Identification of Structural Motifs Using Networks of Hydrogen-Bonded Base Interactions in RNA Crystallographic Structures


Abstract
:1. Introduction
2. Materials and Methods
2.1. Dataset
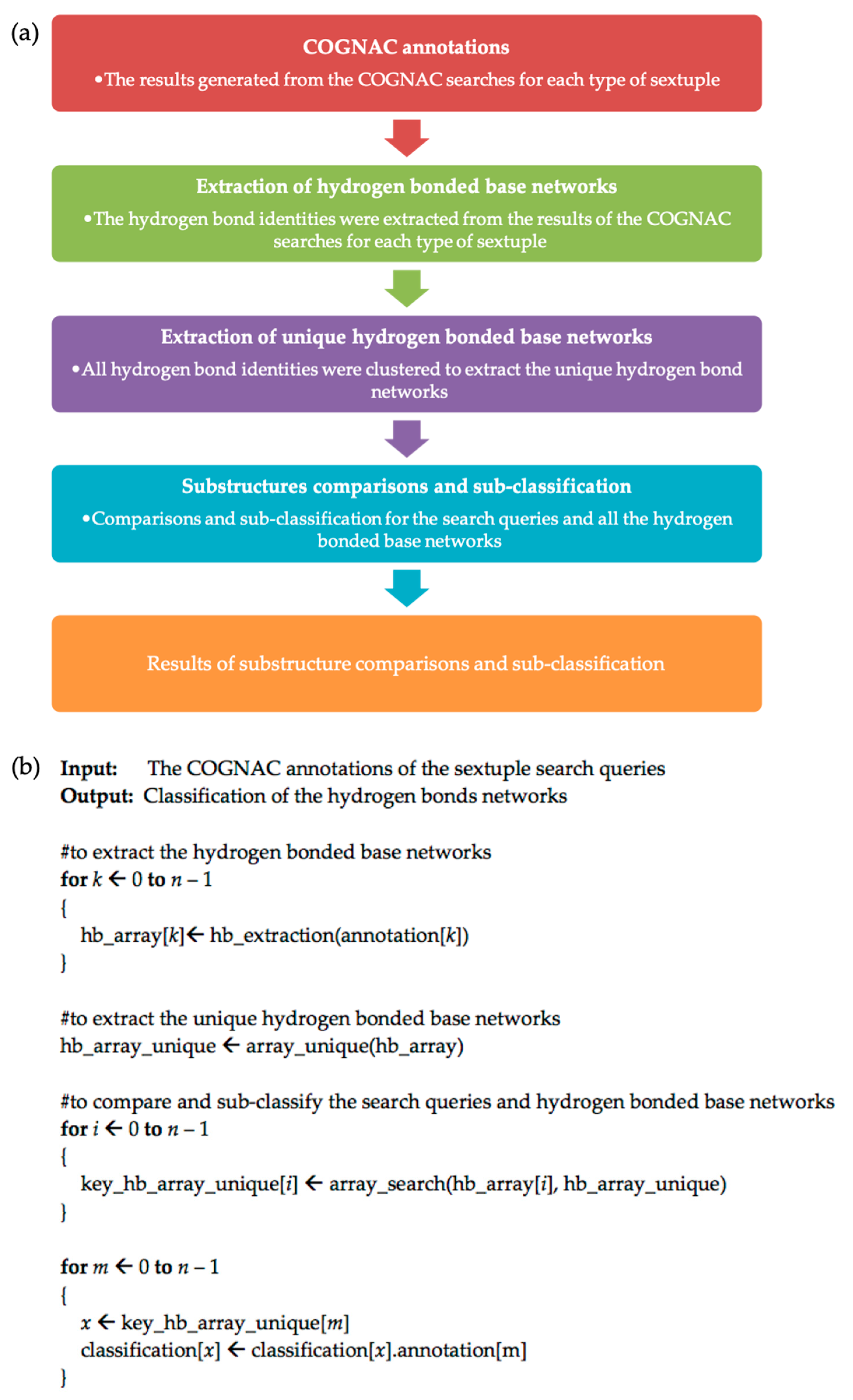
2.2. COGNAC Searches
2.3. Sextuple Sub-Classification
3. Results and Discussion
3.1. Filtering of COGNAC Searches
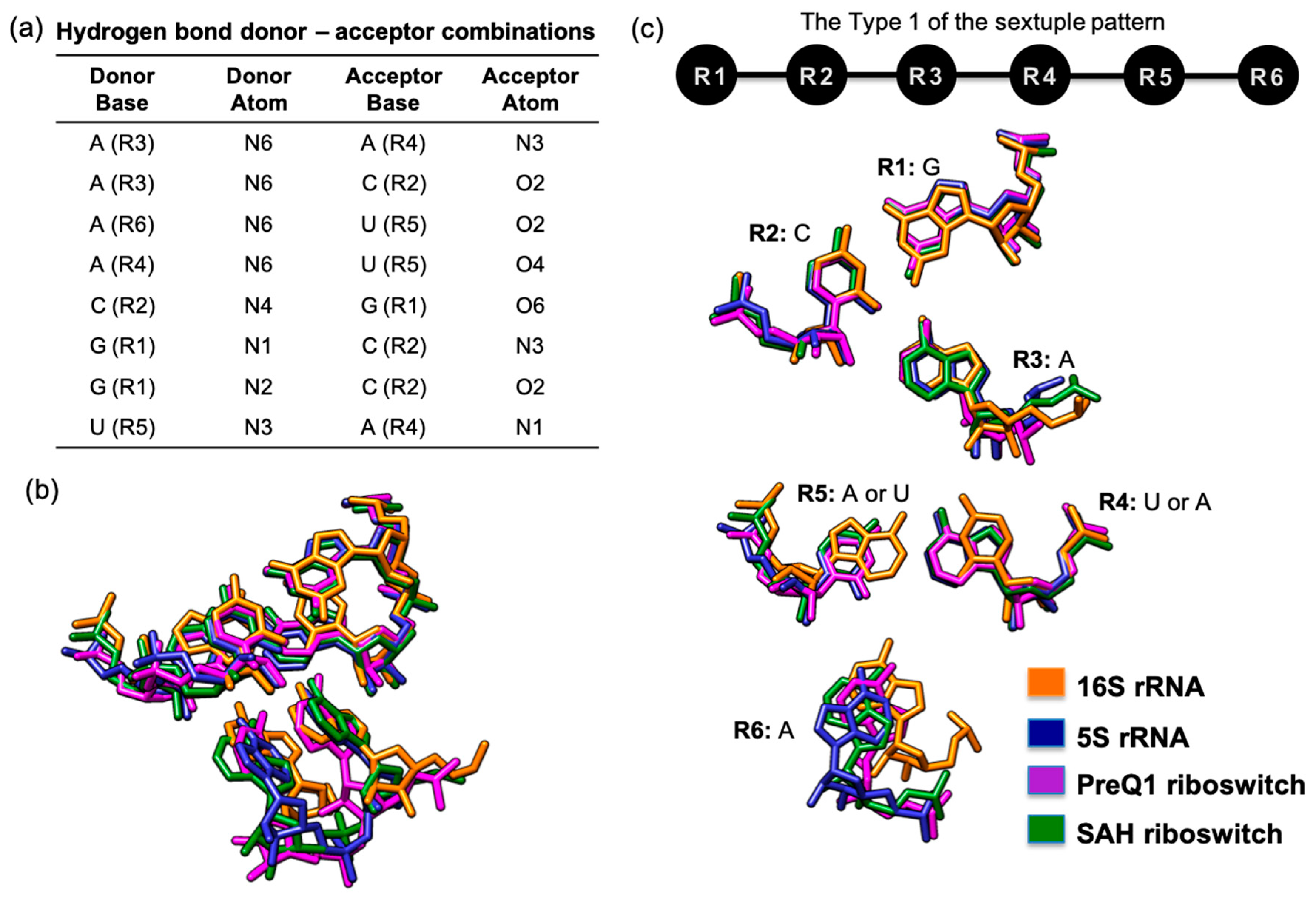
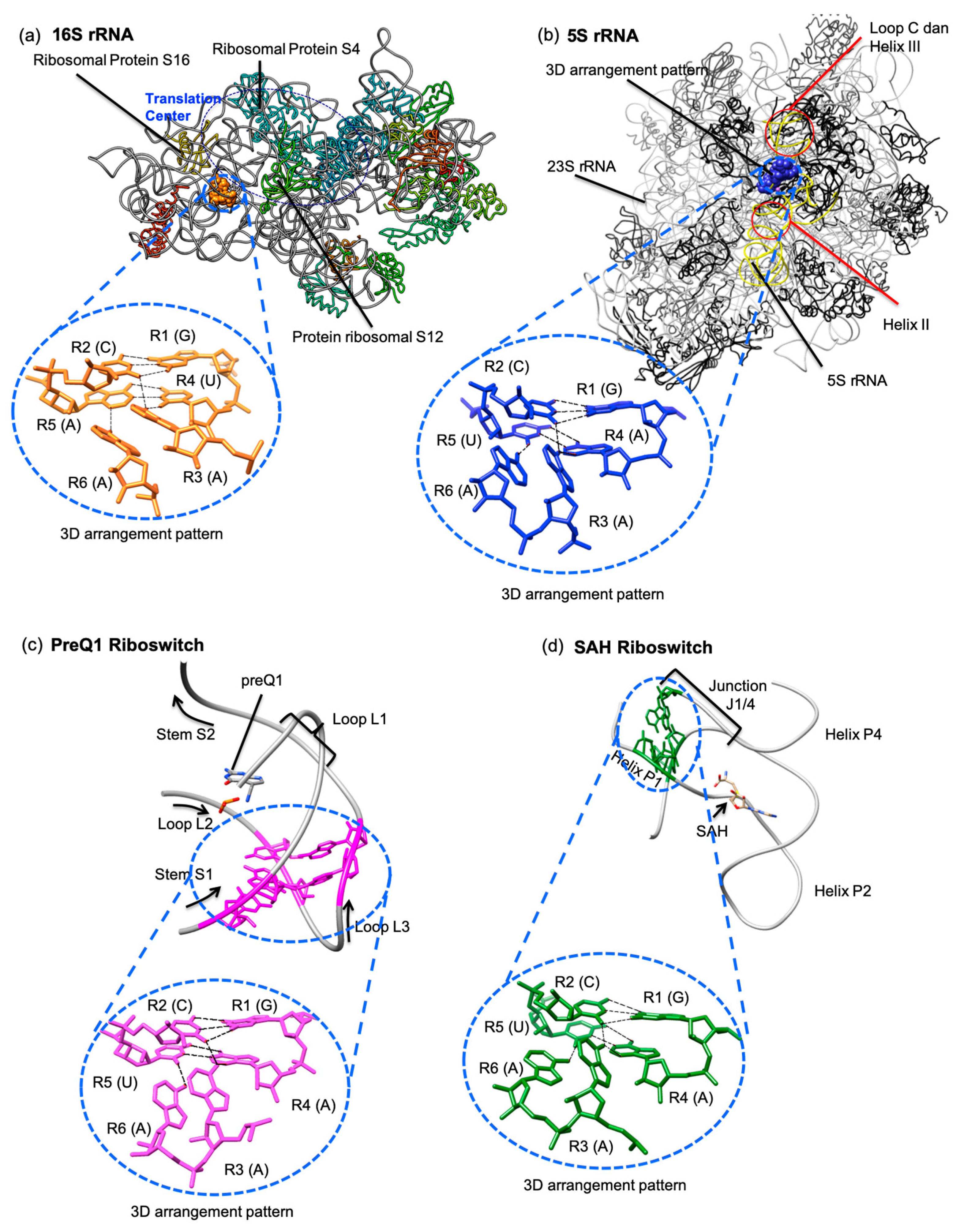
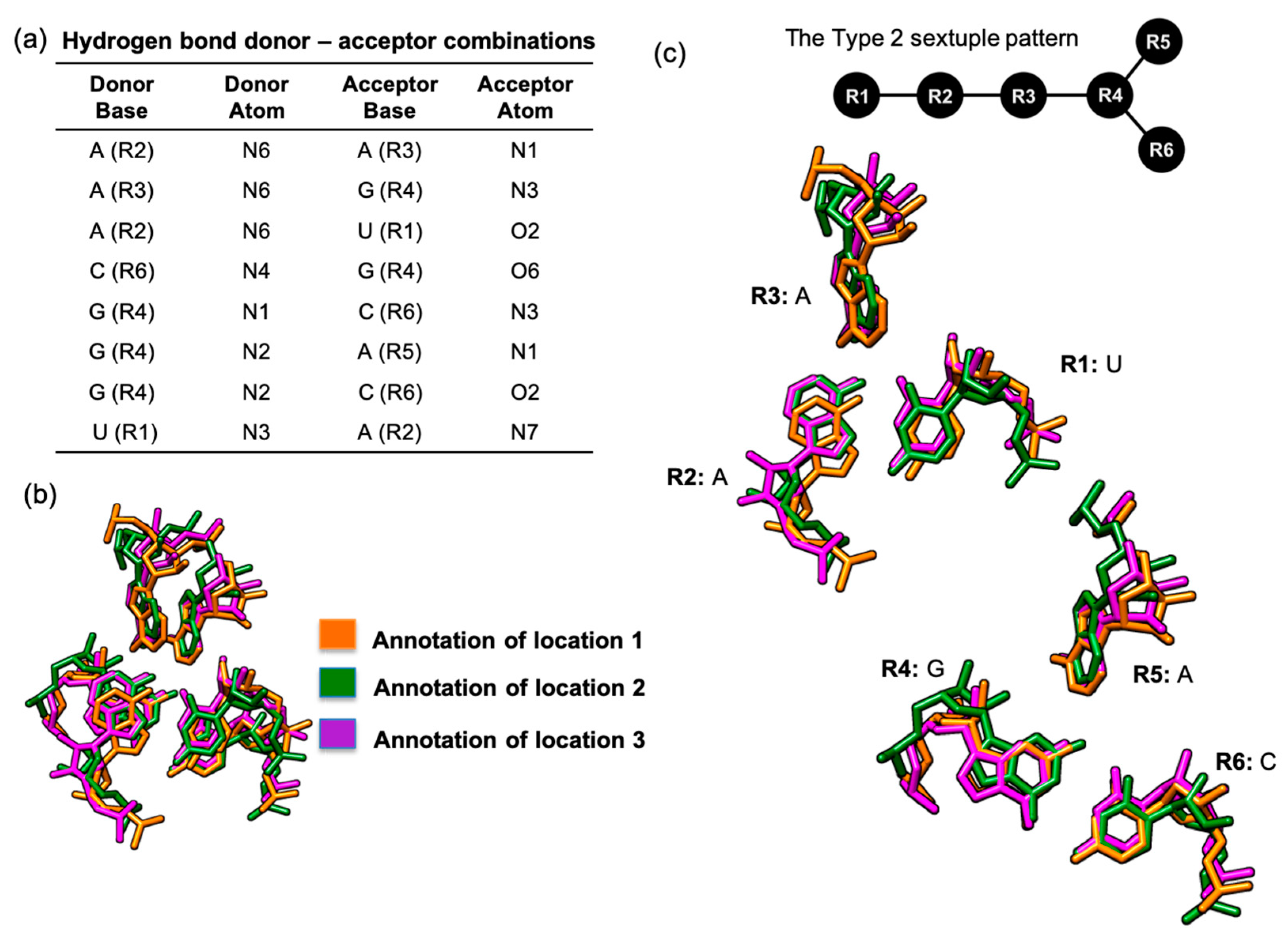
3.2. Hydrogen Bond-Connected Six-Base Interactions as Novel Structural Motifs
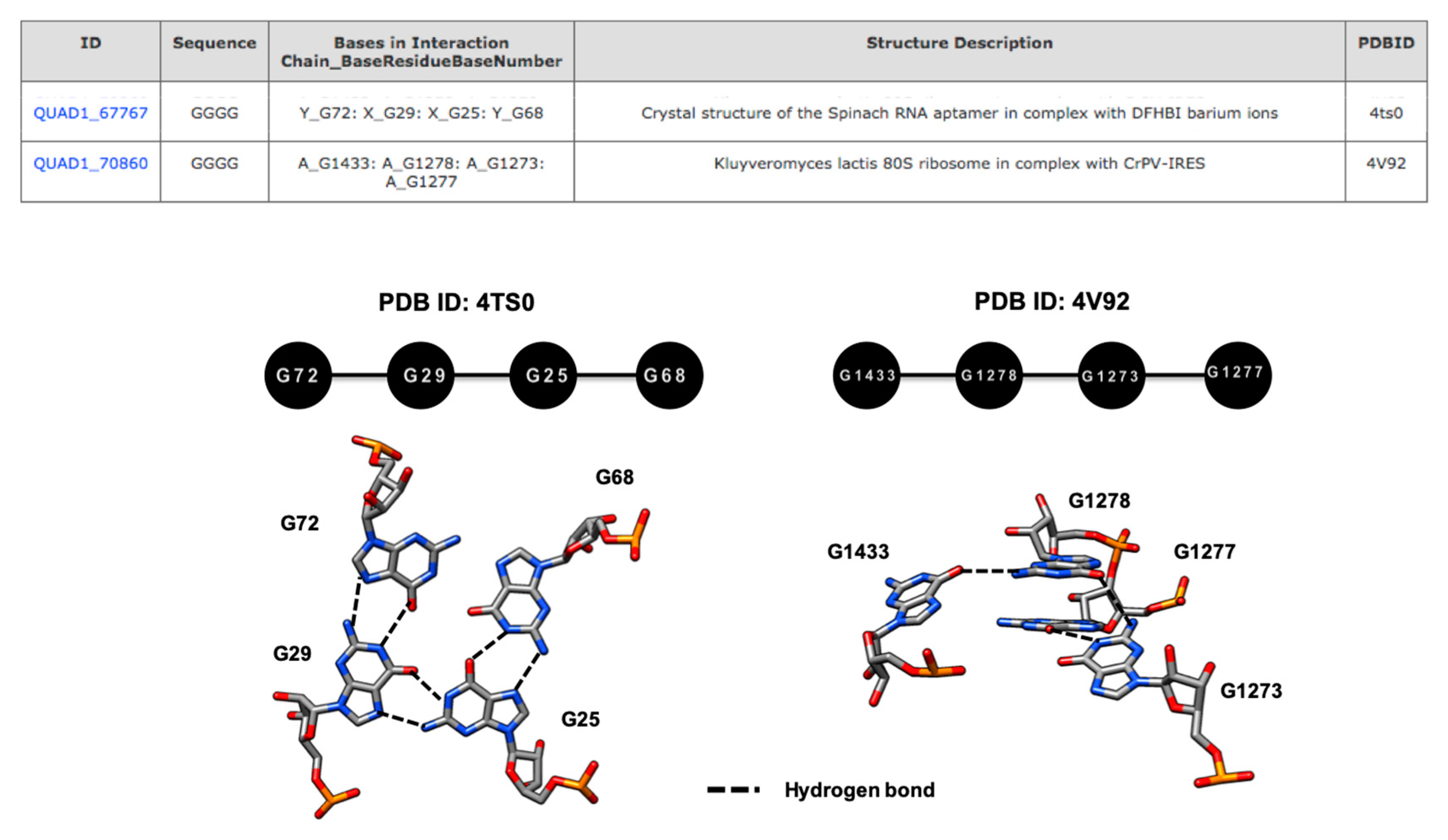
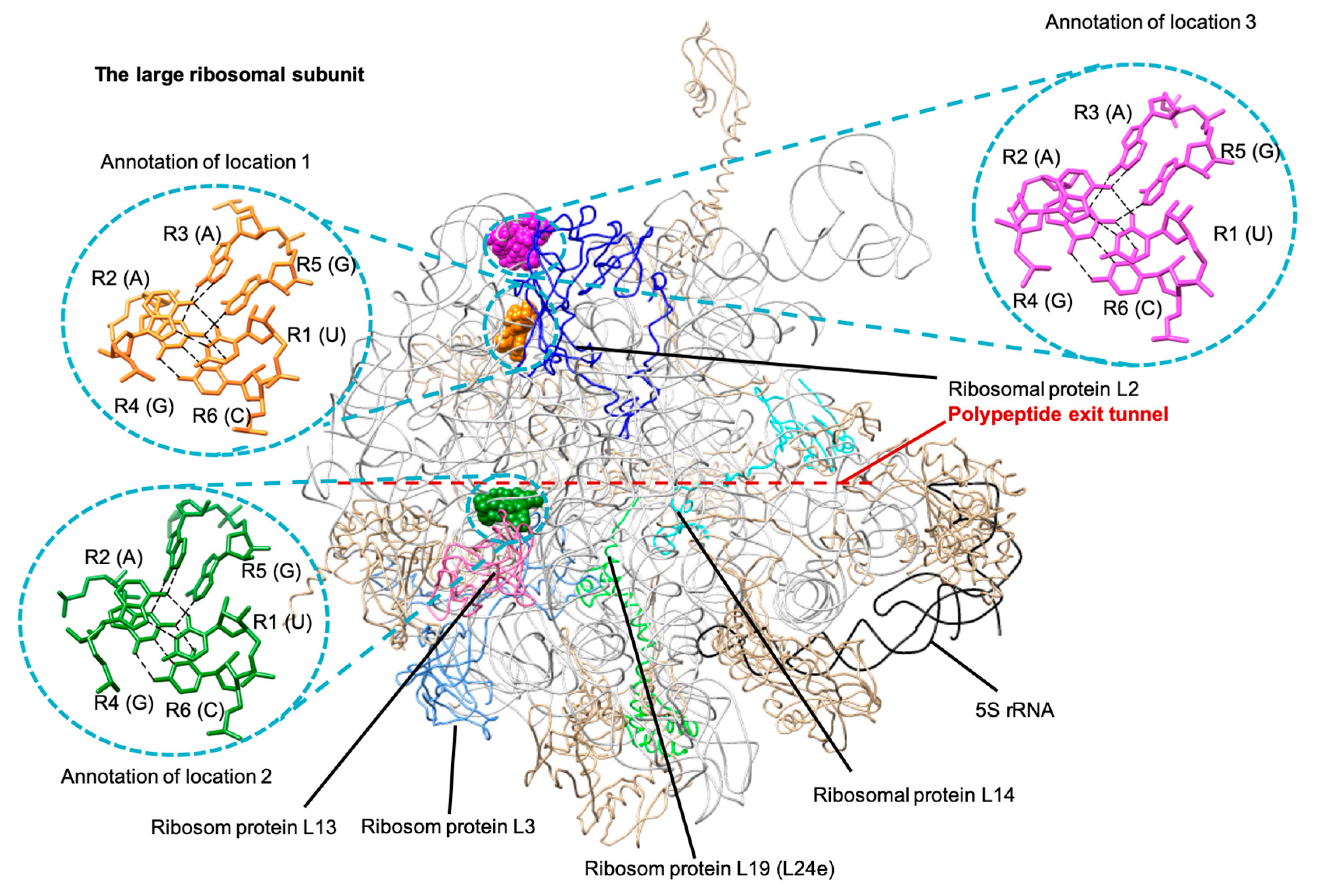
3.2.1. A Base Sextuple Annotated in Different RNA Structures
3.2.2. A Base Sextuple Annotated at Different Locations in the Same Structure
3.3. Presence of Known Motifs within the Sextuples
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Firdaus-Raih, M.; Hamdani, H.Y.; Nadzirin, N.; Ramlan, E.I.; Willett, P.; Artymiuk, P.J. COGNAC: A Web Server for Searching and Annotating Hydrogen-Bonded Base Interactions in RNA Three-Dimensional Structures. Nucleic Acids Res. 2014, 42, W382–W388. [Google Scholar] [CrossRef]
- Hamdani, H.Y.; Artymiuk, P.J.; Firdaus-Raih, M. A Computational Approach for the Annotation of Hydrogen-Bonded Base Interactions in Crystallographic Structures of the Ribozymes. AIP Conf. Proc. 2015, 1678. [Google Scholar] [CrossRef]
- Bhattacharya, S.; Jhunjhunwala, A.; Halder, A.; Bhattacharyya, D.; Mitra, A. Going beyond Base-Pairs: Topology-Based Characterization of Base-Multiplets in RNA. RNA 2019, 25, 573–589. [Google Scholar] [CrossRef]
- Nagaswamy, U. NCIR: A Database of Non-Canonical Interactions in Known RNA Structures. Nucleic Acids Res. 2002, 30, 395–397. [Google Scholar] [CrossRef]
- Firdaus-Raih, M.; Harrison, A.M.; Willett, P.; Artymiuk, P.J. Novel Base Triples in RNA Structures Revealed by Graph Theoretical Searching Methods. BMC Bioinform. 2011, 12 (Suppl. 13). [Google Scholar] [CrossRef]
- Appasamy, S.D.; Hamdani, H.Y.; Ramlan, E.I.; Firdaus-Raih, M. InterRNA: A Database of Base Interactions in RNA Structures. Nucleic Acids Res. 2016, 44, D266–D271. [Google Scholar] [CrossRef] [PubMed]
- Ren, H.; Shen, Y.; Zhang, L. The λ-Turn: A New Structural Motif in Ribosomal RNA. In Lecture Notes in Computer Science (Including Subseries Lecture Notes in Artificial Intelligence and Lecture Notes in Bioinformatics); Springer: Berlin/Heidelberg, Germany, 2015; Volume 9226, pp. 456–466. [Google Scholar] [CrossRef]
- Shen, Y.; Zhang, L. The Hasp Motif: A New Type of RNA Tertiary Interactions. In Lecture Notes in Computer Science (Including Subseries Lecture Notes in Artificial Intelligence and Lecture Notes in Bioinformatics); Springer: Berlin/Heidelberg, Germany, 2017; Volume 10362 LNCS, pp. 441–453. [Google Scholar] [CrossRef]
- Parlea, L.G.; Sweeney, B.A.; Hosseini-Asanjan, M.; Zirbel, C.L.; Leontis, N.B. The RNA 3D Motif Atlas: Computational Methods for Extraction, Organization and Evaluation of RNA Motifs. Methods 2016, 103, 99–119. [Google Scholar] [CrossRef] [PubMed]
- Zahran, M.; Sevim Bayrak, C.; Elmetwaly, S.; Schlick, T. RAG-3D: A Search Tool for RNA 3D Substructures. Nucleic Acids Res. 2015, 43, 9474–9488. [Google Scholar] [CrossRef] [PubMed]
- Piątkowski, P.; Jabłónska, J.; Zyła, A.; Niedziałek, D.; Matelska, D.; Jankowska, E.; Waleń, T.; Dawson, W.K.; Bujnicki, J.M. SupeRNAlign: A New Tool for Flexible Superposition of Homologous RNA Structures and Inference of Accurate Structure-Based Sequence Alignments. Nucleic Acids Res. 2017, 45, e150. [Google Scholar] [CrossRef]
- Hendrix, D.K.; Brenner, S.E.; Holbrook, S.R. RNA Structural Motifs: Building Blocks of a Modular Biomolecule. Q. Rev. Biophys. 2005, 38, 221–243. [Google Scholar] [CrossRef]
- Schroeder, K.T.; Mcphee, S.A.; Ouellet, J.; Lilley, D.M.J. A Structural Database for K-Turn Motifs in RNA. RNA 2010, 16, 1463–1468. [Google Scholar] [CrossRef] [PubMed]
- Lilley, D.M.J. The K-Turn Motif in Riboswitches and Other RNA Species. Biochim. Biophys. Acta Gene Regul. Mech. 2014, 1839, 995–1004. [Google Scholar] [CrossRef] [PubMed]
- Miskiewicz, J.; Szachniuk, M. Discovering Structural Motifs in MiRNA Precursors from the Viridiplantae Kingdom. Molecules 2018, 23, 1367. [Google Scholar] [CrossRef] [PubMed]
- Nissen, P.; Ippolito, J.A.; Ban, N.; Moore, P.B.; Steitz, T.A. RNA Tertiary Interactions in the Large Ribosomal Subunit: The A-Minor Motif. Proc. Natl. Acad. Sci. USA 2001, 98, 4899–4903. [Google Scholar] [CrossRef]
- Lai, C.E.; Tsai, M.Y.; Liu, Y.C.; Wang, C.W.; Chen, K.T.; Lu, C.L. FASTR3D: A Fast and Accurate Search Tool for Similar RNA 3D Structures. Nucleic Acids Res. 2009, 37 (Suppl. 2), W287–W295. [Google Scholar] [CrossRef]
- Hamdani, H.Y.; Appasamy, S.D.; Willett, P.; Artymiuk, P.J.; Firdaus-Raih, M. NASSAM: A Server to Search for and Annotate Tertiary Interactions and Motifs in Three-Dimensional Structures of Complex RNA Molecules. Nucleic Acids Res. 2012, 40, W35–W41. [Google Scholar] [CrossRef]
- Yen, C.Y.; Lin, J.C.; Chen, K.T.; Lu, C.L. R3D-BLAST2: An Improved Search Tool for Similar RNA 3D Substructures. BMC Bioinform. 2017, 18, 574. [Google Scholar] [CrossRef]
- Warner, K.D.; Chen, M.C.; Song, W.; Strack, R.L.; Thorn, A.; Jaffrey, S.R.; Ferré-D’Amaré, A.R. Structural Basis for Activity of Highly Efficient RNA Mimics of Green Fluorescent Protein. Nat. Struct. Mol. Biol. 2014, 21, 658–663. [Google Scholar] [CrossRef]
- Fernández, I.S.; Bai, X.C.; Murshudov, G.; Scheres, S.H.W.; Ramakrishnan, V. Initiation of Translation by Cricket Paralysis Virus IRES Requires Its Translocation in the Ribosome. Cell 2014, 157, 823–831. [Google Scholar] [CrossRef] [Green Version]
- Burley, S.K.; Berman, H.M.; Bhikadiya, C.; Bi, C.; Chen, L.; Di Costanzo, L.; Christie, C.; Dalenberg, K.; Duarte, J.M.; Dutta, S.; et al. RCSB Protein Data Bank: Biological Macromolecular Structures Enabling Research and Education in Fundamental Biology, Biomedicine, Biotechnology and Energy. Nucleic Acids Res. 2019, 47, D464–D474. [Google Scholar] [CrossRef]
- McDonald, I.K.; Thornton, J.M. Satisfying Hydrogen Bonding Potential in Proteins. J. Mol. Biol. 1994, 238, 777–793. [Google Scholar] [CrossRef] [PubMed]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera—A Visualization System for Exploratory Research and Analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef] [PubMed]
- Cocozaki, A.I.; Altman, R.B.; Huang, J.; Buurman, E.T.; Kazmirski, S.L.; Doig, P.; Prince, D.B.; Blanchard, S.C.; Cate, J.H.D.; Ferguson, A.D. Resistance Mutations Generate Divergent Antibiotic Susceptibility Profiles against Translation Inhibitors. Proc. Natl. Acad. Sci. USA 2016, 113, 8188–8193. [Google Scholar] [CrossRef] [PubMed]
- Schluenzen, F.; Tocilj, A.; Zarivach, R.; Harms, J.; Gluehmann, M.; Janell, D.; Bashan, A.; Bartels, H.; Agmon, I.; Franceschi, F.; et al. Structure of Functionally Activated Small Ribosomal Subunit at 3.3 Å Resolution. Cell 2000, 102, 615–623. [Google Scholar] [CrossRef]
- Ogle, J.M.; Brodersen, D.E.; Clemons, W.M., Jr.; Tarry, M.J.; Carter, A.P.; Ramakrishnan, V. Recognition of Cognate Transfer RNA by the 30S Ribosomal Subunit. Science 2001, 292, 897–902. [Google Scholar] [CrossRef]
- Dinman, J.D. 5S RRNA: Structure and Function from Head to Toe. Int. J. Biomed. Sci. 2005, 1, 2–7. [Google Scholar] [PubMed]
- Klein, D.J.; Edwards, T.E.; Ferré-D’Amaré, A.R. Cocrystal Structure of a Class I PreQ1 Riboswitch Reveals a Pseudoknot Recognizing an Essential Hypermodified Nucleobase. Nat. Struct. Mol. Biol. 2009, 16, 343–344. [Google Scholar] [CrossRef]
- Staple, D.W.; Butcher, S.E. Pseudoknots: RNA Structures with Diverse Functions. PLoS Biol. 2005, 3, e213. [Google Scholar] [CrossRef]
- Klein, D.J.; Schmeing, T.M.; Moore, P.B.; Steitz, T.A. The Kink-Turn: A New RNA Secondary Structure Motif. EMBO J. 2001, 20, 4214–4221. [Google Scholar] [CrossRef]
- Edwards, A.L.; Reyes, F.E.; Héroux, A.; Batey, R.T. Structural Basis for Recognition of S-Adenosylhomocysteine by Riboswitches. RNA 2010, 16, 2144–2155. [Google Scholar] [CrossRef]
- Aalberts, D.P.; Hodas, N.O. Asymmetry in RNA Pseudoknots: Observation and Theory. Nucleic Acids Res. 2005, 33, 2210–2214. [Google Scholar] [CrossRef] [PubMed]
- Maehigashi, T.; Dunkle, J.A.; Miles, S.J.; Dunham, C.M. Structural Insights into +1 Frameshifting Promoted by Expanded or Modification-Deficient Anticodon Stem Loops. Proc. Natl. Acad. Sci. USA 2014, 111, 12740–12745. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.; Wongsa, A.; Lee, J.; Shang, L.; Cannone, J.J.; Gutell, R.R. RNA2DMap: A Visual Exploration Tool of the Information in RNA’s Higher-Order Structure. In Proceedings of the 2011 IEEE International Conference on Bioinformatics and Biomedicine, BIBM 2011, Atlanta, GA, USA, 12–15 November 2011; pp. 613–617. [Google Scholar] [CrossRef]
- Borovinskaya, M.A.; Pai, R.D.; Zhang, W.; Schuwirth, B.S.; Holton, J.M.; Hirokawa, G.; Kaji, H.; Kaji, A.; Cate, J.H.D. Structural Basis for Aminoglycoside Inhibition of Bacterial Ribosome Recycling. Nat. Struct. Mol. Biol. 2007, 14, 727–732. [Google Scholar] [CrossRef] [PubMed]
- Maguire, B.A.; Zimmermann, R.A. The Ribosome in Focus. Cell 2001, 104, 813–816. [Google Scholar] [CrossRef]
- Jenner, L.; Starosta, A.L.; Terry, D.S.; Mikolajka, A.; Filonava, L.; Yusupov, M.; Blanchard, S.C.; Wilson, D.N.; Yusupova, G. Structural Basis for Potent Inhibitory Activity of the Antibiotic Tigecycline during Protein Synthesis. Proc. Natl. Acad. Sci. USA 2013, 110, 3812–3816. [Google Scholar] [CrossRef]
- Ban, N.; Nissen, P.; Hansen, J.; Moore, P.B.; Steitz, T.A. The Complete Atomic Structure of the Large Ribosomal Subunit at 2.4 Å Resolution. Science 2000, 289, 905–920. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pattern | Number of Annotations | Number of Unique Arrangements | Pattern | Number of Annotations | Number of Unique Arrangements |
|---|---|---|---|---|---|
 Type 1 | 10168 | 3572 |  Type 2 | 5448 | 2102 |
 Type 3 | 4240 | 1862 |  Type 4 | 525 | 299 |
 Type 5 | 8 | 6 |  Type 6 | 614 | 311 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hamdani, H.Y.; Firdaus-Raih, M. Identification of Structural Motifs Using Networks of Hydrogen-Bonded Base Interactions in RNA Crystallographic Structures. Crystals 2019, 9, 550. https://doi.org/10.3390/cryst9110550
Hamdani HY, Firdaus-Raih M. Identification of Structural Motifs Using Networks of Hydrogen-Bonded Base Interactions in RNA Crystallographic Structures. Crystals. 2019; 9(11):550. https://doi.org/10.3390/cryst9110550
Chicago/Turabian StyleHamdani, Hazrina Yusof, and Mohd Firdaus-Raih. 2019. "Identification of Structural Motifs Using Networks of Hydrogen-Bonded Base Interactions in RNA Crystallographic Structures" Crystals 9, no. 11: 550. https://doi.org/10.3390/cryst9110550
APA StyleHamdani, H. Y., & Firdaus-Raih, M. (2019). Identification of Structural Motifs Using Networks of Hydrogen-Bonded Base Interactions in RNA Crystallographic Structures. Crystals, 9(11), 550. https://doi.org/10.3390/cryst9110550