
Synergistic Effect of Zinc-Chitosan Nanoparticles and Hydroxychloroquine to Inhibit Buffalo Coronavirus
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Synthesis of Hydroxychloroquine/zinc Oxide Nanoparticles Loaded Chitosan Composites
2.3. Morphology, Particle Size, and Surface Charge Determination (ζ)
2.4. Estimation of Zinc Concentration in Formulations
2.5. Fourier Transform Infrared Spectroscopy (FTIR)
2.6. Cytotoxicity of Nanoformulations
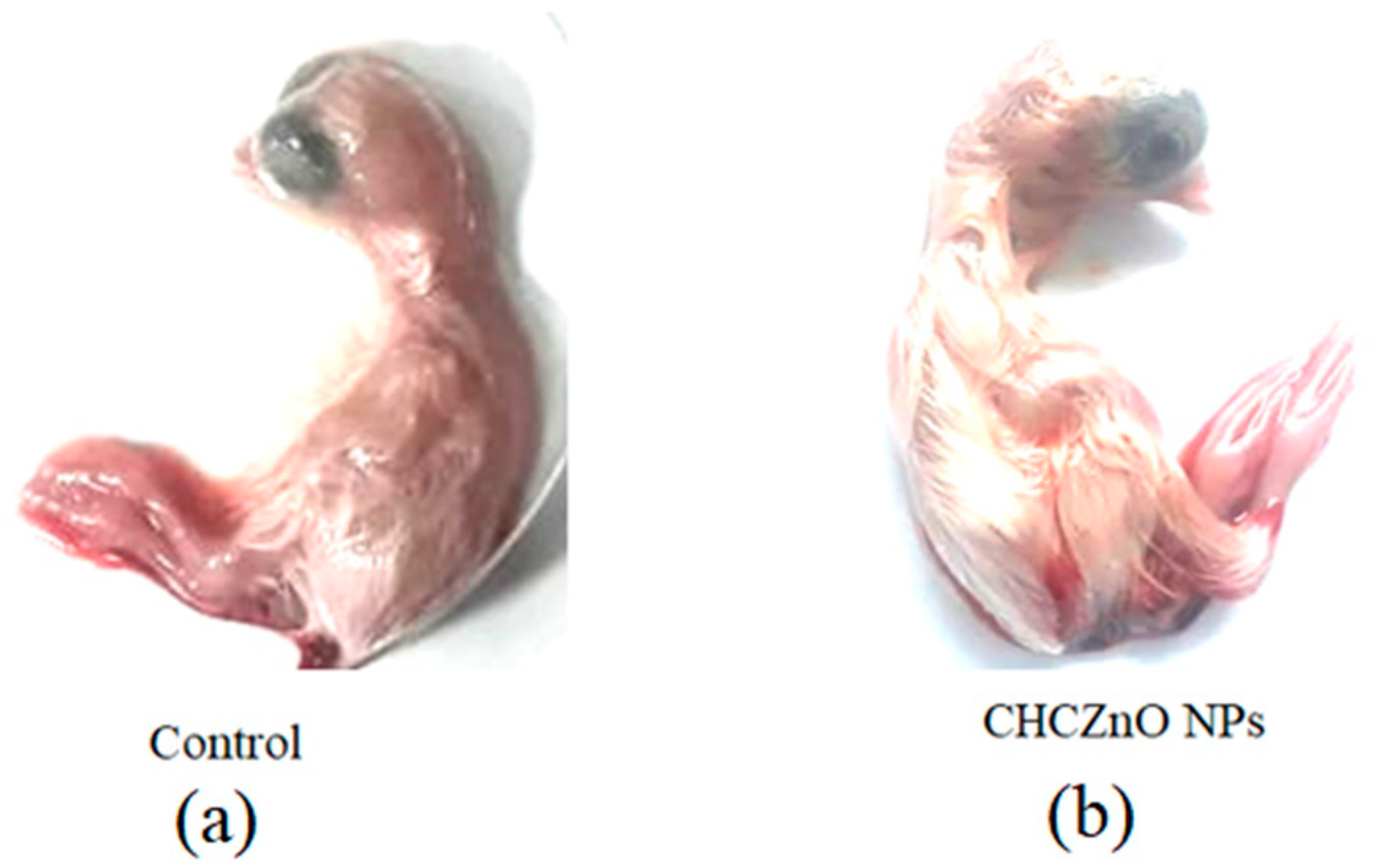
2.7. Biocompatibility in Embryonated Chicken Eggs
2.8. Virus Propagation in African Green Monkey Kidney Cells (Vero Cells)
2.9. RT-PCR
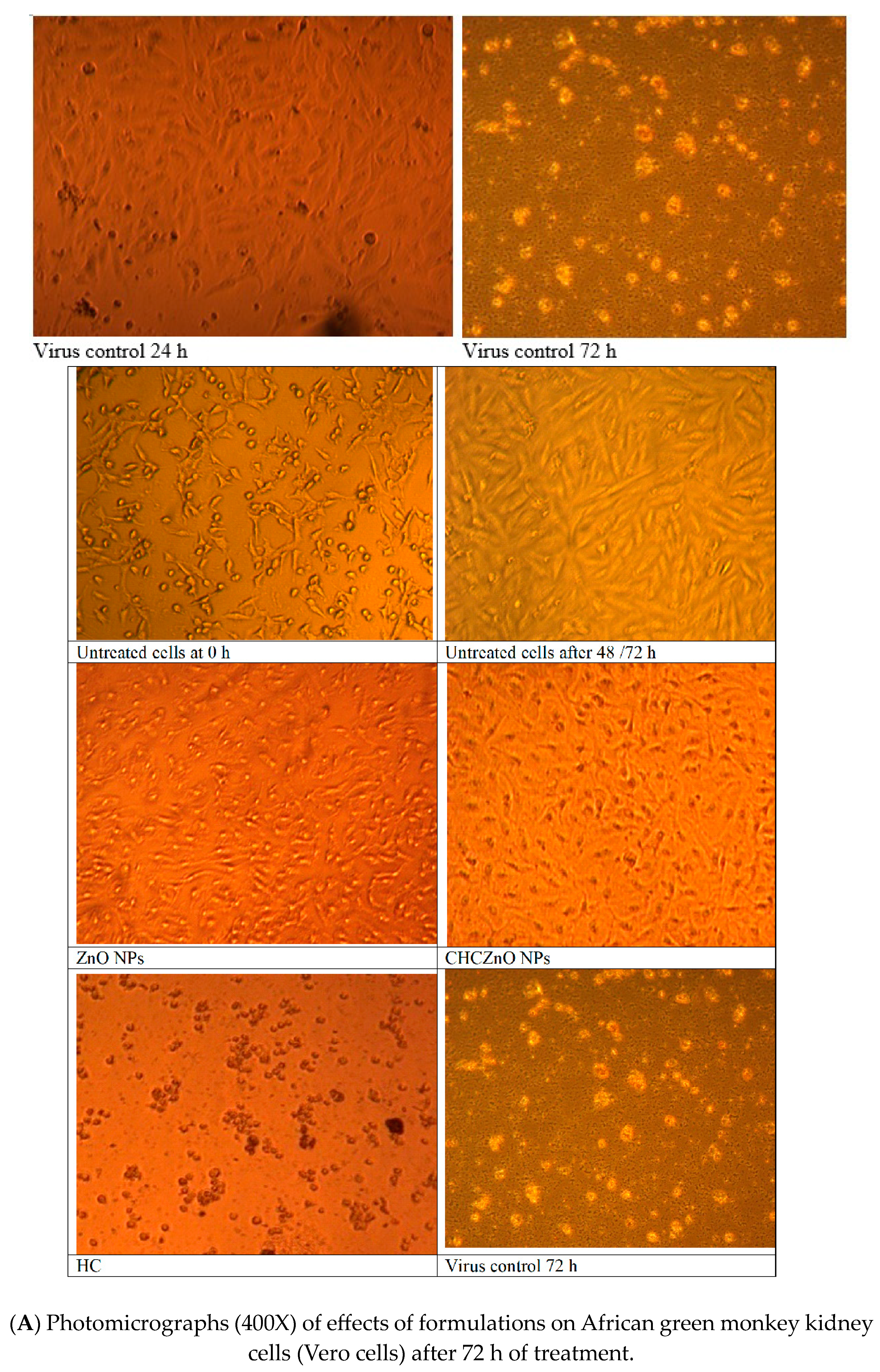
2.10. Effects of Formulations on African Green Monkey Kidney Cells (Vero Cells)
2.11. Real-Time Quantitative PCR
2.12. Intra-Cellular Zinc Uptake
2.13. Statistical Analysis
3. Results and Discussion
3.1. Characterization/Description
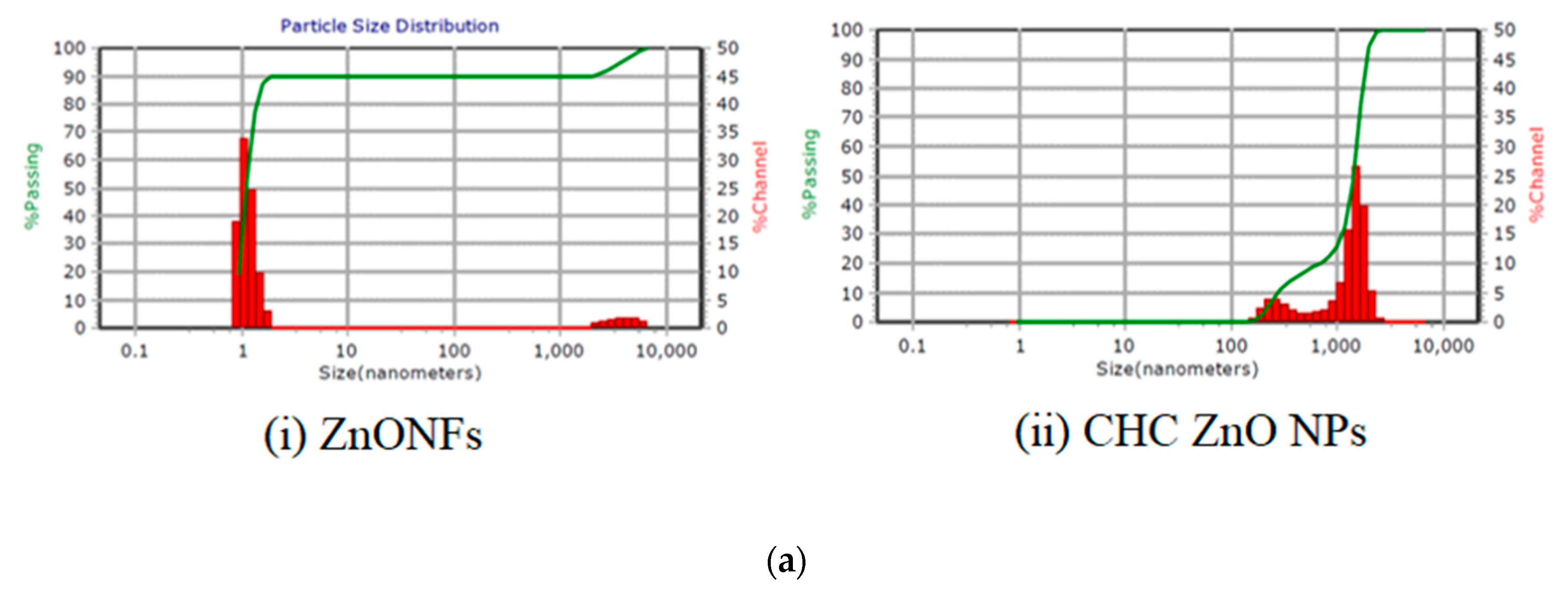
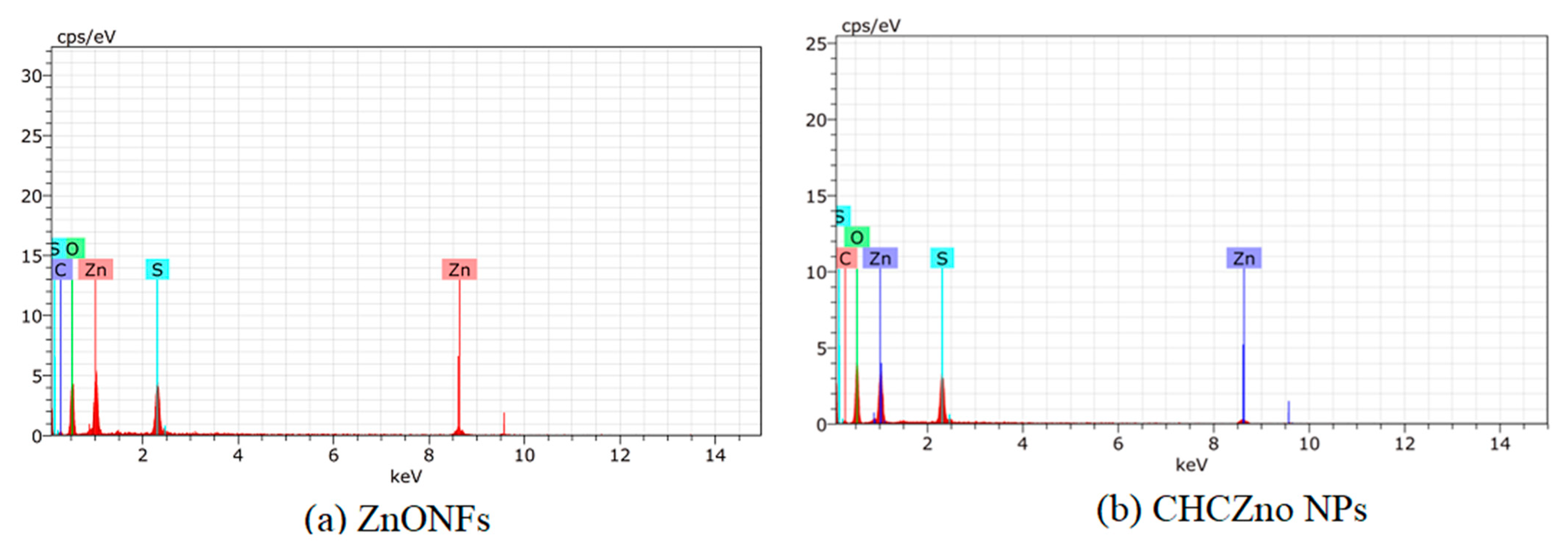
3.1.1. Size, Morphology, and Charge
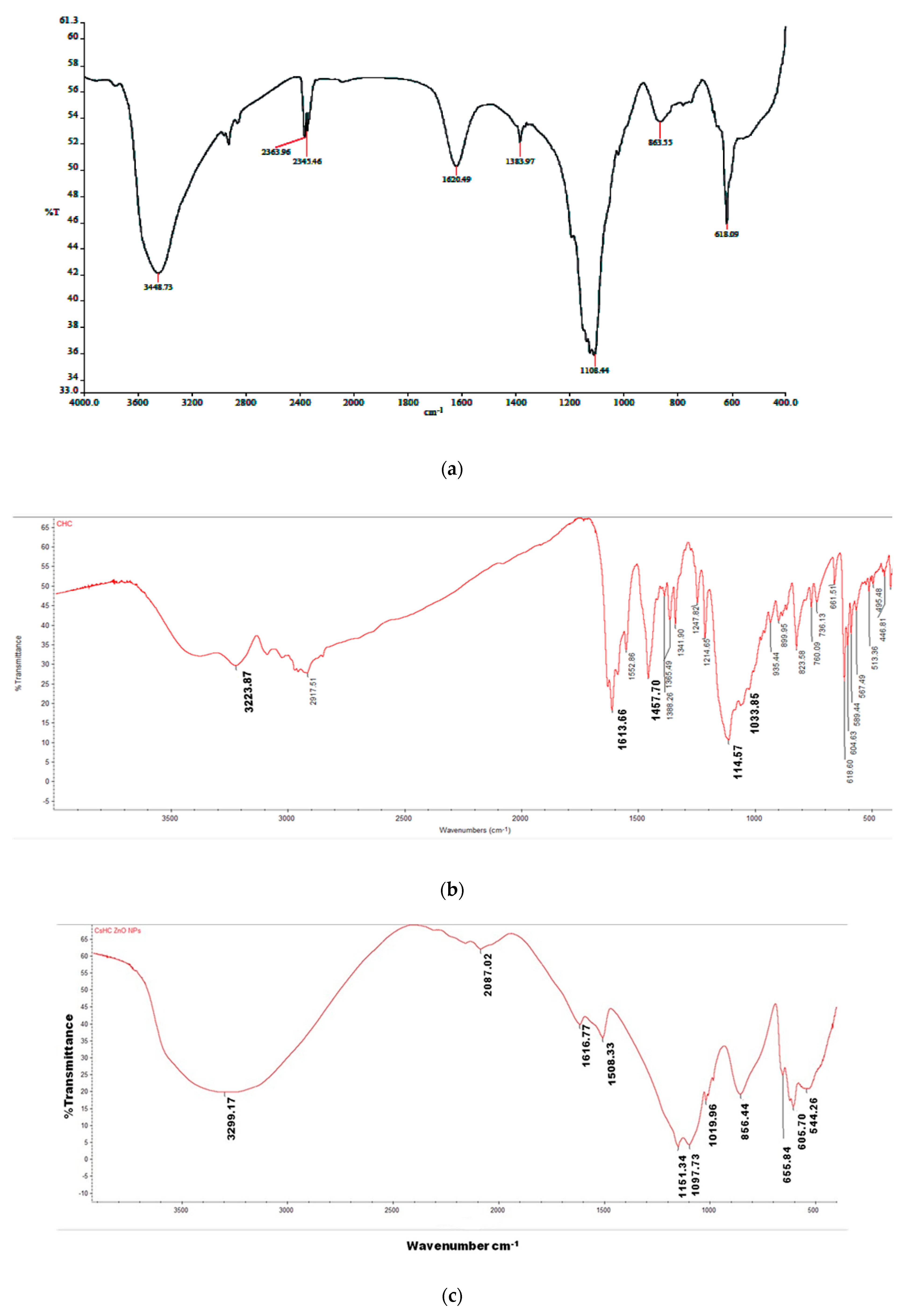
3.1.2. Fourier Transform Infrared Spectroscopy
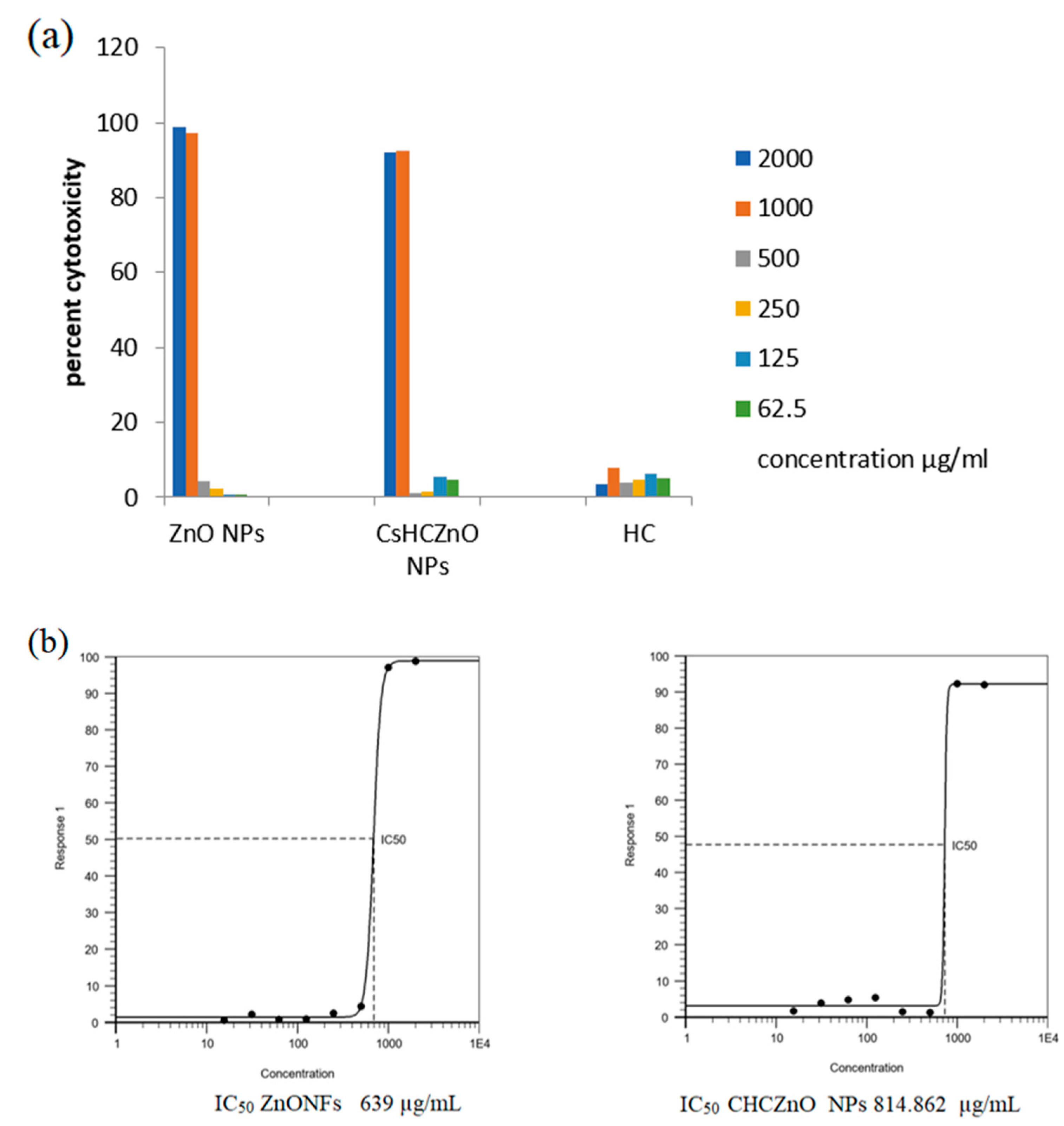
3.2. Cytotoxicity Studies on African Green Monkey Kidney Cells (Vero Cells)
3.3. Biocompatibility in SPF Chick Embryos
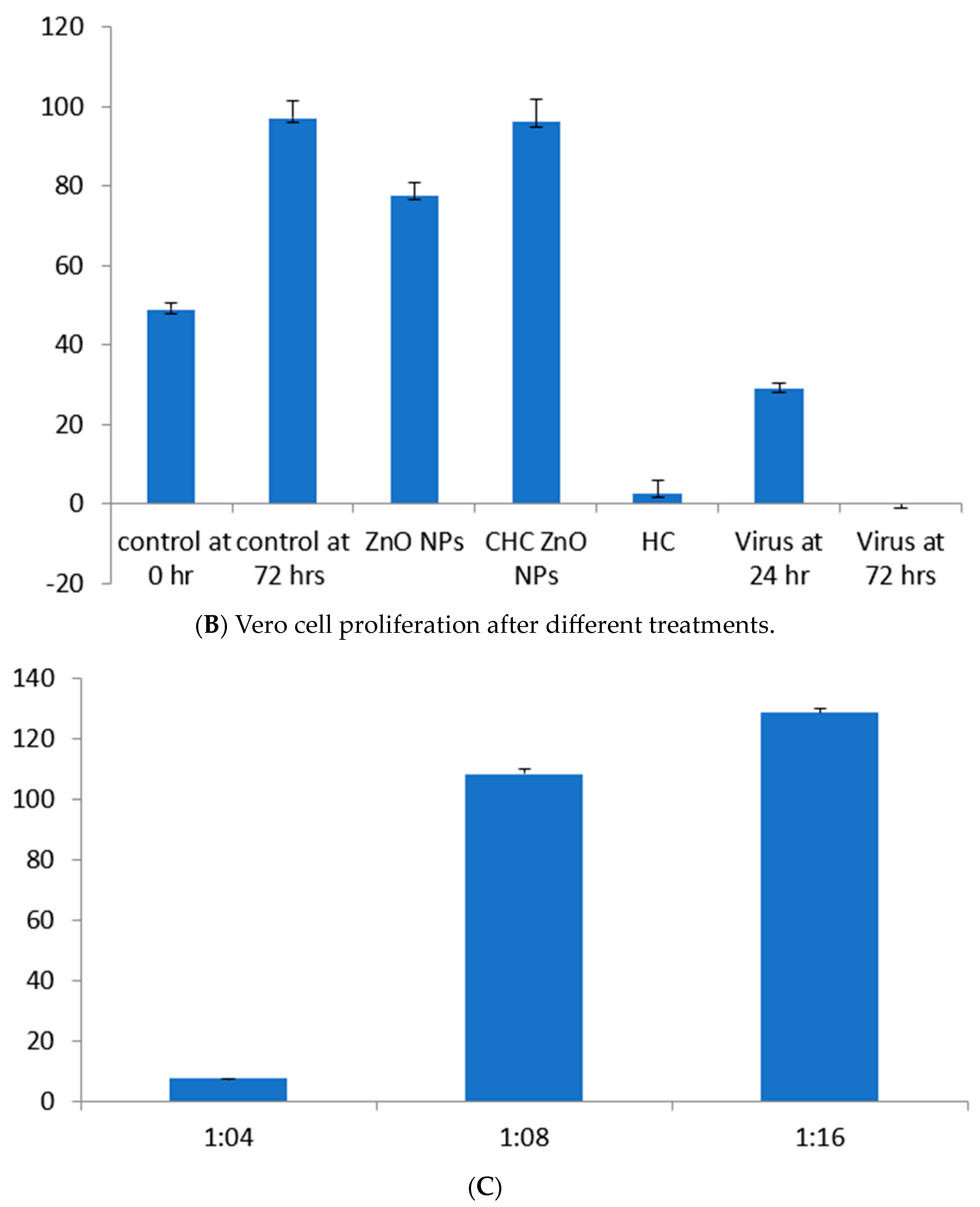
3.4. Effects of Formulations on African Green Monkey Kidney Cells (Vero Cells)
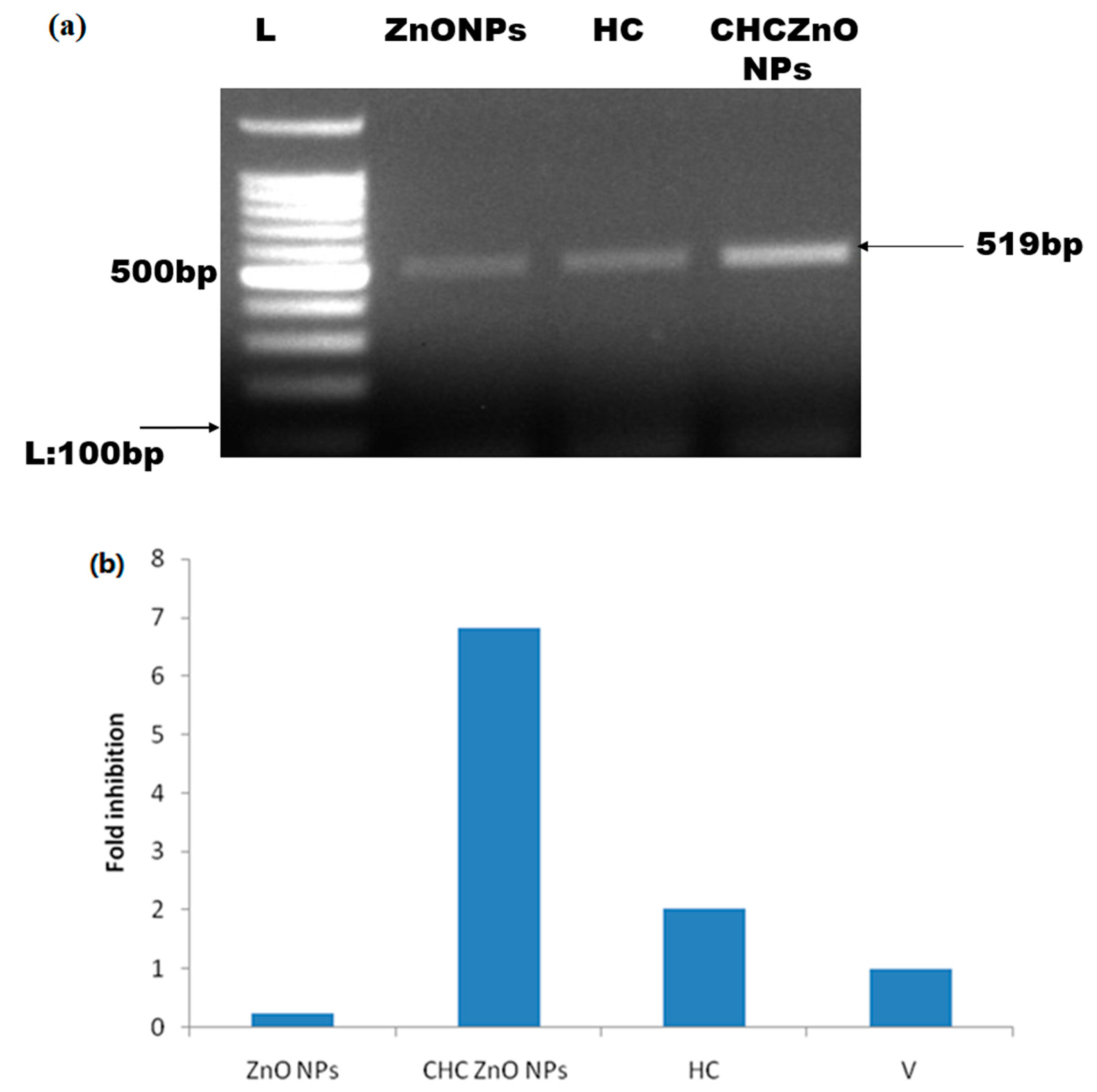
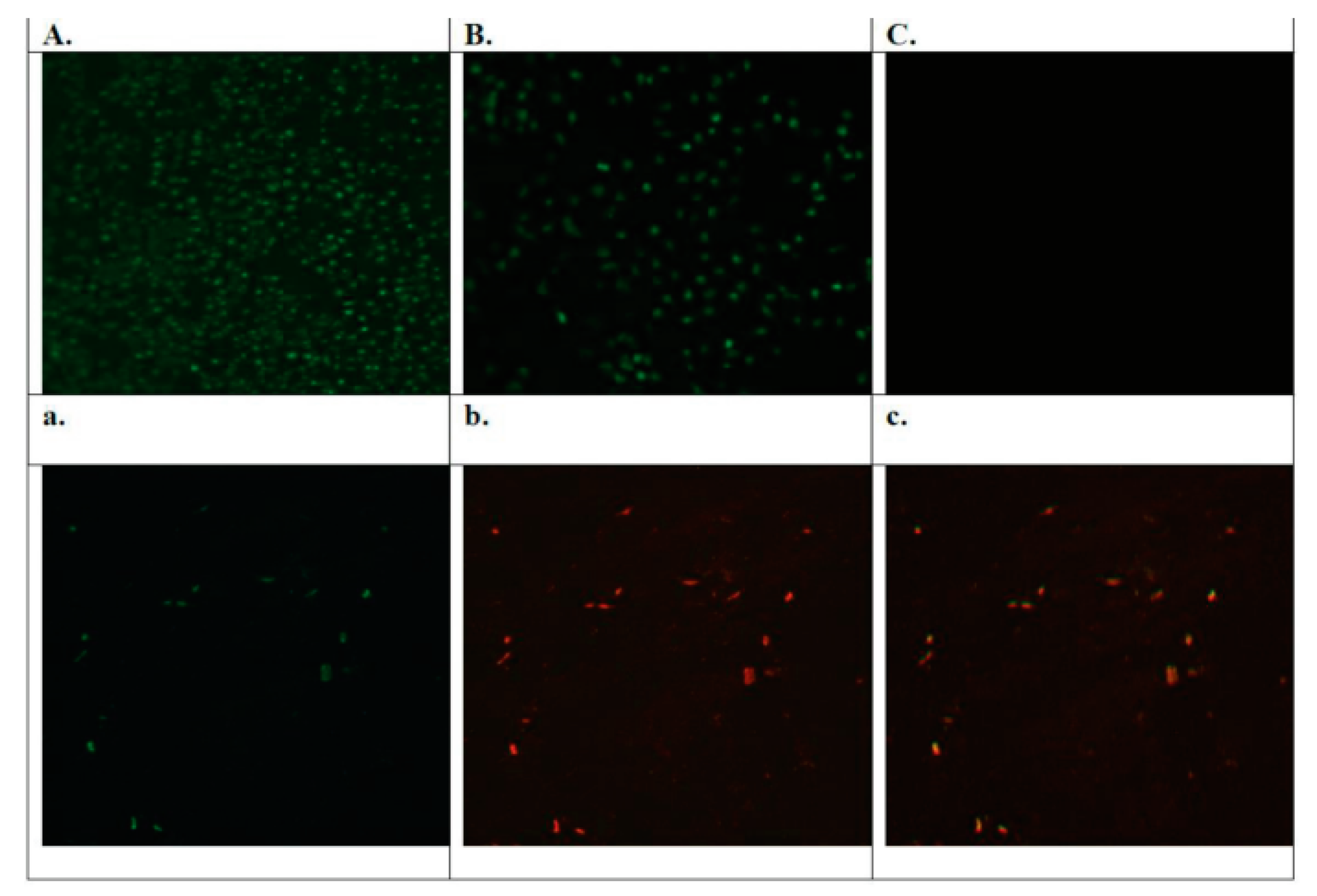
3.5. Buffalo Coronavirus-Infected Vero Cells Exhibited M Protein
3.6. Nanoparticle Uptake in Vero Cells
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Weiss, S.R.; Navas-Martin, S. Coronavirus pathogenesis and the emerging pathogen severe acute respiratory syndrome coronavirus. Microbiol. Mol. Biol. Rev. 2005, 69, 635–664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xue, J.; Moyer, A.; Peng, B.; Wu, J.; Hannafon, B.N.; Ding, W.Q. Chloroquine is a zinc ionophore. PLoS ONE 2014, 9, e109180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, J.; Tian, Z.; Yang, X. Breakthrough: Chloroquine phosphate has shown apparent efficacy in treatment of COVID-19 associated pneumonia in clinical studies. Biosci. Trends 2020, 14, 72–73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collins, K.P.; Jackson, K.M.; Gustafson, D.L. Hydroxychloroquine: A physiologically-based pharmacokinetic model in the context of cancer-related autophagy modulation. J. Pharmacol. Exp. Therap. 2018, 365, 447–459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsai, W.P.; Nara, P.L.; Kung, H.F.; Oroszlan, S. Inhibition of human immunodeficiency virus infectivity by chloroquine. AIDS Res. Hum. Retrovir. 1990, 6, 481–489. [Google Scholar] [CrossRef]
- Ooi, E.E.; Chew, J.S.W.; Loh, J.P.; Chua, R.C.S. In vitro inhibition of human influenza A virus replication by chloroquine. Virol. J. 2006, 3, 39. [Google Scholar] [CrossRef] [Green Version]
- Farias, K.J.S.; Machado, P.R.L.; da Fonseca, B.A.L. Chloroquine inhibits dengue virus type 2 replication in Vero cells but not in C6/36 cells. Sci. World J. 2013, 2013, 282734. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Y.; Xu, Q.; Wu, D.; Ren, H.; Zhao, P.; Lao, W.; Wang, Y.; Tao, Q.; Qian, X.; Wei, Y.-H.; et al. Japanese encephalitis virus enters rat neuroblastoma cells via a pH-dependent, dynamin and caveola-mediated endocytosis pathway. J. Virol. 2012, 86, 13407–13422. [Google Scholar] [CrossRef] [Green Version]
- Boonyasuppayakorn, S.; Reichert, E.D.; Manzano, M.; Nagarajan, K.; Padmanabhan, R. Amodiaquine, an antimalarial drug, inhibits dengue virus type 2 replication and infectivity. Antivir. Res. 2014, 106, 125–134. [Google Scholar] [CrossRef]
- Delvecchio, R.; Higa, L.M.; Pezzuto, P.; Valadao, A.L.; Garcez, P.P.; Monteiro, F.L.; Loiola, E.C.; Dias, A.A.; Silva, F.J.; Aliota, M.T.; et al. Chloroquine, an endocytosis blocking agent, inhibits Zika virus infection in different cell models. Viruses 2016, 8, 322. [Google Scholar] [CrossRef] [Green Version]
- Chatre, C.; Roubille, F.; Vernhet, H.; Jorgensen, C.; Pers, Y.M. Cardiac complications attributed to chloroquine and hydroxychloroquine: A systematic review of the literature. Drug Saf. 2018, 41, 919–931. [Google Scholar] [CrossRef] [PubMed]
- Manuja, A.; Kumar, B.; Chhabra, D. Chloroquine Chaos and COVID-19: Smart delivery perspectives through pH sensitive polymers/micelles and ZnO nanoparticles. Arabian J. Chem. 2022, 30, 104468. [Google Scholar] [CrossRef] [PubMed]
- Manuja, A.; Kumar, B.; Riyesh, T.; Talluri, T.R.; Tripathi, B.N. Microwave assisted fast fabrication of zinc/iron oxides based polymeric nanocomposites and evaluation on equine fibroblasts. Int. J. Biol. Macromol. 2020, 165, 71–81. [Google Scholar] [CrossRef] [PubMed]
- Raguvaran, R.; Manuja, A.; Manuja, B.K.; Riyesh, T.; Singh, S.; Kesavan, M.; Dimri, U. Sodium alginate and gum acacia hydrogels of zinc oxide nanoparticles reduce hemolytic and oxidative stress inflicted by zinc oxide nanoparticles on mammalian cells. Int. J. biol. Macromol. 2017, 101, 967–972. [Google Scholar] [CrossRef]
- Manuja, A.; Raguvaran, R.; Kumar, B.; Kalia, A.; Tripathi, B.N. Accelerated healing of full thickness excised skin wound in rabbits using single application of alginate/acacia based nanocomposites of ZnO nanoparticles. Int. J. Biol. Macromol. 2020, 155, 823–833. [Google Scholar] [CrossRef]
- Available online: https://www.aatbio.com/tools/ic50-calculator-v1theequation (accessed on 7 July 2021).
- Available online: http://www.graphpad.com/quickcalcs/ttest1.cfm (accessed on 3 January 2023).
- Manuja, A.; Kumar, B.; Kumar, R.; Chhabra, D.; Ghosh, M.; Manuja, M.; Brar, B.; Pal, Y.; Tripathi, B.N.; Prasad, M. Metal/metal oxide nanoparticles: Toxicity concerns associated with their physical state and remediation for biomedical applications. Toxicol. Rep. 2021, 8, 1970–1978. [Google Scholar] [CrossRef]
- Neuman, B.W.; Kiss, G.; Kunding, A.H.; Bhella, D.; Baksh, M.F.; Connelly, S.; Droese, B.; Klaus, J.P.; Makino, S.; Sawicki, S.G.; et al. A structural analysis of M protein in coronavirus assembly and morphology. J. Struct. Biol. 2011, 174, 11–22. [Google Scholar] [CrossRef]
- Nal, B.; Chan, C.; Kien, F.; Siu, L.; Tse, J.; Chu, K.; Kam, J.; Staropoli, I.; Crescenzo-Chaigne, B.; Escriou, N.; et al. Differential maturation and subcellular localization of severe acute respiratory syndrome coronavirus surface proteins S, M and E. J. Gen. Virol. 2005, 86, 1423–1434. [Google Scholar] [CrossRef] [Green Version]
- Arndt, A.L.; Larson, B.J.; Hogue, B.G. A conserved domain in the coronavirus membrane protein tail is important for virus assembly. J. Virol. 2010, 84, 11418–11428. [Google Scholar] [CrossRef] [Green Version]
- Wu, A.; Peng, Y.; Huang, B.; Ding, X.; Wang, X.; Niu, P.; Meng, J.; Zhu, Z.; Zhang, Z.; Wang, J.; et al. Genome composition and divergence of the novel coronavirus (2019-nCoV) originating in China. Cell Host Microbe. 2020, 27, 325–328. [Google Scholar] [CrossRef] [Green Version]
- Solomon, V.R.; Lee, H. Chloroquine and its analogs: A new promise of an old drug for effective and safe cancer therapies. Eur. J. Pharmacol. 2009, 625, 220–233. [Google Scholar] [CrossRef] [PubMed]
- Mizushima, N.; Yoshimori, T.; Levine, B. Methods in mammalian autophagy research. Cell 2010, 140, 313–326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, M.; Khambu, B.; Zhang, H.; Kang, J.H.; Chen, X.; Chen, D.; Vollmer, L.; Liu, P.Q.; Vogt, A.; Yin, X.M. Suppression of lysosome function induces autophagy via a feedback down-regulation of MTOR complex 1 (MTORC1) activity. J. Biol. Chem. 2013, 288, 35769–35780. [Google Scholar] [CrossRef] [PubMed] [Green Version]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Element | ZnONFs | CHCZnO NPs |
|---|---|---|
| Oxygen | 35.56 | 43.55 |
| Zinc | 43.00 | 33.79 |
| Carbon | 6.48 | 6.98 |
| Sulfur | 14.96 | 15.68 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Manuja, A.; Kumar, B.; Chhabra, D.; Brar, B.; Thachamvally, R.; Pal, Y.; Prasad, M. Synergistic Effect of Zinc-Chitosan Nanoparticles and Hydroxychloroquine to Inhibit Buffalo Coronavirus. Polymers 2023, 15, 2949. https://doi.org/10.3390/polym15132949
Manuja A, Kumar B, Chhabra D, Brar B, Thachamvally R, Pal Y, Prasad M. Synergistic Effect of Zinc-Chitosan Nanoparticles and Hydroxychloroquine to Inhibit Buffalo Coronavirus. Polymers. 2023; 15(13):2949. https://doi.org/10.3390/polym15132949
Chicago/Turabian StyleManuja, Anju, Balvinder Kumar, Dharvi Chhabra, Basanti Brar, Riyesh Thachamvally, Yash Pal, and Minakshi Prasad. 2023. "Synergistic Effect of Zinc-Chitosan Nanoparticles and Hydroxychloroquine to Inhibit Buffalo Coronavirus" Polymers 15, no. 13: 2949. https://doi.org/10.3390/polym15132949
APA StyleManuja, A., Kumar, B., Chhabra, D., Brar, B., Thachamvally, R., Pal, Y., & Prasad, M. (2023). Synergistic Effect of Zinc-Chitosan Nanoparticles and Hydroxychloroquine to Inhibit Buffalo Coronavirus. Polymers, 15(13), 2949. https://doi.org/10.3390/polym15132949