
Biomedical Applications of Bacteria-Derived Polymers


 and
and
Abstract
:
1. Introduction
2. The History, Contemporary Status, and Future Applications of Bacteria-Derived Polymers
2.1. Polysaccharides
2.1.1. Dextran
Dextran Synthesis and Properties
Dextran as a Potential Biomaterial
2.1.2. Glycogen
Glycogen Properties and Current Research
Concepts, Advantages and Limitations of Glycogen Production by Bacterial Fermentation
2.1.3. Alginate
Structure, Biosynthesis, and Modifications
Potential Applications of Alginate in Biomedicine
2.1.4. Hyaluronic Acid
Properties, Current, and Future Clinical Usage
Past, Current, and Future Manufacturing of Hyaluronic Acid
2.1.5. Gellan
Structure, Composition, and Classification of Gellan Gum
Biomedical Applications of Gellan
2.1.6. Xanthan
Biosynthesis and Industrial Production
Biomedical Properties of Xanthan
2.1.7. Curdlan
Structure and Properties of Curdlan
Biopharmaceutical Applications of Curdlan
2.2. Polyesters
2.2.1. Polyhydroxyalkanoates
PHA as a Biomaterial
2.2.2. Polylactic Acid
Polylactic Variant and Attributes
Production of Polylactic Acid
Polylactic Acid in Biomedical Application
2.3. Polyamide
2.3.1. ε-Poly-l-Lysine
Current Properties and Subsequent Applications
Production of the Polymer by Bacteria
2.3.2. Poly-γ-Glutamate (γ-PGA)
Properties of γ-PGA
Manufacture of Poly-γ-Glutamate
2.4. Polyanhydride
2.4.1. Polyphosphate
Microbial Production of Polyphosphate
Polyphosphate Role in Physiological Processes
3. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- PlasticsEurope. Plastics—The Facts 2019: An Analysis of European Plastics Production, Demand and Waste Data. Belgium. 2019. Available online: https://www.plasticseurope.org/application/files/9715/7129/9584/FINAL_web_version_Plastics_the_facts2019_14102019.pdf (accessed on 15 October 2020).
- Meikle, J.L. American Plastic: A Cultural History; Rutgers University Press: New Brunswick, NJ, USA, 1995. [Google Scholar]
- Potter, K.D. The early history of the resin transfer moulding process for aerospace applications. Compos. Part A Appl. Sci. Manuf. 1999, 30, 619–621. [Google Scholar] [CrossRef]
- Hench, L.L. Biomaterials: A forecast for the future. Biomaterials 1998, 19, 1419–1423. [Google Scholar] [CrossRef]
- Rea, S.; Bonfield, W. Biocomposites for medical applications. J. Australas. Ceram. Soc. 2004, 40, 43–57. [Google Scholar]
- Kane, S.R.; Ashby, P.D.; Pruitt, L.A. Characterization and tribology of PEG-like coatings on UHMWPE for total hip replacements. J. Biomed. Mater. Res. Part A Off. J. Soc. Biomater. Jpn. Soc. Biomater. Aust. Soc. Biomater. Korean Soc. Biomater. 2010, 92, 1500–1509. [Google Scholar] [CrossRef] [PubMed]
- Xing, C.-M.; Meng, F.-N.; Quan, M.; Ding, K.; Dang, Y.; Gong, Y.-K. Quantitative fabrication, performance optimization and comparison of PEG and zwitterionic polymer antifouling coatings. Acta Biomater. 2017, 59, 129–138. [Google Scholar] [CrossRef]
- Prokop, A.; Hunkeler, D.; Powers, A.; Whitesell, R.; Wang, T. Water soluble polymers for immunoisolation II: Evaluation of multicomponent microencapsulation systems. In Microencapsulation Microgels Iniferters; Springer: Berlin/Heidelberg, Germany, 1998; pp. 53–73. [Google Scholar]
- Gupta, M.K.; Walthall, J.M.; Venkataraman, R.; Crowder, S.W.; Jung, D.K.; Shann, S.Y.; Feaster, T.K.; Wang, X.; Giorgio, T.D.; Hong, C.C. Combinatorial polymer electrospun matrices promote physiologically-relevant cardiomyogenic stem cell differentiation. PLoS ONE 2011, 6, e28935. [Google Scholar] [CrossRef] [Green Version]
- Jaidev, L.; Chatterjee, K. Surface functionalization of 3D printed polymer scaffolds to augment stem cell response. Mater. Des. 2019, 161, 44–54. [Google Scholar] [CrossRef]
- Shim, W.J.; Thomposon, R.C. Microplastics in the ocean. Arch. Environ. Contam. Toxicol. 2015, 69, 265–268. [Google Scholar] [CrossRef]
- Astrup, T.; Fruergaard, T.; Christensen, T.H. Recycling of plastic: Accounting of greenhouse gases and global warming contributions. Waste Manag. Res. 2009, 27, 763–772. [Google Scholar] [CrossRef]
- Pirc, U.; Vidmar, M.; Mozer, A.; Kržan, A. Emissions of microplastic fibers from microfiber fleece during domestic washing. Environ. Sci. Pollut. Res. 2016, 23, 22206–22211. [Google Scholar] [CrossRef] [Green Version]
- Valavanidis, A.; Iliopoulos, N.; Gotsis, G.; Fiotakis, K. Persistent free radicals, heavy metals and PAHs generated in particulate soot emissions and residue ash from controlled combustion of common types of plastic. J. Hazard. Mater. 2008, 156, 277–284. [Google Scholar] [CrossRef]
- Rehm, B.H. Bacterial polymers: Biosynthesis, modifications and applications. Nat. Rev. Microbiol. 2010, 8, 578–592. [Google Scholar] [CrossRef]
- Inbaraj, B.S.; Chiu, C.; Ho, G.; Yang, J.; Chen, B. Effects of temperature and pH on adsorption of basic brown 1 by the bacterial biopolymer poly (γ-glutamic acid). Bioresour. Technol. 2008, 99, 1026–1035. [Google Scholar] [CrossRef]
- Lenz, R.W.; Marchessault, R.H. Bacterial polyesters: Biosynthesis, biodegradable plastics and biotechnology. Biomacromolecules 2005, 6, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Li, S.Y.; Dong, C.L.; Wang, S.Y.; Ye, H.M.; Chen, G.-Q. Microbial production of polyhydroxyalkanoate block copolymer by recombinant Pseudomonas putida. Appl. Microbiol. Biotech. 2011, 90, 659–669. [Google Scholar] [CrossRef] [PubMed]
- Reichmann, N.T.; Cassona, C.P.; Gründling, A. Revised mechanism of D-alanine incorporation into cell wall polymers in Gram-positive bacteria. Microbiology 2013, 159, 1868. [Google Scholar] [CrossRef]
- Chen, G.G.-Q. Plastics from Bacteria: Natural Functions and Applications; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2009; Volume 14. [Google Scholar]
- Lopes, M.S.; Jardini, A.; Maciel-Filho, R. Poly (lactic acid) production for tissue engineering applications. Proc. Eng. 2012, 42, 1402–1413. [Google Scholar] [CrossRef] [Green Version]
- Tachibana, Y.; Yamahata, M.; Kimura, S.; Kasuya, K.-I. Synthesis, Physical Properties, and Biodegradability of Biobased Poly (butylene succinate-co-butylene oxabicyclate). ACS Sustain. Chem. Eng. 2018, 6, 10806–10814. [Google Scholar] [CrossRef]
- Tarrahi, R.; Fathi, Z.; Seydibeyoğlu, M.Ö.; Doustkhah, E.; Khataee, A. Polyhydroxyalkanoates (PHA): From production to nanoarchitecture. Int. J. Biol. Macromol. 2020, 146, 596–619. [Google Scholar] [CrossRef]
- Morris, G.; Harding, S. Polysaccharides, microbial. In Encyclopedia of Microbiology; Elsevier Inc.: Amsterdam, The Netherlands, 2009; pp. 482–494. [Google Scholar]
- Oliveira, J.T.; Reis, R.L. 18—Hydrogels from Polysaccharide-Based Materials: Fundamentals and Applications in Regenerative Medicine; Reis, R.L., Neves, N.M., Mano, J.F., Gomes, M.E., Marques, A.P., Azevedo, H.S., Eds.; Woodhead Publishing: Cambridge, UK, 2008; pp. 485–514. [Google Scholar] [CrossRef]
- Sajna, K.V.; Gottumukkala, L.D.; Sukumaran, R.K.; Pandey, A. Chapter 18—White Biotechnology in Cosmetics. In Industrial Biorefineries & White Biotechnology; Pandey, A., Höfer, R., Taherzadeh, M., Nampoothiri, K.M., Larroche, C., Eds.; Elsevier: Amsterdam, The Netherlands, 2015; pp. 607–652. [Google Scholar] [CrossRef]
- Kothari, D.; Das, D.; Patel, S.; Goyal, A. Dextran and food application. In Polysacchccharides; Springer: Berlin, Germany, 2014; pp. 735–752. [Google Scholar]
- Patel, A.; Prajapat, J. Food and health applications of exopolysaccharides produced by lactic acid bacteria. Adv. Dairy Res. 2013, 1–8. [Google Scholar] [CrossRef]
- Selvi, S.S.; Eminagic, E.; Kandur, M.Y.; Ozcan, E.; Kasavi, C.; Oner, E.T. Research and Production of Microbial Polymers for Food Industry. Bioproces. Biomol. Product. 2019, 211–238. [Google Scholar] [CrossRef]
- Pasteur, L. On the viscous fermentation and the butyrous fermentation. Bull. Soc. Chim. 1861, 11, 30–31. [Google Scholar]
- Crescenzi, V. Microbial Polysaccharides of Applied Interest—Ongoing Research Activities in Europe. Biotechnol. Progr. 1995, 11, 251–259. [Google Scholar] [CrossRef]
- Naessens, M.; Cerdobbel, A.; Soetaert, W.; Vandamme, E.J. Leuconostoc dextransucrase and dextran: Production, properties and applications. J. Chem. Technol. Biotech. 2005, 80, 845–860. [Google Scholar] [CrossRef]
- Zhou, Q.; Feng, F.; Yang, Y.; Zhao, F.; Du, R.; Zhou, Z.; Han, Y. Characterization of a dextran produced by Leuconostoc pseudomesenteroides XG5 from homemade wine. Int. J. Biol. Macromol. 2018, 107, 2234–2241. [Google Scholar] [CrossRef] [PubMed]
- Aman, A.; Siddiqui, N.N.; Ul-Qader, S.A. Characterization and potential applications of high molecular weight dextran produced by Leuconostoc mesenteroides AA1. Carbohydr. Polym. 2012, 87, 910–915. [Google Scholar] [CrossRef]
- Feng, F.; Zhou, Q.Q.; Yang, Y.F.; Zhao, F.K.; Du, R.P.; Han, Y.; Xiao, H.Z.; Zhou, Z.J. Characterization of highly branched dextran produced by Leuconostoc citreum B-2 from pineapple fermented product. Int. J. Biol. Macromol. 2018, 113, 45–50. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, A.; Bandopadhyay, R. Use of dextran nanoparticle: A paradigm shift in bacterial exopolysaccharide based biomedical applications. Int. J. Biol. Macromol. 2016, 87, 295–301. [Google Scholar] [CrossRef]
- Maia, J.; Evangelista, M.; Gil, H.; Ferreira, L. Dextran-based materials for biomedical applications. In Carbohydrates Applications in Medicine; Gil, M.H., Ed.; Research Signpost: Kerala, India, 2014; pp. 31–53. [Google Scholar]
- Purama, R.K.; Arun, G. Dextransucrase production by Leuconostoc mesenteroides. Indian J. Microbiol. 2005, 45, 89–101. [Google Scholar]
- Patil, S.B.; Inamdar, S.Z.; Reddy, K.R.; Raghu, A.V.; Soni, S.K.; Kulkarni, R.V. Novel biocompatible poly(acrylamide)-grafted-dextran hydrogels: Synthesis, characterization and biomedical applications. J. Microbiol. Meth. 2019, 159, 200–209. [Google Scholar] [CrossRef] [PubMed]
- Khalikova, E.; Susi, P.; Korpela, T. Microbial dextran-hydrolyzing enzymes: Fundamentals and applications. Microbiol. Mol. Biol. R 2005, 69, 306–325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patel, S.; Majumder, A.; Goyal, A. Potentials of Exopolysaccharides from Lactic Acid Bacteria. Indian J. Microbiol. 2012, 52, 3–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baruah, R.; Maina, N.H.; Katina, K.; Juvonen, R.; Goyal, A. Functional food applications of dextran from Weissella cibaria RBA12 from pummelo (Citrus maxima). Int. J. Food Microbiol. 2017, 242, 124–131. [Google Scholar] [CrossRef] [PubMed]
- Besrour-Aouam, N.; Fhoula, I.; Hernández-Alcántara, A.M.; Mohedano, M.L.; Najjari, A.; Prieto, A.; Ruas-Madiedo, P.; López, P.; Ouzari, H.-I. The role of dextran production in the metabolic context of Leuconostoc and Weissella Tunisian strains. Carbohydr. Polym. 2021, 253, 117254. [Google Scholar] [CrossRef]
- Shukla, R.; Goyal, A. Novel dextran from Pediococcus pentosaceus CRAG3 isolated from fermented cucumber with anti-cancer properties. Int. J. Biol. Macromol. 2013, 62, 352–357. [Google Scholar] [CrossRef]
- Wang, B.; Song, Q.; Zhao, F.; Zhang, L.; Han, Y.; Zhou, Z. Isolation and characterization of dextran produced by Lactobacillus sakei L3 from Hubei sausage. Carbohydr. Polym. 2019, 223, 115111. [Google Scholar] [CrossRef]
- Freitas, F.; Torres, C.A.V.; Reis, M.A.M. Engineering aspects of microbial exopolysaccharide production. Bioresour. Technol. 2017, 245, 1674–1683. [Google Scholar] [CrossRef]
- Gibbons, R.J.; Banghart, S.B. Synthesis of extracellular dextran by cariogenic bacteria and its presence in human dental plaque. Arch. Oral Biol. 1967, 12, 11–24. [Google Scholar] [CrossRef]
- Leathers, T.D. Dextran. Biopolymers 2002, 5, 299–321. [Google Scholar]
- Rahmat-Zohra, R.; Waseem, S.; Aman, A.; Siddiqui, A.; Kahkashan-Kazmi, S.; Rahmat-Zohra, R. Dextran Production by Microbial Biotransformation of Sugarcane Waste. FUUAST J. Biol. 2009, 9, 87–94. [Google Scholar]
- Pescosolido, L.; Vermonden, T.; Malda, J.; Censi, R.; Dhert, W.J.A.; Alhaique, F.; Hennink, W.E.; Matricardi, P. In situ forming IPN hydrogels of calcium alginate and dextran-HEMA for biomedical applications. Acta Biomater. 2011, 7, 1627–1633. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pacelli, S.; di Muzio, L.; Paolicelli, P.; Fortunati, V.; Petralito, S.; Trilli, J.; Casadei, M.A. Dextran-polyethylene glycol cryogels as spongy scaffolds for drug delivery. Int. J. Biol. Macromol. 2020. [Google Scholar] [CrossRef] [PubMed]
- Redasani, V.K.; Bari, S.B. Chapter—Approaches for Prodrugs. In Prodrug Design; Redasani, V.K., Bari, S.B., Eds.; Academic Press: Boston, MA, USA, 2015; pp. 33–49. [Google Scholar] [CrossRef]
- Praveen, B.; Shrivastava, P.; Shrivastava, S. In-Vitro release and pharmacological study of synthesized valproic acid-dextran conjugate. Acta Pharm. Sci. 2009, 51, 169–176. [Google Scholar]
- Cai, L.T.; Li, J.T.; Quan, S.T.; Feng, W.; Yao, J.N.; Yang, M.L.; Li, W.Y. Dextran-based hydrogel with enhanced mechanical performance via covalent and non-covalent cross-linking units carrying adipose-derived stem cells toward vascularized bone tissue engineering. J. Biomed. Mater. Res. Part A 2019, 107, 1120–1131. [Google Scholar] [CrossRef] [PubMed]
- Jain, V.; Shukla, N.; Mahajan, S. Polysaccharides in colon specific drug delivery. J. Transl. Sci. 2015, 1, 3–11. [Google Scholar] [CrossRef]
- Hovgaard, L.; Brondsted, H. Dextran Hydrogels for Colon-Specific Drug-Delivery. J. Control. Release 1995, 36, 159–166. [Google Scholar] [CrossRef]
- Chalasani, K.B.; Russell-Jones, G.J.; Jain, A.K.; Diwan, P.V.; Jain, S.K. Effective oral delivery of insulin in animal models using vitamin B12-coated dextran nanoparticles. J. Control. Release 2007, 122, 141–150. [Google Scholar] [CrossRef]
- Foerster, F.; Bamberger, D.; Schupp, J.; Weilbacher, M.; Kaps, L.; Strobl, S.; Radi, L.; Diken, M.; Strand, D.; Tuettenberg, A.; et al. Dextran-based therapeutic nanoparticles for hepatic drug delivery. Nanomedicine 2016, 11, 2663–2677. [Google Scholar] [CrossRef]
- Froemel, D.; Fitzsimons, S.J.; Frank, J.; Sauerbier, M.; Meurer, A.; Barker, J.H. A Review of Thrombosis and Antithrombotic Therapy in Microvascular Surgery. Eur. Surg. Res. 2013, 50, 32–43. [Google Scholar] [CrossRef]
- Manners, D.J. Recent developments in our understanding of glycogen structure. Carbohydr. Polym. 1991, 16, 37–82. [Google Scholar] [CrossRef]
- El Khadem, H.S. Carbohydrates. In Encyclopedia of Physical Science and Technology; Meyers, R.A., Ed.; Academic Press: London, UK, 2002. [Google Scholar]
- Mischnick, P.; Momcilovic, D. Chemical structure analysis of starch and cellulose derivatives. In Advances in Carbohydrate Chemistry and Biochemistry; Horton, D., Ed.; Elsevier Science: Amsterdam, The Netherlands, 2010; Volume 64. [Google Scholar]
- Brown, A.M.; Tekkök, S.B.; Ransom, B.R. Glycogen regulation and functional role in mouse white matter. J. Physiol. 2003, 549, 501–512. [Google Scholar] [CrossRef] [PubMed]
- Hers, H.G. Mechanisms of blood glucose homeostasis. J. Inherit. Metab. Dis. 1990, 13, 395–410. [Google Scholar] [CrossRef]
- Fricain, J.; Granja, P.; Barbosa, M.; de Jéso, B.; Barthe, N.; Baquey, C. Cellulose phosphates as biomaterials. In vivo biocompatibility studies. Biomaterials 2002, 23, 971–980. [Google Scholar] [CrossRef]
- Klemm, D.; Schumann, D.; Udhardt, U.; Marsch, S. Bacterial synthesized cellulose—Artificial blood vessels for microsurgery. Prog. Polym. Sci. 2001, 26, 1561–1603. [Google Scholar] [CrossRef]
- Torres, F.G.; Commeaux, S.; Troncoso, O.P. Biocompatibility of bacterial cellulose based biomaterials. J. Funct. Biomater. 2012, 3, 864–878. [Google Scholar] [CrossRef] [Green Version]
- Cifuente, J.O.; Comino, N.; D’Angelo, C.; Marina, A.; Gil-Carton, D.; Albesa-Jové, D.; Guerin, M.E. The allosteric control mechanism of bacterial glycogen biosynthesis disclosed by cryoEM. Curr. Res. Struct. Biol. 2020, 2, 89–103. [Google Scholar] [CrossRef]
- Ball, S.; Colleoni, C.; Cenci, U.; Raj, J.N.; Tirtiaux, C. The evolution of glycogen and starch metabolism in eukaryotes gives molecular clues to understand the establishment of plastid endosymbiosis. J. Exp. Bot. 2011, 62, 1775–1801. [Google Scholar] [CrossRef] [Green Version]
- Adeva-Andany, M.M.; González-Lucán, M.; Donapetry-García, C.; Fernández-Fernández, C.; Ameneiros-Rodríguez, E. Glycogen metabolism in humans. BBA Clin. 2016, 5, 85–100. [Google Scholar] [CrossRef] [Green Version]
- Engelking, L.R. Textbook of Veterinary Physiological Chemistry, Updated 2/e; Academic Press: Cambridge, MA, USA, 2010. [Google Scholar]
- Mor, I.; Cheung, E.; Vousden, K. Control of glycolysis through regulation of PFK1: Old friends and recent additions. Proc. Cold Spring Harb. Symp. Quant. Biol. 2011, 76, 211–216. [Google Scholar] [CrossRef]
- Yamashita, T.; Ishibashi, Y.; Nagaoka, I.; Kasuya, K.; Masuda, K.; Warabi, H.; Shiokawa, Y. Studies on glycogen-induced inflammation of mice. Inflammation 1982, 6, 87–101. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.X.; Jones, D.V.; Clemens, M.G. Effect of activation on neutrophil-induced hepatic microvascular injury in isolated rat liver. Shock 1994, 1, 273–278. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, E.M. Hydrogel: Preparation, characterization, and applications: A review. J. Adv. Res. 2015, 6, 105–121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Billiet, T.; Vandenhaute, M.; Schelfhout, J.; van Vlierberghe, S.; Dubruel, P. A review of trends and limitations in hydrogel-rapid prototyping for tissue engineering. Biomaterials 2012, 33, 6020–6041. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Marchant, R.E. Design properties of hydrogel tissue-engineering scaffolds. Expert Rev. Med. Devices 2011, 8, 607–626. [Google Scholar] [CrossRef]
- Patra, P.; Rameshbabu, A.P.; Das, D.; Dhara, S.; Panda, A.B.; Pal, S. Stimuli-responsive, biocompatible hydrogel derived from glycogen and poly (N-isopropylacrylamide) for colon targeted delivery of ornidazole and 5-amino salicylic acid. Polym. Chem. 2016, 7, 5426–5435. [Google Scholar] [CrossRef]
- Patra, P.; Patra, N.; Pal, S. Opposite swelling characteristics through changing the connectivity in a biopolymeric hydrogel based on glycogen and glycine. Polym. Chem. 2020, 11, 2630–2634. [Google Scholar] [CrossRef]
- Evans, N.D.; Minelli, C.; Gentleman, E.; la Pointe, V.; Patankar, S.N.; Kallivretaki, M.; Chen, X.; Roberts, C.J.; Stevens, M.M. Substrate stiffness affects early differentiation events in embryonic stem cells. Eur. Cell Mater. 2009, 18, e13. [Google Scholar] [CrossRef]
- Park, J.S.; Chu, J.S.; Tsou, A.D.; Diop, R.; Tang, Z.; Wang, A.; Li, S. The effect of matrix stiffness on the differentiation of mesenchymal stem cells in response to TGF-β. Biomaterials 2011, 32, 3921–3930. [Google Scholar] [CrossRef] [Green Version]
- Subramony, S.D.; Dargis, B.R.; Castillo, M.; Azeloglu, E.U.; Tracey, M.S.; Su, A.; Lu, H.H. The guidance of stem cell differentiation by substrate alignment and mechanical stimulation. Biomaterials 2013, 34, 1942–1953. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Zhou, J.; Ying, H.; Zhou, Y.; Lai, J.; Chen, J. Glycogen as a Cross-Linking Agent of Collagen and Nanohydroxyapatite To Form Hydrogels for bMSC Differentiation. ACS Sustain. Chem. Eng. 2020, 8, 2106–2114. [Google Scholar] [CrossRef]
- Schlegel, P.N.; Group, H.S. Efficacy and safety of histrelin subdermal implant in patients with advanced prostate cancer. J. Urol. 2006, 175, 1353–1358. [Google Scholar] [CrossRef]
- Soon-Shiong, P.; Heintz, R.E.; Merideth, N.; Yao, Q.X.; Yao, Z.; Zheng, T.; Murphy, M.; Moloney, M.K.; Schmehl, M.; Harris, M. Insulin independence in a type 1 diabetic patient after encapsulated islet transplantation. Lancet 1994, 343, 950. [Google Scholar] [CrossRef]
- Zhang, L.; Chen, J.; Han, C. A multicenter clinical trial of recombinant human GM-CSF hydrogel for the treatment of deep second-degree burns. Wound Repair Regen. 2009, 17, 685–689. [Google Scholar] [CrossRef]
- Han, Y.; Hu, B.; Wang, M.; Yang, Y.; Zhang, L.; Zhou, J.; Chen, J. pH-Sensitive tumor-targeted hyperbranched system based on glycogen nanoparticles for liver cancer therapy. Appl. Mater. Today 2020, 18, 100521. [Google Scholar] [CrossRef]
- Hussain, I.; Sayed, S.M.; Liu, S.; Yao, F.; Oderinde, O.; Fu, G. Hydroxyethyl cellulose-based self-healing hydrogels with enhanced mechanical properties via metal-ligand bond interactions. Eur. Polym. J. 2018, 100, 219–227. [Google Scholar] [CrossRef]
- Hua, J.; Ng, P.F.; Fei, B. High-strength hydrogels: Microstructure design, characterization and applications. J. Polym. Sci. Part B Polym. Phys. 2018, 56, 1325–1335. [Google Scholar] [CrossRef] [Green Version]
- Pourjavadi, A.; Tavakoli, E.; Motamedi, A.; Salimi, H. Facile synthesis of extremely biocompatible double-network hydrogels based on chitosan and poly (vinyl alcohol) with enhanced mechanical properties. J. Appl. Polym. Sci. 2018, 135, 45752. [Google Scholar] [CrossRef]
- Shin, H.; Olsen, B.D.; Khademhosseini, A. The mechanical properties and cytotoxicity of cell-laden double-network hydrogels based on photocrosslinkable gelatin and gellan gum biomacromolecules. Biomaterials 2012, 33, 3143–3152. [Google Scholar] [CrossRef] [Green Version]
- Caballero, A.; Sulejmani, F.; Martin, C.; Pham, T.; Sun, W. Evaluation of transcatheter heart valve biomaterials: Biomechanical characterization of bovine and porcine pericardium. J. Mech. Behav. Biomed. Mater. 2017, 75, 486–494. [Google Scholar] [CrossRef]
- Diba, M.; Spaans, S.; Ning, K.; Ippel, B.D.; Yang, F.; Loomans, B.; Dankers, P.Y.; Leeuwenburgh, S.C. Self-healing biomaterials: From molecular concepts to clinical applications. Adv. Mater. Interfaces 2018, 5, 1800118. [Google Scholar] [CrossRef]
- Tellado, S.F.; Balmayor, E.R.; van Griensven, M. Strategies to engineer tendon/ligament-to-bone interface: Biomaterials, cells and growth factors. Adv. Drug Deliv. Rev. 2015, 94, 126–140. [Google Scholar] [CrossRef]
- Antoine, A.; Tepper, B. Environmental control of glycogen and lipid content of Mycobacterium phlei. Microbiology 1969, 55, 217–226. [Google Scholar] [CrossRef] [Green Version]
- Welles, L.; Lopez-Vazquez, C.; Hooijmans, C.; van Loosdrecht, M.; Brdjanovic, D. Impact of salinity on the anaerobic metabolism of phosphate-accumulating organisms (PAO) and glycogen-accumulating organisms (GAO). Appl. Microbiol. Biotech. 2014, 98, 7609–7622. [Google Scholar] [CrossRef]
- Zhang, C.; Chen, Y.; Liu, Y. The long-term effect of initial pH control on the enrichment culture of phosphorus-and glycogen-accumulating organisms with a mixture of propionic and acetic acids as carbon sources. Chemosphere 2007, 69, 1713–1721. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Wang, X.; Li, X.; Jia, S.; Wang, Q.; Peng, Y. Improvement of partial nitrification endogenous denitrification and phosphorus removal system: Balancing competition between phosphorus and glycogen accumulating organisms to enhance nitrogen removal without initiating phosphorus removal deterioration. Bioresour. Technol. 2019, 281, 382–391. [Google Scholar] [CrossRef] [PubMed]
- Preiss, J. Bacterial glycogen synthesis and its regulation. Annu. Rev. Microbiol. 1984, 38, 419–458. [Google Scholar] [CrossRef] [PubMed]
- Ali, A.A.; Shaban, K.A.; Tantawy, E.A. Effect of poly-β-hydroxybutyrate (PHB) and glycogen producing endophytic bacteria on yield, growth and nutrient. Appl. Sci. Rep. 2014, 8, 134–142. [Google Scholar]
- Birkhed, D.; Tanzer, J. Glycogen synthesis pathway in Streptococcus mutans strain NCTC 10449S and its glycogen synthesis-defective mutant 805. Arch. Oral Biol. 1979, 24, 67–73. [Google Scholar] [CrossRef]
- Braβnta, A.F.; Eandez, C.M.; Dáiaaz, L.A.; Manzanal, M.B.; Hardisson, C. Glycogen and trehalose accumulation during colony development in Streptomyces antibioticus. Microbiology 1986, 132, 1319–1326. [Google Scholar]
- Eydallin, G.; Montero, M.; Almagro, G.; Sesma, M.T.; Viale, A.M.; Munoz, F.J.; Rahimpour, M.; Baroja-Fernández, E.; Pozueta-Romero, J. Genome-wide screening of genes whose enhanced expression affects glycogen accumulation in Escherichia coli. DNA Res. 2010, 17, 61–71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, J.-P.; Ladapo, J.; Whitman, W.B. Pathway of glycogen metabolism in Methanococcus maripaludis. J. Bacteriol. 1994, 176, 325–332. [Google Scholar] [CrossRef] [Green Version]
- Zevenhuizen, L. Cellular glycogen, β-1, 2-glucan, poly-β-hydroxybutyric acid and extracellular polysaccharides in fast-growing species of Rhizobium. Antonie Van Leeuwenhoek 1981, 47, 481–497. [Google Scholar] [CrossRef] [PubMed]
- He, S.; McMahon, K.D. Microbiology of ‘Candidatus Accumulibacter’in activated sludge. Microb. Biotechnol. 2011, 4, 603–619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hickman, J.W.; Kotovic, K.M.; Miller, C.; Warrener, P.; Kaiser, B.; Jurista, T.; Budde, M.; Cross, F.; Roberts, J.M.; Carleton, M. Glycogen synthesis is a required component of the nitrogen stress response in Synechococcus elongatus PCC 7942. Algal Res. 2013, 2, 98–106. [Google Scholar] [CrossRef]
- Shintani, T.; Liu, W.-T.; Hanada, S.; Kamagata, Y.; Miyaoka, S.; Suzuki, T.; Nakamura, K. Micropruina glycogenica gen. nov., sp. nov., a new Gram-positive glycogen-accumulating bacterium isolated from activated sludge. Int. J. Syst. Evol. Microbiol. 2000, 50, 201–207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Preiss, J.; Romeo, T. Physiology, biochemistry and genetics of bacterial glycogen synthesis. In Advances in Microbial Physiology; Elsevier: Amsterdam, The Netherlands, 1990; Volume 30, pp. 183–238. [Google Scholar]
- Aikawa, S.; Nishida, A.; Ho, S.-H.; Chang, J.-S.; Hasunuma, T.; Kondo, A. Glycogen production for biofuels by the euryhaline cyanobacteria Synechococcus sp. strain PCC 7002 from an oceanic environment. Biotechnol. Biofuels 2014, 7, 88. [Google Scholar] [CrossRef] [Green Version]
- Brown, M.J.; Lester, J.N. Comparison of bacterial extracellular polymer extraction methods. Appl. Environ. Microb. 1980, 40, 179–185. [Google Scholar] [CrossRef] [Green Version]
- Iglesias, A.A.; Preiss, J. Bacterial glycogen and plant starch biosynthesis. Biochem. Educ. 1992, 20, 196–203. [Google Scholar] [CrossRef]
- Sambou, T.; Dinadayala, P.; Stadthagen, G.; Barilone, N.; Bordat, Y.; Constant, P.; Levillain, F.; Neyrolles, O.; Gicquel, B.; Lemassu, A. Capsular glucan and intracellular glycogen of Mycobacterium tuberculosis: Biosynthesis and impact on the persistence in mice. Mol. Microbiol 2008, 70, 762–774. [Google Scholar] [CrossRef] [Green Version]
- Quilès, F.; Polyakov, P.; Humbert, F.O.; Francius, G.G. Production of extracellular glycogen by Pseudomonas fluorescens: Spectroscopic evidence and conformational analysis by biomolecular recognition. Biomacromolecules 2012, 13, 2118–2127. [Google Scholar] [CrossRef]
- Celik, G.Y.; Aslim, B.; Beyatli, Y. Characterization and production of the exopolysaccharide (EPS) from Pseudomonas aeruginosa G1 and Pseudomonas putida G12 strains. Carbohydr. Polym. 2008, 73, 178–182. [Google Scholar] [CrossRef]
- Rehm, B.H.; Moradali, M.F. Alginates and Their Biomedical Applications; Springer: Berlin, Germany, 2018. [Google Scholar]
- Bakkevig, K.; Sletta, H.; Gimmestad, M.; Aune, R.; Ertesvåg, H.; Degnes, K.; Christensen, B.E.; Ellingsen, T.E.; Valla, S. Role of the Pseudomonas fluorescens alginate lyase (AlgL) in clearing the periplasm of alginates not exported to the extracellular environment. J. Bacteriol. 2005, 187, 8375–8384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hay, I.D.; Rehman, Z.U.; Moradali, M.F.; Wang, Y.; Rehm, B.H. Microbial alginate production, modification and its applications. Microb. Biotechnol. 2013, 6, 637–650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robles-Price, A.; Wong, T.Y.; Sletta, H.; Valla, S.; Schiller, N.L. AlgX is a periplasmic protein required for alginate biosynthesis in Pseudomonas aeruginosa. J. Bacteriol. 2004, 186, 7369–7377. [Google Scholar] [CrossRef] [Green Version]
- Szekalska, M.; Puciłowska, A.; Szymańska, E.; Ciosek, P.; Winnicka, K. Alginate: Current use and future perspectives in pharmaceutical and biomedical applications. Int. J. Polym. Sci. 2016, 2016, 7697031. [Google Scholar] [CrossRef] [Green Version]
- Schmid, J.; Sieber, V.; Rehm, B. Bacterial exopolysaccharides: Biosynthesis pathways and engineering strategies. Front. Microbiol. 2015, 6, 496. [Google Scholar] [CrossRef] [Green Version]
- Nordgård, C.T.; Nonstad, U.; Olderøy, M.Ø.; Espevik, T.; Draget, K.I. Alterations in mucus barrier function and matrix structure induced by guluronate oligomers. Biomacromolecules 2014, 15, 2294–2300. [Google Scholar] [CrossRef]
- Powell, L.C.; Pritchard, M.F.; Emanuel, C.; Onsøyen, E.; Rye, P.D.; Wright, C.J.; Hill, K.E.; Thomas, D.W. A nanoscale characterization of the interaction of a novel alginate oligomer with the cell surface and motility of Pseudomonas aeruginosa. Am. J. Respir. Cell Mol. Biol. 2014, 50, 483–492. [Google Scholar] [CrossRef] [Green Version]
- Powell, L.C.; Sowedan, A.; Khan, S.; Wright, C.J.; Hawkins, K.; Onsøyen, E.; Myrvold, R.; Hill, K.E.; Thomas, D.W. The effect of alginate oligosaccharides on the mechanical properties of Gram-negative biofilms. Biofouling 2013, 29, 413–421. [Google Scholar] [CrossRef]
- Sun, J.; Tan, H. Alginate-based biomaterials for regenerative medicine applications. Materials 2013, 6, 1285–1309. [Google Scholar] [CrossRef]
- Campa, C.; Holtan, S.; Nilsen, N.; Bjerkan, T.M.; Stokke, B.T.; SKJåK-BRæK, G. Biochemical analysis of the processive mechanism for epimerization of alginate by mannuronan C-5 epimerase AlgE4. Biochem. J. 2004, 381, 155–164. [Google Scholar] [CrossRef] [Green Version]
- Falkeborg, M.; Cheong, L.-Z.; Gianfico, C.; Sztukiel, K.M.; Kristensen, K.; Glasius, M.; Xu, X.; Guo, Z. Alginate oligosaccharides: Enzymatic preparation and antioxidant property evaluation. Food Chem. 2014, 164, 185–194. [Google Scholar] [CrossRef]
- Yang, J.-S.; Xie, Y.-J.; He, W. Research progress on chemical modification of alginate: A review. Carbohydr. Polym. 2011, 84, 33–39. [Google Scholar] [CrossRef]
- Pawar, S.N.; Edgar, K.J. Alginate derivatization: A review of chemistry, properties and applications. Biomaterials 2012, 33, 3279–3305. [Google Scholar] [CrossRef]
- Wong, T.W. Alginate graft copolymers and alginate–co-excipient physical mixture in oral drug delivery. J. Pharm. Pharmacol. 2011, 63, 1497–1512. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.Y.; Mooney, D.J. Alginate: Properties and biomedical applications. Prog. Polym. Sci. 2012, 37, 106–126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bouhadir, K.H.; Alsberg, E.; Mooney, D.J. Hydrogels for combination delivery of antineoplastic agents. Biomaterials 2001, 22, 2625–2633. [Google Scholar] [CrossRef]
- Lucinda-Silva, R.M.; Salgado, H.R.N.; Evangelista, R.C. Alginate–chitosan systems: In vitro controlled release of triamcinolone and in vivo gastrointestinal transit. Carbohydr. Polym. 2010, 81, 260–268. [Google Scholar] [CrossRef]
- Chang, C.-H.; Lin, Y.-H.; Yeh, C.-L.; Chen, Y.-C.; Chiou, S.-F.; Hsu, Y.-M.; Chen, Y.-S.; Wang, C.-C. Nanoparticles incorporated in pH-sensitive hydrogels as amoxicillin delivery for eradication of Helicobacter pylori. Biomacromolecules 2010, 11, 133–142. [Google Scholar] [CrossRef]
- Cao, L.; Mooney, D.J. Spatiotemporal control over growth factor signaling for therapeutic neovascularization. Adv. Drug Deliv. Rev. 2007, 59, 1340–1350. [Google Scholar] [CrossRef] [Green Version]
- Rabbany, S.Y.; Pastore, J.; Yamamoto, M.; Miller, T.; Rafii, S.; Aras, R.; Penn, M. Continuous delivery of stromal cell-derived factor-1 from alginate scaffolds accelerates wound healing. Cell Transplant. 2010, 19, 399–408. [Google Scholar] [CrossRef] [Green Version]
- Lópiz-Morales, Y.; Abarrategi, A.; Ramos, V.; Moreno-Vicente, C.; López-Durán, L.; López-Lacomba, J.L.; Marco, F. In vivo comparison of the effects of rhBMP-2 and rhBMP-4 in osteochondral tissue regeneration. Eur. Cell Mater. 2010, 20, e78. [Google Scholar] [CrossRef] [PubMed]
- Chang, S.C.; Tobias, G.; Roy, A.K.; Vacanti, C.A.; Bonassar, L.J. Tissue engineering of autologous cartilage for craniofacial reconstruction by injection molding. Plast. Reconstr. Surg. 2003, 112, 793–799. [Google Scholar] [CrossRef] [PubMed]
- Zmora, S.; Glicklis, R.; Cohen, S. Tailoring the pore architecture in 3-D alginate scaffolds by controlling the freezing regime during fabrication. Biomaterials 2002, 23, 4087–4094. [Google Scholar] [CrossRef]
- Prang, P.; Müller, R.; Eljaouhari, A.; Heckmann, K.; Kunz, W.; Weber, T.; Faber, C.; Vroemen, M.; Bogdahn, U.; Weidner, N. The promotion of oriented axonal regrowth in the injured spinal cord by alginate-based anisotropic capillary hydrogels. Biomaterials 2006, 27, 3560–3569. [Google Scholar] [CrossRef]
- Meyer, K.; Palmer, J.W. The polysaccharide of the vitreous humor. J. Biol. Chem. 1934, 107, 629–634. [Google Scholar] [CrossRef]
- Fallacara, A.; Baldini, E.; Manfredini, S.; Vertuani, S. Hyaluronic acid in the third millennium. Polymers 2018, 10, 701. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Atkins, E.; Sheehan, J. Structure for hyaluronic acid. Nat. New Biol. 1972, 235, 253–254. [Google Scholar] [CrossRef]
- Weissmann, B.; Meyer, K. Structure of hyaluronic acid. The glucuronidic linkage. J. Am. Chem. Soc. 1952, 74, 4729. [Google Scholar] [CrossRef]
- Ward, P.D.; Thibeault, S.L.; Gray, S.D. Hyaluronic acid: Its role in voice. J. Voice 2002, 16, 303–309. [Google Scholar] [CrossRef]
- Anilkumar, T.; Muhamed, J.; Jose, A.; Jyothi, A.; Mohanan, P.; Krishnan, L.K. Advantages of hyaluronic acid as a component of fibrin sheet for care of acute wound. Biologicals 2011, 39, 81–88. [Google Scholar] [CrossRef]
- Frasca, P.; Harper, R.; Katz, J. Scanning electron microscopy studies of collagen, mineral and ground substance in human cortical bone. Scan. Electron. Microsc. 1981, 109, 339–346. [Google Scholar]
- Mathews, M.B.; Decker, L. Comparative studies of water sorption of hyaline cartilage. Biochim. Biophys. Acta BBA Gen. Subj. 1977, 497, 151–159. [Google Scholar] [CrossRef]
- Reddi, A.; Piez, K.A. Extracellular Matrix Biochemistry; Elsevier: New York, NY, USA, 1984. [Google Scholar]
- Hardingham, T. Chondroitin sulfate and joint disease. Osteoarthr. Cartil. 1998, 6, 3–5. [Google Scholar] [CrossRef] [Green Version]
- Rosines, E.; Schmidt, H.J.; Nigam, S.K. The effect of hyaluronic acid size and concentration on branching morphogenesis and tubule differentiation in developing kidney culture systems: Potential applications to engineering of renal tissues. Biomaterials 2007, 28, 4806–4817. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, X.; Zaia, J. Organ-specific heparan sulfate structural phenotypes. J. Biol. Chem. 2009, 284, 11806–11814. [Google Scholar] [CrossRef] [Green Version]
- Jahn, M.; Baynes, J.W.; Spiteller, G. The reaction of hyaluronic acid and its monomers, glucuronic acid and N-acetylglucosamine, with reactive oxygen species. Carbohydr. Res. 1999, 321, 228–234. [Google Scholar] [CrossRef]
- Kogan, G.; Šoltés, L.; Stern, R.; Gemeiner, P. Hyaluronic acid: A natural biopolymer with a broad range of biomedical and industrial applications. Biotechnol. Lett. 2007, 29, 17–25. [Google Scholar] [CrossRef] [PubMed]
- Greene, G.W.; Zappone, B.; Banquy, X.; Lee, D.W.; Söderman, O.; Topgaard, D.; Israelachvili, J.N. Hyaluronic acid–collagen network interactions during the dynamic compression and recovery of cartilage. Soft Matter 2012, 8, 9906–9914. [Google Scholar] [CrossRef]
- Lai, V.K.; Nedrelow, D.S.; Lake, S.P.; Kim, B.; Weiss, E.M.; Tranquillo, R.T.; Barocas, V.H. Swelling of collagen-hyaluronic acid co-gels: An in vitro residual stress model. Ann. Biomed. Eng. 2016, 44, 2984–2993. [Google Scholar] [CrossRef] [Green Version]
- Miranda, D.G.; Malmonge, S.M.; Campos, D.M.; Attik, N.G.; Grosgogeat, B.; Gritsch, K. A chitosan-hyaluronic acid hydrogel scaffold for periodontal tissue engineering. J. Biomed. Mater. Res. Part B Appl. Biomater. 2016, 104, 1691–1702. [Google Scholar] [CrossRef]
- Sionkowska, A.; Kaczmarek, B.; Lewandowska, K.; Grabska, S.; Pokrywczyńska, M.; Kloskowski, T.; Drewa, T. 3D composites based on the blends of chitosan and collagen with the addition of hyaluronic acid. Int. J. Biol. Macromol. 2016, 89, 442–448. [Google Scholar] [CrossRef] [PubMed]
- Beasley, K.L.; Weiss, M.A.; Weiss, R.A. Hyaluronic acid fillers: A comprehensive review. Facial Plast. Surg. 2009, 25, 086–094. [Google Scholar] [CrossRef]
- Clark, C.P., III. Animal-based hyaluronic acid fillers: Scientific and technical considerations. Plast. Reconstr. Surg. 2007, 120, 27S–32S. [Google Scholar] [CrossRef] [PubMed]
- Edwards, P.C.; Fantasia, J.E. Review of long-term adverse effects associated with the use of chemically-modified animal and nonanimal source hyaluronic acid dermal fillers. Clin. Interv. Aging 2007, 2, 509. [Google Scholar] [CrossRef] [Green Version]
- Romagnoli, M.; Belmontesi, M. Hyaluronic acid–based fillers: Theory and practice. Clin. Dermatol. 2008, 26, 123–159. [Google Scholar] [CrossRef] [PubMed]
- Raeissadat, S.A.; Rayegani, S.M.; Forogh, B.; Abadi, P.H.; Moridnia, M.; Dehgolan, S.R. Intra-articular ozone or hyaluronic acid injection: Which one is superior in patients with knee osteoarthritis? A 6-month randomized clinical trial. J. Pain Res. 2018, 11, 111. [Google Scholar] [CrossRef] [Green Version]
- Lin, W.; Liu, Z.; Kampf, N.; Klein, J. The Role of Hyaluronic Acid in Cartilage Boundary Lubrication. Cells 2020, 9, 1606. [Google Scholar] [CrossRef]
- Das, S.; Banquy, X.; Zappone, B.; Greene, G.W.; Jay, G.D.; Israelachvili, J.N. Synergistic interactions between grafted hyaluronic acid and lubricin provide enhanced wear protection and lubrication. Biomacromolecules 2013, 14, 1669–1677. [Google Scholar] [CrossRef]
- Harrington, S.; Ott, L.; Karanu, F.; Ramachandran, K.; Stehno-Bittel, L. A Versatile Microencapsulation Platform for Hyaluronic Acid and Polyethylene Glycol. Tissue Eng. Part A 2021, 27, 153–164. [Google Scholar] [CrossRef] [Green Version]
- Harrington, S.; Williams, J.; Rawal, S.; Ramachandran, K.; Stehno-Bittel, L. Hyaluronic acid/collagen hydrogel as an alternative to alginate for long-term immunoprotected islet transplantation. Tissue Eng. Part A 2017, 23, 1088–1099. [Google Scholar] [CrossRef] [PubMed]
- Resnick, N.M.; Clarke, M.R.; Siegfried, J.M.; Landreneau, R.; Asman, D.C.; Ge, L.; Kierstead, L.S.; Dougherty, G.D.; Cooper, D.L. Expression of the cell adhesion molecule CD44 in human lung tumors and cell lines. Mol. Diagn. 1998, 3, 93–103. [Google Scholar] [CrossRef]
- Yang, B.; Yang, B.L.; Savani, R.C.; Turley, E.A. Identification of a common hyaluronan binding motif in the hyaluronan binding proteins RHAMM, CD44 and link protein. EMBO J. 1994, 13, 286–296. [Google Scholar] [CrossRef]
- Penno, M.B.; August, J.T.; Baylin, S.B.; Mabry, M.; Linnoila, R.I.; Lee, V.S.; Croteau, D.; Yang, X.L.; Rosada, C. Expression of CD44 in human lung tumors. Cancer Res. 1994, 54, 1381–1387. [Google Scholar]
- Lee, S.Y.; Kang, M.S.; Jeong, W.Y.; Han, D.-W.; Kim, K.S. Hyaluronic Acid-Based Theranostic Nanomedicines for Targeted Cancer Therapy. Cancers 2020, 12, 940. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Le, T.M.D.; Bui, Q.N.; Yang, H.Y.; Lee, D.S. Tumor acidity and CD44 dual targeting hyaluronic acid-coated gold nanorods for combined chemo-and photothermal cancer therapy. Carbohydr. Polym. 2019, 226, 115281. [Google Scholar] [CrossRef]
- Wickens, J.M.; Alsaab, H.O.; Kesharwani, P.; Bhise, K.; Amin, M.C.I.M.; Tekade, R.K.; Gupta, U.; Iyer, A.K. Recent advances in hyaluronic acid-decorated nanocarriers for targeted cancer therapy. Drug Discov. Today 2017, 22, 665–680. [Google Scholar] [CrossRef] [Green Version]
- Yu, M.; Jambhrunkar, S.; Thorn, P.; Chen, J.; Gu, W.; Yu, C. Hyaluronic acid modified mesoporous silica nanoparticles for targeted drug delivery to CD44-overexpressing cancer cells. Nanoscale 2013, 5, 178–183. [Google Scholar] [CrossRef] [PubMed]
- Silvipriya, K.; Kumar, K.K.; Bhat, A.; Kumar, B.D.; John, A.; Lakshmanan, P. Collagen: Animal sources and biomedical application. J. Appl. Pharm. Sci. 2015, 5, 123–127. [Google Scholar] [CrossRef] [Green Version]
- Van Der Laan, L.J.; Lockey, C.; Griffeth, B.C.; Frasier, F.S.; Wilson, C.A.; Onions, D.E.; Hering, B.J.; Long, Z.; Otto, E.; Torbett, B.E. Infection by porcine endogenous retrovirus after islet xenotransplantation in SCID mice. Nature 2000, 407, 90–94. [Google Scholar] [CrossRef] [PubMed]
- Moses, A.E.; Wessels, M.R.; Zalcman, K.; Albertí, S.; Natanson-Yaron, S.; Menes, T.; Hanski, E. Relative contributions of hyaluronic acid capsule and M protein to virulence in a mucoid strain of the group A Streptococcus. Infect. Immun. 1997, 65, 64–71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wessels, M.R.; Moses, A.E.; Goldberg, J.B.; DiCesare, T.J. Hyaluronic acid capsule is a virulence factor for mucoid group A streptococci. Proc. Natl. Acad. Sci. USA 1991, 88, 8317–8321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeng, Y.; Zeng, W.; Zhou, Q.; Jia, X.; Li, J.; Yang, Z.; Hao, Y.; Liu, J. Hyaluronic acid mediated biomineralization of multifunctional ceria nanocomposites as ROS scavengers and tumor photodynamic therapy agents. J. Mat. Chem. B 2019, 7, 3210–3219. [Google Scholar] [CrossRef]
- Gunasekaran, V.; Gowdhaman, D.; Ponnusami, V. Role of membrane proteins in bacterial synthesis of hyaluronic acid and their potential in industrial production. Int. J. Biol. Macromol. 2020, 164, 1916–1926. [Google Scholar] [CrossRef] [PubMed]
- Chong, B.F.; Nielsen, L.K. Aerobic cultivation of Streptococcus zooepidemicus and the role of NADH oxidase. Biochem. Eng. J. 2003, 16, 153–162. [Google Scholar] [CrossRef]
- Mohan, N.; Tadi, S.R.R.; Pavan, S.S.; Sivaprakasam, S. Deciphering the role of dissolved oxygen and N-acetyl glucosamine in governing higher molecular weight hyaluronic acid synthesis in Streptococcus zooepidemicus cell factory. Appl. Microbiol. Biotech. 2020, 104, 3349–3365. [Google Scholar] [CrossRef] [PubMed]
- Arslan, N.P.; Aydogan, M.N. Evaluation of Sheep Wool Protein Hydrolysate and Molasses as Low-Cost Fermentation Substrates for Hyaluronic Acid Production by Streptococcus zooepidemicus ATCC 35246. Waste Biomass Valor. 2020, 12, 925–935. [Google Scholar] [CrossRef]
- Chien, L.-J.; Lee, C.-K. Hyaluronic acid production by recombinant Lactococcus lactis. Appl. Microbiol. Biotech. 2007, 77, 339–346. [Google Scholar] [CrossRef]
- Yu, H.; Stephanopoulos, G. Metabolic engineering of Escherichia coli for biosynthesis of hyaluronic acid. Metab. Eng. 2008, 10, 24–32. [Google Scholar] [CrossRef]
- Li, Y.; Li, G.; Zhao, X.; Shao, Y.; Wu, M.; Ma, T. Regulation of hyaluronic acid molecular weight and titer by temperature in engineered Bacillus subtilis. 3 Biotech. 2019, 9, 225. [Google Scholar] [CrossRef]
- Prajapati, V.D.; Jani, G.K.; Zala, B.S.; Khutliwala, T.A. An insight into the emerging exopolysaccharide gellan gum as a novel polymer. Carbohydr. Polym. 2013, 93, 670–678. [Google Scholar] [CrossRef] [PubMed]
- Manjanna, K. Natural polysaccharide hydrogels as novel excipients for modified drug delivery systems: A review. Int. J. Chemtech. Res. 2010, 2, 509–525. [Google Scholar]
- Pszczola, D.E. Gellan gum wins IFT’s food technology industrial achievement award. Food Technol. 1993, 47, 94–96. [Google Scholar]
- Zhang, H.; Zhang, F.; Yuan, R. Applications of natural polymer-based hydrogels in the food industry. In Hydrogels Based on Natural Polymers; Elsevier: Amsterdam, The Netherlands, 2020; pp. 357–410. [Google Scholar]
- Kang, K.S.; Veeder, G.T.; Mirrasoul, P.J.; Kaneko, T.; Cottrell, I.W. Agar-like polysaccharide produced by a Pseudomonas species: Production and basic properties. Appl. Environ. Microb. 1982, 43, 1086–1091. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morris, E.R.; Nishinari, K.; Rinaudo, M. Gelation of gellan—A review. Food Hydrocolloid 2012, 28, 373–411. [Google Scholar] [CrossRef]
- Osmałek, T.; Froelich, A.; Tasarek, S. Application of gellan gum in pharmacy and medicine. Int. J. Pharm. 2014, 466, 328–340. [Google Scholar] [CrossRef]
- Mahdi, M.H.; Conway, B.R.; Smith, A.M. Development of mucoadhesive sprayable gellan gum fluid gels. Int. J. Pharm. 2015, 488, 12–19. [Google Scholar] [CrossRef] [Green Version]
- Zia, K.M.; Tabasum, S.; Khan, M.F.; Akram, N.; Akhter, N.; Noreen, A.; Zuber, M. Recent trends on gellan gum blends with natural and synthetic polymers: A review. Int. J. Biol. Macromol. 2018, 109, 1068–1087. [Google Scholar] [CrossRef]
- Bajaj, I.B.; Survase, S.A.; Saudagar, P.S.; Singhal, R.S. Gellan gum: Fermentative production, downstream processing and applications. Food Technol. Biotech. 2007, 45, 341–354. [Google Scholar]
- Bacelar, A.H.; Silva-Correia, J.; Oliveira, J.M.; Reis, R.L. Recent progress in gellan gum hydrogels provided by functionalization strategies. J. Mat. Chem. B 2016, 4, 6164–6174. [Google Scholar] [CrossRef] [Green Version]
- Novac, O.; Lisa, G.; Profire, L.; Tuchilus, C.; Popa, M. Antibacterial quaternized gellan gum based particles for controlled release of ciprofloxacin with potential dermal applications. Mater. Sci. Eng. C 2014, 35, 291–299. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Kaur, P.; Bernela, M.; Rani, R.; Thakur, R. Ketoconazole encapsulated in chitosan-gellan gum nanocomplexes exhibits prolonged antifungal activity. Int. J. Biol. Macromol. 2016, 93, 988–994. [Google Scholar] [CrossRef]
- Liu, L.; Wang, B.; Gao, Y.; Bai, T.-C. Chitosan fibers enhanced gellan gum hydrogels with superior mechanical properties and water-holding capacity. Carbohydr. Polym. 2013, 97, 152–158. [Google Scholar] [CrossRef]
- Stevens, L.; Gilmore, K.J.; Wallace, G.G. Tissue engineering with gellan gum. Biomater. Sci. 2016, 4, 1276–1290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lozano, R.; Stevens, L.; Thompson, B.C.; Gilmore, K.J.; Gorkin, R., III; Stewart, E.M.; Panhuis, M.; Romero-Ortega, M.; Wallace, G.G. 3D printing of layered brain-like structures using peptide modified gellan gum substrates. Biomaterials 2015, 67, 264–273. [Google Scholar] [CrossRef] [PubMed]
- Meier, C.; Welland, M.E. Wet-spinning of amyloid protein nanofibers into multifunctional high-performance biofibers. Biomacromolecules 2011, 12, 3453–3459. [Google Scholar] [CrossRef] [PubMed]
- Da Silva, L.P.; Cerqueira, M.T.; Sousa, R.A.; Reis, R.L.; Correlo, V.M.; Marques, A.P. Engineering cell-adhesive gellan gum spongy-like hydrogels for regenerative medicine purposes. Acta Biomater. 2014, 10, 4787–4797. [Google Scholar] [CrossRef]
- Oliveira, J.T.; Gardel, L.S.; Rada, T.; Martins, L.; Gomes, M.E.; Reis, R.L. Injectable gellan gum hydrogels with autologous cells for the treatment of rabbit articular cartilage defects. J. Orthop. Res. 2010, 28, 1193–1199. [Google Scholar] [CrossRef] [Green Version]
- Petri, D.F. Xanthan gum: A versatile biopolymer for biomedical and technological applications. J. Appl. Polym. Sci. 2015, 132. [Google Scholar] [CrossRef] [Green Version]
- Tao, F.; Wang, X.; Ma, C.; Yang, C.; Tang, H.; Gai, Z.; Xu, P. Genome sequence of Xanthomonas campestris JX, an industrially productive strain for Xanthan gum. Am. Soc. Microbiol. 2012. [Google Scholar] [CrossRef]
- Janse, J.D. Phytobacteriology: Principles and Practice; Cabi: Wallingford, UK, 2005. [Google Scholar]
- Patel, J.; Maji, B.; Moorthy, N.H.N.; Maiti, S. Xanthan gum derivatives: Review of synthesis, properties and diverse applications. RSC Adv. 2020, 10, 27103–27136. [Google Scholar] [CrossRef]
- Garcıa-Ochoa, F.; Santos, V.; Casas, J.; Gómez, E. Xanthan gum: Production, recovery, and properties. Biotechnol. Adv. 2000, 18, 549–579. [Google Scholar] [CrossRef]
- Ielpi, L.; Couso, R.; Dankert, M. Sequential assembly and polymerization of the polyprenol-linked pentasaccharide repeating unit of the xanthan polysaccharide in Xanthomonas campestris. J. Bacteriol. 1993, 175, 2490–2500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, G.; Wang, G.; Zhu, X.; Shao, H.; Liu, F.; Yang, P.; Ying, Y.; Wang, F.; Ling, P. Preparation of xanthan gum injection and its protective effect on articular cartilage in the development of osteoarthritis. Carbohydr. Polym. 2012, 87, 1837–1842. [Google Scholar] [CrossRef]
- Palaniraj, A.; Jayaraman, V. Production, recovery and applications of xanthan gum by Xanthomonas campestris. J. Food Eng. 2011, 106, 1–12. [Google Scholar] [CrossRef]
- Camesano, T.A.; Wilkinson, K.J. Single molecule study of xanthan conformation using atomic force microscopy. Biomacromolecules 2001, 2, 1184–1191. [Google Scholar] [CrossRef]
- Carmona, J.A.; Lucas, A.; Ramírez, P.; Calero, N.; Muñoz, J. Nonlinear and linear viscoelastic properties of a novel type of xanthan gum with industrial applications. Rheol. Acta 2015, 54, 993–1001. [Google Scholar] [CrossRef]
- Junyaprasert, V.B.; Manwiwattanakul, G. Release profile comparison and stability of diltiazem–resin microcapsules in sustained release suspensions. Int. J. Pharm. 2008, 352, 81–91. [Google Scholar] [CrossRef]
- Psomas, S.; Liakopoulou-Kyriakides, M.; Kyriakidis, D. Optimization study of xanthan gum production using response surface methodology. Biochem. Eng. J. 2007, 35, 273–280. [Google Scholar] [CrossRef]
- Liu, Z.; Yao, P. Injectable shear-thinning xanthan gum hydrogel reinforced by mussel-inspired secondary crosslinking. RSC Adv. 2015, 5, 103292–103301. [Google Scholar] [CrossRef]
- Bueno, V.B.; Bentini, R.; Catalani, L.H.; Petri, D.F.S. Synthesis and swelling behavior of xanthan-based hydrogels. Carbohydr. Polym. 2013, 92, 1091–1099. [Google Scholar] [CrossRef] [Green Version]
- Hoffman, A.S. Hydrogels for biomedical applications. Adv. Drug Deliv. Rev. 2012, 64, 18–23. [Google Scholar] [CrossRef]
- Alvarez-Mancenido, F.; Landin, M.; Martinez-Pacheco, R. Konjac glucomannan/xanthan gum enzyme sensitive binary mixtures for colonic drug delivery. Eur. J. Pharm. Biopharm. 2008, 69, 573–581. [Google Scholar] [CrossRef]
- Sinha, V.; Kumria, R. Binders for colon specific drug delivery: An in vitro evaluation. Int. J. Pharm. 2002, 249, 23–31. [Google Scholar] [CrossRef]
- Sethi, S.; Kaith, B.S.; Kaur, M.; Sharma, N.; Kumar, V. Cross-linked xanthan gum–starch hydrogels as promising materials for controlled drug delivery. Cellulose 2020, 1–25. [Google Scholar] [CrossRef]
- Hu, X.; Wang, K.; Yu, M.; He, P.; Qiao, H.; Zhang, H.; Wang, Z. Characterization and Antioxidant Activity of a Low-Molecular-Weight Xanthan Gum. Biomolecules 2019, 9, 730. [Google Scholar] [CrossRef] [Green Version]
- Abu-Huwaij, R.; Obaidat, R.M.; Sweidan, K.; Al-Hiari, Y. Formulation and in vitro evaluation of xanthan gum or carbopol 934-based mucoadhesive patches, loaded with nicotine. Aaps Pharmscitech. 2011, 12, 21–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manconi, M.; Mura, S.; Manca, M.L.; Fadda, A.M.; Dolz, M.; Hernandez, M.; Casanovas, A.; Díez-Sales, O. Chitosomes as drug delivery systems for C-phycocyanin: Preparation and characterization. Int. J. Pharm. 2010, 392, 92–100. [Google Scholar] [CrossRef]
- Shiledar, R.R.; Tagalpallewar, A.A.; Kokare, C.R. Formulation and in vitro evaluation of xanthan gum-based bilayered mucoadhesive buccal patches of zolmitriptan. Carbohydr. Polym. 2014, 101, 1234–1242. [Google Scholar] [CrossRef]
- Bueno, V.B.; Bentini, R.; Catalani, L.H.; Barbosa, L.R.; Petri, D.F.S. Synthesis and characterization of xanthan–hydroxyapatite nanocomposites for cellular uptake. Mater. Sci. Eng. C 2014, 37, 195–203. [Google Scholar] [CrossRef]
- Bueno, V.B.; Takahashi, S.H.; Catalani, L.H.; de Torresi, S.I.C.; Petri, D.F.S. Biocompatible xanthan/polypyrrole scaffolds for tissue engineering. Mater. Sci. Eng. C 2015, 52, 121–128. [Google Scholar] [CrossRef] [PubMed]
- Darzi, H.H.; Larimi, S.G.; Darzi, G.N. Synthesis, characterization and physical properties of a novel xanthan gum/polypyrrole nanocomposite. Synth. Met. 2012, 162, 236–239. [Google Scholar] [CrossRef]
- Glaser, T.; Bueno, V.B.; Cornejo, D.R.; Petri, D.F.; Ulrich, H. Neuronal adhesion, proliferation and differentiation of embryonic stem cells on hybrid scaffolds made of xanthan and magnetite nanoparticles. Biomed. Mater. 2015, 10, 045002. [Google Scholar] [CrossRef] [PubMed]
- McIntosh, M.; Stone, B.A.; Stanisich, V.A. Curdlan and other bacterial (1→3)-β-d-glucans. Appl. Microbiol. Biotech. 2005, 68, 163–173. [Google Scholar] [CrossRef] [PubMed]
- Harada, T.; Masada, M.; Fujimori, K.; Maeda, I. Production of a firm, resilient gel-forming polysaccharide by a mutant of Alcaligenes faecalis var. myxogenes 10 C3. Agric. Biol. Chem. 1966, 30, 196–198. [Google Scholar] [CrossRef] [Green Version]
- Zhang, R.; Edgar, K.J. Properties, chemistry, and applications of the bioactive polysaccharide curdlan. Biomacromolecules 2014, 15, 1079–1096. [Google Scholar] [CrossRef]
- US Food and Drug Administration. CFR 172-Food Additives Permitted for Direct Addition to Food for Human Consumption: Curdlan; Federal Register 61; US Food and Drug Administration: White Oak, MD, USA, 2020; pp. 65941–65942.
- Steinbüchel, A.; Hofrichter, M.; Koyama, T.; Vandamme, E.J.; de Baets, S. Biopolymers Online: Biology, Chemistry, Biotechnology, Applications: Polysaccharides 1: Polysaccharides from Prokaryotes, 1st ed.; Wiley: Weinheim, Germany, 2002; Volume 5, pp. 135–158. [Google Scholar]
- Funami, T.; Funami, M.; Tawada, T.; Nakao, Y. Decreasing oil uptake of doughnuts during deep-fat frying using curdlan. J. Food Sci. 1999, 64, 883–888. [Google Scholar] [CrossRef]
- Yotsuzuka, F. Curdlan. In Handbook of Dietary Fiber; CRC Press: Boca Raton, FL, USA, 2001; pp. 737–757. [Google Scholar]
- Kasai, N.; Harada, T. Ultrastructure of Curdlan; ACS Publications: Washington, DC, USA, 1980. [Google Scholar]
- Bohn, J.A.; BeMiller, J.N. (1→3)-β-d-Glucans as biological response modifiers: A review of structure-functional activity relationships. Carbohydr. Polym. 1995, 28, 3–14. [Google Scholar] [CrossRef]
- Vannucci, L.; Krizan, J.; Sima, P.; Stakheev, D.; Caja, F.; Rajsiglova, L.; Horak, V.; Saieh, M. Immunostimulatory properties and antitumor activities of glucans. Int. J. Oncol. 2013, 43, 357–364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stone, B.; Clarke, A. Chemistry and Biology of (1-3)-β-Glucans; La Trobe University Press: Bundoora, Australia, 1992; p. 517. [Google Scholar]
- Kanke, M.; Tanabe, E.; Katayama, H.; Koda, Y.; Yoshitomi, H. Application of curdlan to controlled drug delivery. III. Drug release from sustained release suppositories in vitro. Biol. Pharm. Bull. 1995, 18, 1154–1158. [Google Scholar] [CrossRef] [Green Version]
- Na, K.; Park, K.-H.; Kim, S.W.; Bae, Y.H. Self-assembled hydrogel nanoparticles from curdlan derivatives: Characterization, anti-cancer drug release and interaction with a hepatoma cell line (HepG2). J. Control. Release 2000, 69, 225–236. [Google Scholar] [CrossRef]
- Delatte, S.J.; Evans, J.; Hebra, A.; Adamson, W.; Othersen, H.B.; Tagge, E.P. Effectiveness of beta-glucan collagen for treatment of partial-thickness burns in children. J. Pediatric Surg. 2001, 36, 113–118. [Google Scholar] [CrossRef]
- Basha, R.Y.; Sampath-Kumar, T.; Doble, M. Electrospun nanofibers of curdlan (β-1, 3 glucan) blend as a potential skin scaffold material. Macromol. Mater. Eng. 2017, 302, 1600417. [Google Scholar] [CrossRef]
- Hsieh, W.-C.; Hsu, C.-C.; Shiu, L.-Y.; Zeng, Y.-J. Biocompatible testing and physical properties of curdlan-grafted poly (vinyl alcohol) scaffold for bone tissue engineering. Carbohydr. Polym. 2017, 157, 1341–1348. [Google Scholar] [CrossRef]
- Keshavarz, T.; Roy, I. Polyhydroxyalkanoates: Bioplastics with a green agenda. Curr. Opin. Microbiol. 2010, 13, 321–326. [Google Scholar] [CrossRef]
- Berezina, N.; Martelli, S.M. Polyhydroxyalkanoates: Structure, properties and sources. RSC Green Chem. Ser. 2014, 30, 18. [Google Scholar]
- Martínez-Abad, A.; Cabedo, L.; Oliveira, C.S.; Hilliou, L.; Reis, M.; Lagarón, J.M. Characterization of polyhydroxyalkanoate blends incorporating unpurified biosustainably produced poly (3-hydroxybutyrate-co-3-hydroxyvalerate). J. Appl. Polym. Sci. 2016, 133. [Google Scholar] [CrossRef]
- Tan, G.-Y.A.; Chen, C.-L.; Li, L.; Ge, L.; Wang, L.; Razaad, I.M.N.; Li, Y.; Zhao, L.; Mo, Y.; Wang, J.-Y. Start a Research on Biopolymer Polyhydroxyalkanoate (PHA): A Review. Polymers 2014, 6, 706–754. [Google Scholar] [CrossRef] [Green Version]
- Li, L.Z.; Huang, W.; Wang, B.J.; Wei, W.F.; Gu, Q.; Chen, P. Properties and structure of polylactide/poly (3-hydroxybutyrate-co-3-hydroxyvalerate) (PLA/PHBV) blend fibers. Polymer 2015, 68, 183–194. [Google Scholar] [CrossRef]
- Liu, Q.S.; Zhang, H.X.; Deng, B.Y.; Zhao, X.Y. Poly(3-hydroxybutyrate) and Poly(3-hydroxybutyrate-co-3-hydroxyvalerate): Structure, Property, and Fiber. Int. J. Polym. Sci. 2014, 2014, 374368. [Google Scholar] [CrossRef]
- Ouyang, S.P.; Luo, R.C.; Chen, S.S.; Liu, Q.; Chung, A.; Wu, Q.; Chen, G.Q. Production of polyhydroxyalkanoates with high 3-hydroxydodecanoate monomer content by fadB and fadA knockout mutant of Pseudomonas putida KT2442. Biomacromolecules 2007, 8, 2504–2511. [Google Scholar] [CrossRef]
- Bhatia, S.K.; Gurav, R.; Choi, T.R.; Jung, H.R.; Yang, S.Y.; Song, H.S.; Jeon, J.M.; Kim, J.S.; Lee, Y.K.; Yang, Y.H. Poly(3-hydroxybutyrate-co-3-hydroxyhexanoate) production from engineered Ralstonia eutropha using synthetic and anaerobically digested food waste derived volatile fatty acids. Int. J. Biol. Macromol. 2019, 133, 1–10. [Google Scholar] [CrossRef]
- Budde, C.F.; Riedel, S.L.; Willis, L.B.; Rha, C.; Sinskey, A.J. Production of Poly(3-Hydroxybutyrate-co-3-Hydroxyhexanoate) from Plant Oil by Engineered Ralstonia eutropha Strains. Appl. Environ. Microb. 2011, 77, 2847–2854. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grande, D.; Ramier, J.; Versace, D.L.; Renard, E.; Langlois, V. Design of functionalized biodegradable PHA-based electrospun scaffolds meant for tissue engineering applications. New Biotechnol. 2017, 37, 129–137. [Google Scholar] [CrossRef] [PubMed]
- Mozejko-Ciesielska, J.; Szacherska, K.; Marciniak, P. Pseudomonas Species as Producers of Eco-friendly Polyhydroxyalkanoates. J. Polym. Environ. 2019, 27, 1151–1166. [Google Scholar] [CrossRef] [Green Version]
- Mohapatra, S.; Maity, S.; Dash, H.R.; Das, S.; Pattnaik, S.; Rath, C.C.; Samantaray, D. Bacillus and biopolymer: Prospects and challenges. Biochem. Biophys. Rep. 2017, 12, 206–213. [Google Scholar] [CrossRef]
- Basnett, P.; Marcello, E.; Lukasiewicz, B.; Panchal, B.; Nigmatullin, R.; Knowles, J.C.; Roy, I. Biosynthesis and characterization of a novel, biocompatible medium chain length polyhydroxyalkanoate by Pseudomonas mendocina CH50 using coconut oil as the carbon source. J. Mater. Sci. Mater. M 2018, 29, 179. [Google Scholar] [CrossRef]
- Basnett, P.; Lukasiewicz, B.; Marcello, E.; Gura, H.K.; Knowles, J.C.; Roy, I. Production of a novel medium chain length poly(3-hydroxyalkanoate) using unprocessed biodiesel waste and its evaluation as a tissue engineering scaffold. Microb. Biotechnol. 2017, 10, 1384–1399. [Google Scholar] [CrossRef]
- Lukasiewicz, B.; Basnett, P.; Nigmatullin, R.; Matharu, R.; Knowles, J.C.; Roy, I. Binary polyhydroxyalkanoate systems for soft tissue engineering. Acta Biomater. 2018, 71, 225–234. [Google Scholar] [CrossRef] [Green Version]
- Le Meur, S.; Zinn, M.; Egli, T.; Thony-Meyer, L.; Ren, Q. Production of medium-chain-length polyhydroxyalkanoates by sequential feeding of xylose and octanoic acid in engineered Pseudomonas putida KT2440. BMC Biotechnol. 2012, 12, 53. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Horlamus, F.; Henkel, M.; Kovacic, F.; Schlafle, S.; Hausmann, R.; Wittgens, A.; Rosenau, F. Growth of engineered Pseudomonas putida KT2440 on glucose, xylose, and arabinose: Hemicellulose hydrolysates and their major sugars as sustainable carbon sources. GCB Bioenergy 2019, 11, 249–259. [Google Scholar] [CrossRef] [Green Version]
- Salvachua, D.; Rydzak, T.; Auwae, R.; de Capite, A.; Black, B.A.; Bouvier, J.T.; Cleveland, N.S.; Elmore, J.R.; Huenemann, J.D.; Katahira, R.; et al. Metabolic engineering of Pseudomonas putida for increased polyhydroxyalkanoate production from lignin. Microb. Biotechnol. 2019. [Google Scholar] [CrossRef] [Green Version]
- Marcano, A.; Haidar, N.B.; Marais, S.; Valleton, J.M.; Duncan, A.C. Designing Biodegradable PHA-Based 3D Scaffolds with Antibiofilm Properties for Wound Dressings: Optimization of the Microstructure/Nanostructure. ACS Biomater. Sci. Eng. 2017, 3, 3654–3661. [Google Scholar] [CrossRef] [PubMed]
- Brigham, C.J.; Sinskey, A.J. Application of polyhydroxyalkanoates in the Medical Industry. Int. J. Biotechnol. Wellness Ind. 2012, 1, 53–60. [Google Scholar] [CrossRef]
- Shishatskaya, E.I.; Volova, T.G.; Puzyr, A.P.; Mogilnaya, O.A.; Efremov, S.N. Tissue response to the implantation of biodegradable polyhydroxyalkanoate sutures. J. Mater. Sci. Mater. M 2004, 15, 719–728. [Google Scholar] [CrossRef] [PubMed]
- Elmowafy, E.; Abdal-Hay, A.; Skouras, A.; Tiboni, M.; Casettari, L.; Guarino, V. Polyhydroxyalkanoate (PHA): Applications in drug delivery and tissue engineering. Expert Rev. Med. Devices 2019, 16, 467–482. [Google Scholar] [CrossRef]
- Bassas-Galià, M.; Gonzalez, A.; Micaux, F.; Gaillard, V.; Piantini, U.; Schintke, S.; Zinn, M.; Mathieu, M. Chemical modification of polyhydroxyalkanoates (PHAs) for the preparation of hybrid biomaterials. Chim. Int. J. Chem. 2015, 69, 627–630. [Google Scholar] [CrossRef]
- Sadiku, E.R.; Fasiku, V.O.; Owonubi, S.J.; Mukwevho, E.; Aderibigbe, B.A.; Lemmer, Y.; Abbavaram, B.R.; Manjula, B.; Nkuna, C.; Dludlu, M.K.; et al. Polyhydroxyalkanoates (PHAs) as scaffolds for tissue engineering. In Polyhydroxyalkanoates: Biosynthesis, Chemical Structure and Applications; Williams, H., Kelly, P., Eds.; Nova Science Publishers, Inc.: New York, NY, USA, 2018. [Google Scholar]
- Braunegg, G.; Lefebvre, G.; Genser, K.F. Polyhydroxyalkanoates, biopolyesters from renewable resources: Physiological and engineering aspects. J. Biotechnol. 1998, 65, 127–161. [Google Scholar] [CrossRef]
- Rai, R.; Roether, J.A.; Knowles, J.C.; Mordan, N.; Salih, V.; Locke, I.C.; Gordge, M.P.; McCormick, A.; Mohn, D.; Stark, W.J.; et al. Highly elastomeric poly(3-hydroxyoctanoate) based natural polymer composite for enhanced keratinocyte regeneration. Int. J. Polym. Mater. Polym. Biomater. 2017, 66, 326–335. [Google Scholar] [CrossRef]
- Shishatskaya, E.I.; Nikolaeva, E.D.; Vinogradova, O.N.; Volova, T.G. Experimental wound dressings of degradable PHA for skin defect repair. J. Mater. Sci. Mater. Med. 2016, 27, 165. [Google Scholar] [CrossRef] [Green Version]
- De Souza, L.; Shivakumar, S. Polyhydroxyalkanoates (PHA)—Applications in Wound Treatment and as Precursors for Oral Drugs. In Biotechnological Applications of Polyhydroxyalkanoates; Kalia, V., Ed.; Springer: Singapore, 2019. [Google Scholar]
- Asran, A.S.; Razghandi, K.; Aggarwal, N.; Michler, G.H.; Groth, T. Nanofibers from Blends of Polyvinyl Alcohol and Polyhydroxy Butyrate As Potential Scaffold Material for Tissue Engineering of Skin. Biomacromolecules 2010, 11, 3413–3421. [Google Scholar] [CrossRef] [PubMed]
- Gumel, A.M.; Razaif-Mazinah, M.R.M.; Anis, S.N.S.; Annuar, M.S.M. Poly (3-hydroxyalkanoates)-co-(6-hydroxyhexanoate) hydrogel promotes angiogenesis and collagen deposition during cutaneous wound healing in rats. Biomed. Mater. 2015, 10, 045001. [Google Scholar] [CrossRef] [PubMed]
- Li, X.-T.; Zhang, Y.; Chen, G.-Q. Nanofibrous polyhydroxyalkanoate matrices as cell growth supporting materials. Biomaterials 2008, 29, 3720–3728. [Google Scholar] [CrossRef]
- Lim, J.; You, M.L.; Li, J.; Li, Z.B. Emerging bone tissue engineering via Polyhydroxyalkanoate (PHA)-based scaffolds. Mater. Sci. Eng. C Mater. Biol. Appl. 2017, 79, 917–929. [Google Scholar] [CrossRef] [PubMed]
- Galego, N.; Rozsa, C.; Sánchez, R.; Fung, J.; Analía, V.; Santo-Tomás, J. Characterization and application of poly(β-hydroxyalkanoates) family as composite biomaterials. Polym. Test. 2000, 19, 485–492. [Google Scholar] [CrossRef]
- Zhao, K.; Deng, Y.; Chen, J.C.; Chen, G.Q. Polyhydroxyalkanoate (PHA) scaffolds with good mechanical properties and biocompatibility. Biomaterials 2003, 24, 1041–1045. [Google Scholar] [CrossRef]
- Bretcanu, O.; Chen, Q.; Misra, S.K.; Boccaccini, A.R.; Roy, I.; Verne, E.; Brovarone, C.V. Biodegradable polymer coated 45S5 Bioglassderived glass-ceramic scaffolds for bone tissue engineering. Glass Technol. Eur. J. Glass Sci. Technol. Part A 2007, 48, 227–234. [Google Scholar]
- Francis, L.; Meng, D.; Knowles, J.C.; Roy, I.; Boccaccini, A.R. Multi-functional P (3HB) microsphere/45S5 Bioglass®-based composite scaffolds for bone tissue engineering. Acta Biomater. 2010, 6, 2773–2786. [Google Scholar] [CrossRef]
- Mouriño, V.; Cattalini, J.P.; Roether, J.A.; Dubey, P.; Roy, I.; Boccaccini, A.R. Composite polymer-bioceramic scaffolds with drug delivery capability for bone tissue engineering. Expert Opin. Drug Deliv. 2013, 10, 1353–1365. [Google Scholar] [CrossRef]
- Sodian, R.; Sperling, J.S.; Martin, D.P.; Egozy, A.; Stock, U.; Mayer, J.E.; Vacanti, J.P. Fabrication of a trileaflet heart valve scaffold from a polyhydroxyalkanoate biopolyester for use in tissue engineering. Tissue Eng. 2000, 6, 183–188. [Google Scholar] [CrossRef] [PubMed]
- Cheng, S.T.; Chen, Z.F.; Chen, G.Q. The expression of cross-linked elastin by rabbit blood vessel smooth muscle cells cultured in polyhydroxyalkanoate scaffolds. Biomaterials 2008, 29, 4187–4194. [Google Scholar] [CrossRef] [PubMed]
- Rathbone, S.; Furrer, P.; Lubben, J.; Zinn, M.; Cartmell, S. Biocompatibility of polyhydroxyalkanoate as a potential material for ligament and tendon scaffold material. J. Biomed. Mater. Res. Part. A 2010, 93a, 1391–1403. [Google Scholar] [CrossRef] [PubMed]
- Lizarraga-Valderrama, L.R.; Taylor, C.S.; Aeyssens, F.C.; Haycock, J.W.; Knowles, J.C.; Roy, I. Unidirectional neuronal cell growth and differentiation on aligned polyhydroxyalkanoate blend microfibres with varying diameters. J. Tissue Eng. Regen Med. 2019, 13, 1581–1594. [Google Scholar] [CrossRef] [PubMed]
- Bagdadi, A.V.; Safari, M.; Dubey, P.; Basnett, P.; Sofokleous, P.; Humphrey, E.; Locke, I.; Edirisinghe, M.; Terracciano, C.; Boccaccini, A.R.; et al. Poly(3-hydroxyoctanoate), a promising new material for cardiac tissue engineering. J. Tissue Eng. Regen Med. 2018, 12, e495–e512. [Google Scholar] [CrossRef]
- Lizarraga-Valderrama, L.R.; Nigmatullin, R.; Taylor, C.; Haycock, J.W.; Claeyssens, F.; Knowles, J.C.; Roy, I. Nerve tissue engineering using blends of poly(3-hydroxyalkanoates) for peripheral nerve regeneration. Eng. Life Sci. 2015, 15, 612–621. [Google Scholar] [CrossRef] [Green Version]
- Francis, L.; Meng, D.; Locke, I.C.; Knowles, J.C.; Mordan, N.; Salih, V.; Boccaccini, A.R.; Roy, I. Novel poly(3-hydroxybutyrate) composite films containing bioactive glass nanoparticles for wound healing applications. Polym. Int. 2016, 65, 661–674. [Google Scholar] [CrossRef] [Green Version]
- Gao, S.; Tang, G.; Hua, D.; Xiong, R.; Han, J.; Jiang, S.; Zhang, Q.; Huang, C. Stimuli-responsive bio-based polymeric systems and their applications. J. Mat. Chem. B 2019, 7, 709–729. [Google Scholar] [CrossRef]
- Nigmatullin, R.; Thomas, P.; Lukasiewicz, B.; Puthussery, H.; Roy, I. Polyhydroxyalkanoates, a family of natural polymers, and their applications in drug delivery. J. Chem. Technol. Biotechnol. 2015, 90, 1209–1221. [Google Scholar] [CrossRef]
- Di Mascolo, D.; Basnett, P.; Palange, A.L.; Francardi, M.; Roy, I.; Decuzzi, P. Tuning core hydrophobicity of spherical polymeric nanoconstructs for docetaxel delivery. Polym. Int. 2016, 65, 741–746. [Google Scholar] [CrossRef]
- Shishatskaya, E.; Goreva, A.; Voinova, O.; Inzhevatkin, E.; Khlebopros, R.; Volova, T. Evaluation of antitumor activity of rubomycin deposited in absorbable polymeric microparticles. Bull. Exp. Biol. Med. 2008, 145, 358–361. [Google Scholar] [CrossRef]
- Masood, F.; Chen, P.; Yasin, T.; Fatima, N.; Hasan, F.; Hameed, A. Encapsulation of Ellipticine in poly-(3-hydroxybutyrate-co-3-hydroxyvalerate) based nanoparticles and its in vitro application. Mater. Sci. Eng. C Mater. Biol. Appl. 2013, 33, 1054–1060. [Google Scholar] [CrossRef]
- Loh, X.J.; Ong, S.J.; Tung, Y.T.; Choo, H.T. Dual responsive micelles based on poly (R)-3-hydroxybutyrate and poly(2-(di-methylamino)ethyl methacrylate) for effective doxorubicin delivery. Polym. Chem. 2013, 4, 2564–2574. [Google Scholar] [CrossRef]
- Xiao, L.; Wang, B.; Yang, G.; Gauthier, M. Poly(Lactic Acid)-Based Biomaterials: Synthesis, Modification and Applications. Biomed. Sci. Eng. Technol. 2012. [Google Scholar] [CrossRef] [Green Version]
- Msuya, N.; Katima, J.H.; Masanja, E.; Temu, A.K. Poly (lactic acid) Production from Monomer to Polymer: A Review. Scifed J. Polym. Sci. 2017, 1, 1–15. [Google Scholar]
- Inkinen, S.; Hakkarainen, M.; Albertsson, A.C.; Sodergard, A. From lactic acid to poly(lactic acid) (PLA): Characterization and analysis of PLA and its precursors. Biomacromolecules 2011, 12, 523–532. [Google Scholar] [CrossRef]
- Singhvi, M.S.; Zinjarde, S.S.; Gokhale, D.V. Polylactic acid: Synthesis and biomedical applications. J. Appl. Microbiol. 2019, 127, 1612–1626. [Google Scholar] [CrossRef] [Green Version]
- Jung, Y.K.; Kim, T.Y.; Park, S.J.; Lee, S.Y. Metabolic engineering of Escherichia coli for the production of polylactic acid and its copolymers. Biotechnol. Bioeng. 2010, 105, 161–171. [Google Scholar] [CrossRef]
- Riaz, S.; Fatima, N.; Rasheed, A.; Riaz, M.; Anwar, F.; Khatoon, Y. Metabolic Engineered Biocatalyst: A Solution for PLA Based Problems. Int. J. Biomater. 2018, 2018, 1963024. [Google Scholar] [CrossRef]
- Jung, Y.K.; Lee, S.Y. Efficient production of polylactic acid and its copolymers by metabolically engineered Escherichia coli. J. Biotechnol. 2011, 151, 94–101. [Google Scholar] [CrossRef] [PubMed]
- Elsawy, M.A.; Kim, K.H.; Park, J.W.; Deep, A. Hydrolytic degradation of polylactic acid (PLA) and its composites. Renew. Sustain. Energy Rev. 2017, 79, 1346–1352. [Google Scholar] [CrossRef]
- Pina, S.; Ferreira, J.M.F. Bioresorbable Plates and Screws for Clinical Applications: A Review. J. Healthc. Eng. 2012, 3, 243–260. [Google Scholar] [CrossRef] [Green Version]
- Liu, S.; Qin, S.; He, M.; Zhou, D.; Qin, Q.; Wang, H. Current applications of poly(lactic acid) composites in tissue engineering and drug delivery. Compos. Part B Eng. 2020, 199, 108238. [Google Scholar] [CrossRef]
- Prokop, A.; Jubel, A.; Helling, H.J.; Eibach, T.; Peters, C.; Baldus, S.E.; Rehm, K.E. Soft tissue reactions of different biodegradable polylactide implants. Biomaterials 2004, 25, 259–267. [Google Scholar] [CrossRef]
- Majola, A.; Vainionpaa, S.; Vihtonen, K.; Mero, M.; Vasenius, J.; Tormala, P.; Rokkanen, P. Absorption, biocompatibility, and fixation properties of polylactic acid in bone tissue: An experimental study in rats. Clin. Orthop. Relat. Res. 1991, 268, 260–269. [Google Scholar]
- Shikinami, Y.; Matsusue, Y.; Nakamura, T. The complete process of bioresorption and bone replacement using devices made of forged composites of raw hydroxyapatite particles/poly l-lactide (F-u-HA/PLLA). Biomaterials 2005, 26, 5542–5551. [Google Scholar] [CrossRef]
- Hochuli-Vieira, E.; Cabrini-Gabrielli, M.A.; Pereira-Filho, V.A.; Gabrielli, M.F.; Padilha, J.G. Rigid internal fixation with titanium versus bioresorbable miniplates in the repair of mandibular fractures in rabbits. Int. J. Oral Maxillofac. Surg. 2005, 34, 167–173. [Google Scholar] [CrossRef] [PubMed]
- Lassalle, V.; Ferreira, M.L. PLA Nano- and Microparticles for Drug Delivery: An Overview of the Methods of Preparation. Macromol. Biosci. 2007, 7, 767–783. [Google Scholar] [CrossRef] [PubMed]
- Zeng, X.; Tao, W.; Liu, G.; Mei, L. Polydopamine-based surface modification of copolymeric nanoparticles as a targeted drug delivery system for cancer therapy. J. Control. Release 2017, 259, e150–e151. [Google Scholar] [CrossRef]
- Mi, F.-L.; Shyu, S.-S.; Lin, Y.-M.; Wu, Y.-B.; Peng, C.-K.; Tsai, Y.-H. Chitin/PLGA blend microspheres as a biodegradable drug delivery system: A new delivery system for protein. Biomaterials 2003, 24, 5023–5036. [Google Scholar] [CrossRef]
- Hu, K.; Li, J.; Shen, Y.; Lu, W.; Gao, X.; Zhang, Q.; Jiang, X. Lactoferrin-conjugated PEG–PLA nanoparticles with improved brain delivery: In vitro and in vivo evaluations. J. Control. Release 2009, 134, 55–61. [Google Scholar] [CrossRef] [PubMed]
- Xia, H.; Gao, X.; Gu, G.; Liu, Z.; Hu, Q.; Tu, Y.; Song, Q.; Yao, L.; Pang, Z.; Jiang, X.; et al. Penetratin-functionalized PEG–PLA nanoparticles for brain drug delivery. Int. J. Pharm. 2012, 436, 840–850. [Google Scholar] [CrossRef]
- Chuaponpat, N.; Ueda, T.; Ishigami, A.; Kurose, T.; Ito, H. Morphology, Thermal and Mechanical Properties of Co-Continuous Porous Structure of PLA/PVA Blends by Phase Separation. Polymers 2020, 12, 1083. [Google Scholar] [CrossRef]
- Buzarovska, A.; Dinescu, S.; Chitoiu, L.; Costache, M. Porous poly (L-lactic acid) nanocomposite scaffolds with functionalized TiO 2 nanoparticles: Properties, cytocompatibility and drug release capability. J. Mater. Sci. 2018, 53, 11151–11166. [Google Scholar] [CrossRef]
- Drechsel, E. Anleitung zur Darstellung Physiologisch Chemischer Präparate; Bergmann: Wiesbaden, Germany, 1889. [Google Scholar]
- Shima, S.; Sakai, H. Polylysine produced by Streptomyces. Agric. Biol. Chem. 1977, 41, 1807–1809. [Google Scholar] [CrossRef] [Green Version]
- Singh, M.; Rao, D.M.; Pande, S.; Battu, S.; Dutt, K.R.; Ramesh, M. Medicinal uses of L-lysine: Past and future. Int. J. Res. Pharm. Sci. 2011, 2, 637–642. [Google Scholar]
- Rubia, L.B.; Gomez, R. TLC sensitivity of six modifications of Dragendorff’s reagent. J. Pharm. Sci. 1977, 66, 1656–1657. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Ren, X.; Yu, C.; Wang, J.; Wang, L.; Zhuge, X.; Liu, X. Physiological and Transcriptional Responses of Streptomyces albulus to Acid Stress in the Biosynthesis of ε-Poly-L-lysine. Front. Microbiol. 2020, 11, 1379. [Google Scholar] [CrossRef]
- Hancock, R.E. Peptide antibiotics. Lancet 1997, 349, 418–422. [Google Scholar] [CrossRef]
- Bradshaw, J.P. Cationic antimicrobial peptides. BioDrugs 2003, 17, 233–240. [Google Scholar] [CrossRef] [PubMed]
- Hyldgaard, M.; Mygind, T.; Vad, B.S.; Stenvang, M.; Otzen, D.E.; Meyer, R.L. The antimicrobial mechanism of action of epsilon-poly-l-lysine. Appl. Environ. Microb. 2014, 80, 7758–7770. [Google Scholar] [CrossRef] [Green Version]
- Xu, M.; Song, Q.; Gao, L.; Liu, H.; Feng, W.; Huo, J.; Jin, H.; Huang, L.; Chai, J.; Pei, Y. Single-step fabrication of catechol-ε-poly-L-lysine antimicrobial paint that prevents superbug infection and promotes osteoconductivity of titanium implants. Chem. Eng. J. 2020, 125240. [Google Scholar] [CrossRef]
- Wang, R.; Li, Q.; Chi, B.; Wang, X.; Xu, Z.; Xu, Z.; Chen, S.; Xu, H. Enzyme-induced dual-network ε-poly-L-lysine-based hydrogels with robust self-healing and antibacterial performance. Chem. Commun. 2017, 53, 4803–4806. [Google Scholar] [CrossRef] [PubMed]
- Zou, Y.-J.; He, S.-S.; Du, J.-Z. ε-Poly (L-lysine)-based Hydrogels with Fast-acting and Prolonged Antibacterial Activities. Chin. J. Polym. Sci. 2018, 36, 1239–1250. [Google Scholar] [CrossRef]
- Yang, X.; Wang, B.; Sha, D.; Liu, Y.; Xu, J.; Shi, K.; Yu, C.; Ji, X. Injectable and antibacterial ε-poly (l-lysine)-modified poly (vinyl alcohol)/chitosan/AgNPs hydrogels as wound healing dressings. Polymer 2020, 212, 123155. [Google Scholar] [CrossRef]
- Karimi, M.; Yazdi, F.T.; Mortazavi, S.A.; Shahabi-Ghahfarrokhi, I.; Chamani, J. Development of active antimicrobial poly (l-glutamic) acid-poly (l-lysine) packaging material to protect probiotic bacterium. Polym. Test. 2020, 83, 106338. [Google Scholar] [CrossRef]
- Rapp, M.V.; Maier, G.P.; Dobbs, H.A.; Higdon, N.J.; Waite, J.H.; Butler, A.; Israelachvili, J.N. Defining the catechol–cation synergy for enhanced wet adhesion to mineral surfaces. J. Am. Chem. Soc. 2016, 138, 9013–9016. [Google Scholar] [CrossRef] [Green Version]
- Wang, R.; Li, J.; Chen, W.; Xu, T.; Yun, S.; Xu, Z.; Xu, Z.; Sato, T.; Chi, B.; Xu, H. A biomimetic mussel-inspired ε-poly-l-lysine hydrogel with robust tissue-anchor and anti-infection capacity. Adv. Funct. Mater. 2017, 27, 1604894. [Google Scholar] [CrossRef]
- Li, S.; Chen, N.; Li, Y.; Li, X.; Zhan, Q.; Ban, J.; Zhao, J.; Hou, X.; Yuan, X. Metal-crosslinked ɛ-poly-L-lysine tissue adhesives with high adhesive performance: Inspiration from mussel adhesive environment. Int. J. Biol. Macromol. 2020, 153, 1251–1261. [Google Scholar] [CrossRef]
- Liu, S.; Liu, X.; Ren, Y.; Wang, P.H.; Pu, Y.; Yang, R.; Wang, X.; Tan, X.Y.; Ye, Z.; Maurizot, V. Mussel-inspired Dual-crosslinking Hyaluronic Acid/ε-polylysine Hydrogel with Self-healing and Antibacterial Properties for Wound Healing. ACS Appl. Mater. Interfaces 2020, 12, 27876–27888. [Google Scholar] [CrossRef]
- De Smedt, S.C.; Demeester, J.; Hennink, W.E. Cationic polymer based gene delivery systems. Pharm. Res. 2000, 17, 113–126. [Google Scholar] [CrossRef]
- Deng, J.; Gao, N.; Wang, Y.; Yi, H.; Fang, S.; Ma, Y.; Cai, L. Self-assembled cationic micelles based on PEG-PLL-PLLeu hybrid polypeptides as highly effective gene vectors. Biomacromolecules 2012, 13, 3795–3804. [Google Scholar] [CrossRef]
- Sun, Z.; Song, C.; Wang, C.; Hu, Y.; Wu, J. Hydrogel-based controlled drug delivery for cancer treatment: A review. Mol. Pharm. 2019, 17, 373–391. [Google Scholar] [CrossRef]
- Guo, Z.; Sui, J.; Ma, M.; Hu, J.; Sun, Y.; Yang, L.; Fan, Y.; Zhang, X. pH-Responsive charge switchable PEGylated ε-poly-l-lysine polymeric nanoparticles-assisted combination therapy for improving breast cancer treatment. J. Control. Release 2020, 326, 350–364. [Google Scholar] [CrossRef] [PubMed]
- El Assal, R.; Abou-Elkacem, L.; Tocchio, A.; Pasley, S.; Matosevic, S.; Kaplan, D.L.; Zylberberg, C.; Demirci, U. Bioinspired preservation of natural killer cells for cancer immunotherapy. Adv. Sci. 2019, 6, 1802045. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tarantino, L.M. Agency Response Letter GRAS Notice No. GRN 000135. Available online: https://www.researchgate.net/publication/237593613_Antimicrobial_Activity_of_e-Polylysine_in_Various_Food_Extracts (accessed on 11 January 2021).
- Hiraki, J.; Ichikawa, T.; Ninomiya, S.-I.; Seki, H.; Uohama, K.; Seki, H.; Kimura, S.; Yanagimoto, Y.; Barnett, J.W., Jr. Use of ADME studies to confirm the safety of ε-polylysine as a preservative in food. Regul. Toxicol. Pharmacol. 2003, 37, 328–340. [Google Scholar] [CrossRef]
- Hiraki, J.; Suzuki, E. Process for Producing ε-poly-L-lysine with Immobilized Streptomyces Albulus. U.S. Patent 5,900,363, 4 May 1999. [Google Scholar]
- Zhang, Y.-X.; Perry, K.; Vinci, V.A.; Powell, K.; Stemmer, W.P.; del Cardayré, S.B. Genome shuffling leads to rapid phenotypic improvement in bacteria. Nature 2002, 415, 644–646. [Google Scholar] [CrossRef]
- Li, S.; Li, F.; Chen, X.-S.; Wang, L.; Xu, J.; Tang, L.; Mao, Z.-G. Genome shuffling enhanced ε-poly-l-lysine production by improving glucose tolerance of Streptomyces graminearus. Appl. Biochem. Biotech. 2012, 166, 414–423. [Google Scholar] [CrossRef]
- Li, S.; Ji, J.; Hu, S.; Chen, G. Enhancement of ε-poly-L-lysine production in Streptomyces griseofuscus by addition of exogenous astaxanthin. Bioproc. Biosyst. Eng. 2020, 43, 1813–1821. [Google Scholar] [CrossRef]
- Yamanaka, K.; Maruyama, C.; Takagi, H.; Hamano, Y. ε-Poly-L-lysine dispersity is controlled by a highly unusual nonribosomal peptide synthetase. Nat. Chem. Biol. 2008, 4, 766–772. [Google Scholar] [CrossRef]
- Samadlouie, H.R.; Gharanjik, S.; Tabatabaie, Z.B. Optimization of the Production of ε-Poly-L-Lysine by Novel Producer Lactic Acid Bacteria Isolated from Traditional Dairy Products. Biomed. Res. Int 2020, 2020. [Google Scholar] [CrossRef] [PubMed]
- Kimura, K.; Fujimoto, Z. Enzymatic degradation of poly-gamma-glutamic acid. In Amino-Acid Homopolymers Occurring in Nature; Springer: Berlin, Germany, 2010; pp. 95–117. [Google Scholar]
- Moghaddam, B. Stress activation of glutamate neurotransmission in the prefrontal cortex: Implications for dopamine-associated psychiatric disorders. Biol. Psychiatry 2002, 51, 775–787. [Google Scholar] [CrossRef]
- Peng, L.; Hertz, L.; Huang, R.; Sonnewald, U.; Petersen, S.B.; Westergaard, N.; Larsson, O.; Schousboe, A. Utilization of glutamine and of TCA cycle constituents as precursors for transmitter glutamate and GABA. Dev. Neurosci. 1993, 15, 367–377. [Google Scholar] [CrossRef]
- Shih, L.; Van, Y.-T. The production of poly-(γ-glutamic acid) from microorganisms and its various applications. Bioresour. Technol. 2001, 79, 207–225. [Google Scholar] [CrossRef]
- Kawashima, S.; Kanehisa, M. AAindex: Amino acid index database. Nucleic Acids Res. 2000, 28, 374. [Google Scholar] [CrossRef]
- Tanaka, T.; Yaguchi, T.; Hiruta, O.; Futamura, T.; Uotani, K.; Satoh, A.; Taniguchi, M.; Susumu, O. Screening for microorganisms having poly (γ-glutamic acid) endohydrolase activity and the enzyme production by Myrothecium sp. TM-4222. Biosci. Biotechnol. Biochem. 1993, 57, 1809–1810. [Google Scholar] [CrossRef]
- Liao, Z.-X.; Peng, S.-F.; Ho, Y.-C.; Mi, F.-L.; Maiti, B.; Sung, H.-W. Mechanistic study of transfection of chitosan/DNA complexes coated by anionic poly (γ-glutamic acid). Biomaterials 2012, 33, 3306–3315. [Google Scholar] [CrossRef] [PubMed]
- Su, C.-Y.; Tseng, C.-L.; Wu, S.-H.; Shih, B.-W.; Chen, Y.-Z.; Fang, H.-W. Poly-gamma-glutamic acid functions as an effective lubricant with antimicrobial activity in multipurpose contact lens care solutions. Polymers 2019, 11, 1050. [Google Scholar] [CrossRef] [Green Version]
- Su, C.-Y.; Chen, C.-C.; Chen, H.-Y.; Lin, C.-P.; Lin, F.-H.; Fang, H.-W. Characteristics of an alternative antibacterial biomaterial for mouthwash in the absence of alcohol. J. Dent. Sci. 2019, 14, 192–197. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Song, L.; Zhang, X.; Zhou, R.; Yin, J.; Luan, S. Poly (γ-glutamic acid)-based electrospun nanofibrous mats with photodynamic therapy for effectively combating wound infection. Mater. Sci. Eng. C 2020, 113, 110936. [Google Scholar] [CrossRef] [PubMed]
- Bae, S.-R.; Park, C.; Choi, J.-C.; Poo, H.; Kim, C.-J.; Sung, M.-H. Effects of ultra high molecular weight poly-gamma-glutamic acid from Bacillus subtilis (chungkookjang) on corneal wound healing. J. Microbiol. Biotechnol. 2010, 20, 803–808. [Google Scholar] [PubMed]
- Choi, J.-C.; Uyama, H.; Lee, C.-H.; Sung, M.-H. Promotion effects of ultra-high molecular weight poly-gamma-glutamic acid on wound healing. J. Microbiol. Biotechnol. 2015, 25, 941–945. [Google Scholar] [CrossRef] [PubMed]
- Dissemond, J.; Goos, M.; Wagner, S. The role of oxidative stress in the pathogenesis and therapy of chronic wounds. Hautarzt Z. Dermatol. Venerol. Verwandte Geb. 2002, 53, 718–723. [Google Scholar] [CrossRef] [PubMed]
- Park, S.-J.; Uyama, H.; Kwak, M.-S.; Sung, M.-H. Comparison of the Stability of Poly-γ-Glutamate Hydrogels Prepared by UV and γ-Ray Irradiation. J. Microbiol. Biotechnol. 2019, 29, 1078–1082. [Google Scholar] [CrossRef] [PubMed]
- Hua, J.; Li, Z.; Xia, W.; Yang, N.; Gong, J.; Zhang, J.; Qiao, C. Preparation and properties of EDC/NHS mediated crosslinking poly (gamma-glutamic acid)/epsilon-polylysine hydrogels. Mater. Sci. Eng. C 2016, 61, 879–892. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Ma, Y.; Pan, X.; Chen, S.; Zhuang, H.; Wang, S. A composite hydrogel of chitosan/heparin/poly (γ-glutamic acid) loaded with superoxide dismutase for wound healing. Carbohydr. Polym. 2018, 180, 168–174. [Google Scholar] [CrossRef] [PubMed]
- Stevanović, M.; Bračko, I.; Milenković, M.; Filipović, N.; Nunić, J.; Filipič, M.; Uskoković, D.P. Multifunctional PLGA particles containing poly (l-glutamic acid)-capped silver nanoparticles and ascorbic acid with simultaneous antioxidative and prolonged antimicrobial activity. Acta Biomater. 2014, 10, 151–162. [Google Scholar] [CrossRef]
- Pisani, S.; Dorati, R.; Scocozza, F.; Mariotti, C.; Chiesa, E.; Bruni, G.; Genta, I.; Auricchio, F.; Conti, M.; Conti, B. Preliminary investigation on a new natural based poly (gamma-glutamic acid)/Chitosan bioink. J. Biomed. Mater. Res. Part B Appl. Biomater. 2020, 108, 2718–2732. [Google Scholar] [CrossRef]
- Upadhyay, K.K.; Bhatt, A.N.; Mishra, A.K.; Dwarakanath, B.S.; Jain, S.; Schatz, C.; le Meins, J.-F.; Farooque, A.; Chandraiah, G.; Jain, A.K. The intracellular drug delivery and anti tumor activity of doxorubicin loaded poly (γ-benzyl l-glutamate)-b-hyaluronan polymersomes. Biomaterials 2010, 31, 2882–2892. [Google Scholar] [CrossRef]
- Liao, Z.-X.; Peng, S.-F.; Chiu, Y.-L.; Hsiao, C.-W.; Liu, H.-Y.; Lim, W.-H.; Lu, H.-M.; Sung, H.-W. Enhancement of efficiency of chitosan-based complexes for gene transfection with poly (γ-glutamic acid) by augmenting their cellular uptake and intracellular unpackage. J. Control. Release 2014, 193, 304–315. [Google Scholar] [CrossRef]
- Ivanovics, G.; Erdos, L. Ein beitrag zum wesen der kapselsubstanz des milzbrandbazillus. Z. Immun. 1937, 90, 5–19. [Google Scholar]
- Jang, J.; Cho, M.; Chun, J.-H.; Cho, M.-H.; Park, J.; Oh, H.-B.; Yoo, C.-K.; Rhie, G.-E. The poly-γ-D-glutamic acid capsule of Bacillus anthracis enhances lethal toxin activity. Infect. Immun. 2011, 79, 3846–3854. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Candela, T.; Fouet, A. Poly-gamma-glutamate in bacteria. Mol. Microbiol. 2006, 60, 1091–1098. [Google Scholar] [CrossRef] [PubMed]
- Shurtleff, W.; Aoyagi, A. History of Natto and Its Relatives (1405–2012); Soyinfo Center: Lafayette, CA, USA, 2012. [Google Scholar]
- Luo, Z.; Guo, Y.; Liu, J.; Qiu, H.; Zhao, M.; Zou, W.; Li, S. Microbial synthesis of poly-γ-glutamic acid: Current progress, challenges, and future perspectives. Biotechnol. Biofuels 2016, 9, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Steinkraus, K. Industrialization of Indigenous Fermented Foods, Revised and Expanded; CRC Press: Boca Raton, FL, USA, 2004. [Google Scholar]
- Feng, J.; Gu, Y.; Quan, Y.; Cao, M.; Gao, W.; Zhang, W.; Wang, S.; Yang, C.; Song, C. Improved poly-γ-glutamic acid production in Bacillus amyloliquefaciens by modular pathway engineering. Metab. Eng. 2015, 32, 106–115. [Google Scholar] [CrossRef]
- Cai, D.; He, P.; Lu, X.; Zhu, C.; Zhu, J.; Zhan, Y.; Wang, Q.; Wen, Z.; Chen, S. A novel approach to improve poly-γ-glutamic acid production by NADPH regeneration in Bacillus licheniformis WX-02. Sci. Rep. 2017, 7, 43404. [Google Scholar] [CrossRef]
- Niemeyer, R. Cyclic Condensed Metaphosphates in Plants and the Possible Correlations between Inorganic Polyphosphates and Other Compounds. In Inorganic Polyphosphates: Biochemistry, Biology, Biotechnology; Schröder, H.C., Müller, W.E.G., Eds.; Springer: Berlin/Heidelberg, Germany, 1999; pp. 83–100. [Google Scholar] [CrossRef]
- Achbergerova, L.; Nahalka, J. Polyphosphate—An ancient energy source and active metabolic regulator. Microb. Cell Fact. 2011, 10, 63. [Google Scholar] [CrossRef] [Green Version]
- Ahn, K.H.; Kornberg, A. Polyphosphate Kinase from Escherichia-Coli—Purification and Demonstration of a Phosphoenzyme Intermediate. J. Biol. Chem. 1990, 265, 11734–11739. [Google Scholar] [CrossRef]
- Kornberg, A. Inorganic Polyphosphate—Toward Making a Forgotten Polymer Unforgettable. J. Bacteriol. 1995, 177, 491–496. [Google Scholar] [CrossRef] [Green Version]
- Tewari, K.K.; Singh, M. Acid Soluble and Acid Insoluble Inorganic Polyphosphates in Cuscuta-Reflexa. Phytochemistry 1964, 3, 341–347. [Google Scholar] [CrossRef]
- Christ, J.J.; Blank, L.M. Analytical polyphosphate extraction from Saccharomyces cerevisiae. Anal. Biochem. 2018, 563, 71–78. [Google Scholar] [CrossRef]
- Kulaev, I.S. Biochemistry of Inorganic Polyphosphates. Rev. Physiol. Biochem. Pharmacol. 1975, 73, 131–158. [Google Scholar] [CrossRef]
- Mandala, V.S.; Loh, D.M.; Shepard, S.M.; Geeson, M.B.; Sergeyev, I.V.; Nocera, D.G.; Cummins, C.C.; Hong, M. Bacterial Phosphate Granules Contain Cyclic Polyphosphates: Evidence from 31P Solid-State NMR. J. Am. Chem. Soc. 2020, 142, 18407–18421. [Google Scholar] [CrossRef]
- Chaubal, M.V.; Sen-Gupta, A.; Lopina, S.T.; Bruley, D.F. Polyphosphates and other phosphorus-containing polymers for drug delivery applications. Crit. Rev. Drug 2003, 20, 295–315. [Google Scholar] [CrossRef]
- Liu, J.Y.; Huang, W.; Pang, Y.; Yan, D.Y. Hyperbranched polyphosphates: Synthesis, functionalization and biomedical applications. Chem. Soc. Rev. 2015, 44, 3942–3953. [Google Scholar] [CrossRef]
- Liu, J.Y.; Huang, W.; Pang, Y.; Zhu, X.Y.; Zhou, Y.F.; Yan, D.Y. Hyperbranched Polyphosphates for Drug Delivery Application: Design, Synthesis, and In Vitro Evaluation. Biomacromolecules 2010, 11, 1564–1570. [Google Scholar] [CrossRef]
- Vasiliadis, G.; Duncan, A.; Bayly, R.C.; May, J.W. Polyphosphate Production by Strains of Acinetobacter. FEMS Microbiol. Lett. 1990, 70, 37–40. [Google Scholar] [CrossRef]
- Liang, M.; Frank, S.; Luensdorf, H.; Warren, M.J.; Prentice, M.B. Bacterial microcompartment-directed polyphosphate kinase promotes stable polyphosphate accumulation in E. coli. Biotechnol. J. 2017, 12. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.Y.; Ishige, K.; Kornberg, A. A polyphosphate kinase (PPK2) widely conserved in bacteria. Proc. Natl. Acad. Sci. USA 2002, 99, 16678–16683. [Google Scholar] [CrossRef] [Green Version]
- Kuroda, A.; Kornberg, A. Polyphosphate kinase as a nucleoside diphosphate kinase in Escherichia coli and Pseudomonas aeruginosa. Proc. Natl. Acad. Sci. USA 1997, 94, 439–442. [Google Scholar] [CrossRef] [Green Version]
- Xie, L.H.; Jakob, U. Inorganic polyphosphate, a multifunctional polyanionic protein scaffold. J. Biol. Chem. 2019, 294, 2180–2190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qiu, G.L.; Zuniga-Montanez, R.; Law, Y.Y.; Thi, S.S.; Nguyen, T.Q.N.; Eganathan, K.; Liu, X.H.; Nielsen, P.H.; Williams, R.B.H.; Wuertz, S. Polyphosphate-accumulating organisms in full-scale tropical wastewater treatment plants use diverse carbon sources. Water Res. 2019, 149, 496–510. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Wang, X.M.; Hui, K.M.; Wei, W.; Zhang, W.; Miao, A.J.; Xiao, L.; Yang, L.Y. Highly Effective Polyphosphate Synthesis, Phosphate Removal, and Concentration Using Engineered Environmental Bacteria Based on a Simple Solo Medium-Copy Plasmid Strategy. Environ. Sci. Technol. 2018, 52, 214–222. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.H.; Schroder, H.C.; Muller, W.E.G. Polyphosphate as a metabolic fuel in Metazoa: A foundational breakthrough invention for biomedical applications. Biotechnol. J. 2016, 11, 11–30. [Google Scholar] [CrossRef] [PubMed]
- Kulakovskaya, T.V.; Vagabov, V.M.; Kulaev, I.S. Inorganic polyphosphate in industry, agriculture and medicine: Modern state and outlook. Process. Biochem. 2012, 47, 1–10. [Google Scholar] [CrossRef]
- Maier, S.K.; Scherer, S.; Loessner, M.J. Long-chain polyphosphate causes cell lysis and inhibits Bacillus cereus septum formation, which is dependent on divalent cations. Appl. Environ. Microb. 1999, 65, 3942–3949. [Google Scholar] [CrossRef] [Green Version]
- Jen, C.M.C.; Shelef, L.A. Factors Affecting Sensitivity of Staphylococcus-Aureus 196e to Polyphosphates. Appl. Environ. Microb. 1986, 52, 842–846. [Google Scholar] [CrossRef] [Green Version]
- Mutch, N.J. Regulation of Coagulation by Polyphosphate. Blood 2019, 134. [Google Scholar] [CrossRef]
- Smith, S.A.; Choi, S.H.; Davis-Harrison, R.; Huyck, J.; Boettcher, J.; Reinstra, C.M.; Morrissey, J.H. Polyphosphate exerts differential effects on blood clotting, depending on polymer size. Blood 2010, 116, 4353–4359. [Google Scholar] [CrossRef]
- Travers, R.J.; Smith, S.A.; Morrissey, J.H. Polyphosphate, platelets, and coagulation. Int. J. Lab. Hematol. 2015, 37, 31–35. [Google Scholar] [CrossRef] [Green Version]
- Verhoef, J.J.F.; Barendrecht, A.D.; Nickel, K.F.; Dijkxhoorn, K.; Kenne, E.; Labberton, L.; McCarty, O.J.T.; Schiffelers, R.; Heijnen, H.F.; Hendrickx, A.P.; et al. Polyphosphate nanoparticles on the platelet surface trigger contact system activation. Blood 2017, 129, 1707–1717. [Google Scholar] [CrossRef]
- Wang, Y.; Li, M.; Li, P.; Teng, H.J.; Fan, D.H.; Du, W.N.; Guo, Z.L. Progress and Applications of Polyphosphate in Bone and Cartilage Regeneration. Biomed. Res. Int. 2019, 2019, 5141204. [Google Scholar] [CrossRef] [PubMed]
- Kawazoe, Y.; Shiba, T.; Nakamura, R.; Mizuno, A.; Tsutsumi, K.; Uematsu, T.; Yamaoka, M.; Shindoh, M.; Kohgo, T. Induction of calcification in MC3T3-E1 cells by inorganic polyphosphate. J. Dent. Res. 2004, 83, 613–618. [Google Scholar] [CrossRef] [PubMed]
- Leyhausen, G.; Lorenz, B.; Zhu, H.; Geurtsen, W.; Bohnensack, R.; Muller, W.E.G.; Schroder, H.C. Inorganic polyphosphate in human osteoblast-like cells. J. Bone Min. Res. 1998, 13, 803–812. [Google Scholar] [CrossRef] [PubMed]
- Schroder, H.C.; Kurz, L.; Muller, W.E.G.; Lorenz, B. Polyphosphate in bone. Biochem. Mosc. 2000, 65, 296–303. [Google Scholar]




















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
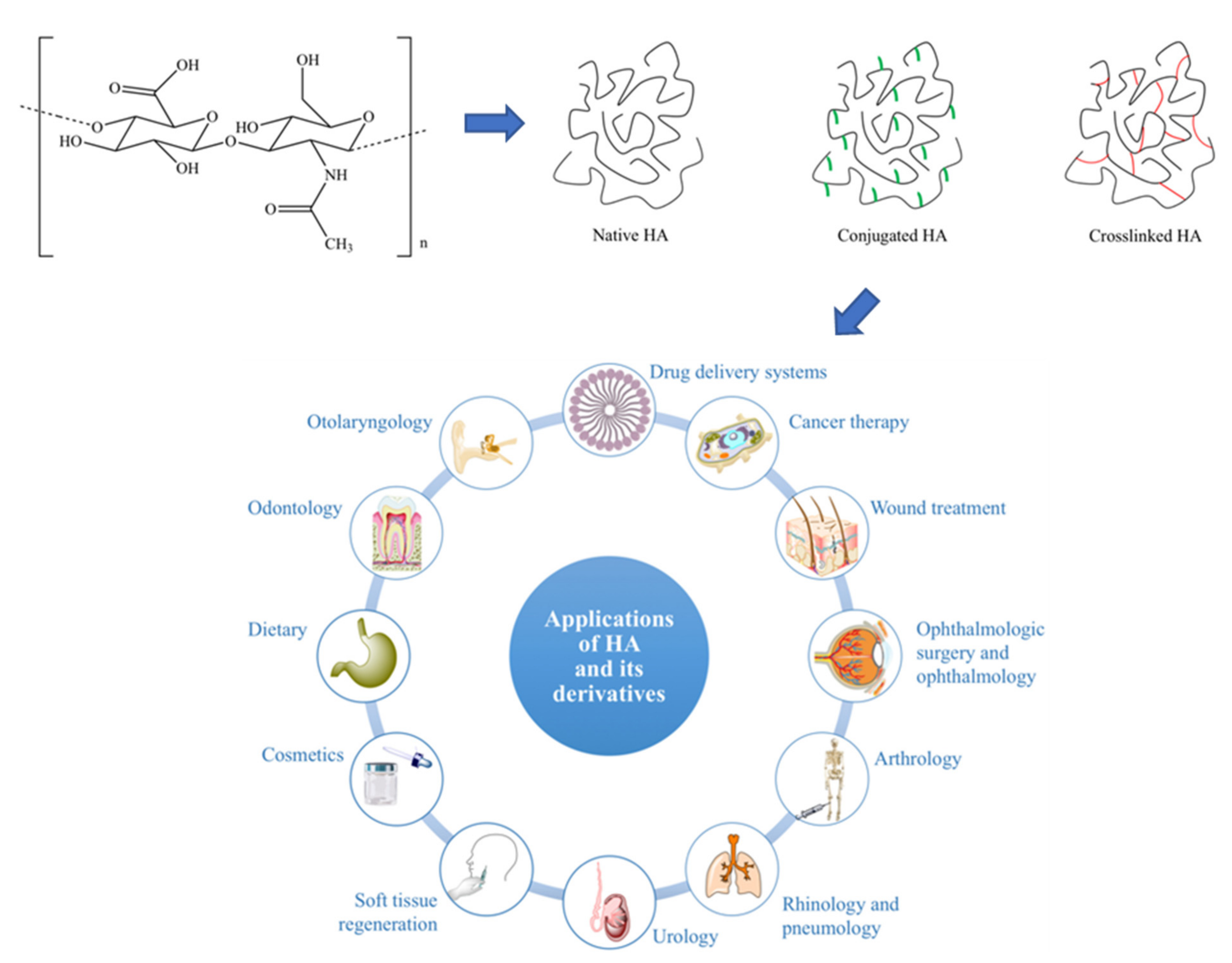{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Polymer | Polymer-Accumulating Bacteria | Biomaterial Properties in Biomedical Application | Ref. |
|---|---|---|---|
| Dextran | Leuconostoc sp., including L. pseudomesenteroides, L. mesenteroides and L. citreum Weisella cibaria, Wiesella confusa, Pediococcus pentosaceus, Lactobacillus satsumensis, and Lactobacillusplantarum | Incorporation of dextran in drug delivery systems takes advantage of its structural integrity in forming hydrogels Dextran-drug conjugates enhance their analgesic and antipyretic properties whilst reducing their constituent drug’s ulcerogenic effect and also possess anticonvulsant properties Serves as drug carrier material in targeting specific organs | [25,26,27,28,29,30,31,32,33,34,35,36,37,38,39,40,41,42,43,44,45,46,47,48,49,50,51,52,53,54,55,56,57,58,59] |
| Glycogen | Genera Streptomyces, Rhizobium, Methanococcus, Streptococcus, Enterobacter, Escherichia, Synechococcus, Micropruina and Candidatus | Tissue engineering applications, as a crosslinker for hydrogels, allow for the generation of multifunctional and self-healing biomaterials Shown to increase elongation at break of polymer structures (at the expense of tensile strength) Controlled-release drug delivery has been trialled, especially in anti-cancer therapies | [60,61,62,63,64,65,66,67,68,69,70,71,72,73,74,75,76,77,78,79,80,81,82,83,84,85,86,87,88,89,90,91,92,93,94,95,96,97,98,99,100,101,102,103,104,105,106,107,108,109,110,111,112,113,114,115] |
| Alginate | Wild-type Alginate Expressors include Pseudomonas aeruginosa and Azotobacter vinelandii Recombined into Escherichia coli | Facilitate appropriate wound moisture retention and wound healing Excellent cell-adhesive and degradation behaviour Successfully used as a minimally invasive delivery system Exceptional sustained release and swelling degree Bind with divalent cations to form crosslinks and susceptible to modifications for tissue engineering applications | [116,117,118,119,120,121,122,123,124,125,126,127,128,129,130,131,132,133,134,135,136,137,138,139,140] |
| Hyaluronic acid | First commercial production in Streptococcus zooepidemicus Genera Streptococcus and Pasteurella | Swelling ability has found use both in hydrogel tissue engineering research and in contemporary plastic surgery polymer expanding filling materials Synergistic lubricative ability has been trialled for the treatment of joint based pathology such as osteoarthritis HA is effective as an immunoisolation material, with avenues in type 1 diabetes treatment Natural affinity for some cancer surface proteins, such as CD44, promoting a drug delivery role | [141,142,143,144,145,146,147,148,149,150,151,152,153,154,155,156,157,158,159,160,161,162,163,164,165,166,167,168,169,170,171,172,173,174,175,176,177,178,179,180,181,182,183,184,185,186] |
| Gellan | Sphingomonas elodea, Sphingomonas paucimobilis and Pseudomonas elodea | Forms stable and self-supporting hydrogel and used as a culture media additive Generally used for oral formulations, as gels or coatings of capsules Protect bioactive molecules from the low pH Mostly applied in nasal, ocular, gastric pharmaceutical delivery systems, and as freeze-dried scaffold or sponges in tissue regeneration | [187,188,189,190,191,192,193,194,195,196,197,198,199,200,201,202,203,204,205] |
| Xanthan | Primarily expressed in Xanthamonas sp., Xantamonas campestris | Resist enzymatic digestion in the stomach or small intestine encouraging in colon and stomach delivery systems Improved drug permeation and bioavailability with nasal gels Excellent biomimicking potential Potential biomolecules and therapeutic carriers because of their stability, protection, and controlled release kinetics | [206,207,208,209,210,211,212,213,214,215,216,217,218,219,220,221,222,223,224,225,226,227,228,229,230,231] |
| Curdlan | First extracted from Alcaligenes faecalis var. myxogenes, (later reclassified as Agrobacterium sp.) | Used as a biological response modifier because of their immunostimulatory properties, anti-infective, and anti-inflammatory Encapsulation vehicle for carrying drugs and other molecules | [232,233,234,235,236,237,238,239,240,241,242,243,244,245,246,247] |
| x | R | Polymer Name | Abbreviation | Type |
|---|---|---|---|---|
| 1 | methyl | Poly-3-hydroxybutyrate | P(3HB) | scl |
| ethyl | Poly-3-hydroxyvalerate | P(3HV) | scl | |
| propyl | Poly-3-hydroxyhexanoate | P(3HHx) | mcl | |
| pentyl | Poly-3-hydroxyoctanoate | P(3HO) | mcl | |
| nonyl | Poly-3-hydroxydodecanoate | P(3HDD) | lcl | |
| 2 | H | Poly-4-hydroxybutyrate | P(4HB) | scl |
| methyl | Poly-3-hydroxyvalerate | P(4HV) | scl | |
| 3 | H | Poly-5-hydroxyvalerate | P(5HV) | scl |
| methyl | Poly-5-hydroxyhexanoate | P(5HHx) | scl | |
| 4 | hexyl | Poly-6-hydroxydodecanoate | P(6HDD) | mcl |
| Polymer | Polymer-Accumulating Bacteria | Biomaterial Properties in Biomedical Application | Ref. |
|---|---|---|---|
| Polyhydroxyalkanoates | First isolated from Bacillus megaterium Multiple strains of Bacillus and Pseudomonas, including P. putida and B. aquamaris | Several aspects have been considered, including wound healing patches by promoting angiogenesis in the healing process, bioresorbable sutures, and in drug delivery with a tailorable material degradation rate Useful in scaffold development for tissue engineering applications, which is biocompatible for a number of tissue types by facilitating cell seeding, adhesion, proliferation, differentiation, and de novo tissue regeneration. | [248,249,250,251,252,253,254,255,256,257,258,259,260,261,262,263,264,265,266,267,268,269,270,271,272,273,274,275,276,277,278,279,280,281,282,283,284,285,286,287,288,289,290,291,292,293,294,295,296,297] |
| Polylactic acid | PLA monomeric components being synthesized by bacteria of the order Lactobacillales Genetically modified Escherichia coli | PLA is bioresorbable, allowing the material to naturally disintegrate as the target site is healing Acts as a scaffold for tissue engineering application and bone fixation purposes Prospective drug delivery material due to its tailorable porosity for controlled adsorption and drug release | [298,299,300,301,302,303,304,305,306,307,308,309,310,311,312,313,314,315,316,317,318] |
| Polymer | Polymer-Accumulating Bacteria | Biomaterial Properties in Biomedical Application | Ref. |
|---|---|---|---|
| ε-poly-l-lysine | Streptococcus albulus Streptococcus graminearus Lactobacillus delbrueckii | Cationic properties make ε-poly-l-lysine and excellent antimicrobial biomaterial and DNA binding for future use in gene therapies Has successfully been used as an antibiotic coating on titanium implants Strongly adhesive properties have suggested ε-poly-l-lysine’s role in adhesive wound healing dressings. | [319,320,321,322,323,324,325,326,327,328,329,330,331,332,333,334,335,336,337,338,339,340,341,342,343,344,345,346,347,348] |
| Poly-γ-glutamate | Multiple strains of genus “Bacillus”, including B. anthracis, B. subtilis, B. licheniformis and B. amyloliquefaciens B. subtilis is responsible for most commercial production. | Proposed applications in antimicrobials due to its resistance to protease virulence factors Has improved the bacterial killing capacity of existing and experimental antimicrobials Pro-inflammatory effect may be beneficial in wound healing | [349,350,351,352,353,354,355,356,357,358,359,360,361,362,363,364,365,366,367,368,369,370,371,372,373,374,375,376] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hinchliffe, J.D.; Parassini Madappura, A.; Syed Mohamed, S.M.D.; Roy, I. Biomedical Applications of Bacteria-Derived Polymers. Polymers 2021, 13, 1081. https://doi.org/10.3390/polym13071081
Hinchliffe JD, Parassini Madappura A, Syed Mohamed SMD, Roy I. Biomedical Applications of Bacteria-Derived Polymers. Polymers. 2021; 13(7):1081. https://doi.org/10.3390/polym13071081
Chicago/Turabian StyleHinchliffe, Jonathan David, Alakananda Parassini Madappura, Syed Mohammad Daniel Syed Mohamed, and Ipsita Roy. 2021. "Biomedical Applications of Bacteria-Derived Polymers" Polymers 13, no. 7: 1081. https://doi.org/10.3390/polym13071081
APA StyleHinchliffe, J. D., Parassini Madappura, A., Syed Mohamed, S. M. D., & Roy, I. (2021). Biomedical Applications of Bacteria-Derived Polymers. Polymers, 13(7), 1081. https://doi.org/10.3390/polym13071081