
Alginate-Induced Disease Resistance in Plants

 and
and 
Abstract
:1. Introduction
2. Plant Immune System against Pathogens
2.1. Systemic Acquired Resistance (SAR)
2.2. Induced Systemic Resistance (ISR)
3. Abiotic Inducers of Disease Resistance in Plants
3.1. Polysaccharides as Plant Defense Inducers
3.2. Alginate and Induction of Resistance against Plant Pathogens
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Al-Ani, L.K.T.; Furtado, E.L. The effect of incompatible plant pathogens on the host plant. In Molecular Aspects of Plant Beneficial Microbes in Agriculture; Academic Press: Cambridge, MA, USA, 2020; pp. 47–57. [Google Scholar] [CrossRef]
- Surico, G. The concepts of plant pathogenicity, virulence/avirulence and effector proteins by a teacher of plant pathology. Phytopathol. Mediterr. 2013, 52, 399–417. [Google Scholar] [CrossRef]
- de León, I.P.; Montesano, M. Activation of defense mechanisms against pathogens in mosses and flowering plants. Int. J. Mol. Sci. 2013, 14, 3178–3200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, S.; Tang, W.; Jiang, L.; Hou, Y.; Yang, F.; Chen, W.; Li, X. Elicitor activity of algino-oligosaccharide and its potential application in protection of rice plant (Oryza sativa L.) against Magnaporthe grisea. Biotechnol. Biotechnol. Equip. 2015, 29, 646–652. [Google Scholar] [CrossRef] [Green Version]
- Guo, J.; Du, G.; Chen, J.; Chen, X.; Li, X. Oligosaccharides act as elicitors to protect plant against crop disease based on knowledge of plant defence response mechanism. In Sugar Beet Crops: Growth, Fertilization and Yield; Nova Science Publishers: New York, NY, USA, 2009. [Google Scholar]
- Conrath, U. Priming of induced plant defense responses. In Advances in Botanical Research; Academic Press: Cambridge, MA, USA, 2009; Volume 51, pp. 361–395. [Google Scholar]
- Aranega-Bou, P.; de la O Leyva, M.; Finiti, I.; García-Agustín, P.; González-Bosch, C. Priming of plant resistance by natural compounds. Hexanoic acid as a model. Front. Plant Sci. 2014, 5, 488. [Google Scholar] [CrossRef] [Green Version]
- Gozzo, F.; Faoro, F. Systemic Acquired Resistance (50 Years after Discovery): Moving from the Lab to the Field. J. Agric. Food Chem. 2013, 61, 12473–12491. [Google Scholar] [CrossRef]
- Cohen, Y.R. β-Aminobutyric Acid-Induced Resistance against Plant Pathogens. Plant Dis. 2002, 86, 448–457. [Google Scholar] [CrossRef] [Green Version]
- Iriti, M.; Castorina, G.; Vitalini, S.; Mignani, I.; Soave, C.; Fico, G.; Faoro, F. Chitosan-induced ethylene-independent resistance does not reduce crop yield in bean. Biol. Control 2010, 54, 241–247. [Google Scholar] [CrossRef]
- Zhou, J.; Sun, A.; Xing, D. Modulation of cellular redox status by thiamine-activated NADPH oxidase confers Arabidopsis resistance to Sclerotinia sclerotiorum. J. Exp. Bot. 2013, 64, 3261–3272. [Google Scholar] [CrossRef]
- Song, G.C.; Choi, H.K.; Ryu, C.-M. The folate precursor para-aminobenzoic acid elicits induced resistance against Cucumber mosaic virus and Xanthomonas axonopodis. Ann. Bot. 2013, 111, 925–934. [Google Scholar] [CrossRef] [Green Version]
- El Hadrami, A.; Adam, L.R.; El Hadrami, I.; Daayf, F. Chitosan in Plant Protection. Mar. Drugs 2010, 8, 968–987. [Google Scholar] [CrossRef]
- Rasul, S.; Dubreuil-Maurizi, C.; Lamotte, O.; Koen, E.; Poinssot, B.; Alcaraz, G.; Wendehenne, D.; Jeandroz, S. Nitric oxide production mediates oligogalacturonide-triggered immunity and resistance to Botrytis cinerea in Arabidopsis thaliana. Plant Cell Environ. 2012, 35, 1483–1499. [Google Scholar] [CrossRef] [PubMed]
- Moenne, A.; González, A. Chitosan-, alginate-carrageenan-derived oligosaccharides stimulate defense against biotic and abiotic stresses, and growth in plants: A historical perspective. Carbohydr. Res. 2021, 503, 108298. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Zhu, L.; Lu, G.; Zhan, X.-B.; Lin, C.-C.; Zheng, Z.-Y. Curdlan β-1,3-Glucooligosaccharides Induce the Defense Responses against Phytophthora infestans Infection of Potato (Solanum tuberosum L. cv. McCain G1) Leaf Cells. PLoS ONE 2014, 9, e97197. [Google Scholar] [CrossRef] [PubMed]
- Shibuya, N.; Minami, E. Oligosaccharide signalling for defence responses in plant. Physiol. Mol. Plant Pathol. 2001, 59, 223–233. [Google Scholar] [CrossRef] [Green Version]
- Vera, J.; Castro, J.; Gonzalez, A.; Moenne, A. Seaweed Polysaccharides and Derived Oligosaccharides Stimulate Defense Responses and Protection Against Pathogens in Plants. Mar. Drugs 2011, 9, 2514–2525. [Google Scholar] [CrossRef] [PubMed]
- Stengel, D.B.; Connan, S. Marine algae: A source of biomass for biotechnological applications. In Natural Products from Marine Algae; Humana Press: New York, NY, USA, 2015; pp. 1–37. [Google Scholar]
- van Aubel, G.; Buonatesta, R.; Van Cutsem, P. COS-OGA: A novel oligosaccharidic elicitor that protects grapes and cucumbers against powdery mildew. Crop Prot. 2014, 65, 129–137. [Google Scholar] [CrossRef]
- van Aubel, G.; Cambier, P.; Dieu, M.; Van Cutsem, P. Plant immunity induced by COS-OGA elicitor is a cumulative process that involves salicylic acid. Plant Sci. 2016, 247, 60–70. [Google Scholar] [CrossRef]
- Guarnizo, N.; Oliveros, D.; Murillo-Arango, W.; Bermúdez-Cardona, M.B. Oligosaccharides: Defense Inducers, Their Recognition in Plants, Commercial Uses and Perspectives. Molecules 2020, 25, 5972. [Google Scholar] [CrossRef]
- Hu, X.; Jiang, X.; Hwang, H.; Liu, S.; Guan, H. Promotive effects of alginate-derived oligosaccharide on maize seed germination. J. Appl. Phycol. 2004, 16, 73–76. [Google Scholar] [CrossRef]
- Xu, X.; Iwamoto, Y.; Kitamura, Y.; Oda, T.; Muramatsu, T. Root Growth-promoting Activity of Unsaturated Oligomeric Uronates from Alginate on Carrot and Rice Plants. Biosci. Biotechnol. Biochem. 2003, 67, 2022–2025. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Howlader, P.; Liu, T.; Sun, X.; Jia, X.; Zhao, X.; Shen, P.; Qin, Y.; Wang, W.; Yin, H. Alginate Oligosaccharide (AOS) induced resistance to Pst DC3000 via salicylic acid-mediated signaling pathway in Arabidopsis thaliana. Carbohydr. Polym. 2019, 225, 115221. [Google Scholar] [CrossRef] [PubMed]
- Popletaeva, S.B.; Statsyuk, N.V.; Voinova, T.M.; Arslanova, L.R.; Zernov, A.L.; Bonartsev, A.P.; Bonartseva, G.A.; Dzhavakhiya, V.G. Evaluation of eliciting activity of peptidil prolyl cys/trans isomerase from Pseudonomas fluorescens encapsulated in sodium alginate regarding plant resistance to viral and fungal pahogens. AIMS Microbiol. 2018, 4, 192–208. [Google Scholar] [CrossRef] [PubMed]
- Dey, P.; Ramanujam, R.; Venkatesan, G.; Nagarathnam, R. Sodium alginate potentiates antioxidant defense and PR proteins against early blight disease caused by Alternaria solani in Solanum lycopersicum Linn. PLoS ONE 2019, 14, e0223216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akimoto, C.; Aoyagi, H.; Dicosmo, F.; Tanaka, H. Synergistic effect of active oxygen species and alginate on chitinase production by Wasabia japonica cells and its application. J. Biosci. Bioeng. 2000, 89, 131–137. [Google Scholar] [CrossRef]
- Ausubel, F.M. Are innate immune signaling pathways in plants and animals conserved? Nat. Immunol. 2005, 6, 973–979. [Google Scholar] [CrossRef] [PubMed]
- Bittel, P.; Robatzek, S. Microbe-associated molecular patterns (MAMPs) probe plant immunity. Curr. Opin. Plant Biol. 2007, 10, 335–341. [Google Scholar] [CrossRef] [Green Version]
- Boller, T.; Felix, G. A Renaissance of Elicitors: Perception of Microbe-Associated Molecular Patterns and Danger Signals by Pattern-Recognition Receptors. Annu. Rev. Plant Biol. 2009, 60, 379–406. [Google Scholar] [CrossRef]
- Jones, J.D.G.; Dangl, J.L. The plant immune system. Nature 2006, 444, 323–329. [Google Scholar] [CrossRef] [Green Version]
- Abramovitch, R.; Janjusevic, R.; Stebbins, E.; Martin, G.B. Type III effector AvrPtoB requires intrinsic E3 ubiquitin ligase activity to suppress plant cell death and immunity. Proc. Natl. Acad. Sci. USA 2006, 103, 2851–2856. [Google Scholar] [CrossRef] [Green Version]
- Cui, H.; Wang, Y.; Xue, L.; Chu, J.; Yan, C.; Fu, J.; Chen, M.; Innes, R.W.; Zhou, J.-M. Pseudomonas syringae Effector Protein AvrB Perturbs Arabidopsis Hormone Signaling by Activating MAP Kinase 4. Cell Host Microbe 2010, 7, 164–175. [Google Scholar] [CrossRef] [Green Version]
- Grant, S.R.; Fisher, E.J.; Chang, J.H.; Mole, B.M.; Dangl, J.L. Subterfuge and Manipulation: Type III Effector Proteins of Phytopathogenic Bacteria. Annu. Rev. Microbiol. 2006, 60, 425–449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, J.-M.; Chai, J. Plant pathogenic bacterial type III effectors subdue host responses. Curr. Opin. Microbiol. 2008, 11, 179–185. [Google Scholar] [CrossRef] [PubMed]
- Denoux, C.; Galletti, R.; Mammarella, N.; Gopalan, S.; Werck, D.; De Lorenzo, G.; Ferrari, S.; Ausubel, F.M.; Dewdney, J. Activation of Defense Response Pathways by OGs and Flg22 Elicitors in Arabidopsis Seedlings. Mol. Plant 2008, 1, 423–445. [Google Scholar] [CrossRef] [Green Version]
- Lotze, M.T.; Zeh, H.J.; Rubartelli, A.; Sparvero, L.J.; Amoscato, A.; Washburn, N.R.; Devera, M.E.; Liang, X.; Tor, M.; Billiar, T. The grateful dead: Damage-associated molecular pattern molecules and reduction/oxidation regulate immunity. Immunol. Rev. 2007, 220, 60–81. [Google Scholar] [CrossRef] [PubMed]
- Thomma, B.P.H.J.; Nürnberger, T.; Joosten, M.H.A.J. Of PAMPs and Effectors: The Blurred PTI-ETI Dichotomy. Plant Cell 2011, 23, 4–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nicaise, V.; Roux, M.; Zipfel, C. Recent Advances in PAMP-Triggered Immunity against Bacteria: Pattern Recognition Receptors Watch over and Raise the Alarm. Plant Physiol. 2009, 150, 1638–1647. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Loon, L.; Rep, M.; Pieterse, C. Significance of Inducible Defense-related Proteins in Infected Plants. Annu. Rev. Phytopathol. 2006, 44, 135–162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bent, A.F.; Mackey, D. Elicitors, Effectors, and R Genes: The New Paradigm and a Lifetime Supply of Questions. Annu. Rev. Phytopathol. 2007, 45, 399–436. [Google Scholar] [CrossRef] [Green Version]
- Zipfel, C. Early molecular events in PAMP-triggered immunity. Curr. Opin. Plant Biol. 2009, 12, 414–420. [Google Scholar] [CrossRef]
- Baysal, Ö.; Gürsoy, Y.Z.; Ornek, H.; Duru, A. Enhanced tomato resistance to bacterial canker by application of turtle oil. J. Gen. Plant Pathol. 2005, 71, 204–210. [Google Scholar] [CrossRef]
- Choudhary, D.K.; Prakash, A.; Johri, B.N. Induced systemic resistance (ISR) in plants: Mechanism of action. Indian J. Microbiol. 2007, 47, 289–297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Durrant, W.E.; Dong, X. Systemic acquired resistance. Annu. Rev. Phytopathol. 2004, 42, 185–209. [Google Scholar] [CrossRef] [PubMed]
- Ryals, J.A.; Neuenschwander, U.H.; Willits, M.G.; Molina, A.; Steiner, H.-Y.; Hunt, M.D. Systemic acquired resistance. Plant Cell 1996, 8, 1809. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sticher, L.; Mauch-Mani, B.; Métraux, J.P. Systemic acquired resistance. Annu. Rev. Phytopathol. 1997, 35, 235–270. [Google Scholar] [CrossRef]
- Zhang, Y.; Xu, S.; Ding, P.; Wang, D.; Cheng, Y.T.; He, J.; Gao, M.; Xu, F.; Li, Y.; Zhu, Z.; et al. Control of salicylic acid synthesis and systemic acquired resistance by two members of a plant-specific family of transcription factors. Proc. Natl. Acad. Sci. USA 2010, 107, 18220–18225. [Google Scholar] [CrossRef] [Green Version]
- David, L.; Kang, J.N.; Dufresne, D.; Zhu, D.; Chen, S.X. Multi-Omics Revealed Molecular Mechanisms Underlying Guard Cell Systemic Acquired Resistance. Int. J. Mol. Sci. 2021, 22, 191. [Google Scholar] [CrossRef]
- Wang, C.; El-Shetehy, M.; Shine, M.B.; Yu, K.; Navarre, D.; Wendehenne, D.; Kachroo, A.; Kachroo, P. Free radicals mediate systemic acquired resistance. Cell Rep. 2014, 7, 348–355. [Google Scholar] [CrossRef]
- Conrath, U.; Pieterse, C.M.J.; Mauch-Mani, B. Priming in plant–pathogen interactions. Trends Plant Sci. 2002, 7, 210–216. [Google Scholar] [CrossRef] [Green Version]
- Bakker, P.A.H.M.; Pieterse, C.M.J.; van Loon, L.C. Induced Systemic Resistance by Fluorescent Pseudomonas spp. Phytopathology 2007, 97, 239–243. [Google Scholar] [CrossRef] [Green Version]
- De Vleesschauwer, D.; Höfte, M. Rhizobacteria-induced systemic resistance. In Advances in Botanical Research; Academic Press: Cambridge, MA, USA, 2009; Volume 51, pp. 223–281. [Google Scholar]
- Van Loon, L. Systemic induced resistance. In Mechanisms of Resistance to Plant Diseases; Springer: Berlin/Heidelberg, Germany, 2000; pp. 521–574. [Google Scholar]
- Heil, M. Induced systemic resistance (ISR) against pathogens–a promising field for ecological research. Perspect. Plant Ecol. Evol. Syst. 2001, 4, 65–79. [Google Scholar] [CrossRef] [Green Version]
- Romera, F.J.; García, M.J.; Lucena, C.; Martinez-Medina, A.; Aparicio, M.A.; Ramos, J.; Alcántara, E.; Angulo, M.; Pérez-Vicente, R. Induced Systemic Resistance (ISR) and Fe Deficiency Responses in Dicot Plants. Front. Plant Sci. 2019, 10, 287. [Google Scholar] [CrossRef] [PubMed]
- Pieterse, C.M.; Zamioudis, C.; Berendsen, R.L.; Weller, D.M.; Van Wees, S.C.; Bakker, P.A. Induced Systemic Resistance by Beneficial Microbes. Annu. Rev. Phytopathol. 2014, 52, 347–375. [Google Scholar] [CrossRef] [Green Version]
- Dong, X. NPR1, all things considered. Curr. Opin. Plant Biol. 2004, 7, 547–552. [Google Scholar] [CrossRef] [PubMed]
- Mauch-Mani, B.; Baccelli, I.; Luna, E.; Flors, V. Defense Priming: An Adaptive Part of Induced Resistance. Annu. Rev. Plant Biol. 2017, 68, 485–512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verhagen, B.W.M.; Trotel-Aziz, P.; Couderchet, M.; Höfte, M.; Aziz, A. Pseudomonas spp.-induced systemic resistance to Botrytis cinerea is associated with induction and priming of defence responses in grapevine. J. Exp. Bot. 2009, 61, 249–260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takishita, Y.; Charron, J.-B.; Smith, D.L. Biocontrol Rhizobacterium Pseudomonas sp. 23S Induces Systemic Resistance in Tomato (Solanum lycopersicum L.) Against Bacterial Canker Clavibacter michiganensis subsp. michiganensis. Front. Microbiol. 2018, 9, 2119. [Google Scholar] [CrossRef] [PubMed]
- Siddiqui, I.A.; Shaukat, S.S. Pseudomonas aeruginosa-mediated induction of systemic resistance in tomato against root-knot nematode. Plant Pathol. J. 2005, 4, 21–25. [Google Scholar]
- Chen, C.; Bélanger, R.R.; Benhamou, N.; Paulitz, T.C. Induced systemic resistance (ISR) by Pseudomonas spp. impairs pre-and post-infection development of Pythium aphanidermatum on cucumber roots. Eur. J. Plant Pathol. 1998, 104, 877–886. [Google Scholar] [CrossRef]
- Xing, Z.; Wu, X.; Zhao, J.; Zhao, X.; Zhu, X.; Wang, Y.; Fan, H.; Chen, L.; Liu, X.; Duan, Y. Isolation and identification of induced systemic resistance determinants from Bacillus simplex Sneb545 against Heterodera glycines. Sci. Rep. 2020, 10, 11586. [Google Scholar] [CrossRef]
- Akram, W.; Anjum, T.; Ali, B. Searching ISR determinant/s from Bacillus subtilis IAGS174 against Fusarium wilt of tomato. BioControl 2014, 60, 271–280. [Google Scholar] [CrossRef]
- Nie, P.; Li, X.; Wang, S.; Guo, J.; Zhao, H.; Niu, D. Induced Systemic Resistance against Botrytis cinerea by Bacillus cereus AR156 through a JA/ET- and NPR1-Dependent Signaling Pathway and Activates PAMP-Triggered Immunity in Arabidopsis. Front. Plant Sci. 2017, 8, 238. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Medina, A.; Fernández, I.; Sánchez-Guzmán, M.J.; Jung, S.C.; Pascual, J.A.; Pozo, M.J. Deciphering the hormonal signalling network behind the systemic resistance induced by Trichoderma harzianum in tomato. Front. Plant Sci. 2013, 4, 206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tucci, M.; Ruocco, M.; De Masi, L.; De Palma, M.; Lorito, M. The beneficial effect of Trichoderma spp. on tomato is modulated by the plant genotype. Mol. Plant Pathol. 2010, 12, 341–354. [Google Scholar] [CrossRef] [PubMed]
- Alkooranee, J.T.; Aledan, T.R.; Ali, A.K.; Lu, G.; Zhang, X.; Wu, J.; Fu, C.; Li, M. Detecting the Hormonal Pathways in Oilseed Rape behind Induced Systemic Resistance by Trichoderma harzianum TH12 to Sclerotinia sclerotiorum. PLoS ONE 2017, 12, e0168850. [Google Scholar] [CrossRef] [Green Version]
- Pozo de la Hoz, J.; Rivero, J.; Azcón-Aguilar, C.; Urrestarazu, M.; Pozo, M.J. Mycorrhiza-induced resistance against foliar pathogens is uncoupled of nutritional effects under different light Intensities. J. Fungi 2021, 7, 402. [Google Scholar] [CrossRef]
- Mustafa, G.; Khong, N.G.; Tisserant, B.; Randoux, B.; Fontaine, J.; Magnin-Robert, M.; Reignault, P.; Sahraoui, A.L.-H. Defence mechanisms associated with mycorrhiza-induced resistance in wheat against powdery mildew. Funct. Plant Biol. 2017, 44, 443–454. [Google Scholar] [CrossRef] [Green Version]
- Kauss, H.; Theisinger-Hinkel, E.; Mindermann, R.; Conrath, U. Dichloroisonicotinic and salicylic acid, inducers of systemic acquired resistance, enhance fungal elicitor responses in parsley cells. Plant J. 1992, 2, 655–660. [Google Scholar] [CrossRef]
- Walters, D.; Walsh, D.; Newton, A.; Lyon, G. Induced Resistance for Plant Disease Control: Maximizing the Efficacy of Resistance Elicitors. Phytopathology 2005, 95, 1368–1373. [Google Scholar] [CrossRef] [Green Version]
- Oostendorp, M.; Kunz, W.; Dietrich, B.; Staub, T. Induced Disease Resistance in Plants by Chemicals. Eur. J. Plant Pathol. 2001, 107, 19–28. [Google Scholar] [CrossRef]
- Worrall, D.; Holroyd, G.H.; Moore, J.P.; Glowacz, M.; Croft, P.; Taylor, J.E.; Paul, N.D.; Roberts, M.R. Treating seeds with activators of plant defence generates long-lasting priming of resistance to pests and pathogens. New Phytol. 2011, 193, 770–778. [Google Scholar] [CrossRef] [Green Version]
- Sillero, J.; Rojas-Molina, M.; Avila, C.; Rubiales, D. Induction of systemic acquired resistance against rust, ascochyta blight and broomrape in faba bean by exogenous application of salicylic acid and benzothiadiazole. Crop Prot. 2012, 34, 65–69. [Google Scholar] [CrossRef] [Green Version]
- Orober, M.; Siegrist, J.; Buchenauer, H. Mechanisms of Phosphate-induced Disease Resistance in Cucumber. Eur. J. Plant Pathol. 2002, 108, 345–353. [Google Scholar] [CrossRef]
- Gottstein, H.D.; Kuc, J. Induction of systemic resistance to anthracnose in cucumber by phosphates. Phytopathology 1989, 79, 176–179. [Google Scholar] [CrossRef] [Green Version]
- Mitchell, A.F.; Walters, D.R. Potassium phosphate induces systemic protection in barley to powdery mildew infection. Pest Manag. Sci. 2004, 60, 126–134. [Google Scholar] [CrossRef] [PubMed]
- Zhou, M.; Wang, W. Recent Advances in Synthetic Chemical Inducers of Plant Immunity. Front. Plant Sci. 2018, 9, 1613. [Google Scholar] [CrossRef] [Green Version]
- Czerwoniec, P.; Lewandowski, P.; Smiglak, M. Derivatives of Isonicotinic Acid as New Efficient Systemic Acquired Resistance (SAR) Inducers. ChemistrySelect 2020, 5, 10759–10764. [Google Scholar] [CrossRef]
- Métraux, J.; Ahlgoy, P.; Staub, T.; Speich, J.; Steinemann, A.; Ryals, J.; Ward, E. Induced systemic resistance in cucumber in response to 2, 6-dichloro-isonicotinic acid and pathogens. In Advances in Molecular Genetics of Plant-Microbe Interactions; Springer: Berlin/Heidelberg, Germany, 1991; Volume 1, pp. 432–439. [Google Scholar]
- Ogawa, M.; Kadowaki, A.; Yamada, T.; Kadooka, O. Applied development of a novel fungicide isotianil (STOUT). Sumitomo Kagaku 2011, 1, 1–15. [Google Scholar]
- Krämer, W.; Schirmer, U. Modern Crop Protection Compounds, 3 Volume Set; John Wiley & Sons Ltd.: Hoboken, NJ, USA, 2007. [Google Scholar]
- Ton, J.; Mauch-Mani, B. β-aminobutyric acid induced resistance against necrotrophic pathogens is based on ABA-dependent priming for callose. Plant J. 2004, 38, 119–130. [Google Scholar] [CrossRef]
- Zhang, C.; Wang, W.; Zhao, X.; Wang, H.; Yin, H. Preparation of alginate oligosaccharides and their biological activities in plants: A review. Carbohydr. Res. 2020, 494, 108056. [Google Scholar] [CrossRef]
- Macho, A.P.; Zipfel, C. Plant PRRs and the Activation of Innate Immune Signaling. Mol. Cell 2014, 54, 263–272. [Google Scholar] [CrossRef] [Green Version]
- Rachidi, F.; Benhima, R.; Kasmi, Y.; Sbabou, L.; El Arroussi, H. Evaluation of microalgae polysaccharides as biostimulants of tomato plant defense using metabolomics and biochemical approaches. Sci. Rep. 2021, 11, 930. [Google Scholar] [CrossRef] [PubMed]
- Yakhin, O.I.; Lubyanov, A.A.; Yakhin, I.A.; Brown, P.H. Biostimulants in Plant Science: A Global Perspective. Front. Plant Sci. 2017, 7, 2049. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wiesel, L.; Newton, A.C.; Elliott, I.; Booty, D.; Gilroy, E.M.; Birch, P.R.J.; Hein, I. Molecular effects of resistance elicitors from biological origin and their potential for crop protection. Front. Plant Sci. 2014, 5, 655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kocira, S.; Szparaga, A.; Kocira, A.; Czerwińska, E.; Wójtowicz, A.; Bronowicka-Mielniczuk, U.; Koszel, M.; Findura, P. Modeling Biometric Traits, Yield and Nutritional and Antioxidant Properties of Seeds of Three Soybean Cultivars Through the Application of Biostimulant Containing Seaweed and Amino Acids. Front. Plant Sci. 2018, 9, 388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stadnik, M.J.; Freitas, M.B.d. Algal polysaccharides as source of plant resistance inducers. Trop. Plant Pathol. 2014, 39, 111–118. [Google Scholar] [CrossRef] [Green Version]
- Pettongkhao, S.; Bilanglod, A.; Khompatara, K.; Churngchow, N. Sulphated Polysaccharide from Acanthophora spicifera Induced Hevea brasiliensis Defense Responses Against Phytophthora palmivora Infection. Plants 2019, 8, 73. [Google Scholar] [CrossRef] [Green Version]
- Cluzet, S.; Torregrosa, C.; Jacquet, C.; Lafitte, C.; Fournier, J.; Mercier, L.; Salamagne, S.; Briand, X.; EsquerrÉ-TugayÉ, M.T.; Dumas, B. Gene expression profiling and protection of Medicago truncatula against a fungal infection in response to an elicitor from green algae Ulva spp. Plant Cell Environ. 2004, 27, 917–928. [Google Scholar] [CrossRef]
- Scala, V.; Pietricola, C.; Farina, V.; Beccaccioli, M.; Zjalic, S.; Quaranta, F.; Fornara, M.; Zaccaria, M.; Momeni, B.; Reverberi, M.; et al. Tramesan Elicits Durum Wheat Defense against the Septoria Disease Complex. Biomolecules 2020, 10, 608. [Google Scholar] [CrossRef] [Green Version]
- Riseh, R.S.; Skorik, Y.A.; Thakur, V.K.; Pour, M.M.; Tamanadar, E.; Noghabi, S.S. Encapsulation of Plant Biocontrol Bacteria with Alginate as a Main Polymer Material. Int. J. Mol. Sci. 2021, 22, 11165. [Google Scholar] [CrossRef]
- Riseh, R.S.; Ebrahimi-Zarandi, M.; Vazvani, M.G.; Skorik, Y.A. Reducing Drought Stress in Plants by Encapsulating Plant Growth-Promoting Bacteria with Polysaccharides. Int. J. Mol. Sci. 2021, 22, 12979. [Google Scholar] [CrossRef]
- de Moura, F.A.; Macagnan, F.T.; da Silva, L.P. Oligosaccharide production by hydrolysis of polysaccharides: A review. Int. J. Food Sci. Technol. 2015, 50, 275–281. [Google Scholar] [CrossRef]
- Klarzynski, O.; Plesse, B.; Joubert, J.-M.; Yvin, J.-C.; Kopp, M.; Kloareg, B.; Fritig, B. Linear β-1,3 Glucans Are Elicitors of Defense Responses in Tobacco. Plant Physiol. 2000, 124, 1027–1038. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mani, S.D.; Nagarathnam, R. Sulfated polysaccharide from Kappaphycus alvarezii (Doty) Doty ex P.C. Silva primes defense responses against anthracnose disease of Capsicum annuum Linn. Algal Res. 2018, 32, 121–130. [Google Scholar] [CrossRef]
- Abasalizadeh, F.; Moghaddam, S.V.; Alizadeh, E.; Akbari, E.; Kashani, E.; Fazljou, S.M.B.; Torbati, M.; Akbarzadeh, A. Alginate-based hydrogels as drug delivery vehicles in cancer treatment and their applications in wound dressing and 3D bioprinting. J. Biol. Eng. 2020, 14, 8, Correction in 2020, 14, 17. [Google Scholar] [CrossRef] [PubMed]
- Venegas, M.; Matsuhiro, B.; Edding, M.E. Alginate Composition of Lessonia trabeculata (Phaeophyta: Laminariales) Growing in Exposed and Sheltered Habitats. Bot. Mar. 1993, 36, 47–51. [Google Scholar] [CrossRef]
- Hu, X.; Gao, C. Photoinitiating polymerization to prepare biocompatible chitosan hydrogels. J. Appl. Polym. Sci. 2008, 110, 1059–1067. [Google Scholar] [CrossRef]
- Ifkovits, J.L.; Burdick, J.A. Review: Photopolymerizable and Degradable Biomaterials for Tissue Engineering Applications. Tissue Eng. 2007, 13, 2369–2385. [Google Scholar] [CrossRef]
- Varghese, S.; Hwang, N.S.; Canver, A.C.; Theprungsirikul, P.; Lin, D.W.; Elisseeff, J. Chondroitin sulfate based niches for chondrogenic differentiation of mesenchymal stem cells. Matrix Biol. 2008, 27, 12–21. [Google Scholar] [CrossRef]
- Garagorri, N.; Fermanian, S.; Thibault, R.; Ambrose, W.M.; Schein, O.D.; Chakravarti, S.; Elisseeff, J. Keratocyte behavior in three-dimensional photopolymerizable poly(ethylene glycol) hydrogels. Acta Biomater. 2008, 4, 1139–1147. [Google Scholar] [CrossRef] [Green Version]
- Goh, C.H.; Heng, P.W.S.; Chan, L.W. Alginates as a useful natural polymer for microencapsulation and therapeutic applications. Carbohydr. Polym. 2012, 88, 1–12. [Google Scholar] [CrossRef]
- González, A.; Castro, J.; Vera, J.; Moenne, A. Seaweed Oligosaccharides Stimulate Plant Growth by Enhancing Carbon and Nitrogen Assimilation, Basal Metabolism, and Cell Division. J. Plant Growth Regul. 2012, 32, 443–448. [Google Scholar] [CrossRef] [Green Version]
- Salgado, L.T.; Tomazetto, R.; Cinelli, L.P.; Farina, M.; Filho, G.M.A. The influence of brown algae alginates on phenolic compounds capability of ultraviolet radiation absorption in vitro. Braz. J. Oceanogr. 2007, 55, 145–154. [Google Scholar] [CrossRef]
- Lawrence, C.B.; Singh, N.P.; Qiu, J.; Gardner, R.G.; Tuzun, S. Constitutive hydrolytic enzymes are associated with polygenic resistance of tomato to Alternaria solani and may function as an elicitor release mechanism. Physiol. Mol. Plant Pathol. 2000, 57, 211–220. [Google Scholar] [CrossRef]
- Keefe, A.J.; Jiang, S. Poly(zwitterionic)protein conjugates offer increased stability without sacrificing binding affinity or bioactivity. Nat. Chem. 2011, 4, 59–63. [Google Scholar] [CrossRef] [PubMed]
- Roberts, M.; Bentley, M.; Harris, J. Chemistry for peptide and protein PEGylation. Adv. Drug Deliv. Rev. 2002, 54, 459–476. [Google Scholar] [CrossRef]
- Shaw, P.E. Peptidyl-prolyl isomerases: A new twist to transcription. EMBO Rep. 2002, 3, 521–526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rizhsky, L.; Liang, H.; Shuman, J.; Shulaev, V.; Davletova, S.; Mittler, R. When Defense Pathways Collide. The Response of Arabidopsis to a Combination of Drought and Heat Stress. Plant Physiol. 2004, 134, 1683–1696. [Google Scholar] [CrossRef] [Green Version]
- Apel, K.; Hirt, H. Reactive oxygen species: Metabolism, oxidative stress, and signal transduction. Annu. Rev. Plant Biol. 2004, 55, 373–399. [Google Scholar] [CrossRef] [Green Version]
- Smirnoff, N.; Arnaud, D. Hydrogen peroxide metabolism and functions in plants. New Phytol. 2018, 221, 1197–1214. [Google Scholar] [CrossRef]
- Arnaud, D.; Lee, S.; Takebayashi, Y.; Choi, D.; Choi, J.; Sakakibara, H.; Hwang, I. Cytokinin-Mediated Regulation of Reactive Oxygen Species Homeostasis Modulates Stomatal Immunity in Arabidopsis. Plant Cell 2017, 29, 543–559. [Google Scholar] [CrossRef] [Green Version]
- Ribeiro, D.M.; Desikan, R.; Bright, J.; Confraria, A.; Harrison, J.; Hancock, J.T.; Barros, R.S.; Neill, S.J.; Wilson, I.D. Differential requirement for NO during ABA-induced stomatal closure in turgid and wilted leaves. Plant Cell Environ. 2008, 32, 46–57. [Google Scholar] [CrossRef] [PubMed]
- Ramanathan, A.; Vidhasekaran, P.; Samiyappan, R. Induction of defense mechanisms in greengram leaves and suspension-cultured cells by Macrophomina phaseolina and its elicitors. J. Plant Dis. Prot. 2000, 107, 245–257. [Google Scholar]
- Aitouguinane, M.; Bouissil, S.; Mouhoub, A.; Rchid, H.; Fendri, I.; Abdelkafi, S.; El-Hadj, M.D.O.; Boual, Z.; Dubessay, P.; Gardarin, C.; et al. Induction of Natural Defenses in Tomato Seedlings by Using Alginate and Oligoalginates Derivatives Extracted from Moroccan Brown Algae. Mar. Drugs 2020, 18, 521. [Google Scholar] [CrossRef] [PubMed]
- Xiang, S.; Lv, X.; He, L.; Shi, H.; Liao, S.; Liu, C.; Huang, Q.; Li, X.; He, X.; Chen, H.; et al. Dual-Action Pesticide Carrier That Continuously Induces Plant Resistance, Enhances Plant Anti-Tobacco Mosaic Virus Activity, and Promotes Plant Growth. J. Agric. Food Chem. 2019, 67, 10000–10009. [Google Scholar] [CrossRef] [PubMed]
- Sano, Y. Antiviral activity of alginate against infection by tobacco mosaic virus. Carbohydr. Polym. 1999, 38, 183–186. [Google Scholar] [CrossRef]
- Zhuo, R.; Li, B.; Tian, S. Alginate oligosaccharide improves resistance to postharvest decay and quality in kiwifruit (Actinidia deliciosa cv. Bruno). Hortic. Plant J. 2021, 8, 44–52. [Google Scholar] [CrossRef]
- Bouissil, S.; Guérin, C.; Roche, J.; Dubessay, P.; El Alaoui-Talibi, Z.; Pierre, G.; Michaud, P.; Mouzeyar, S.; Delattre, C.; El Modafar, C. Induction of defense gene expression and the resistance of date palm to Fusarium oxysporum f.sp. albedinis in response to alginate extracted from Bifurcaria bifurcata. Mar. Drugs 2022, 20, 88. [Google Scholar] [CrossRef]
- Ben Salah, I.; Aghrouss, S.; Douira, A.; Aissam, S.; El Alaoui-Talibi, Z.; Filali-Maltouf, A.; El Modafar, C. Seaweed polysaccharides as bio-elicitors of natural defenses in olive trees against verticillium wilt of olive. J. Plant Interact. 2018, 13, 248–255. [Google Scholar] [CrossRef]
- An, Q.D.; Zhang, G.L.; Wu, H.T.; Zhang, Z.C.; Zheng, G.S.; Luan, L.; Murata, Y.; Li, X. Alginate-deriving oligosaccharide production by alginase from newly isolated Flavobacterium sp. LXA and its potential application in protection against pathogens. J. Appl. Microbiol. 2009, 106, 161–170. [Google Scholar] [CrossRef]
- Han, J.; Zhao, L.; Zhu, H.; Dhanasekaran, S.; Zhang, X.; Zhang, H. Study on the effect of alginate oligosaccharide combined with Meyerozyma guilliermondii against Penicillium expansum in pears and the possible mechanisms involved. Physiol. Mol. Plant Pathol. 2021, 115, 101654. [Google Scholar] [CrossRef]
- Liu, H.; Zhang, Y.-H.; Yin, H.; Wang, W.-X.; Zhao, X.-M.; Du, Y.-G. Alginate oligosaccharides enhanced Triticum aestivum L. tolerance to drought stress. Plant Physiol. Biochem. 2013, 62, 33–40. [Google Scholar] [CrossRef] [PubMed]
- Tang, J.; Zhou, Q.; Chu, H.; Nagata, S. Characterization of Alginase and Elicitor-Active Oligosaccharides from Gracilibacillus A7 in Alleviating Salt Stress for Brassica campestris L. J. Agric. Food Chem. 2011, 59, 7896–7901. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.J.; Li, X.M.; Bu, N.; Li, N. An alginate-derived oligosaccharide enhanced wheat tolerance to cadmium stress. Plant Growth Regul. 2010, 62, 71–76. [Google Scholar] [CrossRef]





{kind=link}
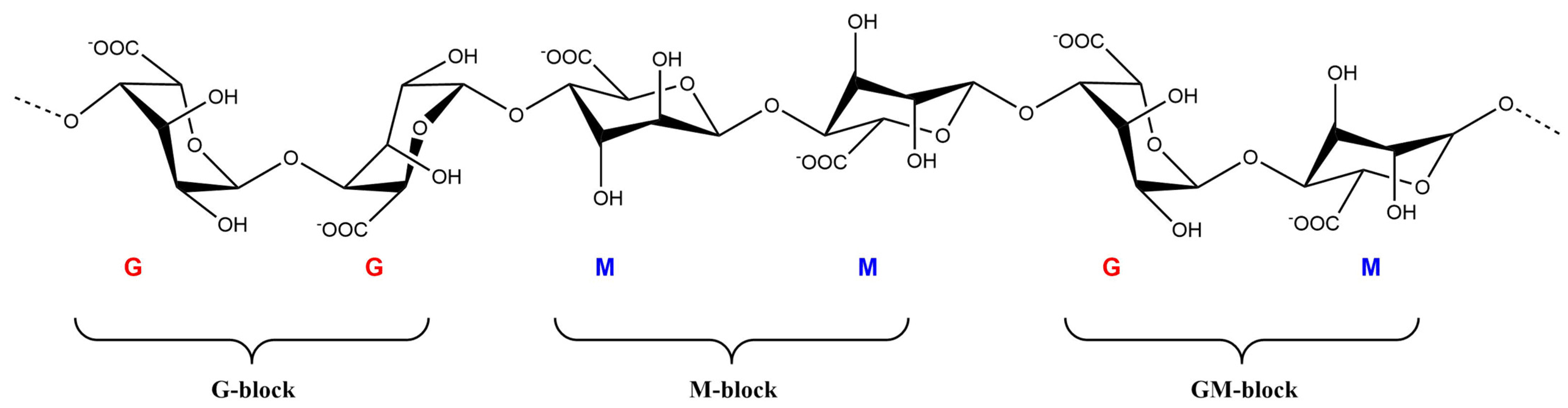{kind=link}
{kind=link}
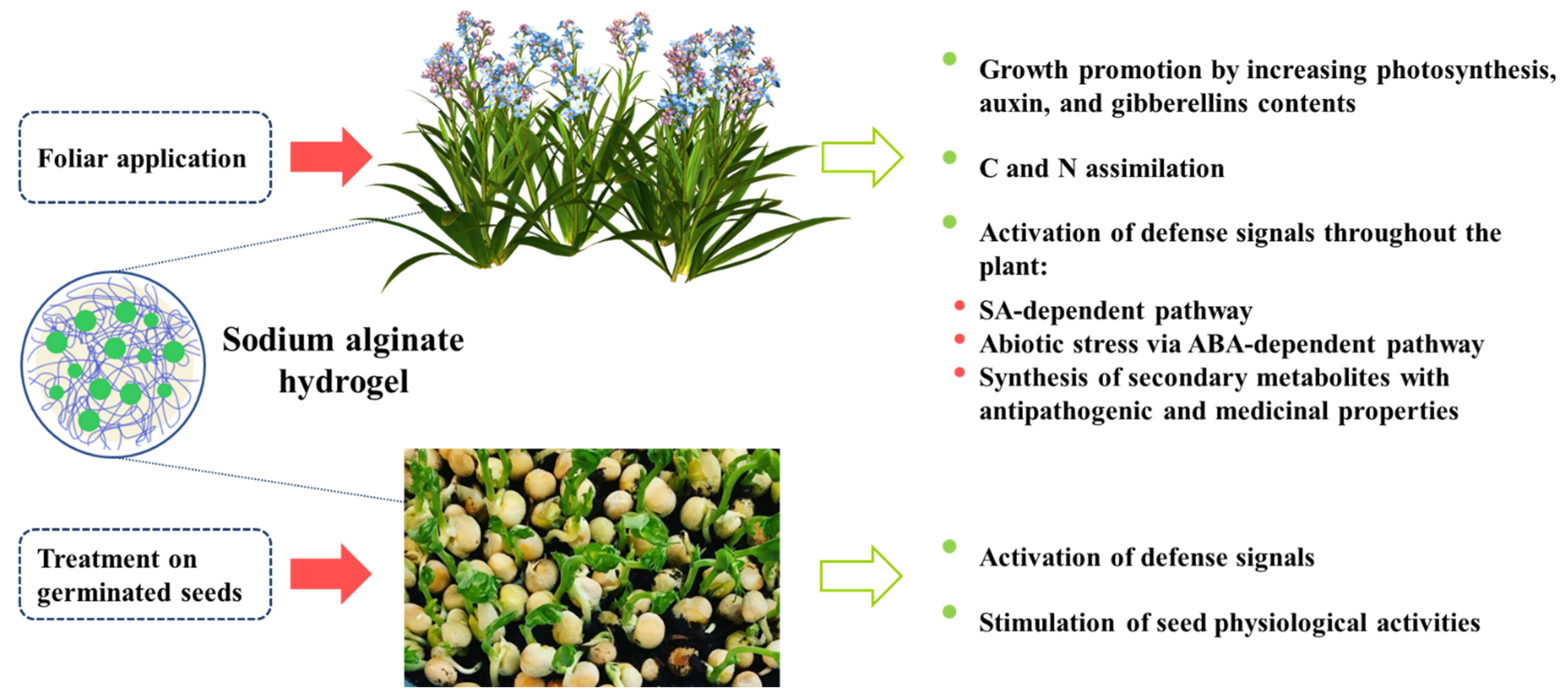{kind=link}
| Biological Inducer | Pathogens | Host | Mechanism | Reference |
|---|---|---|---|---|
| Pseudomonas spp. | Botrytis cinerea | Grapevine | Oxidative burst and phytoalexin accumulation in grape cells and leaves. | [61] |
| Clavibacter michiganensis | Tomato | Increase in levels of PR1a and ACO transcripts and SA signaling pathways. | [62] | |
| Meloidogyne spp. | Tomato | SA production by bacteria. | [63] | |
| Pythium aphanidermatum | Cucumber | Reduced pathogen spread. | [64] | |
| Bacillus spp. | Heterodera glycines | Soybean | Expression of defense-related genes involved in the SA and JA pathways. | [65] |
| Fusarium sp. | Tomato | Production of phthalic acid methyl ester by Bacillus. | [66] | |
| Botrytis cinerea | Arabidopsis | Activation of the JA–ET signaling pathway. | [67] | |
| Trichoderma spp. | Botrytis cinerea | Tomato | Activation of the JA, SA, and ABA signaling pathways. | [68] |
| Enhanced activation of jasmonate-responsive genes. | [69] | |||
| Sclerotinia sclerotiorum | Brassica napus | Induction of SA- and JA–ET-dependent defenses and decreased disease symptoms. | [70] | |
| Mycorrhizal fungi | Botrytis cinerea | Lettuce | Provision of biotic stress protection with no nutritional or growth benefits. | [71] |
| Blumeria graminis f.sp. tritici | Wheat | Accumulation of phenolic compounds and H2O2, upregulation of genes encoding several defense markers (POD, PAL, chitinase 1) | [72] |
| Abiotic Component | Pathogen/Plant Disease | Type of Plant | Mechanism | Reference |
|---|---|---|---|---|
| Dibasic and tribasic phosphate salts | Colletotrichum lagenarium | Cucumber | Influences the activity of apoplastic enzymes, such as polygalacturonases, thereby releasing elicitor-active oligogalacturonides from plant cell walls. | [78,79] |
| Preceded by a rapid generation of superoxide and hydrogen peroxide. | ||||
| Blumeria graminis f.sp. hordei | Barley | Reduces powdery mildew infection by 89%. | [80] | |
| SA Derivatives | TMV | Tomato Tobacco | Establishes plant immunity by an accumulation of PR proteins. | [81] |
| Isonicotinic acid derivatives | TMV | Tobacco | Decreases the necrotic area on leaves. | [82] |
| Colletotrichum lagenarium | Cucumber | Induces chitinase and modifies the physiology of the host. | [83] | |
| Thiadiazole and isothiazole derivative | Powdery mildew, anthracnose, and bacterial leaf spot Alternaria leaf spot, anthracnose, bacterial shot hole | Wheat | Promotes the expression of defense-related genes and SA catabolism. Induces plant defense responses. | [84] [85] |
| Pumpkin | ||||
| Cucumber | ||||
| Chinese cabbage | ||||
| Strawberry | ||||
| Peach | ||||
| β-Aminobutyric acid | Alternaria brassicicola, Plectosphaerella cucumerina | Arabidopsis | Promotes callose accumulation by an ABA-dependent defense pathway. | [86] |
| ALG Concentration | Pathogen | Plant | Mechanism | Reference |
|---|---|---|---|---|
| 5 g/L | Tobacco mosaic virus (TMV) | Tobacco (on leaves) | The antiviral activity of ALG on infectivity of TMV on blocking the decapsulation process of TMV protein on the cell membrane surface. | [123] |
| 50 g/L | Botrytis cinerea | Kiwifruit (on fruit) | Reduction in the incidence of gray mold and diameter of lesions of kiwifruit during storage; enhancing the activity of polyphenol oxidase, l-phenylalanine ammonia-lyase (PAL), and β-1,3-glucanase related to pathogen defense. | [124] |
| 1 g/L | Fusarium oxysporum f.sp. albedinis | Date Palm (on roots) | The stimulation of PAL activity in roost; the increased transcriptional level; stimulates expression of the genes involved in phenolic metabolism and burst oxidation. | [125] |
| 2 g/L | Verticillium dahliae | Olive (on twigs of 10 cm in length with 16 leaves) | Increase in the enzymatic activity of PAL in the stem; inhibitory rates on mycelial growth of the fungus in vitro. | [126] |
| 0.3 g/L | Erwinia carotovora Xanthomonas campestris | soybean cotyledon | The accumulation of phytoalexin and inducing PAL in soybean cotyledon. | [127] |
| 5 g/L AOS combined with Meyerozyma guilliermondii | Penicillium expansum | Pears (on Fruits) | Increase in the activities of superoxide dismutase (SOD), catalase (CAT), polyphenol oxidase (PPO), peroxidase. (POD), phenylalanine ammonia-lyase (PAL), chitinase (CHI), total phenol content, and flavonoid content in pears; reduce spore germination rate and inhibit the germ tube elongation of P. expansum. | [128] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Saberi Riseh, R.; Gholizadeh Vazvani, M.; Ebrahimi-Zarandi, M.; Skorik, Y.A. Alginate-Induced Disease Resistance in Plants. Polymers 2022, 14, 661. https://doi.org/10.3390/polym14040661
Saberi Riseh R, Gholizadeh Vazvani M, Ebrahimi-Zarandi M, Skorik YA. Alginate-Induced Disease Resistance in Plants. Polymers. 2022; 14(4):661. https://doi.org/10.3390/polym14040661
Chicago/Turabian StyleSaberi Riseh, Roohallah, Mozhgan Gholizadeh Vazvani, Marzieh Ebrahimi-Zarandi, and Yury A. Skorik. 2022. "Alginate-Induced Disease Resistance in Plants" Polymers 14, no. 4: 661. https://doi.org/10.3390/polym14040661
APA StyleSaberi Riseh, R., Gholizadeh Vazvani, M., Ebrahimi-Zarandi, M., & Skorik, Y. A. (2022). Alginate-Induced Disease Resistance in Plants. Polymers, 14(4), 661. https://doi.org/10.3390/polym14040661