Molecular Dynamics Analysis of Synergistic Effects of Ions and Winter Flounder Antifreeze Protein Adjacent to Ice-Solution Surfaces


Abstract
:1. Introduction
2. Computational Procedures
2.1. Assumptions
2.2. Governing Equations
2.3. Temperature Scaling
2.4. Production of Mixtures
2.4.1. In the Case of Ice Layers with Prism Faces
2.4.2. In the Cases of Ice Layers with Pyramidal Faces
2.5. Potential Functions of Water Molecules
2.6. Model of HPLC6
2.7. Model of Ions
2.8. Computational Conditions
3. Statistical Quantities
3.1. Radial Distribution Function (RDF)
3.2. Hydrogen Bond Correlation Function (HBCF)
3.3. Diffusion Coefficient
3.4. Tetrahedricity Parameter
4. Results and Discussion
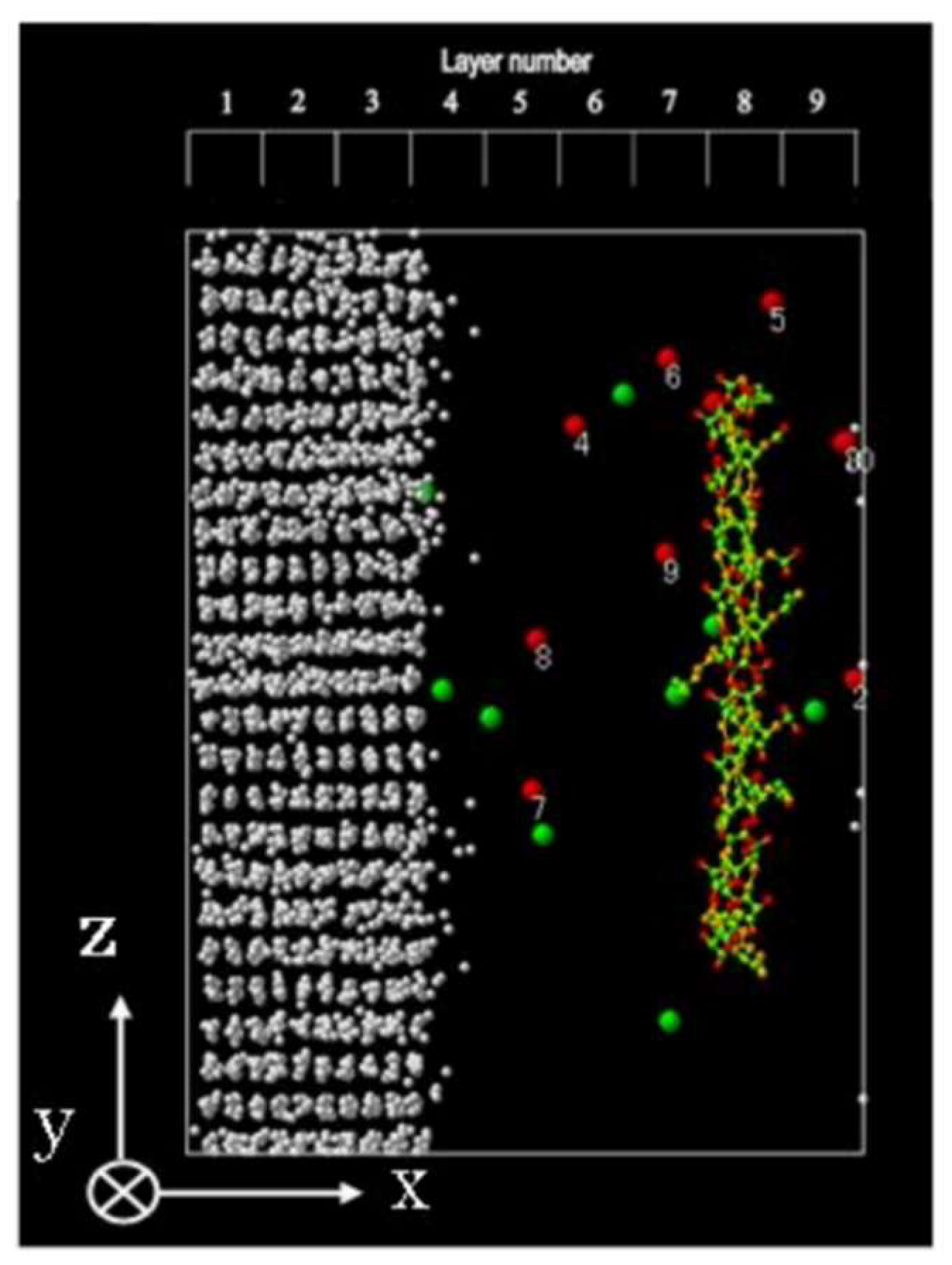
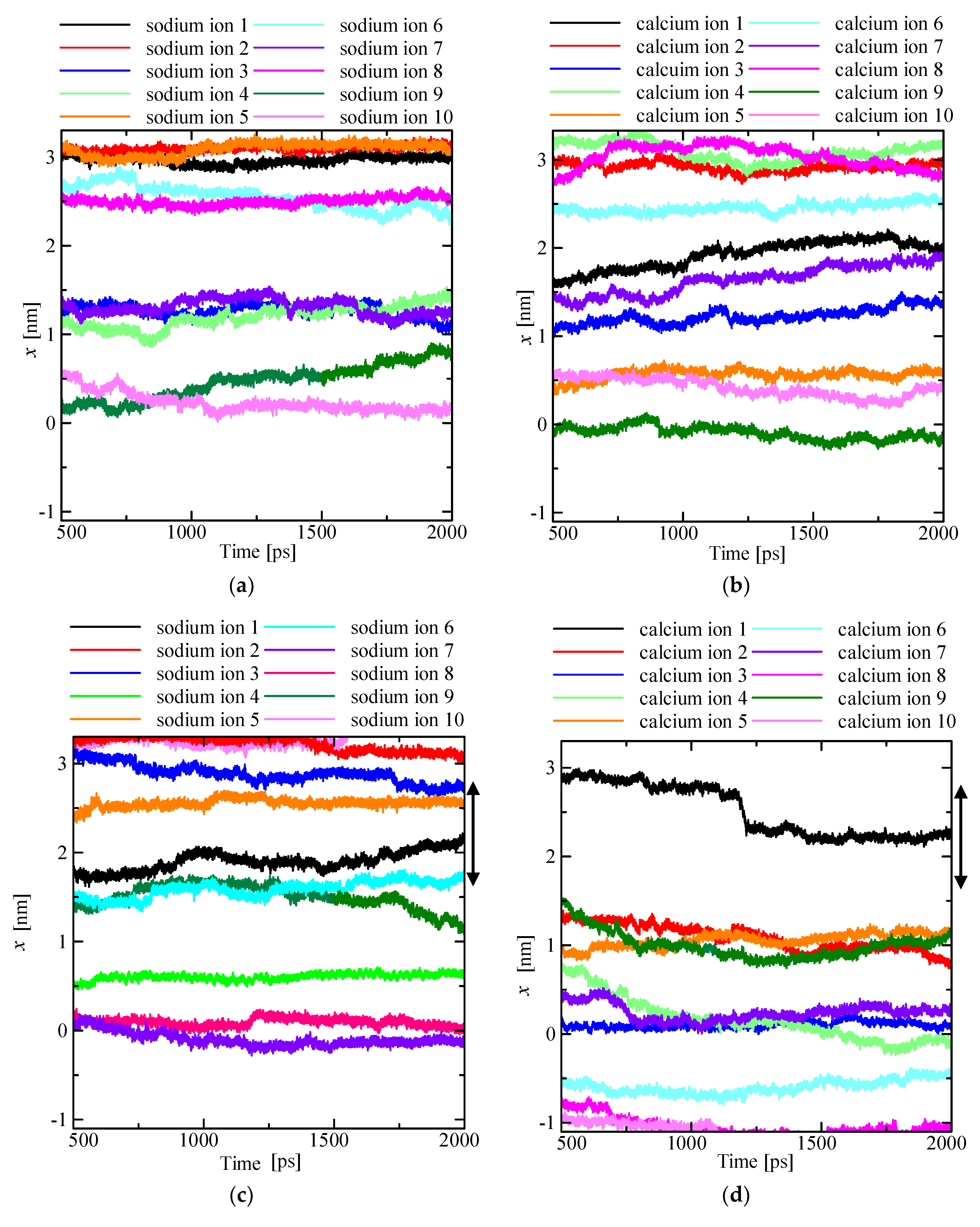
4.1. Time Changes in the Ion Positions
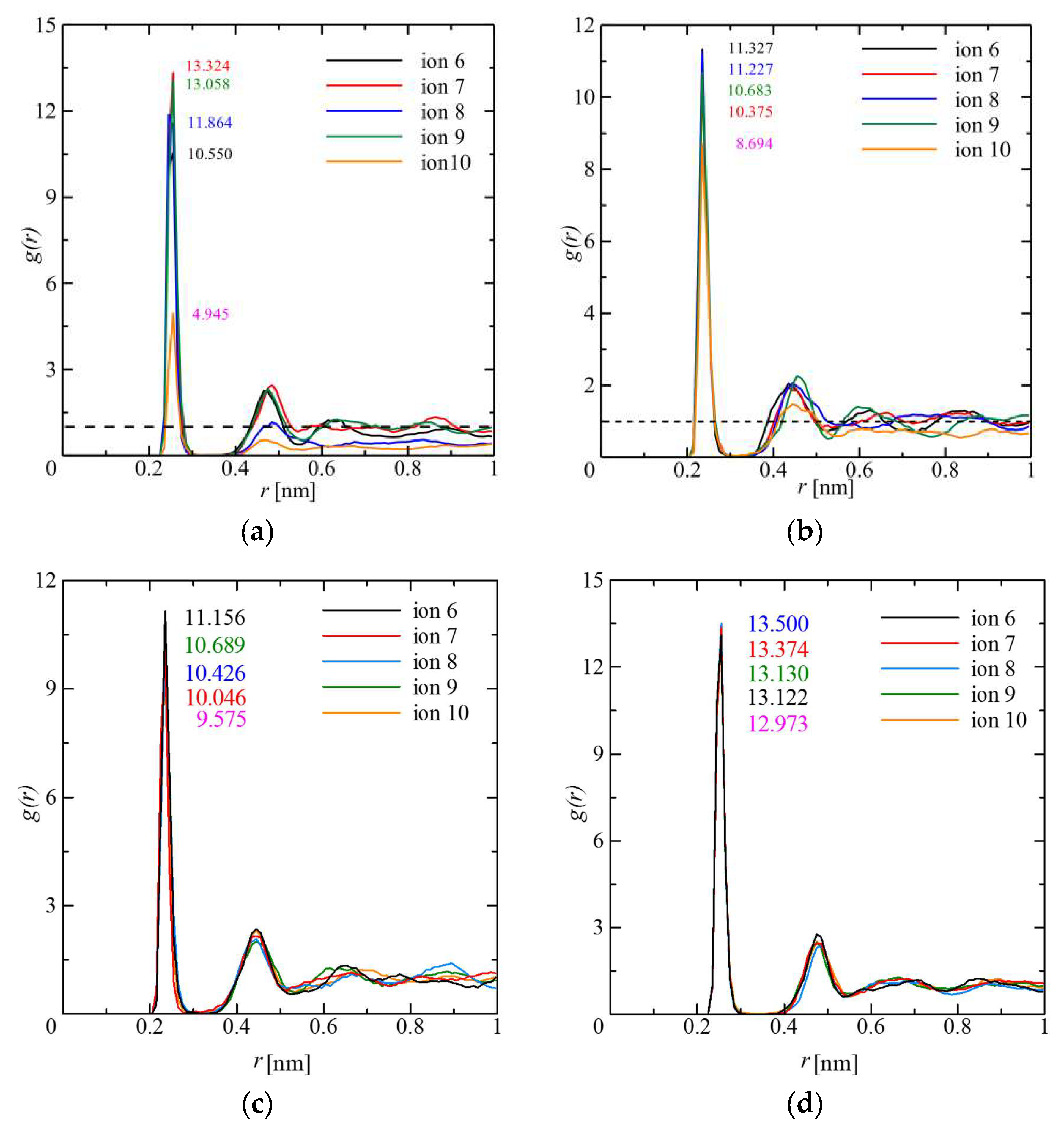
4.2. Radial Distribution Function (RDF)
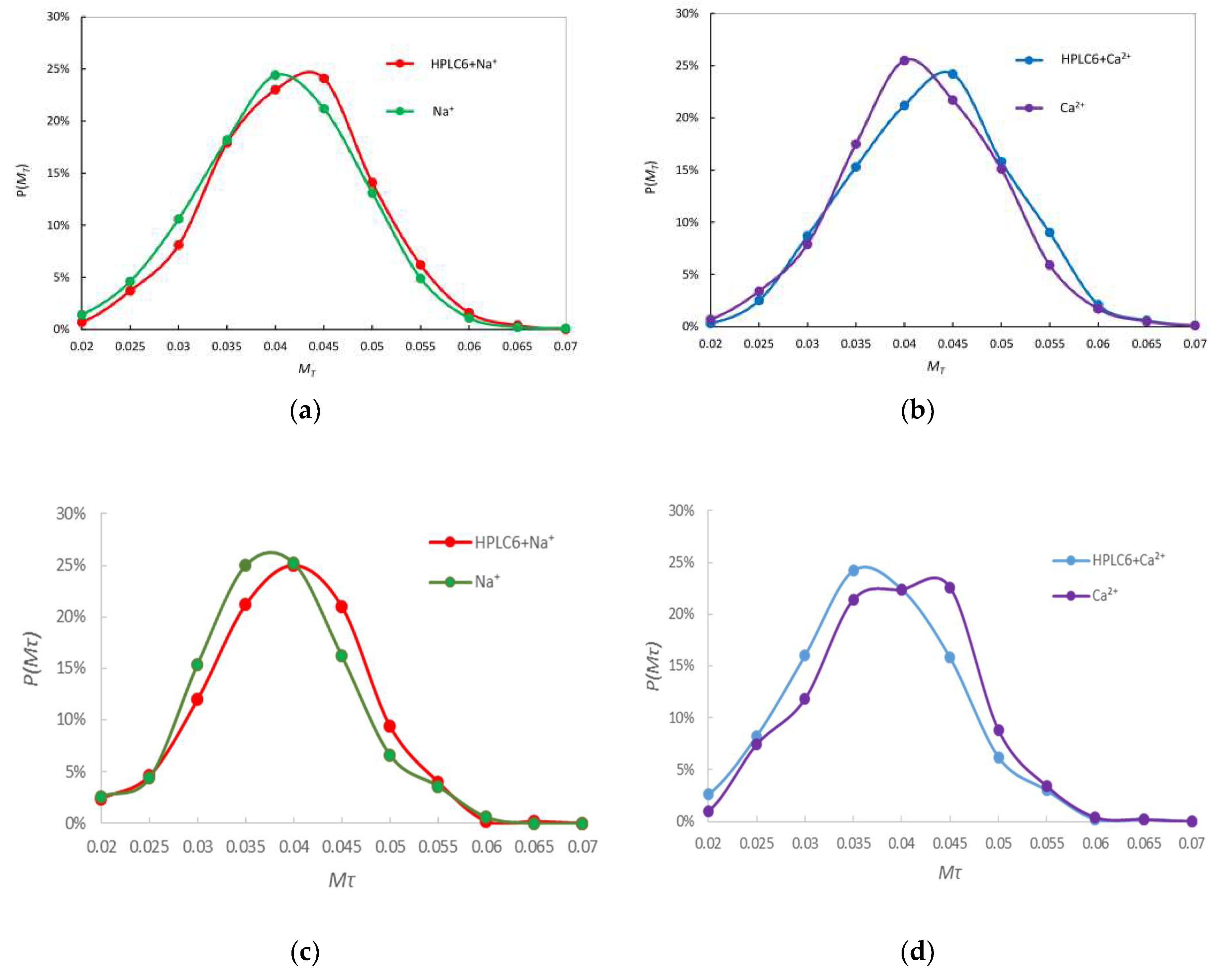
4.3. Tetrahedricity Parameter
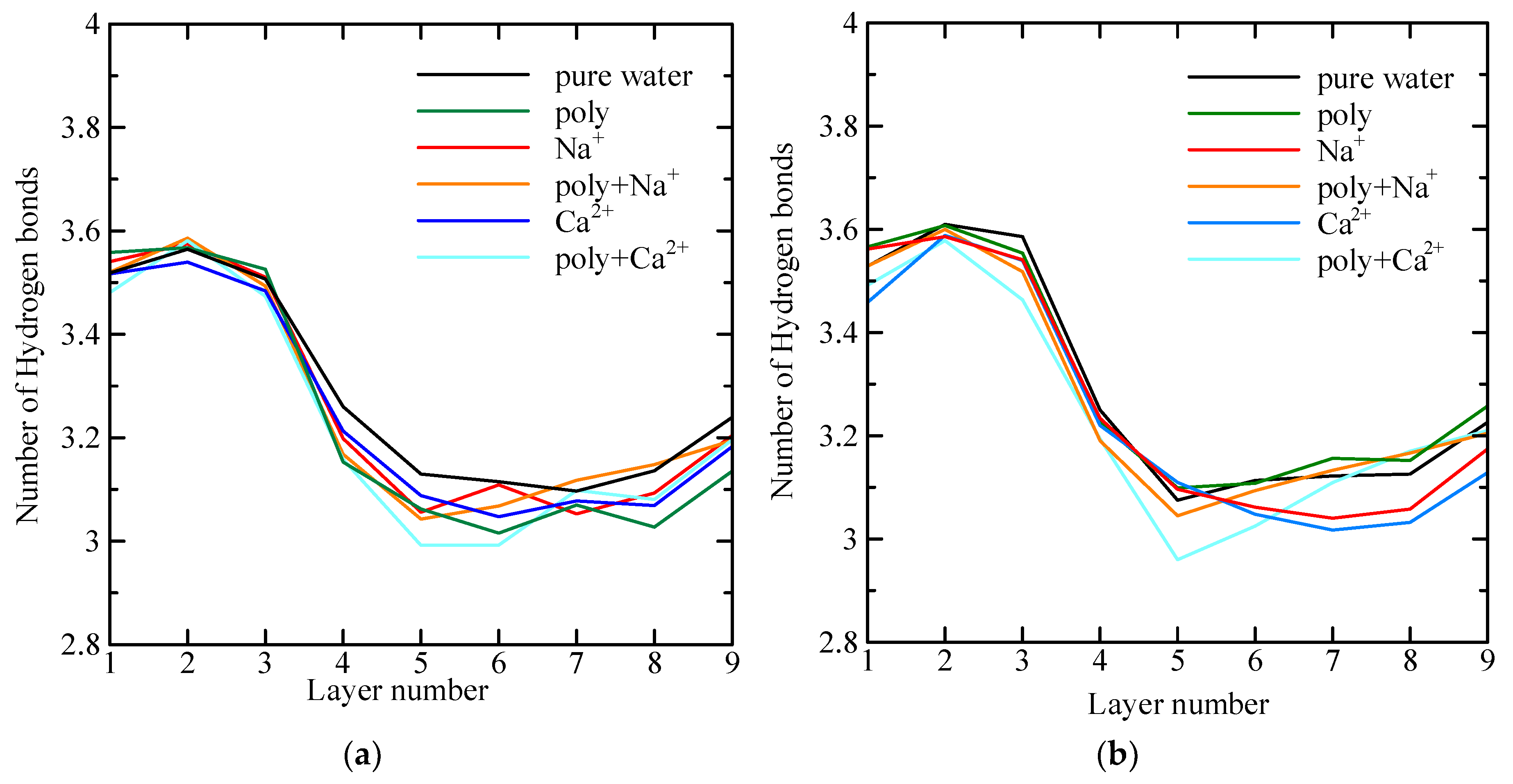
4.4. Average Number of Hydrogen Bonds
- (A)
- The distance between oxygen atoms is lower than 3.5 Å.
- (B)
- The angle between the line connecting two oxygen atoms and the line connecting one of the two oxygen atoms and a covalent-bonded hydrogen atom is within ±15°.
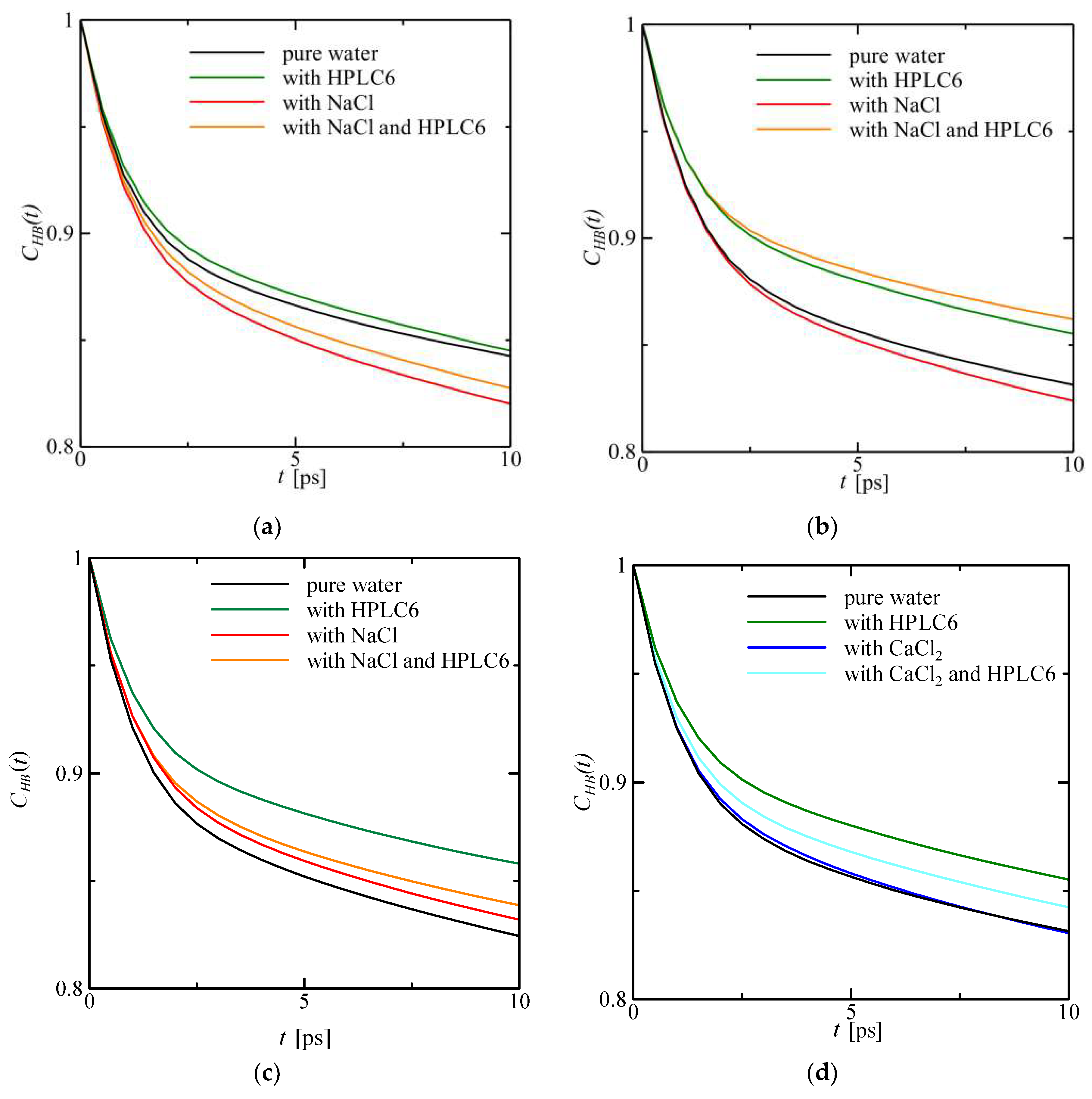
4.5. Hydrogen Bond Correlation Function (HBCF)
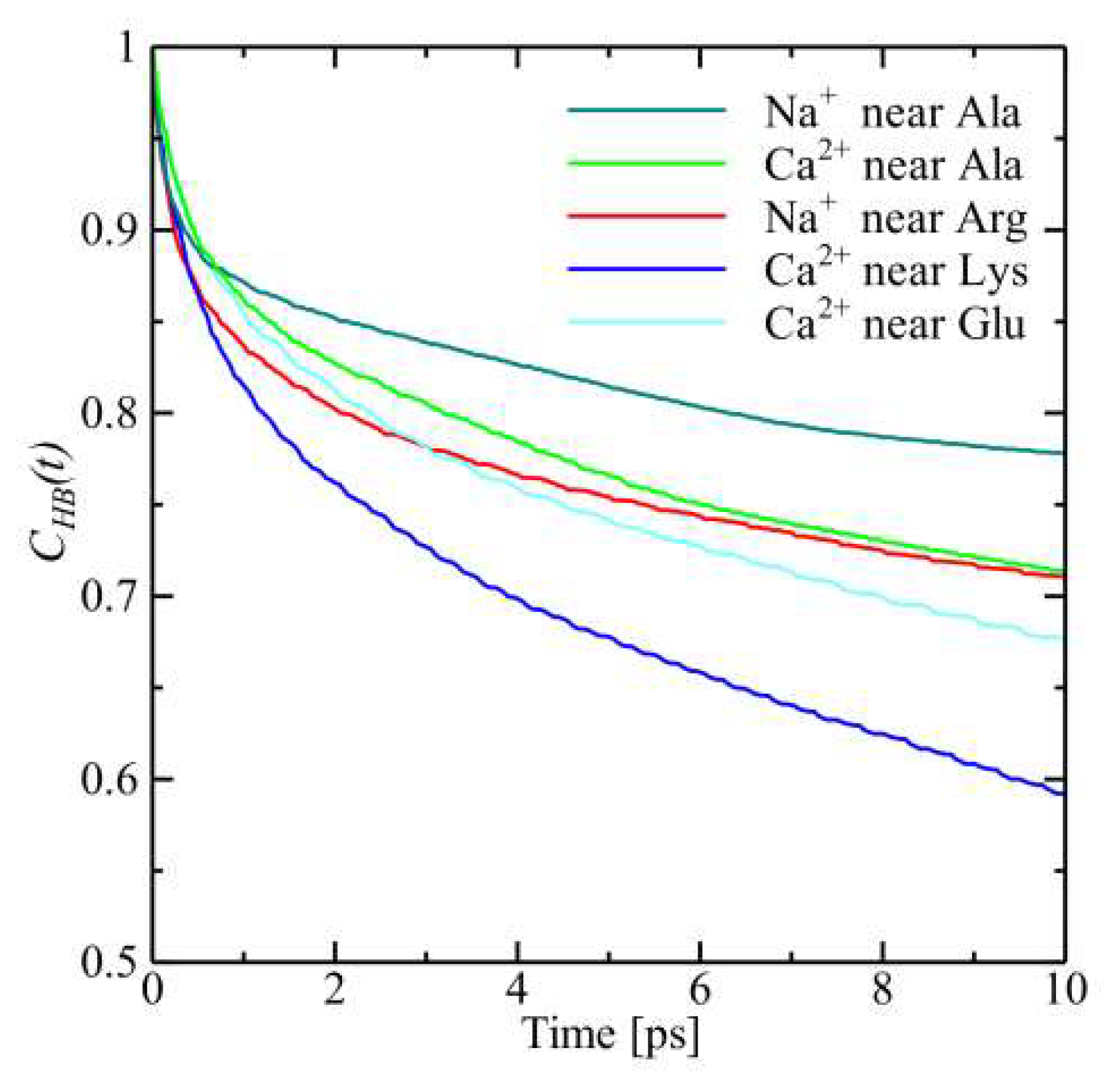
4.6. Interaction between Cations and Amino Acid Residues
4.6.1. Surface-Bound Water Molecules
4.6.2. Diffusion Coefficient
4.6.3. Hydrogen Bond Correlation Function (HBCF)
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ustun, N.S.; Turhan, S. Antifreeze proteins: Characteristics, function, mechanism of action, sources and application to foods. J. Food Process. Preserv. 2015, 39, 3189–3197. [Google Scholar] [CrossRef]
- Li, B.; Sun, D.-W. Novel methods for rapid freezing and thawing of foods—A review. J. Food Eng. 2002, 54, 175–182. [Google Scholar] [CrossRef]
- Amir, G.; Rubinsky, B.; Basheer, S.Y.; Horowitz, L.; Jonathan, L.; Feinberg, M.S.; Smolinsky, A.K.; Lavee, L. Improved viability and reduced apoptosis in sub-zero 21-hour preservation of transplanted rat hearts using anti-freeze proteins. J. Heart Lung Transpl. 2005, 24, 1915–1929. [Google Scholar] [CrossRef] [PubMed]
- Shitzer, A. Cryosurgery: Analysis and experimentation of cryoprobes in phase changing media. J. Heat Transf. 2011, 133. [Google Scholar] [CrossRef]
- Grandum, S.; Yabe, A.; Tanaka, M.; Takemura, F.; Nakagomi, K. Characteristics of ice slurry containing antifreeze protein for ice storage applications. J. Thermophys. Heat Transf. 1997, 11, 461–466. [Google Scholar] [CrossRef]
- Yang, D.S.C.; Sax, M.; Chakrabatty, A.; Hew, C.L. Crystal structure of an antifreeze polypeptide and its mechanistic implications. Nature 1988, 333, 232–237. [Google Scholar] [CrossRef] [PubMed]
- Chao, H.; Houston, M.E., Jr.; Hodges, R.S.; Kay, C.M.; Sykes, B.D.; Loewen, M.C.; Davies, P.L.; Sönnichsen, F.D. A diminished role for hydrogen bonds in antifreeze protein binding to ice. Biochemistry 1997, 36, 14652–14660. [Google Scholar] [CrossRef] [PubMed]
- Sicheri, F.; Yang, D.S.C. Ice-binding structure and mechanism of an antifreeze protein from winter flounder. Nature 1995, 375, 427–431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haymet, A.D.J.; Ward, L.G.; Harding, M.M. Winter flounder “antifreeze” proteins: Synthesis and ice growth inhibition of analogues that probe the relative importance of hydrophobic and hydrogen-bonding interactions. J. Am. Chem. Soc. 1999, 121, 941–948. [Google Scholar] [CrossRef]
- Baardsnes, J.; Kondejewski, L.H.; Hodges, R.S.; Chao, H.; Kay, C.; Davies, P.L. New ice-binding face for type I antifreeze protein. FEBS Lett. 1999, 463, 87–91. [Google Scholar] [CrossRef] [Green Version]
- Davies, P.L.; Baardsnes, J.; Kuiper, M.J.; Walker, V.K. Structure and function of antifreeze proteins. Philos. Trans. R. Soc. Lond. B 2002, 357, 927–935. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jorov, A.; Zhorov, B.S.; Yang, D.S.C. Theoretical study of interaction of winter flounder antifreeze protein with ice. Protein Sci. 2004, 13, 1524–1537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dalal, P.; Knickelbein, J.; Haymet, A.D.J.; Sönnichsen, F.D.; Madura, J. Hydrogen bond analysis of type 1 antifreeze protein in water and the ice/water interface. PhysChemComm 2001, 4, 32–36. [Google Scholar] [CrossRef]
- Wierzbicki, A.; Dalal, P.; Cheatham, T.E., III; Knickelbein, J.E.; Haymet, A.D.J.; Madura, J.D. Antifreeze proteins at the ice/water interface: Three calculated discriminating properties for orientation of type I proteins. Biophys. J. 2007, 93, 1442–1451. [Google Scholar] [CrossRef] [PubMed]
- Nada, H.; Furukawa, Y. Growth inhibition mechanism of an ice-water interface by a mutant of winter flounder antifreeze protein: A molecular dynamics study. J. Phys. Chem. B 2008, 112, 7111–7119. [Google Scholar] [CrossRef] [PubMed]
- Todde, G.; Hovmöller, S.; Laaksonen, A. Influence of antifreeze proteins on the ice/water interface. J. Phys. Chem. B 2015, 119, 3407–3413. [Google Scholar] [CrossRef] [PubMed]
- Evans, R.P.; Hobbs, R.S.; Goddard, S.V.; Fletcher, G.L. The importance of dissolved salts to the in vivo efficacy of antifreeze proteins. Comp. Biochem. Physiol. Part A 2007, 148, 556–561. [Google Scholar] [CrossRef] [PubMed]
- Kristiansen, E.; Pedersen, S.A.; Zachariassen, K.E. Salt-induced enhancement of antifreeze protein activity: A salting-out effect. Cryobiology 2008, 57, 122–129. [Google Scholar] [CrossRef] [PubMed]
- Hagiwara, Y.; Aomatsu, H. Supercooling enhancement by adding antifreeze protein and ions to water in a narrow space. Int. J. Heat Mass Transf. 2015, 86, 55–64. [Google Scholar] [CrossRef]
- Hayakari, K.; Hagiwara, Y. Effects of ions on winter flounder antifreeze protein and water molecules near an ice/water interface. Mol. Simul. 2012, 38, 26–37. [Google Scholar] [CrossRef]
- Nada, H.; Furukawa, Y. Anisotropy in growth kinetics at interfaces between proton-disordered hexagonal ice and water: A molecular dynamics study using the six-site model of H2O. J. Cryst. Growth 2005, 283, 242–256. [Google Scholar] [CrossRef]
- Yokoyama, T.; Hagiwara, Y. Molecular dynamics simulation for the mixture of water and an ice nucleus. Mol. Simul. 2003, 29, 235–248. [Google Scholar] [CrossRef]
- Iwasaki, K.; Hagiwara, Y. Inhibition of ice nucleus growth in water by alanine dipeptide. Mol. Simul. 2004, 30, 487–500. [Google Scholar] [CrossRef]
- Nobekawa, T.; Taniguchi, H.; Hagiwara, Y. Interaction between a twelve-residue segment of antifreeze protein type I, or its mutants, and water molecules. Mol. Simul. 2008, 34, 309–325. [Google Scholar] [CrossRef]
- Nobekawa, T.; Hagiwara, Y. Interaction among the twelve-residue segment of antifreeze protein type I, or its mutants, water and a hexagonal ice crystal. Mol. Simul. 2008, 34, 591–610. [Google Scholar] [CrossRef]
- Nohara, Y.; Hagiwara, Y. Diffusion of cations in salt solutions between ice walls. Mol. Simul. 2015, 41, 980–985. [Google Scholar] [CrossRef]
- Gear, C.W. Numerical Initial Value Problems in Ordinary Differential Equations; Prentice-Hall: Upper Saddle River, NJ, USA, 1971. [Google Scholar]
- Nosé, S. A molecular dynamics method for simulations in the canonical ensemble. Mol. Phys. 1984, 52, 255–268. [Google Scholar] [CrossRef]
- Grandum, S.; Yabe, A.; Nakagomi, K.; Tanaka, M.; Takemura, F.; Kobayashi, Y.; Frivik, P.-E. Analysis of ice crystal growth for a crystal surface containing adsorbed antifreeze proteins. J. Cryst. Growth 1999, 205, 382–390. [Google Scholar] [CrossRef]
- Abascal, J.L.F.; Sanz, E.; Fernández, R.G.; Vega, C. A potential model for the study of ices and amorphous water: TIP4P/Ice. J. Chem. Phys. 2005, 122. [Google Scholar] [CrossRef] [PubMed]
- Jorgensen, W.J.; Chandrasekhar, J.; Madura, J.D.; Impey, R.W.; Klein, M.L. Comparison of simple potential functions for simulating liquid water. J. Chem. Phys. 1983, 79, 926–935. [Google Scholar] [CrossRef]
- Frenkel, D.; Smit, B. Understanding Molecular Simulation; Academic Press: San Diego, CA, USA, 1996. [Google Scholar]
- Ryckaert, J.P.; Ciccotti, G.; Berendsen, H.J.C. Numerical integration of the Cartesian equations of motion of a system with constraints: Molecular dynamics of n-alkanes. J. Comput. Phys. 1977, 23, 327–341. [Google Scholar] [CrossRef]
- Cheng, A.; Merz, K.M., Jr. Ice-binding mechanism of winter flounder antifreeze proteins. Biophys. J. 1997, 73, 2851–2873. [Google Scholar] [CrossRef] [Green Version]
- Jorgensen, W.L.; Tirado-Rives, J. The OPLS potential functions for proteins: Energy minimizations for crystals of cyclic peptides and crambin. J. Am. Chem. Soc. 1988, 110, 1657–1666. [Google Scholar] [CrossRef] [PubMed]
- Chowdhuri, S.; Chandra, A. Molecular dynamics simulations of aqueous NaCl and KCl solutions: Effects of ion concentration on the single-particle pair, and collective dynamical properties of ions and water molecules. J. Chem. Phys. 2001, 115, 3732–3741. [Google Scholar] [CrossRef]
- Chialvo, A.A. The structure of CaCl2 aqueous solutions over a wide range of concentration. Interpretation of diffraction experiments via molecular simulation. J. Chem. Phys. 2003, 119, 8052–8061. [Google Scholar] [CrossRef]
- Allen, M.P.; Tildesley, D.J. Computer Simulation of Liquid; Oxford Science: Oxford, UK, 1987. [Google Scholar]
- Habasaki, J.; Ngai, K.L. Heterogeneous dynamics of ionic liquids from molecular dynamics simulations. J. Chem. Phys. 2008, 129, 194501. [Google Scholar] [CrossRef] [PubMed]
- Holzmann, J.; Ludwig, R.; Geiger, A.; Paschek, D. Pressure and salt effects in simulated water: Two sides of the same coin? Angew. Chem. Int. Ed. 2007, 46, 8907–8911. [Google Scholar] [CrossRef] [PubMed]
- Luzar, A.; Chandler, D. Hydrogen-bond kinetics in liquid water. Nature 1996, 379, 55–57. [Google Scholar] [CrossRef]
- Baştuğ, T.; Kuyucak, S. Temperature dependence of the transport coefficients of ions from molecular dynamics simulations. Chem. Phys. Lett. 2005, 408, 84–88. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Periods [ps] | 0–50 | 50–100 | 100–150 | 150–200 | 200–250 | 250–300 | 300–350 | 350– |
| Tpd for ice [K] | 10 | 50 | 100 | 150 | 200 | 250 | 260 | 250 |
| Tpd for water [K] | 370 | 370 | 340 | 310 | 290 | 280 | 260 | 250 |
| Periods [ps] | 0–50 | 50–60 | 60–65 | 65–70 | 70–75 | 75–80 | 80–85 | 85–90 | 90–95 | 95–100 |
| Tpd for ice [K] | 0 | 0 | 0 | 0 | 0 | 50 | 100 | 150 | 200 | 250 (200) |
| Tpd for water [K] | 370 | 350 | 330 | 310 | 290 | 280 | 280 | 280 | 280 | 280 |
| Primary Structure | Molecular Weight | |
|---|---|---|
| Winter flounder AFP | DTASDAAAAAALTAANAKAAAELTAANAAAAAAATAR | 3243 Da |
| Dalal [13] | Wierzbicki [14] | Nada [15] | Hayakari [20] | Todde [16] | Present Study | ||||
|---|---|---|---|---|---|---|---|---|---|
| Ice Face | Pyramidal | Pyramidal | Pyramidal | Pyramidal | Prism | Pyramidal | Prism | Pyramidal | |
| Domain size [nm3] | 8.4 × 4.0 × 7.8 | 5.6 × 12.0 × 7.6 | 7.0 × 5.7 × 7.0 | 6.7 × 7.0 × 8.8 | 6.1 × 21.7 × 5.9 | 18.1 × 6.7 × 7.5 | 6.7 × 4.7 × 8.8 | 6.7 × 7.0 × 8.8 | |
| Ice thickness [nm] | 1.4 | 3.8 | 1.4 | 2.3 | 11 | 9 | 2.3 | 2.3 | |
| Number of H2O molecules | liquid solid | 5484 2841 | 11,292 5313 | 6520 2160 | 8570 4185 | 12,288 12288 | 14,724 14076 | 5537 1 2880 | 8541 2 4185 |
| Number of AFP molecules | 1 | 2 | 1 | 1 | 1 | 1 | 1 | 1 | |
| AFP conc. [mg/mL] | 32 | 31 | 26 | 19 | 13 | 11 | 29 | 19 | |
| Number of cations | - | - | - | 24 | - | - | 10 | 15 | |
| Ion conc. [mol/L] | - | - | - | 0.15 | - | - | 0.10 | 0.10 | |
| Temperature [K] | 190 | 165 | 268 | 260 | 228 | 228 | 250 | 250 | |
| Ensemble | NVT | NVT | NPT | NV | NPT | NPT | NVT | NVT | |
| Potential functions | TIP3P | TIP3P | 6site | TIP4P/Ice | TIP4P | TIP4P | TIP4P/Ice | TIP4P/Ice | |
| Solution | NaCl | NaCl + HPLC6 | CaCl2 | CaCl2 + HPLC6 |
|---|---|---|---|---|
| 1000–1050 ps | 0.041 | 0.044 | 0.040 | 0.039 |
| 1500–1550 ps | 0.038 | 0.039 | 0.039 | 0.040 |
| 1950–2000 ps | 0.038 | 0.039 | 0.039 | 0.037 |
| Na+ | Ca2+ | |||||
|---|---|---|---|---|---|---|
| Near Arginine | Nohara | Baştuğ | Near Lysine | Near Glutamic Acid | Nohara | Baştuğ |
| 0.18 | 0.46 | 0.42 | 0.19 | 1.06 | 0.28 | 0.31 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yasui, T.; Kaijima, T.; Nishio, K.; Hagiwara, Y. Molecular Dynamics Analysis of Synergistic Effects of Ions and Winter Flounder Antifreeze Protein Adjacent to Ice-Solution Surfaces. Crystals 2018, 8, 302. https://doi.org/10.3390/cryst8070302
Yasui T, Kaijima T, Nishio K, Hagiwara Y. Molecular Dynamics Analysis of Synergistic Effects of Ions and Winter Flounder Antifreeze Protein Adjacent to Ice-Solution Surfaces. Crystals. 2018; 8(7):302. https://doi.org/10.3390/cryst8070302
Chicago/Turabian StyleYasui, Tatsuya, Tadashi Kaijima, Ken Nishio, and Yoshimichi Hagiwara. 2018. "Molecular Dynamics Analysis of Synergistic Effects of Ions and Winter Flounder Antifreeze Protein Adjacent to Ice-Solution Surfaces" Crystals 8, no. 7: 302. https://doi.org/10.3390/cryst8070302
APA StyleYasui, T., Kaijima, T., Nishio, K., & Hagiwara, Y. (2018). Molecular Dynamics Analysis of Synergistic Effects of Ions and Winter Flounder Antifreeze Protein Adjacent to Ice-Solution Surfaces. Crystals, 8(7), 302. https://doi.org/10.3390/cryst8070302