High-Resolution Crystal Structure of RpoS Fragment including a Partial Region 1.2 and Region 2 from the Intracellular Pathogen Legionella pneumophila


Abstract
:1. Introduction
2. Materials and Methods
2.1. Cloning, Expression and Purification of LpRpoS
2.2. Crystallization and Data Collection
2.3. Structure Determination and Refinement
3. Results
3.1. Crystallization of LpRpoS 95–195
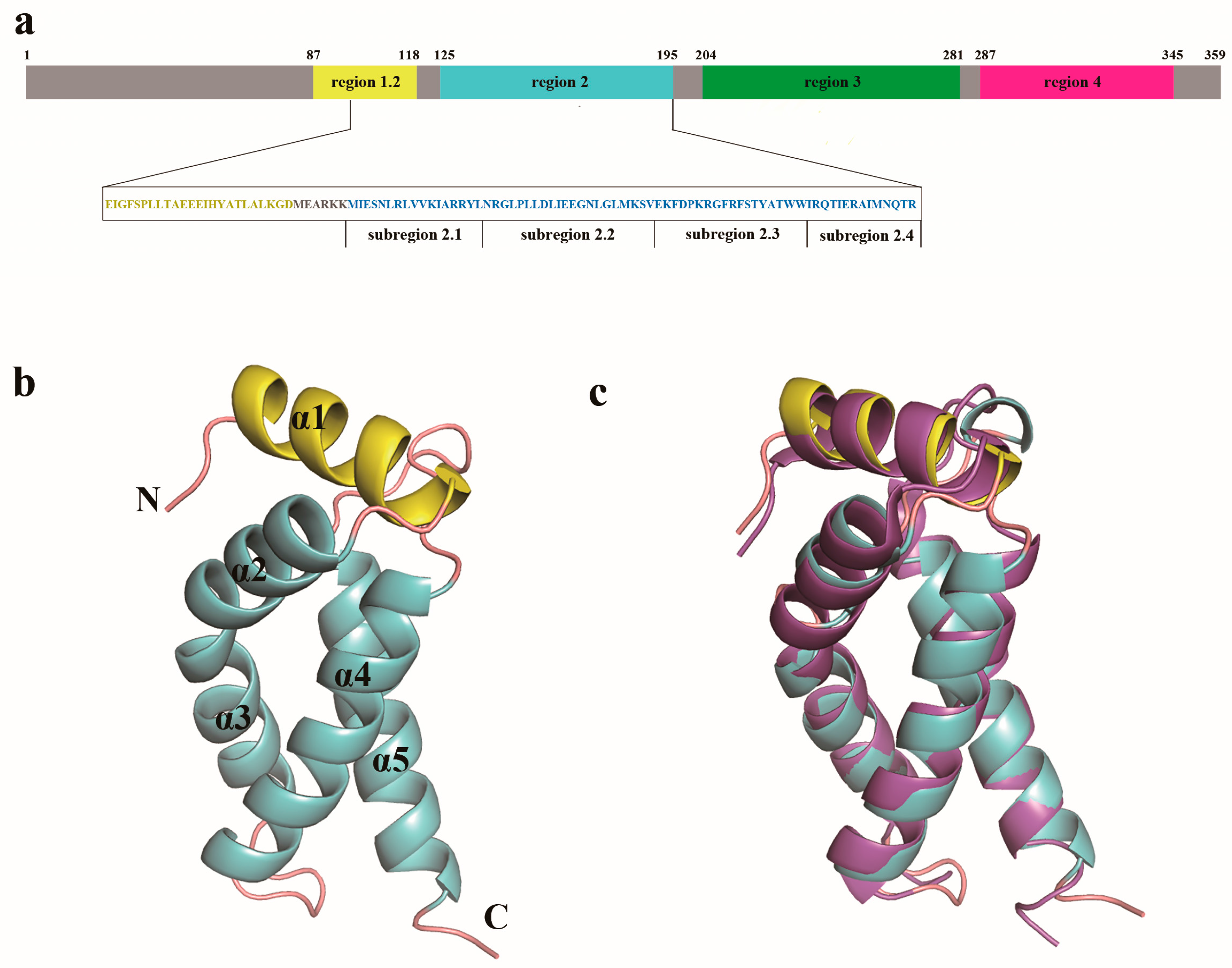
3.2. Overall Structure of LpRpoS 95–195
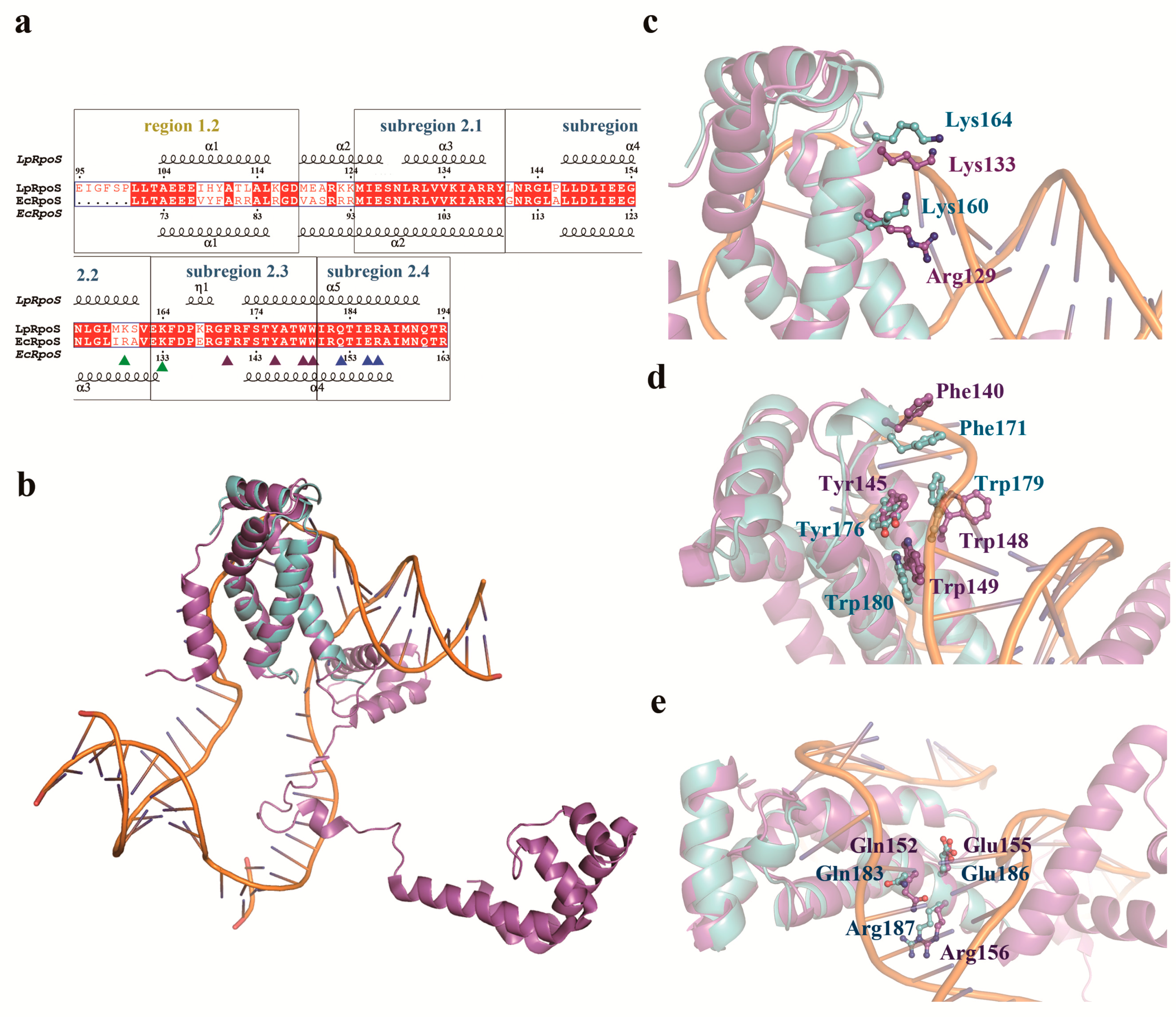
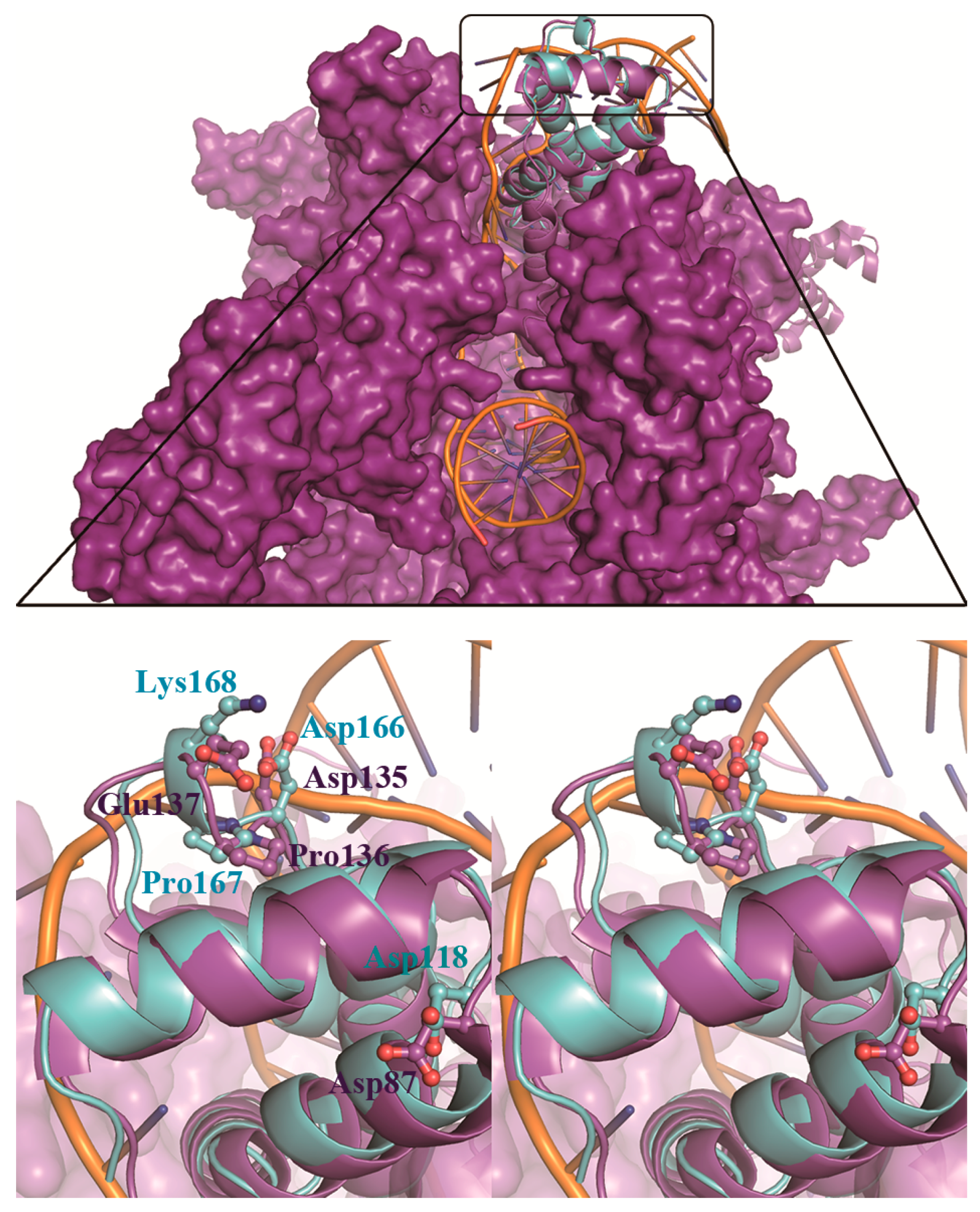
3.3. LpRpoS Show a Different Regulatory Factor Binding Site from EcRpoS
4. Discussion
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Dong, T.; Schellhorn, H.E. Role of rpos in virulence of pathogens. Infect. Immun. 2010, 78, 887–897. [Google Scholar] [CrossRef] [PubMed]
- Isberg, R.R.; O’Connor, T.J.; Heidtman, M. The legionella pneumophila replication vacuole: Making a cosy niche inside host cells. Nat. Rev. Microbiol. 2009, 7, 13–24. [Google Scholar] [CrossRef] [PubMed]
- Abu-Zant, A.; Asare, R.; Graham, J.E.; Abu Kwaik, Y. Role for rpos but not rela of legionella pneumophila in modulation of phagosome biogenesis and adaptation to the phagosomal microenvironment. Infect. Immun. 2006, 74, 3021–3026. [Google Scholar] [CrossRef] [PubMed]
- Bachman, M.A.; Swanson, M.S. Genetic evidence that legionella pneumophila rpos modulates expression of the transmission phenotype in both the exponential phase and the stationary phase. Infect. Immun. 2004, 72, 2468–2476. [Google Scholar] [CrossRef] [PubMed]
- Gruber, T.M.; Gross, C.A. Multiple sigma subunits and the partitioning of bacterial transcription space. Annu. Rev. Microbiol. 2003, 57, 441–466. [Google Scholar] [CrossRef] [PubMed]
- Haugen, S.P.; Ross, W.; Manrique, M.; Gourse, R.L. Fine structure of the promoter-sigma region 1.2 interaction. Proc. Natl. Acad. Sci. USA 2008, 105, 3292–3297. [Google Scholar] [CrossRef] [PubMed]
- Feklistov, A.; Darst, S.A. Structural basis for promoter-10 element recognition by the bacterial rna polymerase sigma subunit. Cell 2011, 147, 1257–1269. [Google Scholar] [CrossRef] [PubMed]
- Campbell, E.A.; Muzzin, O.; Chlenov, M.; Sun, J.L.; Olson, C.A.; Weinman, O.; Trester-Zedlitz, M.L.; Darst, S.A. Structure of the bacterial rna polymerase promoter specificity sigma subunit. Mol. Cell 2002, 9, 527–539. [Google Scholar] [CrossRef]
- Maeda, H.; Fujita, N.; Ishihama, A. Competition among seven escherichia coli sigma subunits: Relative binding affinities to the core rna polymerase. Nucleic Acids Res. 2000, 28, 3497–3503. [Google Scholar] [CrossRef] [PubMed]
- England, P.; Westblade, L.F.; Karimova, G.; Robbe-Saule, V.; Norel, F.; Kolb, A. Binding of the unorthodox transcription activator, crl, to the components of the transcription machinery. J. Biol. Chem. 2008, 283, 33455–33464. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Pan, Q.; Yang, L.; Zhou, H.; Xu, C.; Yu, F.; Wang, Q.; Huang, S.; He, J. Automatic crystal centring procedure at the ssrf macromolecular crystallography beamline. J. Synchrotron Radiat. 2016, 23, 1323–1332. [Google Scholar] [CrossRef] [PubMed]
- Battye, T.G.; Kontogiannis, L.; Johnson, O.; Powell, H.R.; Leslie, A.G. iMOSFLM: A new graphical interface for diffraction-image processing with MOSFLM. Acta Crystallogr. D Biol. Crystallogr. 2011, 67, 271–281. [Google Scholar] [CrossRef] [PubMed]
- Murakami, K.S. X-ray crystal structure of escherichia coli rna polymerase sigma(70) holoenzyme. J. Biol. Chem. 2013, 288, 9126–9134. [Google Scholar] [CrossRef] [PubMed]
- McCoy, A.J.; Grosse-Kunstleve, R.W.; Adams, P.D.; Winn, M.D.; Storoni, L.C.; Read, R.J. Phaser crystallographic software. J. Appl. Crystallogr. 2007, 40, 658–674. [Google Scholar] [CrossRef] [PubMed]
- Murshudov, G.N.; Skubak, P.; Lebedev, A.A.; Pannu, N.S.; Steiner, R.A.; Nicholls, R.A.; Winn, M.D.; Long, F.; Vagin, A.A. Refmac5 for the refinement of macromolecular crystal structures. Acta Crystallogr. D Biol. Crystallogr. 2011, 67, 355–367. [Google Scholar] [CrossRef] [PubMed]
- Emsley, P.; Cowtan, K. Coot: Model-building tools for molecular graphics. Acta Crystallogr. D Biol. Crystallogr. 2004, 60, 2126–2132. [Google Scholar] [CrossRef] [PubMed]
- Adams, P.D.; Afonine, P.V.; Bunkoczi, G.; Chen, V.B.; Davis, I.W.; Echols, N.; Headd, J.J.; Hung, L.W.; Kapral, G.J.; Grosse-Kunstleve, R.W.; et al. Phenix: A comprehensive python-based system for macromolecular structure solution. Acta Crystallogr. D Biol. Crystallogr. 2010, 66, 213–221. [Google Scholar] [CrossRef] [PubMed]
- Davis, I.W.; Leaver-Fay, A.; Chen, V.B.; Block, J.N.; Kapral, G.J.; Wang, X.; Murray, L.W.; Arendall, W.B.; Snoeyink, J.; Richardson, J.S.; et al. Molprobity: All-atom contacts and structure validation for proteins and nucleic acids. Nucleic Acids Res. 2007, 35, W375–W383. [Google Scholar] [CrossRef] [PubMed]
- Corpet, F. Multiple sequence alignment with hierarchical clustering. Nucleic Acids Res. 1988, 16, 10881–10890. [Google Scholar] [CrossRef] [PubMed]
- Gouet, P.; Courcelle, E.; Stuart, D.I.; Metoz, F. Espript: Analysis of multiple sequence alignments in postscript. Bioinformatics 1999, 15, 305–308. [Google Scholar] [CrossRef] [PubMed]
- Krissinel, E.; Henrick, K. Inference of macromolecular assemblies from crystalline state. J. Mol. Biol. 2007, 372, 774–797. [Google Scholar] [CrossRef] [PubMed]
- Holm, L.; Sander, C. Protein structure comparison by alignment of distance matrices. J. Mol. Biol. 1993, 233, 123–138. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Zuo, Y.H.; Steitz, T.A. Structures of e-coli sigma(s)-transcription initiation complexes provide new insights into polymerase mechanism. Proc. Natl. Acad. Sci. USA 2016, 113, 4051–4056. [Google Scholar] [CrossRef] [PubMed]
- Tomsic, M.; Tsujikawa, L.; Panaghie, G.; Wang, Y.; Azok, J.; de Haseth, P.L. Different roles for basic and aromatic amino acids in conserved region 2 of escherichia coli sigma(70) in the nucleation and maintenance of the single-stranded DNA bubble in open rna polymerase-promoter complexes. J. Biol. Chem. 2001, 276, 31891–31896. [Google Scholar] [CrossRef] [PubMed]
- Osterberg, S.; del Peso-Santos, T.; Shingler, V. Regulation of alternative sigma factor use. Annu. Rev. Microbiol. 2011, 65, 37–55. [Google Scholar] [CrossRef] [PubMed]
- Banta, A.B.; Chumanov, R.S.; Yuan, A.H.; Lin, H.L.; Campbell, E.A.; Burgess, R.R.; Gourse, R.L. Key features of sigma(s) required for specific recognition by crl, a transcription factor promoting assembly of rna polymerase holoenzyme. Proc. Natl. Acad. Sci. USA 2013, 110, 15955–15960. [Google Scholar] [CrossRef] [PubMed]
- Cavaliere, P.; Levi-Acobas, F.; Mayer, C.; Saul, F.A.; England, P.; Weber, P.; Raynal, B.; Monteil, V.; Bellalou, J.; Haouz, A.; et al. Structural and functional features of crl proteins and identification of conserved surface residues required for interaction with the rpos/sigmas subunit of rna polymerase. Biochem. J. 2014, 463, 215–224. [Google Scholar] [CrossRef] [PubMed]
- Cavaliere, P.; Sizun, C.; Levi-Acobas, F.; Nowakowski, M.; Monteil, V.; Bontems, F.; Bellalou, J.; Mayer, C.; Norel, F. Binding interface between the salmonella sigma(s)/rpos subunit of rna polymerase and crl: Hints from bacterial species lacking crl. Sci. Rep. 2015, 5, 13564. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| LpRpoS 95–195 | |
|---|---|
| SSRF beamline | BL17U |
| Wavelength (Å) | 0.97915 |
| Space group | P212121 |
| Molecules/ASU | 2 |
| Cell parameters | |
| a/b/c (Å) | 37.93/68.84/70.38 |
| á/â/ã (°) | 90.00/90.00/90.00 |
| Resolution range (Å) | 33.4–1.45 (1.53–1.45) |
| No. of unique reflections | 30,122 (3739) |
| Corresponding % solvent | 39.8 |
| Rpim a (%) | 3.8 (24.4) |
| CC1/2 | 0.99 (0.86) |
| Average I/ó(I) | 12 (3.1) |
| Redundancy | 13.8 (14) |
| Completeness (%) | 90.7 (78.5) |
| Refinement statistics | |
| Resolution range (Å) | 20–1.45 (1.50–1.45) |
| R-factor b (%) | 13.7 (20.8) |
| R-free c (%) | 17.7 (22.9) |
| RMSD d bond length (Å) | 0.007 |
| RMSD bond angles (°) | 1.08 |
| No. of non-H atoms | |
| Protein | 1616 |
| Solvent | 192 |
| Average of B factors (Å2) | |
| Protein | 21.5 |
| Solvent | 36.0 |
| Ramachandran plot f (%) | |
| Ramachandran favored | 99 |
| Ramachandran Outliers | 0 |
| PDB ID code | 5H6X |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, N.; Chen, X.; Gong, X.; Li, T.; Xie, Z.; Hameed, M.F.; Wang, M.; Ge, H. High-Resolution Crystal Structure of RpoS Fragment including a Partial Region 1.2 and Region 2 from the Intracellular Pathogen Legionella pneumophila. Crystals 2018, 8, 54. https://doi.org/10.3390/cryst8020054
Zhang N, Chen X, Gong X, Li T, Xie Z, Hameed MF, Wang M, Ge H. High-Resolution Crystal Structure of RpoS Fragment including a Partial Region 1.2 and Region 2 from the Intracellular Pathogen Legionella pneumophila. Crystals. 2018; 8(2):54. https://doi.org/10.3390/cryst8020054
Chicago/Turabian StyleZhang, Nannan, Xiaofang Chen, Xiaojian Gong, Tao Li, Zhiyuan Xie, Muhammad Fazal Hameed, Mingzhu Wang, and Honghua Ge. 2018. "High-Resolution Crystal Structure of RpoS Fragment including a Partial Region 1.2 and Region 2 from the Intracellular Pathogen Legionella pneumophila" Crystals 8, no. 2: 54. https://doi.org/10.3390/cryst8020054
APA StyleZhang, N., Chen, X., Gong, X., Li, T., Xie, Z., Hameed, M. F., Wang, M., & Ge, H. (2018). High-Resolution Crystal Structure of RpoS Fragment including a Partial Region 1.2 and Region 2 from the Intracellular Pathogen Legionella pneumophila. Crystals, 8(2), 54. https://doi.org/10.3390/cryst8020054