

Acceleration of Polybutylene Succinate Biodegradation by Terribacillus sp. JY49 Isolated from a Marine Environment

 ,
,
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Preparing Solid Media Containing Plastic
2.3. Screening Microorganisms for Their Ability to Degrade PBS
2.4. Characterizing the PBS-Degrading Strain
2.5. Solid and Liquid Culture to Monitor PBS Degradation
2.6. GC-MS Analysis
2.7. Analyzing the Physical Properties of PBS Films
2.8. Confirming the PBS Monomer Effect
2.9. Esterase Activity Assay with p-Nitrophenyl Esters
3. Results
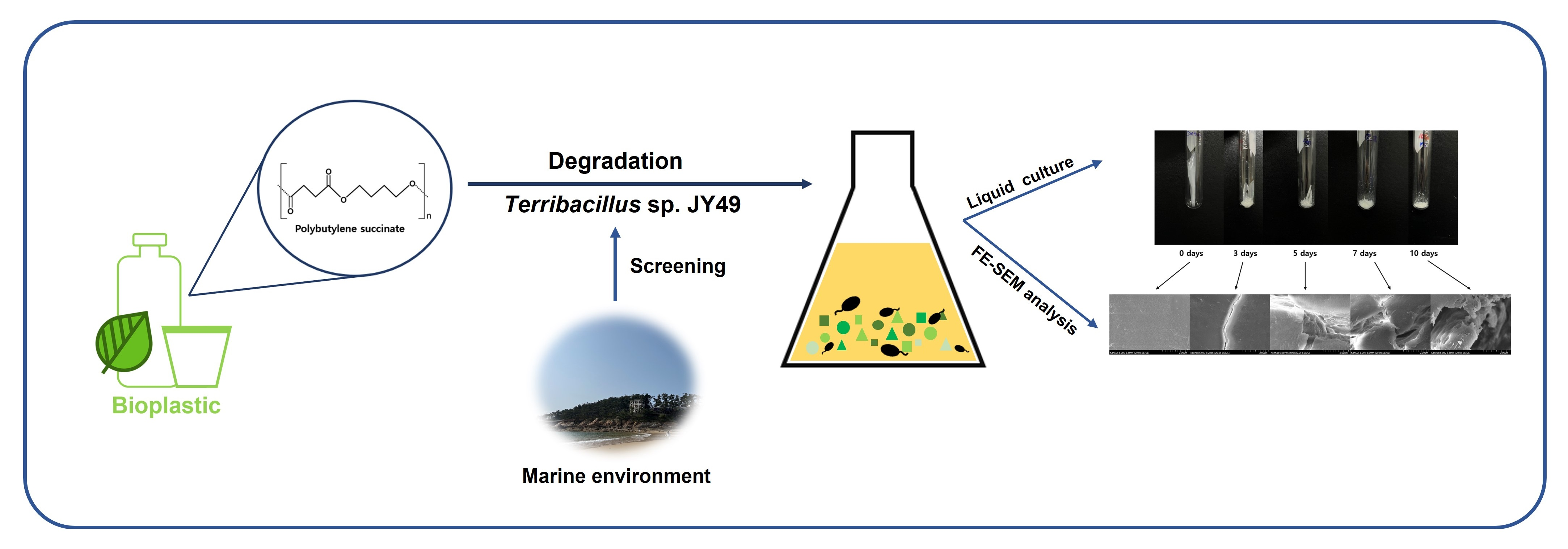
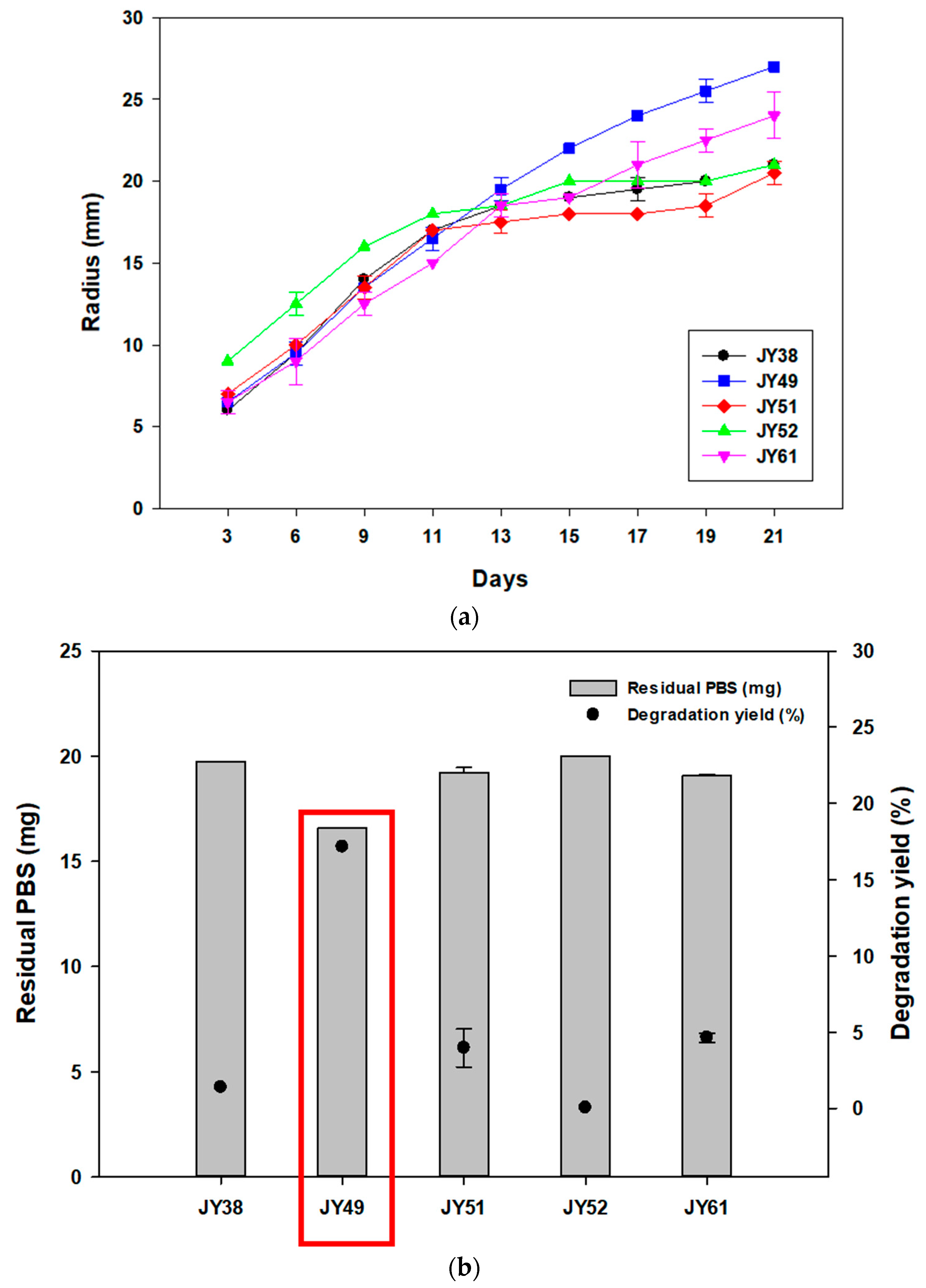
3.1. Screening Polybutylene Succinate (PBS)-Degrading Strains from Marine Samples
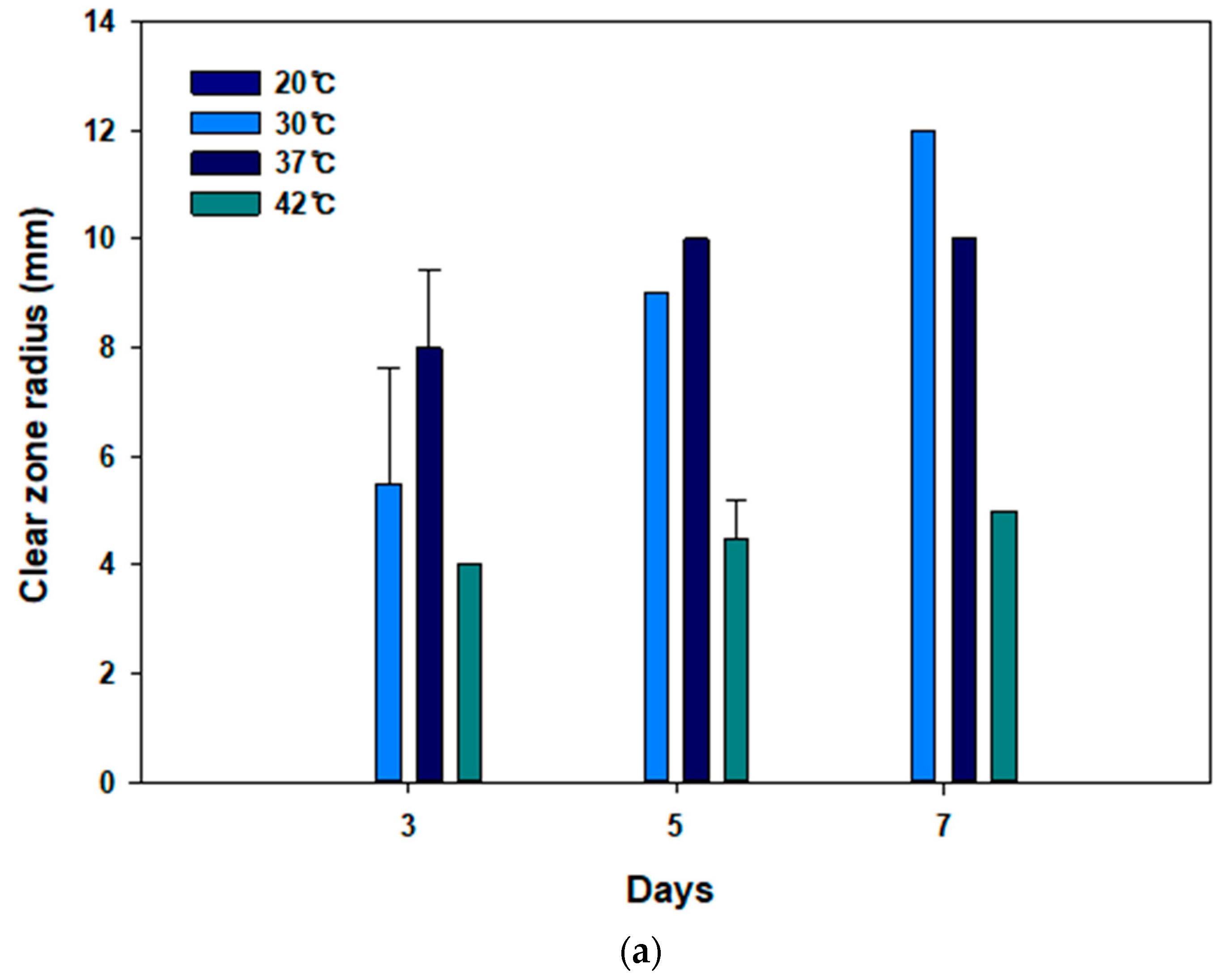
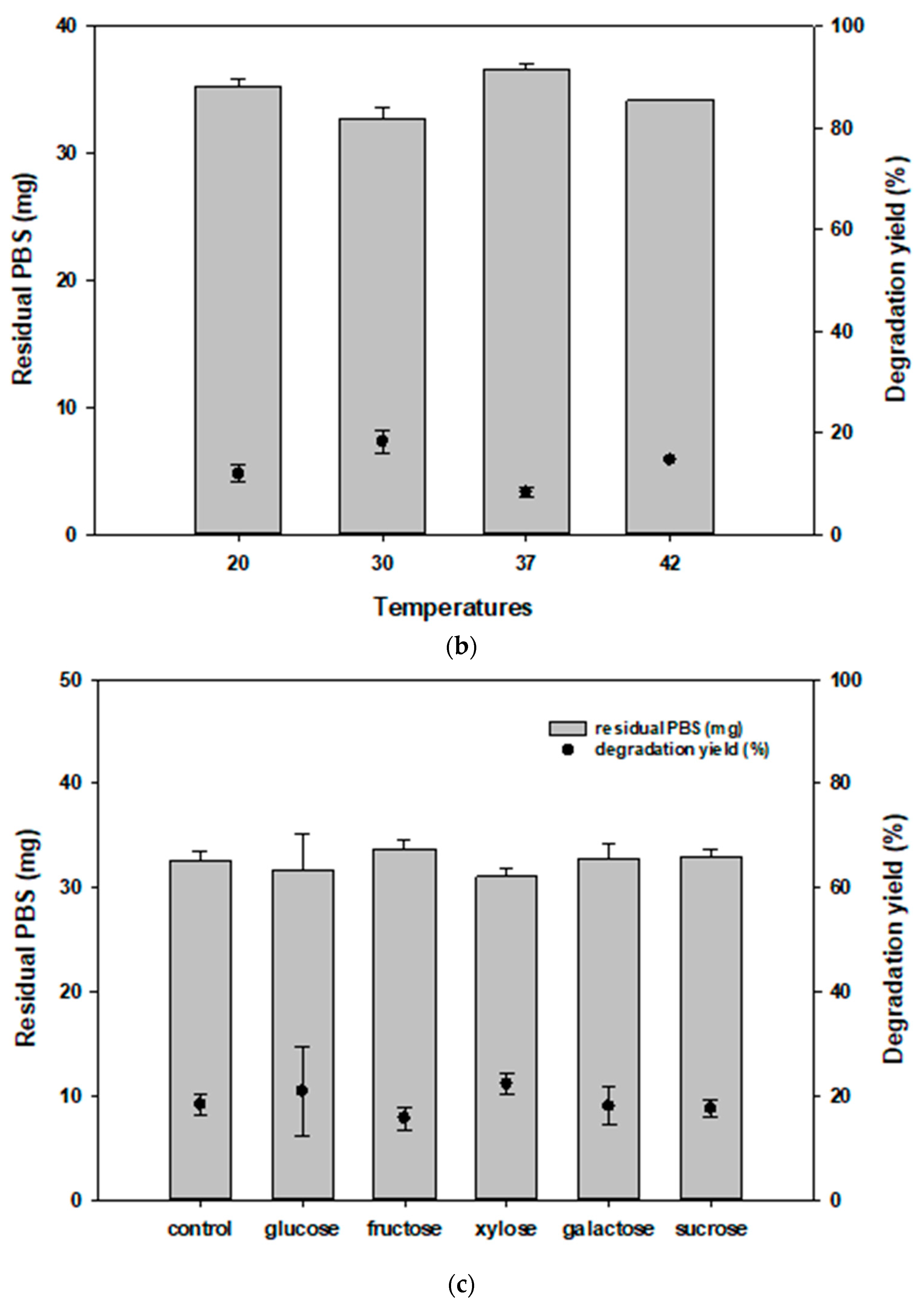
3.2. Optimizing Temperature Conditions for PBS Degradation by JY49
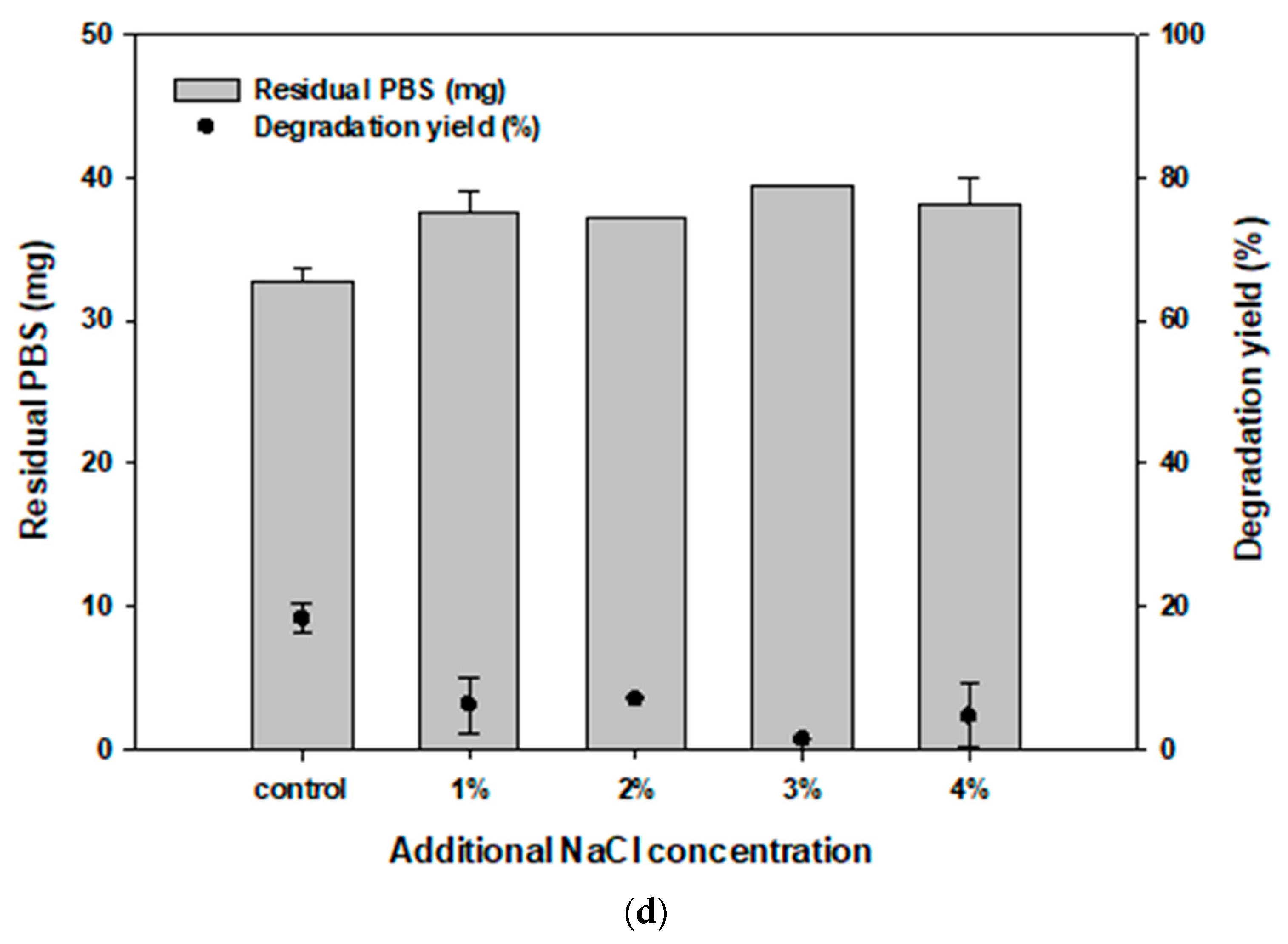
3.3. Effect of Carbon and Salt Concentration on PBS Degradation
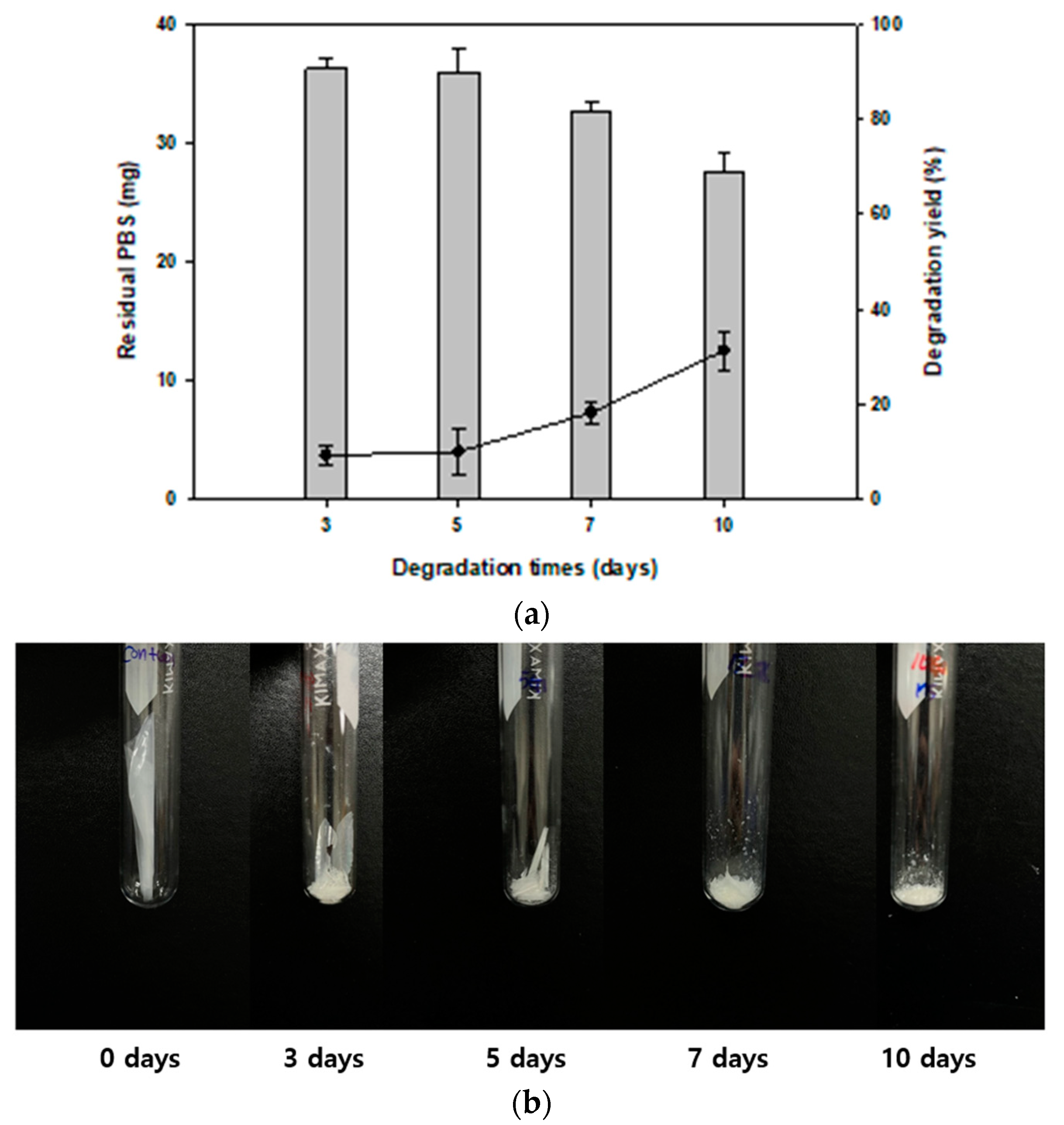
3.4. Time-Dependent Monitoring of PBS Degradation
3.5. Changes in Physical Properties of PBS Films
3.6. Effect of PBS Monomers such as Succinic Acid and 1,4-Butanediol on the Growth of JY49
3.7. Degradability of Other Bioplastics by Terribacillus sp. JY49
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Landrigan, P.J.; Stegeman, J.J.; Fleming, L.E.; Allemand, D.; Anderson, D.M.; Backer, L.C.; Brucker-Davis, F.; Chevalier, N.; Corra, L.; Czerucka, D.; et al. Human Health and Ocean Pollution. Ann. Glob. Health. 2020, 86, 151. [Google Scholar] [CrossRef] [PubMed]
- Bahl, S.; Dolma, J.; Singh, J.J.; Sehgal, S. Biodegradation of Plastics: A State of the Art Review. Mater. Today Proc. 2021, 39, 31–34. [Google Scholar] [CrossRef]
- Evode, N.; Qamar, S.A.; Bilal, M.; Barceló, D.; Iqbal, H.M.N. Plastic Waste and Its Management Strategies for Environmental Sustainability. Case Stud. Chem. Environ. Eng. 2021, 4, 100142. [Google Scholar] [CrossRef]
- Chia, W.Y.; Tang, D.Y.Y.; Khoo, K.S.; Lup, A.N.K.; Chew, K.W. Nature’s Fight against Plastic Pollution: Algae for Plastic Biodegradation and Bioplastics Production. Environ. Sci. Ecotechnol. 2020, 4, 100065. [Google Scholar] [CrossRef]
- Sivan, A. New Perspectives in Plastic Biodegradation. Curr. Opin. Biotechnol. 2011, 22, 422–426. [Google Scholar] [CrossRef] [PubMed]
- DiGregorio, B.E. Biobased Performance Bioplastic: Mirel. Chem. Biol. 2009, 16, 1–2. [Google Scholar] [CrossRef]
- Nilani, P.; Raveesha, P.; Kasthuribai, B.R.N.N.; Duraisamy, B.; Dhamodaran, P.; Elango, K. Formulation and Evaluation of Polysaccharide Based Biopolymer—An Ecofriendly Alternative for Synthetic Polymer. J. Pharm. Sci. Res. 2010, 2, 178–184. [Google Scholar]
- Muneer, F.; Rasul, I.; Azeem, F.; Siddique, M.H.; Zubair, M.; Nadeem, H. Microbial Polyhydroxyalkanoates (PHAs): Efficient Replacement of Synthetic Polymers. J. Polym. Environ. 2020, 28, 2301–2323. [Google Scholar] [CrossRef]
- Shah, M.; Rajhans, S.; Pandya, H.A.; Mankad, A.U. Bioplastic for Future: A Review Then and Now. World J. Adv. Res. Rev. 2021, 9, 56–67. [Google Scholar] [CrossRef]
- Shah, A.; Alshehrei, F. Biodegradation of Synthetic and Natural Plastic by Microorganisms. J. Appl. Environ. Microbiol. 2017, 5, 8–19. [Google Scholar]
- Sharma, S.R. Bioremediation of Polythenes and Plastics: A Microbial Approach. In Nanotechnology in the Life Sciences; Springer: Cham, Switzerland, 2018; pp. 97–114. [Google Scholar]
- Bano, N.; Younas, T.; Shoaib, F.; Rashid, D.; Jaffri, N. Plastic: Reduce, Recycle, and Environment. In Environmentally-Benign Energy Solutions; Springer: Cham, Switzerland, 2020; pp. 191–208. [Google Scholar]
- Ben, M.; Mato, T.; Lopez, A.; Vila, M.; Kennes, C.; Veiga, M.C. Bioplastic Production Using Wood Mill Effluents as Feedstock. Water Sci. Technol. 2011, 63, 1196–1202. [Google Scholar] [CrossRef] [PubMed]
- Chin, I.-J. Biodegradable Polymers and Plastics. Chem. Int.-Newsmag. IUPAC 2004, 26, 28–29. [Google Scholar]
- Scaffaro, R.; Maio, A.; Sutera, F.; Gulino, E.F.; Morreale, M. Degradation and Recycling of Films Based on Biodegradable Polymers: A Short Review. Polymers 2019, 11, 651. [Google Scholar] [CrossRef] [Green Version]
- Patel, M.; Bastioli, C.; Marini, L.; Würd, D.E. Biopolymers Online—Environmental Assessment of Bio-Based Polymers and Natural Fibres; Utrecht University: Utrecht, The Netherlands, 2005. [Google Scholar]
- Dilshad, E.; Waheed, H.; Ali, U.; Amin, A.; Ahmed, I. General Structure and Classification of Bioplastics and Biodegradable Plastics. In Bioplastics for Sustainable Development; Kuddus, M., Roohi, Eds.; Springer Nature: Singapore, 2021; pp. 61–82. [Google Scholar]
- Jiang, L.; Zhang, J. Biodegradable and Biobased Polymers. In Applied Plastics Engineering Handbook: Processing, Materials, and Applications, 2nd ed.; William Andrew Publishing: Norwich, NY, USA, 2017; pp. 127–143. [Google Scholar] [CrossRef]
- Pinazo, J.M.; Domine, M.E.; Parvulescu, V.; Petru, F. Sustainability Metrics for Succinic Acid Production: A Comparison between Biomass-Based and Petrochemical Routes. Catal. Today 2015, 239, 17–24. [Google Scholar] [CrossRef]
- Datta, J.; Kasprzyk, P.; Błażek, K.; Włoch, M. Synthesis, Structure and Properties of Poly(Ester-Urethane)s Obtained Using Bio-Based and Petrochemical 1,3-Propanediol and 1,4-Butanediol. J. Therm. Anal. Calorim. 2017, 130, 261–276. [Google Scholar] [CrossRef]
- Kumari Pallathadka, P.; Koh, X.Q.; Khatta, A.; Luckachan, G.E.; Mittal, V. Characteristics of Biodegradable Poly(Butylene Succinate) Nanocomposites with Thermally Reduced Graphene Nanosheets. Polym. Compos. 2017, 38, E42–E48. [Google Scholar] [CrossRef]
- Rafiqah, S.A.; Khalina, A.; Harmaen, A.S.; Tawakkal, I.A.; Zaman, K.; Asim, M.; Nurrazi, M.N.; Lee, C.H. A Review on Properties and Application of Bio-based Poly(Butylene Succinate). Polymers 2021, 13, 1436. [Google Scholar] [CrossRef] [PubMed]
- Sheikholeslami, S.N.; Rafizadeh, M.; Taromi, F.A.; Shirali, H.; Jabbari, E. Material Properties of Degradable Poly(Butylene Succinate-Co-Fumarate) Copolymer Networks Synthesized by Polycondensation of Pre-Homopolyesters. Polymer 2016, 98, 70–79. [Google Scholar] [CrossRef]
- Kong, X.; Qi, H.; Curtis, J.M. Synthesis and Characterization of High-Molecular Weight Aliphatic Polyesters from Monomers Derived from Renewable Resources. J. Appl. Polym. Sci. 2014, 131, 4401–4404. [Google Scholar] [CrossRef]
- Adhikari, D.; Mukai, M.; Kubota, K.; Kai, T.; Kaneko, N.; Araki, K.S.; Kubo, M. Degradation of Bioplastics in Soil and Their Degradation Effects on Environmental Microorganisms. J. Agric. Chem. Environ. 2016, 5, 23. [Google Scholar] [CrossRef]
- Mao, H.; Liu, H.; Gao, Z.; Su, T.; Wang, Z. Biodegradation of Poly(Butylene Succinate) by Fusarium Sp. FS1301 and Purification and Characterization of Poly(Butylene Succinate) Depolymerase. Polym. Degrad. Stab. 2015, 114, 1–7. [Google Scholar] [CrossRef]
- Mei, X.; Tian, C.; Dong, Q.; Liang, Y. Influencing Factors and Process on In Situ Degradation of Poly(Butylene Succinate) Film by Strain Bionectria ochroleuca BFM-X1 in Soil. J. Environ. Prot. 2012, 3, 523–532. [Google Scholar] [CrossRef] [Green Version]
- Ishii, N.; Inoue, Y.; Tagaya, T.; Mitomo, H.; Nagai, D.; Kasuya, K.I. Isolation and Characterization of Poly(Butylene Succinate)-Degrading Fungi. Polym. Degrad. Stab. 2008, 93, 883–888. [Google Scholar] [CrossRef]
- Abe, M.; Kobayashi, K.; Honma, N.; Nakasaki, K. Microbial Degradation of Poly(Butylene Succinate) by Fusarium Solani in Soil Environments. Polym. Degrad. Stab. 2010, 95, 138–143. [Google Scholar] [CrossRef]
- Zhao, J.H.; Wang, X.Q.; Zeng, J.; Yang, G.; Shi, F.H.; Yan, Q. Biodegradation of Poly(Butylene Succinate) in Compost. J. Appl. Polym. Sci. 2005, 97, 2273–2278. [Google Scholar] [CrossRef]
- Uchida, H.; Nakajima-Kambe, T.; Shigeno-Akutsu, Y.; Nomura, N.; Tokiwa, Y.; Nakahara, T. Properties of a Bacterium Which Degrades Solid Poly(Tetramethylene Succinate)-Co-Adipate, a Biodegradable Plastic. FEMS Microbial Lett. 2000, 189, 25–29. [Google Scholar] [CrossRef] [PubMed]
- Park, S.L.; Cho, J.Y.; Kim, S.H.; Bhatia, S.K.; Gurav, R.; Park, S.H.; Park, K.; Yang, Y.H. Isolation of Microbulbifer Sp. Sol66 with High Polyhydroxyalkanoate-Degrading Activity from the Marine Environment. Polymers 2021, 13, 4257. [Google Scholar] [CrossRef]
- Park, S.L.; Cho, J.Y.; Choi, T.R.; Song, H.S.; Bhatia, S.K.; Gurav, R.; Park, S.H.; Park, K.; Joo, J.C.; Hwang, S.Y.; et al. Improvement of Polyhydroxybutyrate (PHB) Plate-Based Screening Method for PHB Degrading Bacteria Using Cell-Grown Amorphous PHB and Recovered by Sodium Dodecyl Sulfate (SDS). Int. J. Biol. Macromol. 2021, 177, 413–421. [Google Scholar] [CrossRef]
- Park, Y.L.; Bhatia, S.K.; Gurav, R.; Choi, T.R.; Kim, H.J.; Song, H.S.; Park, J.Y.; Han, Y.H.; Lee, S.M.; Park, S.L.; et al. Fructose Based Hyper Production of Poly-3-Hydroxybutyrate from Halomonas Sp. YLGW01 and Impact of Carbon Sources on Bacteria Morphologies. Int. J. Biol. Macromol. 2020, 154, 929–936. [Google Scholar] [CrossRef]
- Lee, S.M.; Lee, H.J.; Kim, S.H.; Suh, M.J.; Cho, J.Y.; Ham, S.; Jeon, J.M.; Yoon, J.J.; Bhatia, S.K.; Gurav, R.; et al. Screening of the Strictly Xylose-Utilizing Bacillus Sp. SM01 for Polyhydroxybutyrate and Its Co-Culture with Cupriavidus Necator NCIMB 11599 for Enhanced Production of PHB. Int. J. Biol. Macromol. 2021, 181, 410–417. [Google Scholar] [CrossRef]
- Cho, J.Y.; Park, S.L.; Lee, H.J.; Kim, S.H.; Suh, M.J.; Ham, S.; Bhatia, S.K.; Gurav, R.; Park, S.H.; Park, K.; et al. Polyhydroxyalkanoates (PHAs) Degradation by the Newly Isolated Marine Bacillus Sp. JY14. Chemosphere 2021, 283, 131172. [Google Scholar] [CrossRef] [PubMed]
- Bhatia, S.K.; Yoon, J.J.; Kim, H.J.; Hong, J.W.; Hong, Y.G.; Song, H.S.; Moon, Y.M.; Jeon, J.M.; Kim, Y.G.; Yang, Y.H. Engineering of Artificial Microbial Consortia of Ralstonia Eutropha and Bacillus Subtilis for Poly(3-Hydroxybutyrate-Co-3-Hydroxyvalerate) Copolymer Production from Sugarcane Sugar without Precursor Feeding. Bioresour. Technol. 2018, 257, 92–101. [Google Scholar] [CrossRef] [PubMed]
- Jung, H.R.; Choi, T.R.; Han, Y.H.; Park, Y.L.; Park, J.Y.; Song, H.S.; Yang, S.Y.; Bhatia, S.K.; Gurav, R.; Park, H.A.; et al. Production of Blue-Colored Polyhydroxybutyrate (PHB) by One-Pot Production and Coextraction of Indigo and PHB from Recombinant Escherichia Coli. Dye. Pigment. 2020, 173, 107889. [Google Scholar] [CrossRef]
- Choi, T.-R.; Park, Y.-L.; Song, H.-S.; Lee, S.M.; Park, S.L.; Lee, H.S.; Kim, H.-J.; Bhatia, S.K.; Gurav, R.; Choi, K.-Y.; et al. Fructose-Based Production of Short-Chain-Length and Medium-Chain-Length Polyhydroxyalkanoate Copolymer by Arctic Pseudomonas Sp. B14-6 Tae-Rim. Polymers 2021, 13, 1398. [Google Scholar] [CrossRef] [PubMed]
- Cho, J.Y.; Park, S.L.; Kim, S.H.; Jung, H.J.; Cho, D.H.; Kim, B.C.; Bhatia, S.K.; Gurav, R.; Park, S.H.; Park, K.; et al. Novel Poly(Butylene Adipate-Co-Terephthalate)-Degrading Bacillus Sp. JY35 from Wastewater Sludge and Its Broad Degradation of Various Bioplastics. Waste Manag. 2022, 144, 1–10. [Google Scholar] [CrossRef]
- Yun, H.J.; Lee, Y.J.; Yeo, S.H.; Choi, H.S.; Park, H.Y.; Park, H.D.; Baek, S.Y. The Isolation and Culture Characterization of a Lipolytic Enzyme Producing Strain from Meju. Korean J. Microbiol. Biotechnol. 2012, 40, 98–103. [Google Scholar] [CrossRef]
- An, S.Y.; Asahara, M.; Goto, K.; Kasai, H.; Yokota, A. Terribacillus Saccharophilus Gen. Nov., Sp. Nov. and Terribacillus Halophilus Sp. Nov., Spore-Forming Bacteria Isolated from Field Soil in Japan. Int. Syst. Evol. Microbiol. 2007, 57, 51–55. [Google Scholar] [CrossRef]
- Essghaier, B. Antimicrobial Behavior of Intracellular Proteins from Two Moderately Halophilic Bacteria: Strain J31 of Terribacillus Halophilus and Strain M3-23 of Virgibacillus Marismortui. J. Plant. Pathol Microbiol 2014, 5, 1. [Google Scholar] [CrossRef]
- Liu, W.; Jiang, L.; Guo, C.; Yang, S.S. Terribacillus Aidingensis Sp. Nov., a Moderately Halophilic Bacterium. Int. J. Syst. Evol. Microbiol. 2010, 60, 2940–2945. [Google Scholar] [CrossRef]
- Krishnamurthi, S.; Chakrabarti, T. Proposal for Transfer of Pelagibacillus Goriensis Kim et al. 2007 to the Genus Terribacillus as Terribacillus Goriensis Comb. Nov. Int. J. Syst. Evol. Microbiol. 2008, 58, 2287–2291. [Google Scholar] [CrossRef]
- Kim, Y.G.; Hwang, C.Y.; Yoo, K.W.; Moon, H.T.; Yoon, J.H.; Cho, B.C. Pelagibacillus Goriensis Gen. Nov., Sp. Nov., a Moderately Halotolerant Bacterium Isolated from Coastal Water off the East Coast of Korea. Int. J. Syst. Evol. Microbiol. 2007, 57, 1554–1560. [Google Scholar] [CrossRef] [PubMed]
- Nagaroor, V.; Gummadi, S.N. Biochemical Characterization of an Esterase from Clostridium Acetobutylicum with Novel GYSMG Pentapeptide Motif at the Catalytic Domain. J. Ind. Microbiol. Biotechnol. 2020, 47, 169–181. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Type | Temp | Period (Days) | Weight Loss (%) | Biodegradation Percentage (%) | Condition | Reference |
|---|---|---|---|---|---|---|---|
| Fusarium sp. FS1301 | Fungi | 30 °C | 21 | 80% | - | Liquid | [26] |
| Bionectria ochroleuca BFM-X1 | Fungi | 30 °C | 30 | 60% | - | Soil | [27] |
| Aspergillus fumigatus | Fungi | 30 °C | 30 | 80% | - | Soil | [28] |
| Fusarium solani | Fungi | - | 14 | - | 2.8% | Soil | [29] |
| Aspergillus versicolor, Penicillium, Bacillus, Thermopolyarpora | Consortia | - | 90 | - | 71.9% (powder) 60.7% (film) 14.1% (granule) | Soil | [30] |
| Terribacillus goriensis | Bacteria | 30 °C | 10 | - | 31.4% | Liquid | This paper |
| Strains | Strain No. | Remark | Isolated Site | Reference |
|---|---|---|---|---|
| Terribacillus saccharophilus | KCTC 13936, DSM 21620 | Characterization * | Field soil | [42] |
| Terribacillus halophilus | KCTC 13936, DSM 21620 | Characterization *, Antimicrobial behavior ** | Field soil | [42,43] |
| Terribacillus aidingensis | DSM 28352, CGMCC 1.8913 | Characterization * | Soil from lake | [44] |
| Terribacillus goriensis | KCCM 42329, DSM 18252 | Characterization * | Surface of sea water | [45] |
| Day | Mn × 104 | Mw × 104 | PDI |
|---|---|---|---|
| 0 | 3.87 | 9.49 | 2.45 |
| 3 | 3.57 | 8.56 | 2.40 |
| 5 | 3.38 | 8.31 | 2.46 |
| 7 | 3.11 | 7.63 | 2.45 |
| 10 | 2.88 | 6.82 | 2.37 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, S.H.; Cho, J.Y.; Cho, D.H.; Jung, H.J.; Kim, B.C.; Bhatia, S.K.; Park, S.-H.; Park, K.; Yang, Y.-H. Acceleration of Polybutylene Succinate Biodegradation by Terribacillus sp. JY49 Isolated from a Marine Environment. Polymers 2022, 14, 3978. https://doi.org/10.3390/polym14193978
Kim SH, Cho JY, Cho DH, Jung HJ, Kim BC, Bhatia SK, Park S-H, Park K, Yang Y-H. Acceleration of Polybutylene Succinate Biodegradation by Terribacillus sp. JY49 Isolated from a Marine Environment. Polymers. 2022; 14(19):3978. https://doi.org/10.3390/polym14193978
Chicago/Turabian StyleKim, Su Hyun, Jang Yeon Cho, Do Hyun Cho, Hee Ju Jung, Byung Chan Kim, Shashi Kant Bhatia, See-Hyoung Park, Kyungmoon Park, and Yung-Hun Yang. 2022. "Acceleration of Polybutylene Succinate Biodegradation by Terribacillus sp. JY49 Isolated from a Marine Environment" Polymers 14, no. 19: 3978. https://doi.org/10.3390/polym14193978
APA StyleKim, S. H., Cho, J. Y., Cho, D. H., Jung, H. J., Kim, B. C., Bhatia, S. K., Park, S. -H., Park, K., & Yang, Y. -H. (2022). Acceleration of Polybutylene Succinate Biodegradation by Terribacillus sp. JY49 Isolated from a Marine Environment. Polymers, 14(19), 3978. https://doi.org/10.3390/polym14193978