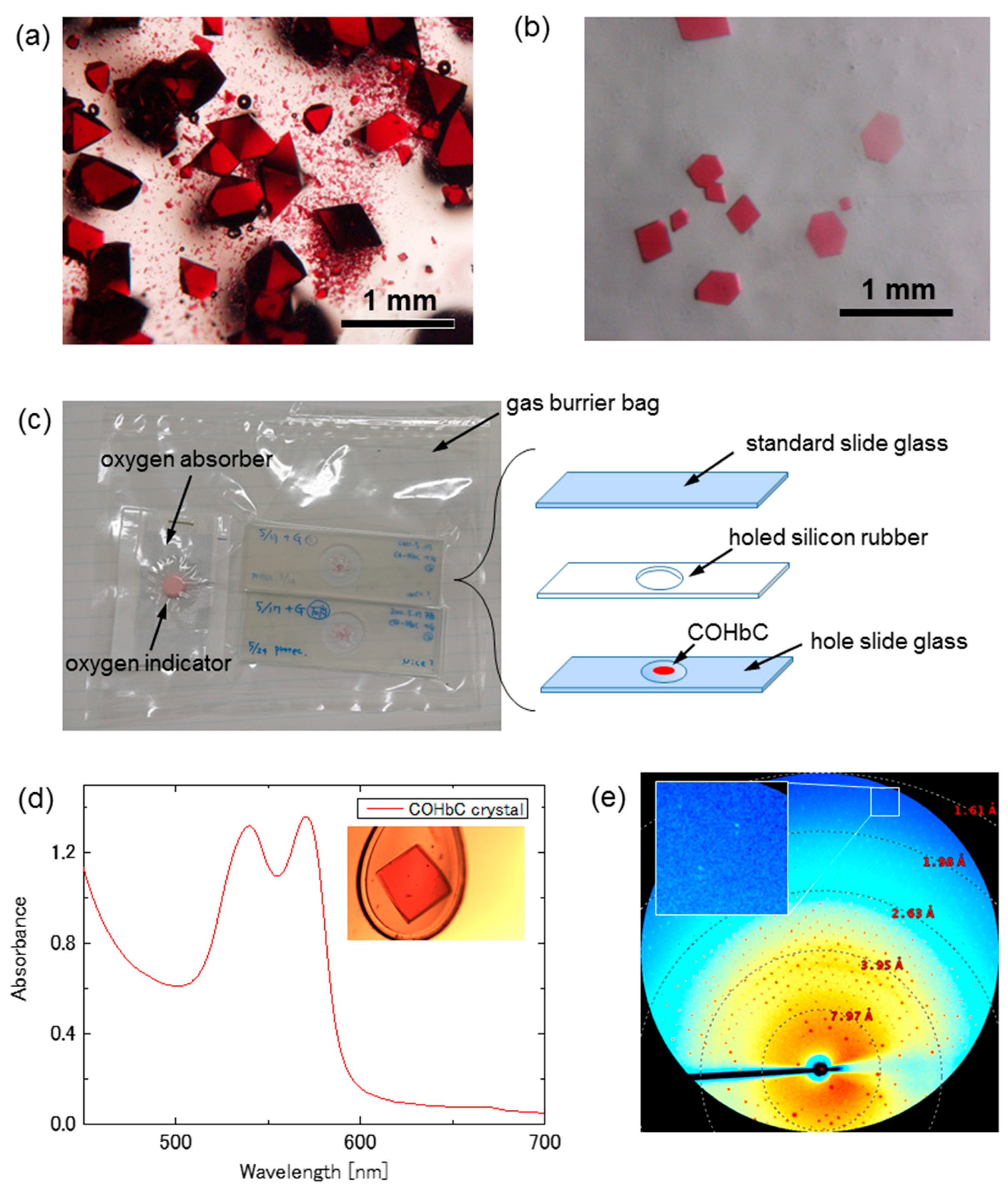
2.1.1. Human COHbC (β6 Glu→Lys) Crystal (R-State)
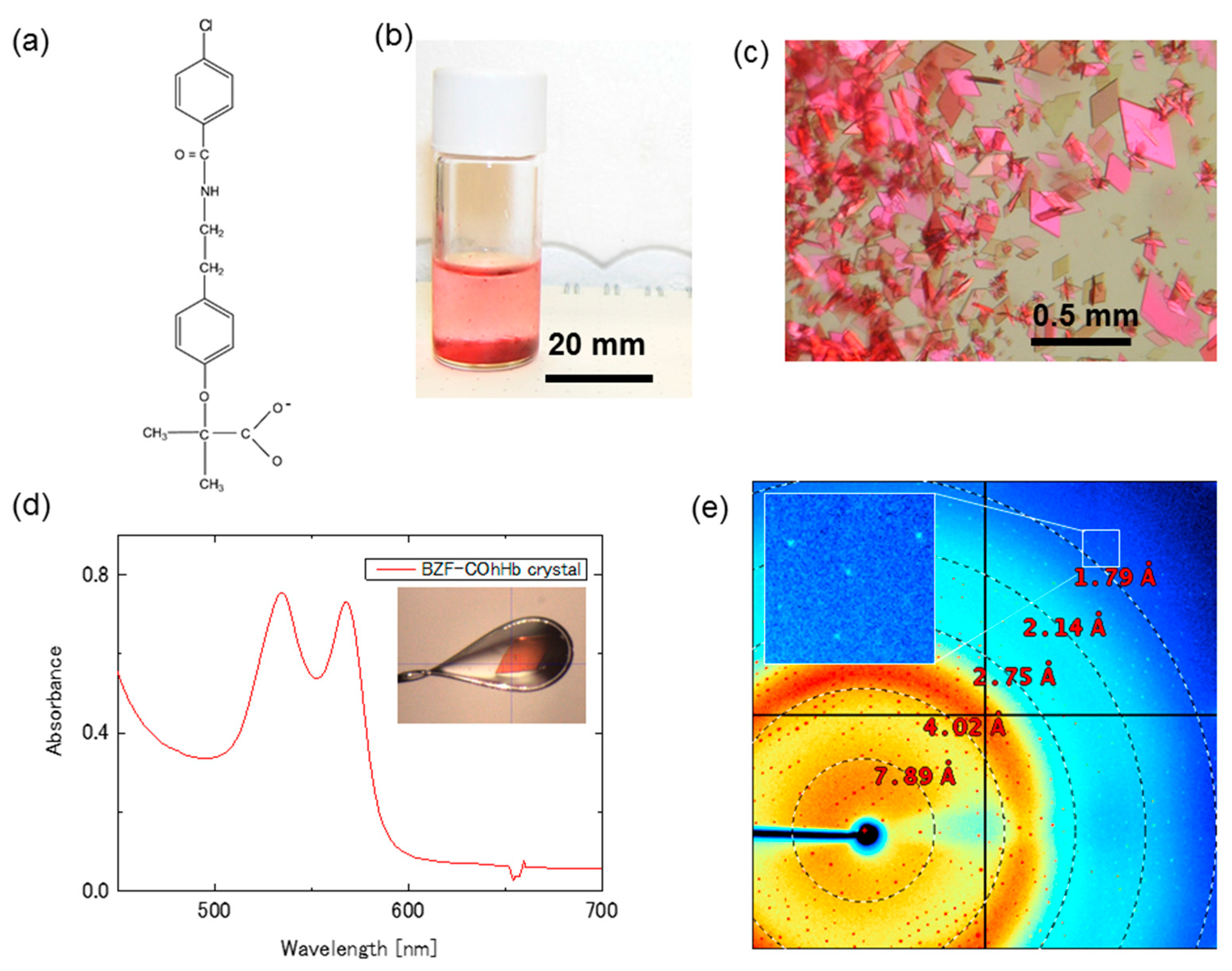
2.1.2. Bezafibrate-Bound Horse COHb (BZF-COhHb) Crystal (R-State)
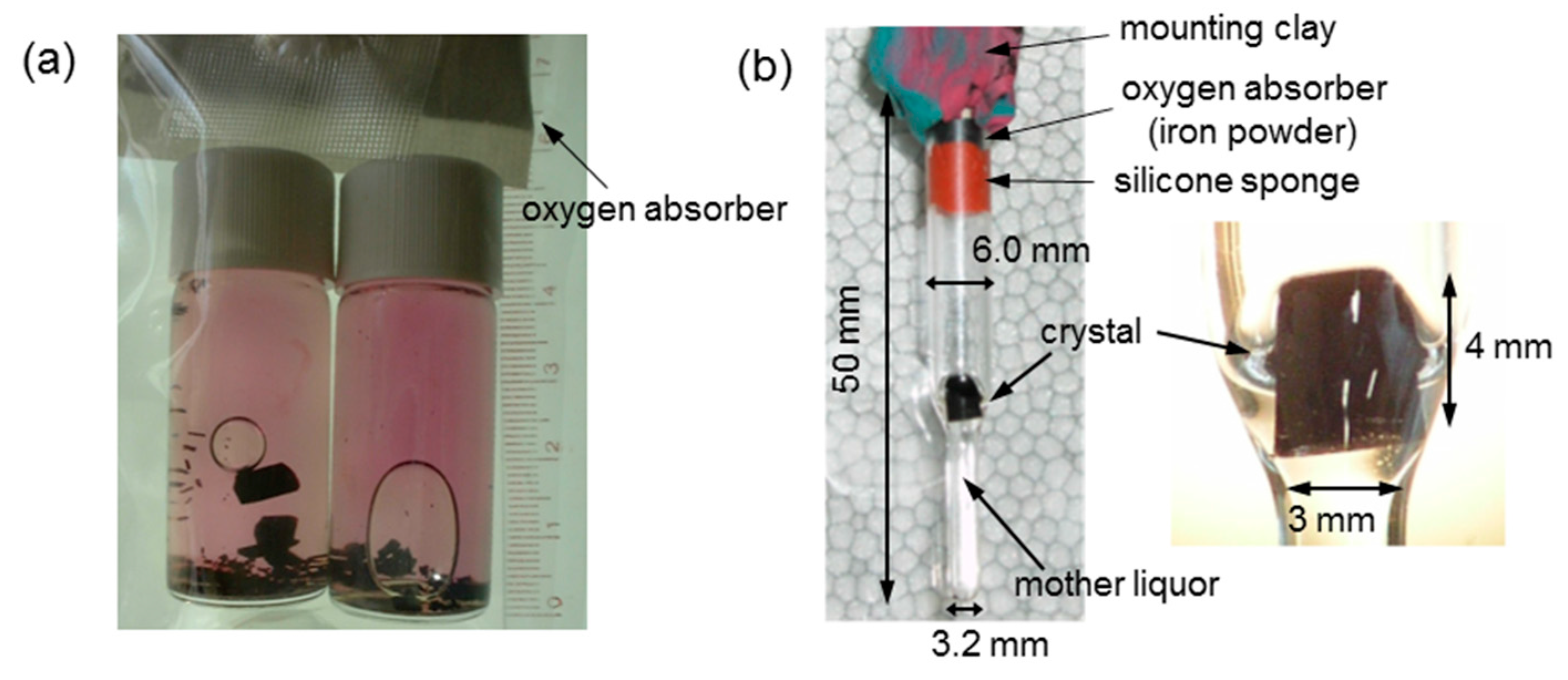
2.2. Large-Volume (>20 mm3) Hb Crystal Growth for Neutron Crystallgoraphy (NC)
2.2.1. DeoxyHbA Crystal (T-State)
2.2.2. COHbA Crystal (R-State)
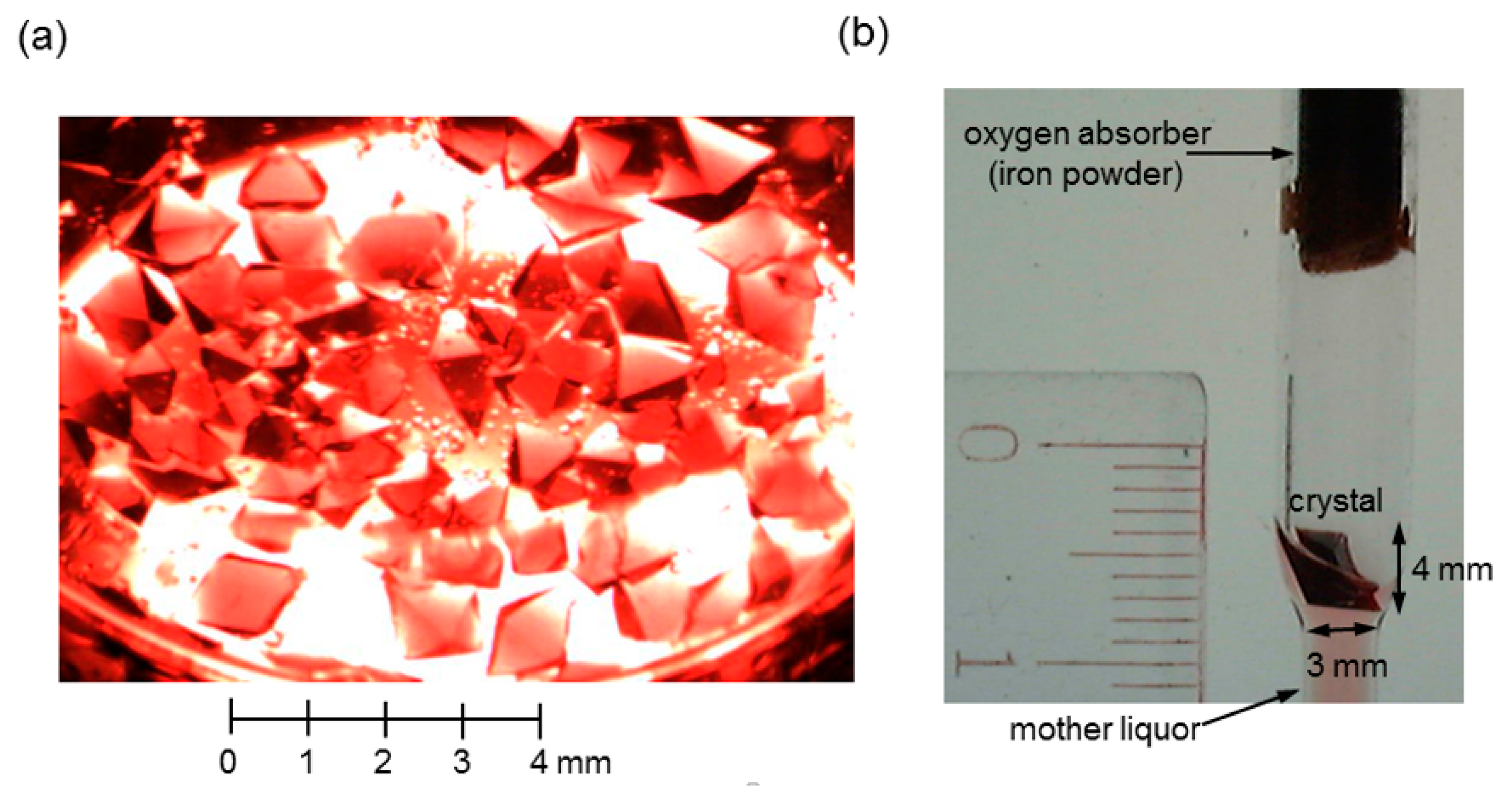
2.3. Extremely Large-Volume (>100 mm3) Hb Crystal Growth for X-ray Fluorescence Holography (XFH)
3. Materials and Methods
3.1. Preparation of HbA, HbC, and Horse Hb
3.2. Microspectrophotometry and Diffraction Image Collection for the COHbC Crystal
3.3. Microspectrophotometry and Diffraction Image Collection for the BZF-COhHb Crystal
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Kendrew, J.C.; Bodo, G.; Dintzis, H.M.; Parrish, R.G.; Wyckoff, H.; Phillips, D.C. A three-dimensional model of the myoglobin molecule obtained by X-ray analysis. Nature 1958, 181, 662–666. [Google Scholar] [CrossRef] [PubMed]
- Pertz, M.F.; Rossmann, M.G.; Cullis, A.F.; Muirhead, H.; Will, G.; North, A.C. Structure of haemoglobin: A three-dimensional Fourier synthesis at 5.5-A. resolution, obtained by X-ray analysis. Nature 1960, 185, 416–422. [Google Scholar] [CrossRef]
- Kendrew, J.C.; Dickerson, R.E.; Strandberg, B.E.; Hart, R.G.; Davies, D.R.; Phillips, D.C.; Shore, V.C. Structure of myoglobin: A three-dimensional Fourier synthesis at 2 Å resolution. Nature 1960, 185, 422–427. [Google Scholar] [CrossRef] [PubMed]
- RCSB Protein Data Bank. Available online: https://www.rcsb.org (accessed on 2 September 2017).
- Shibayama, N.; Morimoto, H.; Kitagawa, T. Properties of chemically modified Ni(II)-Fe(II) hybrid hemoglobins: Ni(II) protoporphyrin IX as a model for a permanent deoxy-heme. J. Mol. Biol. 1986, 192, 331–336. [Google Scholar] [CrossRef]
- Shibayama, N.; Imai, K.; Hirata, H.; Hiraiwa, H.; Morimoto, H.; Saigo, S. Oxygen equilibrium properties of highly purified human adult hemoglobin cross-linked between 82β1 and 82β2 lysyl residues by bis(3,5-dibromosalicyl)fumarate. Biochemistry 1991, 30, 8158–8165. [Google Scholar] [CrossRef] [PubMed]
- Adachi, S.; Park, S.-Y.; Tame, J.R.H.; Shiro, Y.; Shibayama, N. Direct observation of photolysis-induced tertiary structural changes in hemoglobin. Proc. Natl. Acad. Sci. USA 2003, 100, 517–520. [Google Scholar] [CrossRef] [PubMed]
- Schotte, F.; Cho, H.S.; Soman, J.; Wulff, M.; Olson, J.S.; Anfinrud, P.A. Real-time tacking of CO migration and binding in the α and β subunits of human hemoglobin via 150-ps time-resolved Laue crystallography. Chem. Phys. 2013, 422, 98–106. [Google Scholar] [CrossRef] [PubMed]
- Mozzarelli, A.; Rivetti, C.; Rossi, G.L.; Henry, E.R.; Eaton, W.A. Crystals of haemoglobin with the T quaternary structure bind oxygen noncooperatively with no Bohr effect. Nature 1991, 351, 416–419. [Google Scholar] [CrossRef] [PubMed]
- Shibayama, N.; Sugiyama, K.; Park, S.Y. Structures and oxygen affinities of crystalline human hemoglobin C (β6 Glu→Lys) in the R and R2 quaternary structures. J. Biol. Chem. 2011, 286, 33661–33668. [Google Scholar] [CrossRef] [PubMed]
- Blakeley, M.P.; Langan, P.; Niimura, N.; Podjarny, A. Neutron crystallography: Opportunities, challenges, and limitations. Curr. Opin. Struct. Biol. 2008, 18, 593–600. [Google Scholar] [CrossRef] [PubMed]
- Niimura, N.; Bau, R. Neutron protein crystallography: Beyond the folding structure of biological macromolecules. Acta Crystallogr. 2008, 64, 12–22. [Google Scholar] [CrossRef] [PubMed]
- Sato-Tomita, A.; Shibayama, S.; Happo, N.; Kimura, K.; Okabe, T.; Matsushita, T.; Park, S.-Y.; Sasaki, Y.C.; Hayashi, K. Development of an X-ray fluorescence holographic measurement system for protein crystals. Rev. Sci. Instrum. 2016, 87. [Google Scholar] [CrossRef] [PubMed]
- Dickerson, R.E.; Geis, I. Hemoglobin: Structure, Function, Evolution, and Pathology, 1st ed.; The Benjamin/Cummings Publishing Company: Menlo Park, CA, USA, 1983; pp. 76–116. ISBN 0-805-32411-9. [Google Scholar]
- Baldwin, J.; Chothia, C. Haemoglobin: The structural changes related to ligand binding and its allosteric mechanism. J. Mol. Biol. 1979, 129, 175–220. [Google Scholar] [CrossRef]
- Ren, Z. Reaction trajectory revealed by a joint analysis of protein data bank. PLoS ONE 2013, 8, e77141. [Google Scholar] [CrossRef] [PubMed]
- Schlichting, I.; Berendzen, J.; Phillips, G.N., Jr.; Sweet, R.M. Crystal structure of photolysed carbonmonoxy-myoglobin. Nature 1994, 371, 808–812. [Google Scholar] [CrossRef] [PubMed]
- Teng, T.Y.; Srajer, V.; Moffat, K. Photolysis-induced structural changes in single crystals of carbonmonoxy myoglobin at 40 K. Nat. Struct. Biol. 1994, 1, 701–705. [Google Scholar] [CrossRef] [PubMed]
- Ostermann, A.; Waschipky, R.; Parak, F.G.; Nienhaus, G.U. Ligand binding and conformational motions in myoglobin. Nature 2000, 404, 205–208. [Google Scholar] [CrossRef] [PubMed]
- Tomita, A.; Sato, T.; Ichiyanagi, K.; Nozawa, S.; Ichikawa, H.; Chollet, M.; Kawai, F.; Park, S.Y.; Tsuduki, T.; Yamato, T.; et al. Visualizing breathing motion of internal cavities in concert with ligand migration in myoglobin. Proc. Natl. Acad. Sci. USA 2009, 106, 2612–2616. [Google Scholar] [CrossRef] [PubMed]
- Sawicki, C.A.; Gibson, Q.H. Dependence of the quantum efficiency for photolysis of carboxyhemoglobin on the degree of ligation. J. Biol. Chem. 1979, 254, 4058–4062. [Google Scholar] [PubMed]
- Hirsch, R.E.; Juszczak, L.J.; Fataliev, N.A.; Friedman, J.M.; Nagel, R.L. Solution-active structural alterations in liganded hemoglobins C (β6 Glu→Lys) and S (β6 Glu→Val). J. Biol. Chem. 1999, 274, 13777–13782. [Google Scholar] [CrossRef] [PubMed]
- Diggs, L.W.; Kraus, A.P.; Morrison, D.B.; Rudnicki, R.P. Intraerythrocytic crystals in a white patient with hemoglobin C in the absence of other types of hemoglobin. Blood 1954, 9, 1172–1184. [Google Scholar] [PubMed]
- Hirsch, R.E.; Raventos-Suarez, C.; Olson, J.A.; Nagel, R.L. Ligand state of intraerythrocytic circulating HbC crystals in homozygote CC patients. Blood 1985, 66, 775–777. [Google Scholar] [PubMed]
- Perutz, M.F.; Fermi, G.; Abraham, D.J.; Poyart, C.; Bursaux, E. Hemoglobin as a receptor of drugs and peptides: X-ray studies of the stereochemistry of binding. J. Am. Chem. Soc. 1986, 108, 1064–1078. [Google Scholar] [CrossRef]
- Shibayama, N.; Miura, S.; Tame, J.R.H.; Yonetani, T.; Park, S.-Y. Crystal structure of horse carbonmonoxyhemoglobin-bezafibrate complex at 1.55-Å resolution. A novel allosteric binding site in R-state hemoglobin. J. Biol. Chem. 2002, 277, 38791–38796. [Google Scholar] [CrossRef] [PubMed]
- Parak, F.; Knapp, E.W.; Kucheida, D. Protein dynamics: Mössbauer spectroscopy on deoxymyoglobin crystals. J. Mol. Biol. 1982, 161, 177–194. [Google Scholar] [CrossRef]
- Doster, W.; Cusack, S.; Petry, W. Dynamical transition of myoglobin revealed by inelastic neutron scattering. Nature 1989, 337, 754–756. [Google Scholar] [CrossRef] [PubMed]
- Monod, J.; Wyman, J.; Changeux, J.-P. On the nature of allosteric transitions: A plausible model. J. Mol. Biol. 1965, 12, 88–118. [Google Scholar] [CrossRef]
- Ng, J.D.; Baird, J.K.; Coates, L.; García-Ruiz, J.M.; Hodge, T.A.; Huang, S. Large-volume protein crystal growth for neutron macromolecular crystallography. Acta Crystallogr. F Struct. Biol. Commun. 2015, 71, 358–370. [Google Scholar] [CrossRef] [PubMed]
- Blakeley, M.P.; Hasnain, S.S.; Antonyuk, S.V. Sub-atomic resolution X-ray crystallography and neutron crystallography: promise, challenges and potential. IUCrJ 2015, 2, 464–474. [Google Scholar] [CrossRef] [PubMed]
- Pertz, M.F. Preparation of hemoglobin crystals. J. Cryst. Growth 1968, 2, 54–56. [Google Scholar] [CrossRef]
- Chatake, T.; Shibayama, N.; Park, S.-Y.; Kurihara, K.; Tamada, T.; Tanaka, I.; Niimura, N.; Kuroki, R.; Morimoto, Y. Protonation states of buried histidine residues in human deoxyhemoglobin revealed by neutron crystallography. J. Am. Chem. Soc. 2007, 129, 14840–14841. [Google Scholar] [CrossRef] [PubMed]
- Imai, K.; Yonetani, T. PH dependence of the Adair constants of human hemoglobin. Nonuniform contribution of successive oxygen bindings to the alkaline Bohr effect. J. Biol. Chem. 1975, 250, 2227–2231. [Google Scholar] [PubMed]
- Benvenuti, M.; Mangani, S. Crystallization of soluble proteins in vapor diffusion for X-ray crystallography. Nat. Protoc. 2007, 2, 1633–1651. [Google Scholar] [CrossRef] [PubMed]
- García-Ruiz, J.M. Counter diffusion methods for macromolecular crystallization. Methods Enzymol. 2003, 368, 130–154. [Google Scholar] [PubMed]
- Tegze, M.; Faigel, G. X-ray holography with atomic resolution. Nature 1996, 380, 49–51. [Google Scholar] [CrossRef]
- Gog, T.; Len, P.M.; Materlik, G.; Bahr, D.; Fadley, C.S.; Sanchez-Hanke, C. Multiple-energy X-ray holography: Atomic images of hematite (Fe2O3). Phys. Rev. Lett. 1996, 76, 3132–3135. [Google Scholar] [CrossRef] [PubMed]
- Sakai, K.; Matsui, Y.; Kouyama, T.; Shiro, Y.; Adachi, S. Optical monitoring of freeze-trapped reaction intermediates in protein crystals: a microspectro-photometer for cryogenic protein crystallography. J. Appl. Cryst. 2002, 35, 270–273. [Google Scholar] [CrossRef]
- Nozawa, S.; Adachi, S.; Takahashi, J.; Tazaki, R.; Guérin, L.; Daimon, M.; Tomita, A.; Sato, T.; Chollet, M.; Collet, E.; et al. Developing 100 ps-resolved X-ray structural analysis capabilities on beamline NW14A at the Photon Factory Advanced Ring. J. Synchrotron Radiat. 2007, 14, 313–319. [Google Scholar] [CrossRef] [PubMed]
- Orville, A.M.; Buono, R.; Cowan, M.; Heroux, A.; Shea-McCarthy, G.; Schneider, D.K.; Skinner, J.M.; Skinner, M.J.; Stoner-Ma, D.; Sweet, R.M. Correlated single-crystal electronic absorption spectroscopy and X-ray crystallography at NSLS beamline X26-C. J. Synchrotron Radiat. 2011, 18, 358–366. [Google Scholar] [CrossRef] [PubMed]







© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sato-Tomita, A.; Shibayama, N. Size and Shape Controlled Crystallization of Hemoglobin for Advanced Crystallography. Crystals 2017, 7, 282. https://doi.org/10.3390/cryst7090282
Sato-Tomita A, Shibayama N. Size and Shape Controlled Crystallization of Hemoglobin for Advanced Crystallography. Crystals. 2017; 7(9):282. https://doi.org/10.3390/cryst7090282
Chicago/Turabian StyleSato-Tomita, Ayana, and Naoya Shibayama. 2017. "Size and Shape Controlled Crystallization of Hemoglobin for Advanced Crystallography" Crystals 7, no. 9: 282. https://doi.org/10.3390/cryst7090282
APA StyleSato-Tomita, A., & Shibayama, N. (2017). Size and Shape Controlled Crystallization of Hemoglobin for Advanced Crystallography. Crystals, 7(9), 282. https://doi.org/10.3390/cryst7090282