Antibacterial Activity of Four Human Beta-Defensins: HBD-19, HBD-23, HBD-27, and HBD-29

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Experimental Section
2.1. Predicting Mature Defensin Amino and Carboxy Termini
2.2. Reduction and Refolding of Synthetic Defensins
2.3. Antibacterial Activity Assays
2.4. Cellular Toxicity Assay
3. Results
3.1. Predicting Mature Defensin Amino and Carboxy Termini

3.2. Refolding Synthetic Defensins

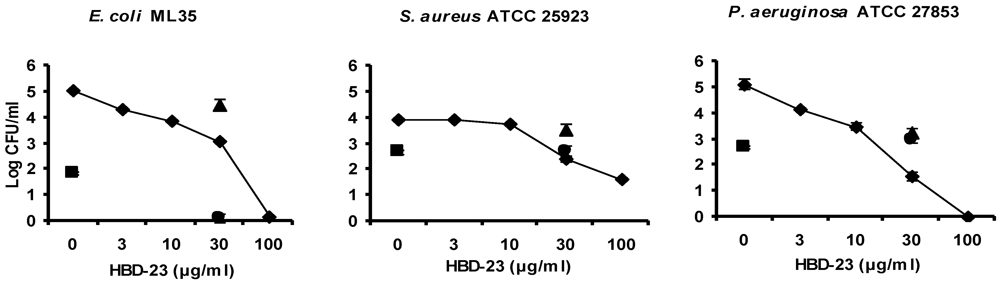
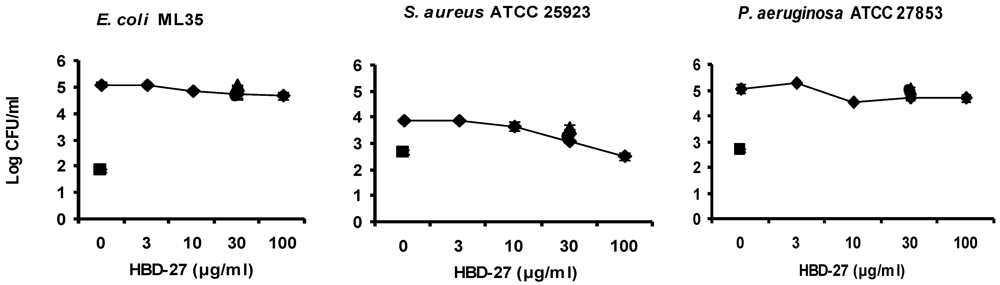
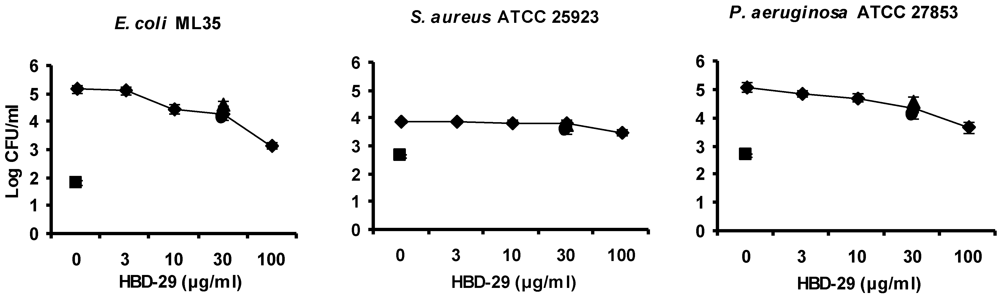
3.3. Antibacterial Activity Assay of Synthetic Defensins




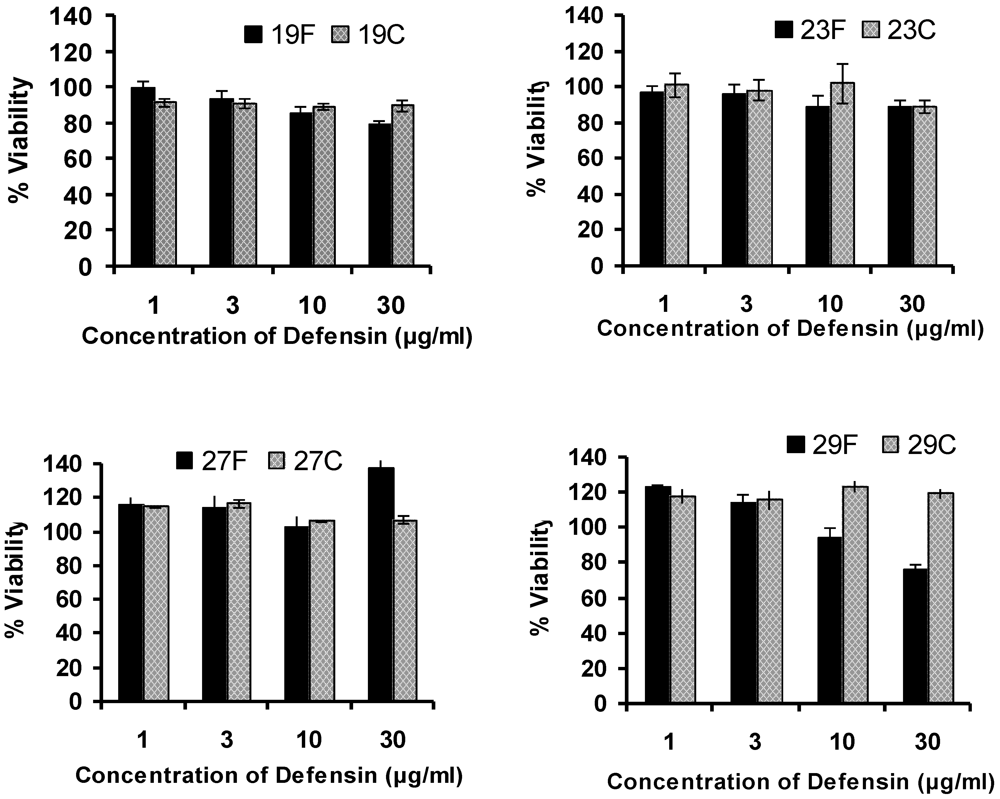
3.4. Cellular Toxicity in Peripheral Blood Mononuclear Cells

3.5. Discussion
4. Conclusions
Acknowledgments
References
- Ganz, T.; Selsted, M.E.; Szklarek, D.; Harwig, S.S.; Daher, K.; Bainton, D.F.; Lehrer, R.I. Defensins. Natural peptide antibiotics of human neutrophils. J. Clin. Invest. 1985, 76, 1427–1435. [Google Scholar] [CrossRef]
- Lehrer, R.I.; Daher, K.; Ganz, T.; Selsted, M.E. Direct inactivation of viruses by MCP-1 and MCP-2, natural peptide antibiotics from rabbit leukocytes. J. Virol. 1985, 54, 467–472. [Google Scholar]
- Lehrer, R.I.; Lichtenstein, A.K.; Ganz, T. Defensins: Antimicrobial and cytotoxic peptides of mammalian cells. Annu. Rev. Immunol. 1993, 11, 105–128. [Google Scholar] [CrossRef]
- Seidel, A.; Ye, Y.; de Armas, L.R.; Soto, M.; Yarosh, W.; Marcissin, R.; Tran, P.; Selsted, M.; Camerini, D. Cyclic and acyclic defensins inhibit human immunodeficiency virus type-1 replication by different mechanisms. PLoS One 2010, 5. [Google Scholar] [CrossRef]
- Selsted, M.E.; Szklarek, D.; Oppenheim, J.J.; Hehlgans, T. Purification and antibacterial activity of antimicrobial peptides of rabbit granulocytes. Infect. Immun. 1984, 45, 150–154. [Google Scholar]
- García, J.R.; Jaumann, F.; Schulz, S.; Krause, A.; Rodríguez-Jiménez, F.J.; Forssmann, U.; Adermann, K.; Klüver, E.; Vogelmeier, C.; Becker, D.; et al. Identification of a novel, multifunctional beta-defensin (human beta-defensin 3) with specific antimicrobial activity. Its interaction with plasma membranes of Xenopus oocytes and the induction of macrophage chemoattraction. Cell Tissue Res. 2001, 306, 257–264. [Google Scholar] [CrossRef]
- García, J.R.; Krause, A.; Schulz, S.; Rodríguez-Jiménez, F.J.; Klüver, E.; Adermann, K.; Forssmann, U.; Frimpong-Boateng, A.; Bals, R.; Forssmann, W.G. Human beta-defensin 4: A novel inducible peptide with a specific salt-sensitive spectrum of antimicrobial activity. FASEB J. 2001, 15, 1819–1821. [Google Scholar]
- Maisetta, G.; Batoni, G.; Esin, S.; Luperini, F.; Pardini, M.; Bottai, D.; Florio, W.; Giuca, M.R.; Gabriele, M.; Campa, M. Activity of human beta-defensin 3 alone or combined with other antimicrobial agents against oral bacteria. Antimicrob. Agents Chemother. 2003, 47, 3349–3351. [Google Scholar]
- Tomita, T.; Hitomi, S.; Nagase, T.; Matsui, H.; Matsuse, T.; Kimura, S.; Ouchi, Y. Effect of ions on antibacterial activity of human beta defensin 2. Microbiol. Immunol. 2000, 44, 749–754. [Google Scholar]
- Valore, E.V.; Park, C.H.; Quayle, A.J.; Wiles, K.R.; McCray, P.B., Jr.; Ganz, T. Human beta-defensin 1: An antimicrobial peptide of urogenital tissues. J. Clin. Invest. 1998, 101, 1633–1642. [Google Scholar] [CrossRef]
- Hancock, R.E.; Lehrer, R. Cationic peptides: A new source of antibiotics. Trends. Biotechnol. 1998, 16, 82–88. [Google Scholar] [CrossRef]
- Kagan, B.L.; Selsted, M.E.; Ganz, T.; Lehrer, R.I. Antimicrobial defensin peptides form voltage-dependent ion permeable channels in planar lipid bilayer membranes. Proc. Natl. Acad. Sci. USA 1990, 87, 210–214. [Google Scholar]
- Harder, J.; Bartels, J.; Christophers, E.; Schroder, J.M. Isolation and characterization of human beta-defensin-3, a novel human inducible peptide antibiotic. J. Biol. Chem. 2001, 276, 5707–5713. [Google Scholar]
- Shimoda, M.; Ohki, K.; Shimamoto, Y.; Kohashi, O. Morphology of defensin-treated Staphylococcus aureus. Infect. Immun. 1995, 63, 2886–2891. [Google Scholar]
- Chertov, O.; Michiel, D.F.; Xu, L.; Wang, J.M.; Tani, K.; Murphy, W.J.; Longo, D.L.; Taub, D.D.; Oppenheim, J.J. Identification of defensin-1, defensin-2, and CAP37/azurocidin as T-cell chemoattractant proteins released from interleukin-8-stimulated neutrophils. J. Biol. Chem. 1996, 271, 2935–2940. [Google Scholar]
- Niyonsaba, F.; Ogawa, H.; Nagaoka, I. Human beta-defensin-2 functions as a chemotactic agent for tumour necrosis factor-alpha-treated human neutrophils. Immunology 2004, 111, 273–281. [Google Scholar] [CrossRef]
- Rorhl, J.; Yang, D.; Lehrer, R.I. Human beta-defensin 2 and 3 and their mouse orthologs induce chemotaxis through interaction with CCR2. J. Immunol. 2010, 184, 6688–6694. [Google Scholar] [CrossRef]
- Yang, D.; Chertov, O.; Bykovskaia, S.N.; Chen, Q.; Buffo, M.J.; Shogan, J.; Anderson, M.; Schröder, J.M.; Wang, J.M.; Howard, O.M.; et al. Beta-defensins: Linking innate and adaptive immunity through dendritic and T cell CCR6. Science 1999, 286, 525–528. [Google Scholar]
- Radhakrishnan, Y.; Hamil, K.G.; Yenugu, S.; Young, S.L.; French, F.S.; Hall, S.H. Identification, characterization, and evolution of a primate beta-defensin gene cluster. Genes Immun. 2005, 6, 203–210. [Google Scholar] [CrossRef]
- Rodríguez-Jiménez, F.J. Identification and characterization of three new human beta-defensins: hBD23, hBD27, hBD29. Ph.D. dissertation, University of Hanover, Hannover, Germany, 2003. [Google Scholar]
- Rodríguez-Jiménez, F.J.; Krause, A.; Schulz, S.; Forssmann, W.G.; Conejo-Garcia, J.R.; Schreeb, R.; Motzkus, D. Distribution of new human beta-defensin genes clustered on chromosome 20 in functionally different segments of epididymis. Genomics 2003, 81, 175–183. [Google Scholar]
- SignalP 4.0 Server. Available online: http://www.cbs.dtu.dk/services/SignalP (accessed on 1 February 2012).
- Llenado, R.A.; Weeks, C.S.; Cocco, M.J.; Ouellette, A. Electropositive charge in alpha-defensin bactericidal activity: Functional effects of Lys-for-Arg substitutions vary with peptide primary structure. Infect. Immun. 2009, 77, 5035–5043. [Google Scholar] [CrossRef]
- Com, E.; Bourgeon, F.; Evrard, B.; Ganz, T.; Colleu, D.; Jégou, B.; Pineau, C. Expression of antimicrobial defensins in the male reproductive tract of rats, mice, and humans. Biol. Reprod. 2003, 68, 95–104. [Google Scholar]
- Hoover, D.M.; Rajashankar, K.R.; Blumenthal, R.; Puri, A.; Oppenheim, J.J.; Chertov, O.; Lubkowski, J. The structure of the human beta-defensin-2 shows evidence of higher order oligomerization. J. Biol. Chem. 2000, 275, 32911–32918. [Google Scholar]
- Zou, G.; de Leeuw, E.; Li, C.; Pazgier, M.; Li, C.; Zeng, P.; Lu, W.Y.; Lubkowski, J.; Lu, W. Toward understanding the cationicitiy of defensins. Arg and Lys versus their noncoded analogs. J. Biol. Chem. 2007, 27, 19653–19665. [Google Scholar]
- Boman, H.G. Innate immunity and the normal microflora. Immunol. Rev. 2000, 173, 5–16. [Google Scholar]
- Matsuzaki, K.; Horikiri, C. Interactions of amyloid beta-peptide (1-40) with ganglioside-containing membranes. Biochemistry 1999, 38, 4137–4142. [Google Scholar] [CrossRef]
- Tanabe, H.; Ayabe, T.; Maemoto, A.; Ishikawa, C.; Inaba, Y.; Sato, R.; Moriichi, K.; Okamoto, K.; Watari, J.; Kono, T.; et al. Denatured human alpha-defensin attenuates the bactericidal activity and the stability against enzymatic digestion. Biochem Biophys Res Commun. 2007, 358, 349–355. [Google Scholar] [CrossRef]
- Hwang, P.M.; Vogel, H.J. Structure-function relationships of antimicrobial peptides. Biochem. Cell. Biol. 1998, 76, 235–246. [Google Scholar] [CrossRef]
- Quinones-Mateu, M.E.; Lederman, M.M.; Feng, Z.; Chakraborty, B.; Weber, J.; Rangel, H.R.; Marotta, M.L.; Mirza, M.; Jiang, B.; Kiser, P.; et al. Human epithelial beta-defensins 2 and 3 inhibit HIV-1 replication. AIDS 2003, 17, F39–F48. [Google Scholar]
- Sun, L.; Finnegan, C.M.; Kish-Catalone, T.; Blumenthal, R.; Garzino-Demo, P.; La Terra Maggiore, G.M.; Berrone, S.; Kleinman, C.; Wu, Z.; Abdelwahab, S.; et al. Human beta-defensins suppress human immunodeficiency virus infection: Potential role in mucosal protection. J. Virol. 2005, 79, 14318–14329. [Google Scholar]
- Territo, M.C.; Ganz, T.; Selsted, M.E.; Lehrer, R. Monocyte-chemotactic activity of defensins from human neutrophils. J. Clin. Invest. 1989, 84, 2017–2020. [Google Scholar] [CrossRef]
- Funderburg, N.; Lederman, M.M.; Feng, Z.; Drage, M.G.; Jadlowsky, J.; Harding, C.V.; Weinberg, A.; Sieg, S.F. Human beta-defensin-3 activates professional antigen-presenting cells via Toll-like receptors 1 and 2. Proc. Natl. Acad. Sci. USA 2007, 104, 18631–18635. [Google Scholar]
- Niyonsaba, F.; Ushio, H.; Nagaoka, I.; Okumura, K.; Ogawa, H. The human beta-defensins (-1, -2, -3, -4) and cathelicidin LL-37 induce IL-18 secretion through p38 and ERK MAPK activation in primary human keratinocytes. J. Immunol. 2005, 175, 1776–1784. [Google Scholar]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Chow, B.T.; Soto, M.; Lo, B.L.; Crosby, D.C.; Camerini, D. Antibacterial Activity of Four Human Beta-Defensins: HBD-19, HBD-23, HBD-27, and HBD-29. Polymers 2012, 4, 747-758. https://doi.org/10.3390/polym4010747
Chow BT, Soto M, Lo BL, Crosby DC, Camerini D. Antibacterial Activity of Four Human Beta-Defensins: HBD-19, HBD-23, HBD-27, and HBD-29. Polymers. 2012; 4(1):747-758. https://doi.org/10.3390/polym4010747
Chicago/Turabian StyleChow, Bryan T., Maira Soto, Bonnie L. Lo, David C. Crosby, and David Camerini. 2012. "Antibacterial Activity of Four Human Beta-Defensins: HBD-19, HBD-23, HBD-27, and HBD-29" Polymers 4, no. 1: 747-758. https://doi.org/10.3390/polym4010747
APA StyleChow, B. T., Soto, M., Lo, B. L., Crosby, D. C., & Camerini, D. (2012). Antibacterial Activity of Four Human Beta-Defensins: HBD-19, HBD-23, HBD-27, and HBD-29. Polymers, 4(1), 747-758. https://doi.org/10.3390/polym4010747