

Carboxymethyl Cellulose-Based Polymers as Promising Matrices for Ficin Immobilization


 ,
,  , , ,
, , ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Synthesis and Characterization of the Enzyme Carrier
2.3. Fourier-Transform Infrared Spectroscopy (FTIR)
2.4. Molecular Docking
2.5. Ficin Immobilization
2.6. Protein Content Assay
2.7. Protease Activity Assay
2.8. Amidase Activity Assay
2.9. Kinetic Properties
2.10. Statistical Analysis
3. Results and Discussions
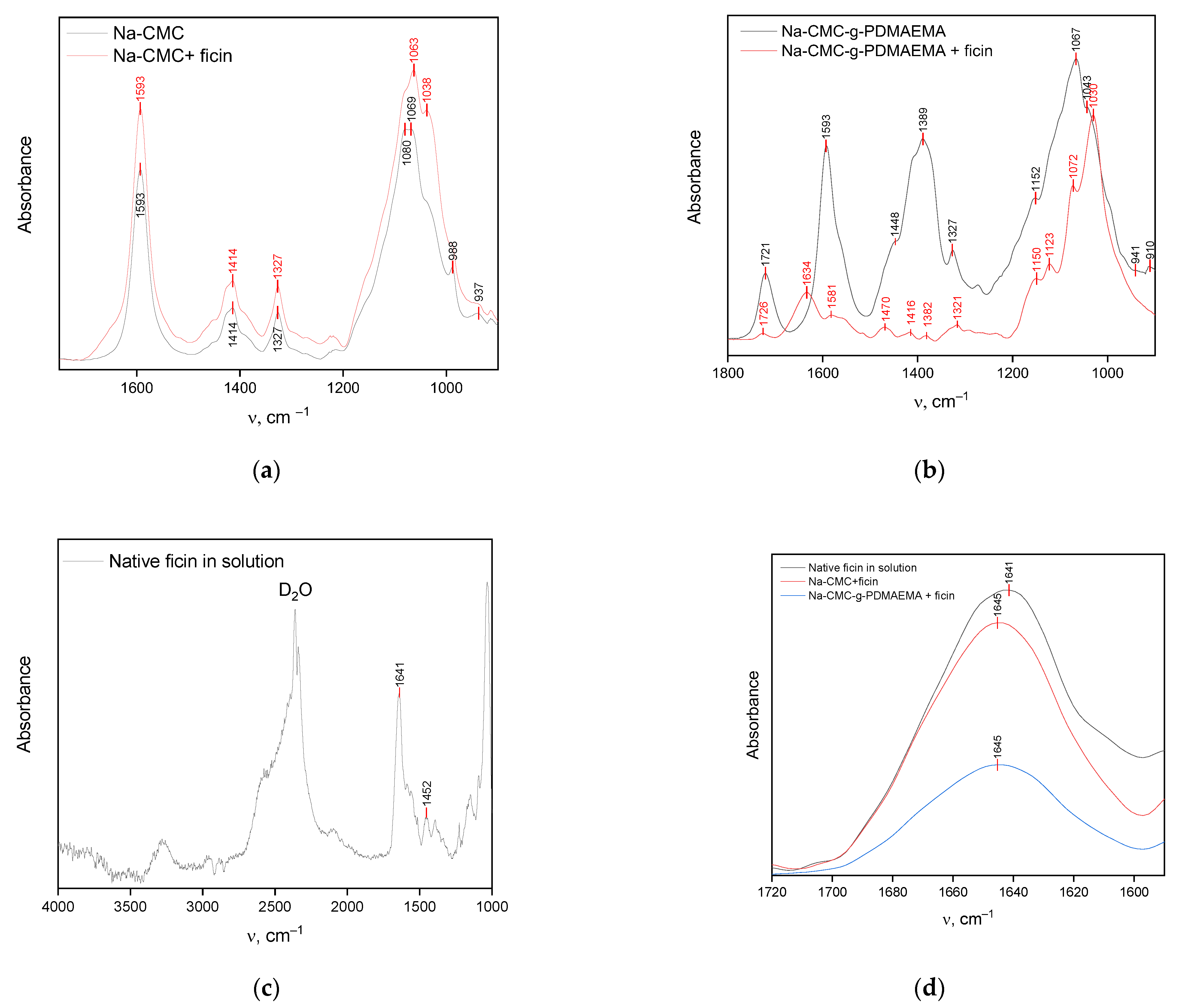
3.1. Researching the Interactions between Ficin and Na-CMC and Na-CMC-g-PDMAEMA
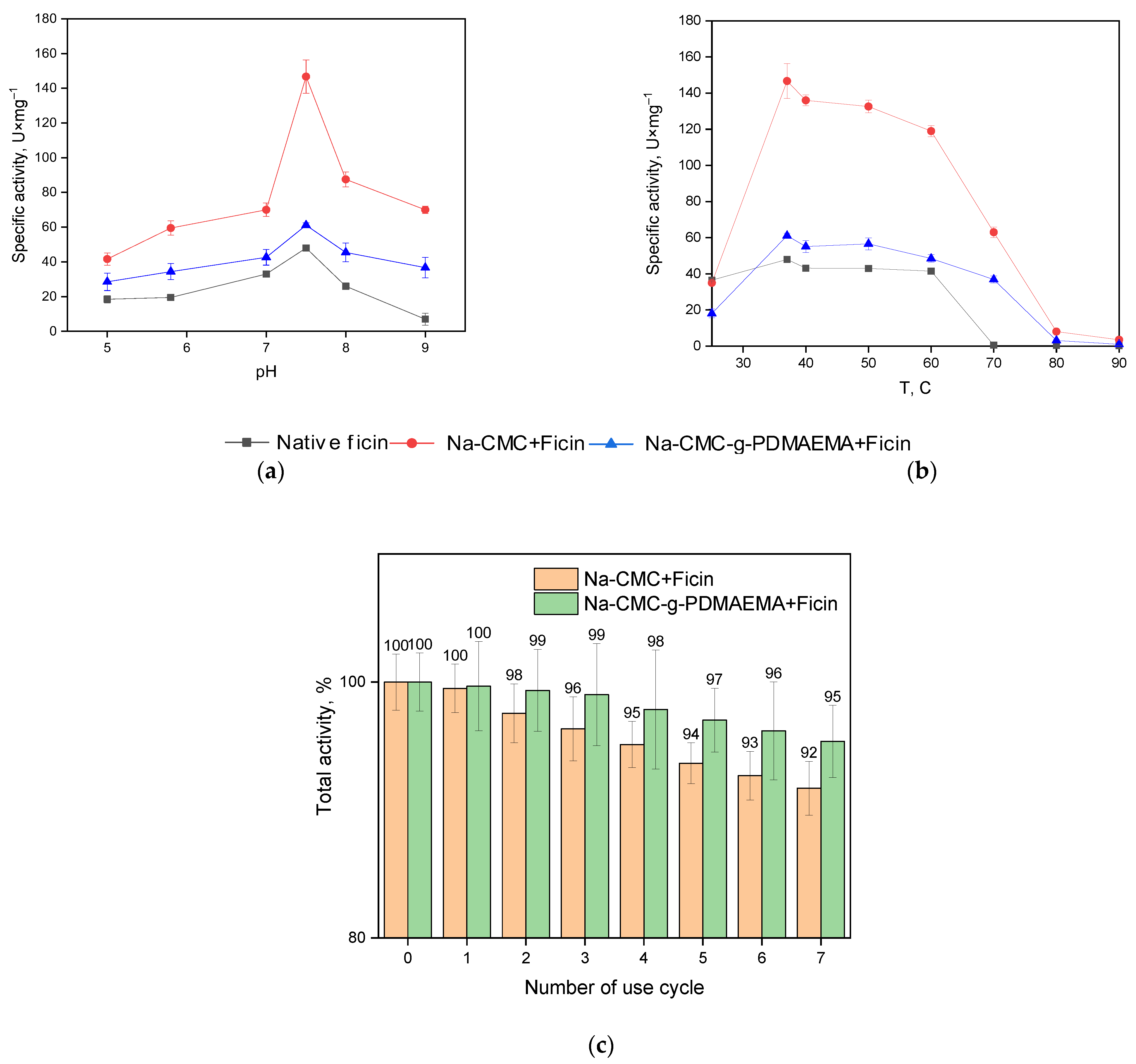
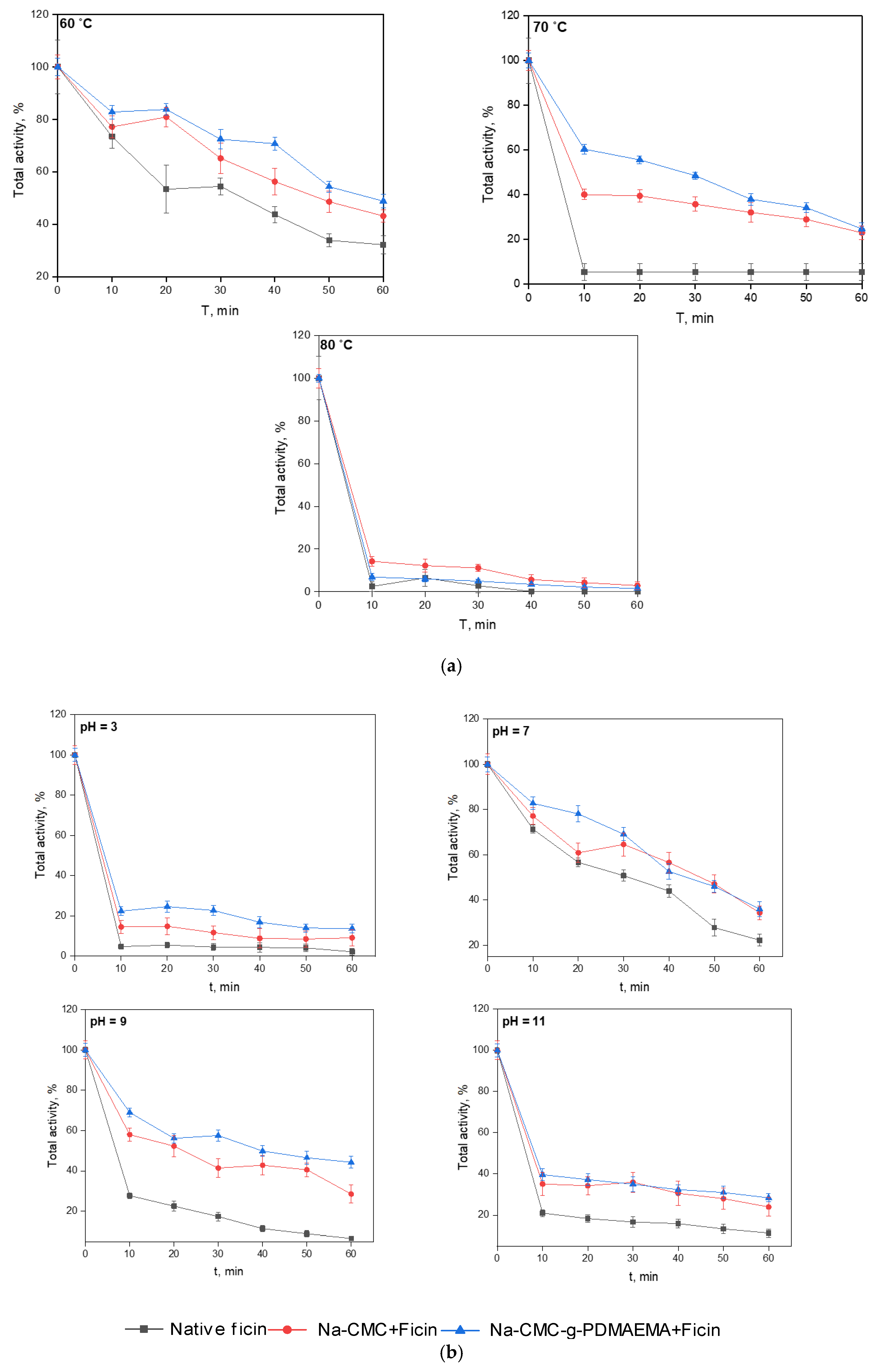
3.2. Properties of the Conjugated Ficin
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Khurd, A.S.; Kandasubramanian, B. A systematic review of cellulosic material for green electronics devices. Carbohyd. Polym. Technol. Sci. Appl. 2022, 4, 100234. [Google Scholar] [CrossRef]
- Navya, P.V.; Gayathri, V.; Samanta, D.; Sampath, S. Bacterial cellulose: A promising biopolymer with interesting properties and applications. Int. J. Biol. Macromol. 2022, 220, 435–461. [Google Scholar] [CrossRef]
- Baghel, R.S.; Reddy, C.R.K.; Singh, R.P. Seaweed-based cellulose: Applications, and future perspectives. Carbohyd. Polym. 2021, 267, 118241. [Google Scholar] [CrossRef] [PubMed]
- Xie, L.; Shen, M.; Wang, Z.; Xie, J. Structure, function and food applications of carboxymethylated polysaccharides: A comprehensive review. Trends Food Sci. Technol. 2021, 118A, 539–557. [Google Scholar] [CrossRef]
- Pinto, E.; Aggrey, W.N.; Boakye, P.; Amenuvor, G.; Sokama-Neuyam, Y.A.; Fokuo, M.K.; Karimaie, H.; Sarkodie, K.; Adenutsi, C.D.; Erzuah, S.; et al. Cellulose processing from biomass and its derivatization into carboxymethylcellulose: A review. Sci. Afr. 2022, 15, e01078. [Google Scholar] [CrossRef]
- Chakka, V.P.; Zhou, T. Carboxymethylation of polysaccharides: Synthesis and bioactivities. Int. J. Biol. Macromol. 2020, 165B, 2425–2431. [Google Scholar] [CrossRef]
- Yildirim-Yalcin, M.; Tornuk, F.; Toker, O.S. Recent advances in the improvement of carboxymethyl cellulose-based edible films. Trends Food Sci. Technol. 2022, 129, 179–193. [Google Scholar] [CrossRef]
- Javanbakht, S.; Shaabani, A. Carboxymethyl cellulose-based oral delivery systems. Int. J. Biol. Macromol. 2019, 133, 21–29. [Google Scholar] [CrossRef]
- Berlin, A.A.; Kislenko, V.N. Kinetics and mechanism of radical graft polymerization of monomers onto polysaccharides. Prog. Polym. Sci. 1992, 17, 765–825. [Google Scholar] [CrossRef]
- Bhattacharya, A.; Misra, B. Grafting: A versatile means to modify polymers: Techniques, factors and applications. Prog. Polym. Sci. 2004, 29, 767–814. [Google Scholar] [CrossRef]
- Glasing, J.; Champagne, P.; Cunningham, M.F. Graft modification of chitosan, cellulose and alginate using reversible deactivation radical polymerization (RDRP). Curr. Opin. Green Sustain. Chem. 2016, 2, 15–21. [Google Scholar] [CrossRef]
- Kuznetsov, V.A.; Sorokin, A.V.; Lavlinskaya, M.S.; Sinelnikov, A.A.; Bykovskiy, D.V. Graft copolymers of carboxymethyl cellulose with N-vinylimidazole: Synthesis and application for drug delivery. Polym. Bull. 2019, 76, 4929–4949. [Google Scholar] [CrossRef]
- Sorokin, A.V.; Kuznetsov, V.A.; Lavlinskaya, M.S. Synthesis of graft copolymers of carboxymethyl cellulose and N,N-dimethylaminoethyl methacrylate and their study as Paclitaxel carriers. Polym. Bull. 2021, 78, 2975–2992. [Google Scholar] [CrossRef]
- Iiyama, S.; Fukaya, K.; Yamaguchi, Y.; Watanabe, A.; Yamamoto, H.; Mochizuki, S.; Saio, R.; Noguchi, T.; Oishi, T.; Sato, T.; et al. Total Synthesis of Paclitaxel. Org. Lett. 2022, 24, 202–206. [Google Scholar] [CrossRef] [PubMed]
- Sorokin, A.V.; Olshannikova, S.S.; Lavlinskaya, M.S.; Holyavka, M.G.; Faizullin, D.A.; Zuev, Y.F.; Artukhov, V.G. Chitosan Graft Copolymers with N-Vinylimidazole as Promising Matrices for Immobilization of Bromelain, Ficin, and Papain. Polymers 2022, 14, 2279. [Google Scholar] [CrossRef]
- Baidamshina, D.R.; Koroleva, V.A.; Trizna, E.Y.; Pankova, S.M.; Agafonova, M.N.; Chirkova, M.N.; Vasilieva, O.S.; Akhmetova, N.; Shubina, V.V.; Porfiryev, A.G.; et al. Anti-biofilm and wound-healing activity of chitosan-immobilized Ficin. Int. J. Biol. Macromol. 2020, 164, 4205–4217. [Google Scholar] [CrossRef]
- Holyavka, M.; Faizullin, D.; Koroleva, V.; Olshannikova, S.; Zachartchenko, N.; Zuev, Y.; Kondratyev, M.; Zakharova, E.; Artukhov, V. Novel biotechnological formulations of cysteine proteases, immobilized on chitosan. Structure, stability and activity. Int. J. Biol. Macromol. 2021, 180, 161–176. [Google Scholar] [CrossRef]
- Morellon-Sterling, R.; El-Siar, H.; Tavano, O.L.; Berenguer-Murcia, Á.; Fernández-Lafuente, R. Ficin: A protease extract with relevance in biotechnology and biocatalysis. Int. J. Biol. Macromol. 2020, 160, 394–404. [Google Scholar] [CrossRef]
- Fernández-Lucas, J.; Castañeda, D.; Hormigo, D. New trends for a classical enzyme: Papain, a biotechnological success story in the food industry. Trends Food Sci. Technol. 2017, 68, 91–101. [Google Scholar] [CrossRef]
- Bié, J.; Sepodes, B.; Fernandes, P.C.B.; Ribeiro, M.H.L. Enzyme Immobilization and Co-Immobilization: Main Framework, Advances and Some Applications. Processes 2022, 10, 494. [Google Scholar] [CrossRef]
- Nunes, Y.L.; de Menezes, F.L.; de Sousa, I.G.; Cavalcante, A.L.G.; Cavalcante, F.T.T.; da Silva Moreira, K.; de Oliveira, A.L.B.; Mota, G.F.; da Silva Souza, J.R.; de Aguiar Falcão, I.R.; et al. Chemical and physical Chitosan modification for designing enzymatic industrial biocatalysts: How to choose the best strategy? Int. J. Biol. Macromol. 2021, 181, 1124–1170. [Google Scholar] [CrossRef] [PubMed]
- Mateo, C.; Palomo, J.M.; Fernandez-Lorente, G.; Guisan, J.M.; Fernandez-Lafuente, R. Improvement of enzyme activity, stability and selectivity via immobilization techniques. Enzyme Microb. Technol. 2007, 40, 1451–1463. [Google Scholar] [CrossRef]
- Sulaiman, S.; Mokhtar, M.N.; Naim, M.N.; Baharuddin, A.S.; Sulaiman, A. A Review: Potential Usage of Cellulose Nanofibers (CNF) for Enzyme Immobilization via Covalent Interactions. Appl. Biochem. Biotechnol. 2015, 175, 1817–1842. [Google Scholar] [CrossRef]
- Liu, Y.; Chen, J.Y. Enzyme immobilization on cellulose matrixes. J. Bioact. Compat. Polym. 2016, 31, 553–567. [Google Scholar] [CrossRef]
- Holyavka, M.G.; Makin, S.M.; Kondratyev, M.S.; Abdullatypov, A.V.; Kovaleva, T.A.; Artyukhov, V.G. Supramolecular Organization of Inulinases from Aspergillus awamori, Aspergillus ficuum and Kluyveromyces marxianus: A Comparative Aspect. Biophysics 2018, 63, 866–875. [Google Scholar] [CrossRef]
- Lowry, O.H.; Rosebrough, N.J.; Faar, A.L.; Randall, R.J. Protein measurement with Folin-phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Artyukhov, V.G.; Kovaleva, T.A.; Kholyavka, M.G.; Bityutskaya, L.A.; Grechkina, M.V. Thermal inactivation of free and immobilized inulinase. Appl. Biochem. Microbiol. 2010, 46, 385–389. [Google Scholar] [CrossRef]
- Sabirova, A.R.; Rudakova, N.L.; Balaban, N.P.; Ilyinskaya, O.N.; Demidyuk, I.V.; Kostrov, S.V.; Rudenskaya, G.N.; Sharipova, M.R. A novel secreted metzincin metalloproteinase from Bacillus intermedius. FEBS Lett. 2010, 584, 4419–4425. [Google Scholar] [CrossRef] [Green Version]
- Drenth, J.; Jansonius, J.N.; Koekoek, R.; Wolthers, B.G. The structure of papain. Adv. Protein Chem. 1971, 25, 79–115. [Google Scholar] [CrossRef]
- Kamphuis, I.G.; Kalk, K.H.; Swarte, M.B.A.; Drenth, J. Structure of papain refined at 1.65 Å resolution. J. Mol. Biol. 1984, 179, 233–256. [Google Scholar] [CrossRef]
- Novinec, M.; Lenarcic, B. Papain-like peptidases: Structure, function, and evolution. Biomol. Concepts 2013, 4, 287–308. [Google Scholar] [CrossRef] [PubMed]
- Vernet, T.; Tessier, D.C.; Chatellier, J.; Plouffe, C.; Lee, T.S.; Thomas, D.Y.; Storer, A.C.; Menard, R. Structural and functional roles of asparagine 175 in the cysteine protease papain. J. Biol. Chem. 1995, 270, 16645–16652. [Google Scholar] [CrossRef] [Green Version]
- Habibi, N. Preparation of biocompatible magnetite-carboxymethyl cellulose nanocomposite: Characterization of nanocomposite by FTIR, XRD, FESEM and TEM. Spectrochim. Acta A Mol. Biomol. Spectrosc. 2014, 131, 55–58. [Google Scholar] [CrossRef] [PubMed]
- Karkhari, R.; Javanbakht, V. A polyurethane foam membrane filled with double cross-linked chitosan/carboxymethyl cellulose gel and decorated with ZSM-5 nano zeolite: Simultaneous dye removal. Int. J. Biol. Macromol. 2022, 213, 699–717. [Google Scholar] [CrossRef] [PubMed]
- Sorokin, A.; Sukhanov, P.; Popov, V.; Kannykin, S.; Lavlinskaya, M. A new approach to increasing the equilibrium swelling ratio of the composite superabsorbents based on carboxymethyl cellulose sodium salt. Cellulose 2022, 29, 159–173. [Google Scholar] [CrossRef]
- Barth, A.; Zscherp, C. What vibrations tell us about proteins. Q. Rev. Biophys. 2022, 35, 369–430. [Google Scholar] [CrossRef]
- Jackson, M.; Mantsch, H.H. The Use and Misuse of FTIR Spectroscopy in the Determination of Protein Structure. Crit. Rev. Biochem. Mol. Biol. 1995, 30, 95–120. [Google Scholar] [CrossRef]
- Milošević, J.; Vrhovac, L.; Đurković, F.; Janković, B.; Malkov, S.; Lah, J.; Polović, N.Đ. Isolation, identification, and stability of Ficin 1c isoform from fig latex. New J. Chem. 2020, 44, 15716–15723. [Google Scholar] [CrossRef]
- Ding, S.; Cargill, A.A.; Medintz, I.L.; Claussen, J.C. Increasing the activity of immobilized enzymes with nanoparticle conjugation. Curr. Opin. Biotechnol. 2015, 34, 242–250. [Google Scholar] [CrossRef] [Green Version]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Binding Site Number | Affinity, kcal/mol | Amino Acid Residues Forming | |
|---|---|---|---|
| H-Bonds, Length, Å | Other Interactions | ||
| Amino acids interacting with Na-CMC: | |||
| 1 | −6.7 | Gly20, 2.73; Cys22, 2.9; Ser66, 3.09; Glu145 (αR2), 2.66 and 3.10; Asp161, 2.76 and 3.06 | Gln19, Arg21, Gly23, Cys25 (αL1), Tyr60, Gly140, Asn187, Trp188 |
| 2 | −6.5 | Gly140, 3.13; Leu160, 2.70 and 3.34 | Arg21, Tyr60, Gly68, Trp69, Thr136 (βR), Glu145 (αR2), Asp161, Gln212 |
| 3 | −6.3 | Arg118, 2.96; Thr158, 2.97 and 2.77; Met211, 2.97 | Trp69, Met70 (αL3), Thr71 (αL3), Lys72 (αL3), Arg118, Thr158, Lys159, Leu160, Met211, Gln212 |
| 4 | −6.3 | Ser213, 3.16 | His115 (βR), Val116, Pro117 |
| 5 | −6.3 | - | His115 (βR) |
| Amino acids interacting with Na-CMC-g-PDMAEMA: | |||
| 1 | −5.0 | Cys65, 3.28; Ser66, 2.80; Gly68, 3.25; Asp161, 2.79 | Asn18, Arg21, Cys22, Gly23, Cys25 (αL1), Trp26, Tyr60, Cys65, Ser66, Gly67, Gly68, Trp69, Met70 (αL3), Thr136 (βR), Glu138, Gly140, Gly141, Glu145 (αR2), Leu146, Thr158, Lys159, Leu160, Asp161, His162 (βR), Trp184, Asn187, Trp188, Gln212 |
| 2 | −3.7 | Asn14, 2.79; Asp43, 2.93; Glu44, 2.10; Leu45, 3.11; Ser47, 3.33; | Arg8, Ile9, Gly11, Val13, Asn14, Pro15, Ile16, Arg17, Asn18, Asp43, Glu44, Leu45, Pro46, Ser47, Gln86, Ser87, Pro90, Tyr91 Val107, Gly185, Thr186, Arg191 |
| 3 | −3.6 | Asn80, 2.79 and 2.76; Ser104, 2.94, 3.01, 3.10 and 3.07; Ile106, 3.16 | Ser38 (αL1), Val41 (αL1), Thr42 (αL1), Glu44, Pro46, Ile78 (αL3), Lys79 (αL3), Asn80, Gly81, Lys100 (αR1), Asp101 (αR1), Ser104, Gln105, Ile106, Val107, Ala108, Thr109, Ile110, Asp111 |
| 4 | −3.7 | Thr92, 2.93 | Gly20, Arg21, Gln51 (αL2), Asp55 (αL2), Ser87, Asn88, Tyr89, Pro90, Thr92, Ala93, Lys94, Gly96, Glu97, Cys98, Asn99, Lys100 (αR1), Asp101 (αR1), Leu102 (αR1) |
| 5 | −3.9 | Trp69, 2.81; Val116, 3.13 and 3.28; Arg118, 3.28; Thr71 (αL3), 3.31 | Thr58, Ser59, Tyr60, Lys61, Trp69, Thr71 (αL3), Lys72 (αL3), Glu114 (βR), His115 (βR), Val116, Pro117, Arg118, Leu160, Met211, Gln212,Tyr215 (βR) |
| Structure Elements | Structure Content | |||
|---|---|---|---|---|
| Ficin X-ray [38] | Ficin in Solution | Ficin with Na-CMC-g-PDMAEMA | Ficin with Na-CMC | |
| α-helices | 0.27 | 0.26 | 0.19 | 0.15 |
| β-sheets | 0.19 | 0.22 | 0.46 | 0.21 |
| Others | 0.54 | 0.52 | 0.35 | 0.64 |
| Enzyme Formulations | Km, μM | Vmax, μM mg−1 min−1 | kcat, min−1 |
|---|---|---|---|
| Ficin in solution | 61 ± 10 | 1314 ± 215 | 25 ± 3 |
| Ficin + Na-CMC | 19 ± 2 | 3950 ± 270 | 77 ± 9 |
| Ficin + Na-CMC-g-PDMAEMA | 50 ± 8 | 1580 ± 221 | 29 ± 3 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sorokin, A.V.; Goncharova, S.S.; Lavlinskaya, M.S.; Holyavka, M.G.; Faizullin, D.A.; Kondratyev, M.S.; Kannykin, S.V.; Zuev, Y.F.; Artyukhov, V.G. Carboxymethyl Cellulose-Based Polymers as Promising Matrices for Ficin Immobilization. Polymers 2023, 15, 649. https://doi.org/10.3390/polym15030649
Sorokin AV, Goncharova SS, Lavlinskaya MS, Holyavka MG, Faizullin DA, Kondratyev MS, Kannykin SV, Zuev YF, Artyukhov VG. Carboxymethyl Cellulose-Based Polymers as Promising Matrices for Ficin Immobilization. Polymers. 2023; 15(3):649. https://doi.org/10.3390/polym15030649
Chicago/Turabian StyleSorokin, Andrey V., Svetlana S. Goncharova, Maria S. Lavlinskaya, Marina G. Holyavka, Dzhigangir A. Faizullin, Maxim S. Kondratyev, Sergey V. Kannykin, Yuriy F. Zuev, and Valeriy G. Artyukhov. 2023. "Carboxymethyl Cellulose-Based Polymers as Promising Matrices for Ficin Immobilization" Polymers 15, no. 3: 649. https://doi.org/10.3390/polym15030649
APA StyleSorokin, A. V., Goncharova, S. S., Lavlinskaya, M. S., Holyavka, M. G., Faizullin, D. A., Kondratyev, M. S., Kannykin, S. V., Zuev, Y. F., & Artyukhov, V. G. (2023). Carboxymethyl Cellulose-Based Polymers as Promising Matrices for Ficin Immobilization. Polymers, 15(3), 649. https://doi.org/10.3390/polym15030649