2. A Single Semiflexible Filament
Semiflexible filaments can be categorized into natural, mainly biological ones such as DNA, actin, collagen, microtubules, intermediate filaments, etc., and the synthetic filaments such as carbon nanotubes or stiff polymer chains. The main physical feature of this class of polymer chains is that there is a finite bending rigidity, such that their mechanical properties are determined by the competition between the elastic potential energy of bending and the entropic free energy arising from thermal fluctuations of their conformations. There is a possibility to have elastic energy of extension added to a model, but our first aim is to examine filaments that are inextensible.
The wormlike chain (WLC) model, proposed by Kratky and Porod in 1949 [
12], is widely used for describing the elasticity of a single semiflexible filament, since the work of Fixman and Kovac in 1973 [
13]. As sketched in
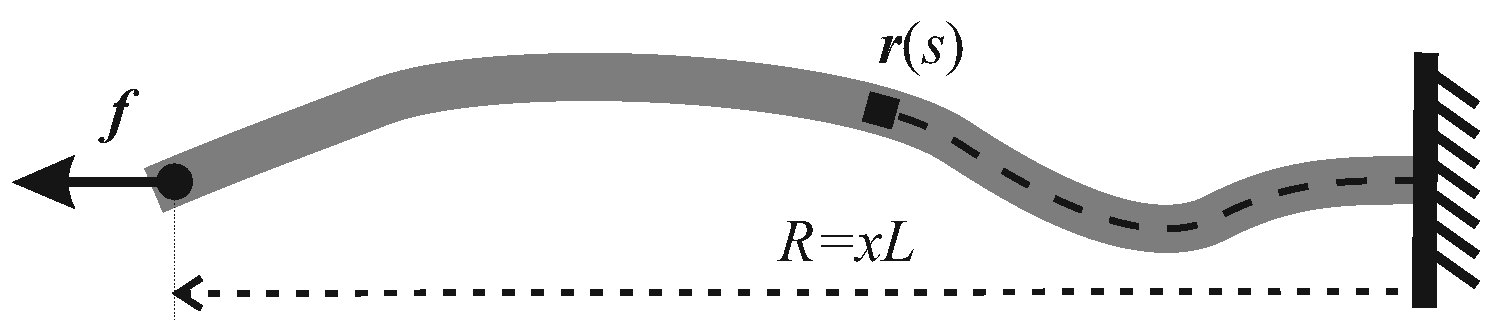
Figure 1, a filament with its contour length as
L is coordinated by
, where
is the arc-length coordinate along the chain. By assuming a finite bending modulus of the filament, its Hamiltonian can be expressed as
where
denotes for local curvature of a curve, and
κ is the bending modulus. In classical elasticity, the bending modulus of a filament is given by the Young modulus of its material and the shape of its cross-section:
, where the second moment of filament cross-section
; for an elastic tube of outer radius
a and inner radius
b (the wall thickness
) the bending modulus will be
. Persistence length, defined as
, is a key quantity for describing whether a chain is regarded as flexible (
), semiflexible (
), or rigid (
). As the persistence length is a function of the temperature, a chain can become more flexible under high temperature, and vice versa. As noticed from how the conformation is parameterized in WLC model by the coordinate
, the semiflexible chain is naturally assumed as locally inextensible, i.e.,
, for all
s.
As mentioned at the beginning, except for the bending energy defined above, the other important energy source of a semiflexible chain is thermal fluctuation of its conformations, or simply the conformational entropy. This can be more clearly understood by formulating the free energy of a semiflexible chain,
, as a function of its end-to-end separation,
, which is useful in analyzing its mechanic properties, such as the force-extension relation. Alternatively, it can be re-expressed as a function of the non-dimensional ’end-to-end factor’
. The free energy of a semiflexible chain is
where the probability
is the partition function at a constrained end-to-end distance, expressed by the standard ’Edwards path integral’:
where the local inextensibility is incorporated as a delta-function constraint in the path integral. However, it is difficult to obtain an analytic free energy expression of an inextensible chain defined above: although the bending Hamiltonian is quadratic in the field
, the inextensibility constraint makes this classical problem highly non-linear. One of the ways for resolving this difficulty is by using the concept of mean extensibility (often called the ’mean field approximation’ in this context), where the total length of the chain
L is conserved, while local fluctuations are allowed. Although mathematically this is an approximation – physically this limit is perhaps a more reasonable model of a semiflexible chain or filament. The idea of mean extensibility is simple: the Fourier transformation of
δ-function in Equation (
4) is
where
is introduced as an auxiliary field, imposing the original constraint at each point
s along the filament. If we replace
with a constant value
ϕ (the ’mean field’), then the chain can be extensible locally, but with the average inextensibility will be maintained:
. The validity of this approximation can be checked by calculating the fluctuations of the local length. As discussed in Ref. [
14], if a chain is divided into
N segments with equal length as
, then the fluctuation in the local arc-length of an individual segment, Δ, can be expressed as
, which is equal to
for
; the fluctuations are well-bounded. With this approximation, the probability function (
4) can be reduced to a compact integral in infinite limits:
where the single non-dimensional parameter
measures the deviation of the chain from its full extension, and at the same time – encodes the effective stiffness of the filament [
14]. It turns out that a simple analytical expression
captures the full range of Equation (
6). A numerical calculation of the probability function is shown in
Figure 2a. This numerical integration is quite non-trivial; it uses the fact that the imaginary part of the integrand in Equation (
6) is antisymmetric in
ϕ, while the (symmetric) real part has several points on the
ϕ-axis where an artefact of discontinuous sign-reversal occurs in numerical evaluation. So the integral has to be evaluated piece-wise, correcting for these artefacts. The analytical interpolation Equation (
7) is also plotted in
Figure 2a, shown to follow the numerical result with high precision. The close match across the whole range of
γ suggests that this is, in fact, an exact analytical result of integration (which we simply didn’t know how to calculate). This allows an immediate evaluation of the free energy of semiflexible filament
:
where the non-dimensional stiffness parameter
is the single factor that determines the properties of an inextensible filament. Note that this expression has no approximations (apart from the ’mean-field’ approximation for an auxiliary field
ϕ), and so it is equally valid in the limit of flexible chains (
), giving the classical entropic-spring result, in the limit of highly stretched flexible chain (
), and also in the limit of rigid athermal elastic rod (
).
2.1. Models of the Force-Extension Relation
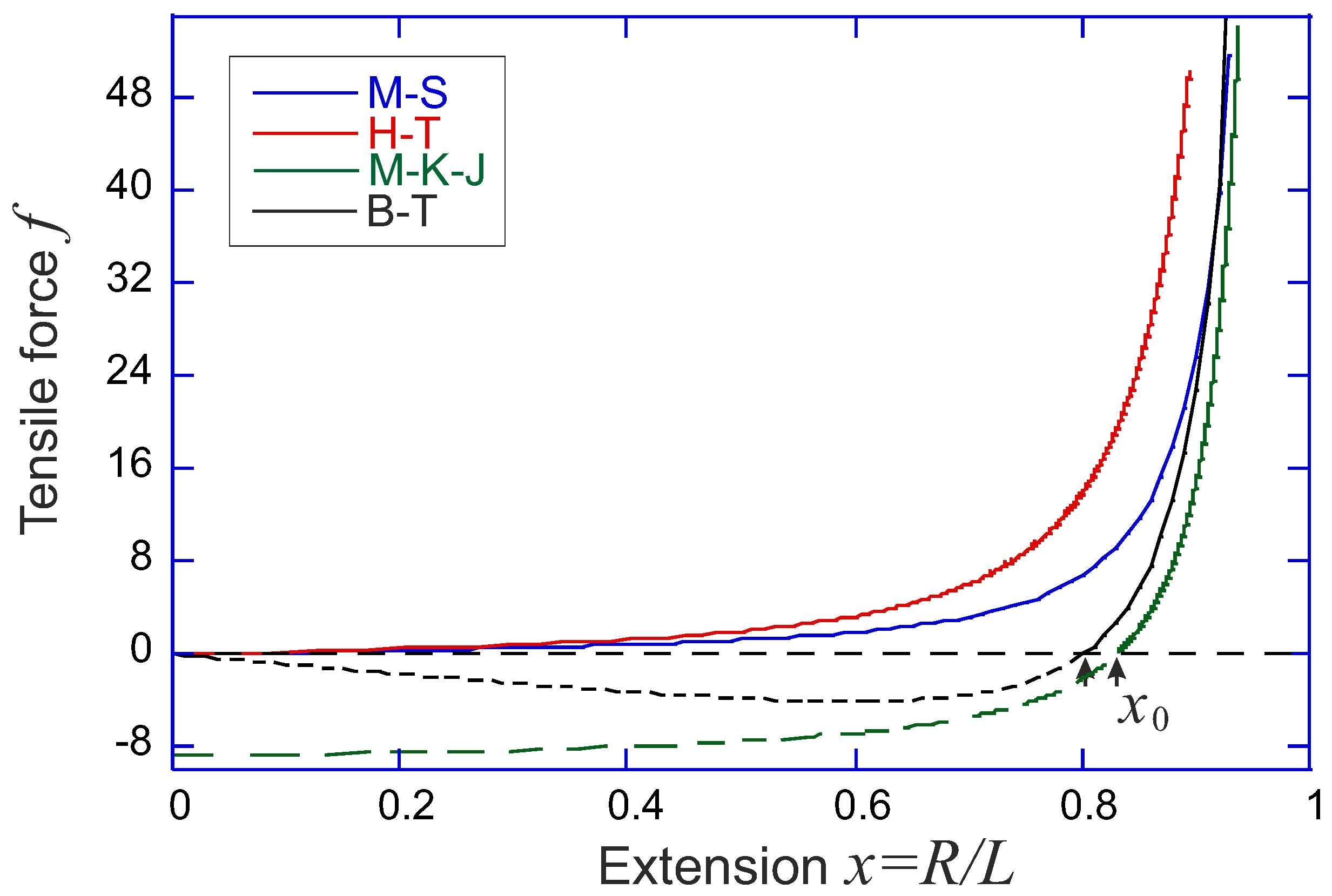
Over the years, several famous analytical forms of the force-extension relation of a semiflexible chain as a function of its end-to-end distance have been developed (comparison shown in
Figure 3). We discuss these force-extension expressions here, while the corresponding free energy in each case could be obtained by an integral of the force over the extension. In particular, when we compare with the numerical result of Equation (
6) in
Figure 2a, the constrained free energy expressions are obtained by integration, and then raised to exponent to get a non-normalized probability plotted.
Marko-Siggia model [
10]. In 1995, Marko and Siggia proposed an approximate force-expansion relation as:
It originates from the high-extension limit of the WLC model, which was interpolated such that it also matches the low-force limit of entropic spring. This expression works very well for chains near full extension, and has a long and successful history of application in many experimental situations [
15]. For instance, most values of the persistence length of various biological filaments in the literature are obtained by fitting force-extension curves with the Marko-Siggia expression (
9). When we examine this expression, it turns out that it is valid for a long, flexible chain in the highly stretched limit; in other words, at
in Equations (
7) and (
8).
Ha-Thirumalai model [
16]. In 1997, Ha and Thirumalai proposed another approximate force-expansion relation by numerically solving:
This expression, as well as the Marko-Siggia model, and also the Odijk model [
17], all predict the limiting behavior of
at
, with a slightly different numerical factor each. This is an inherent feature of the inextensible WLC model in the limit of high extension when the transverse excursions of the chain are small (certainly no ’overhangs’ are possible in the chain configuration). However, slightly different values of
would result by equally successfully fitting the force-extension curves of chains and filaments by these expressions. In terminology of Equations (
7) and (
8), these limits correspond to the regime
, that is: a flexible chain at high extension. The validity of Ha-Thirumalai model is expanded to a wider range of
γ compared with that by Marko and Siggia, as shown in
Figure 2a.
MacKintosh-Käs-Janmey model [
18]. Another way of evaluating the equilibrium elastic response of filaments to tensile force
f, starting from the same WLC model, is by applying equipartition to each bending mode in Fourier space, which is nicely reviewed in [
19] by Broedersz and Mackintosh, and in [
20] by Palmer and Boyce. Again, using the mean-field constraint on chain extensibility, but a different way of performing the summation of Fourier modes, the end-to-end factor is expressed as a function of applied force:
where the shorthand
. The force-extension relation can be obtained by numerically inverting the relation between
x and
f (Palmer and Boyce [
20] offer a rather accurate analytical interpolation using Padè approximation, valid in the tensile deformation regime). In the limit of high extensions, the divergent tensile force follows the same characteristic limit as all other models:
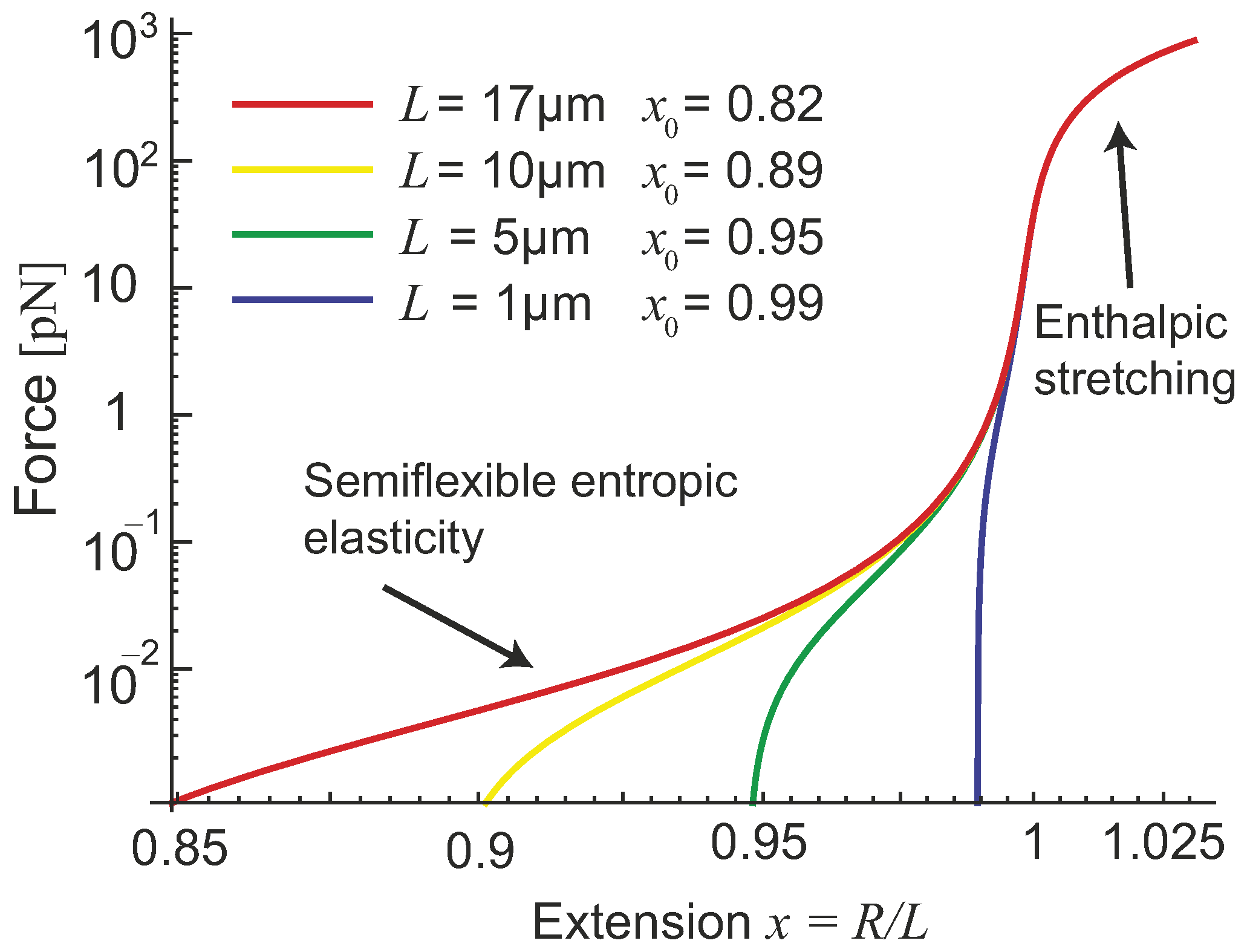
. A very important factor, missing in earlier models, is the natural length of the filament: the force-free average end-to-end extension we labelled as
in
Figure 2b. Palmer and Boyce give it as
for this model (e.g., at
we have
). In the linear regime (at small deformations around
) the filament responds with the spring constant 90
.
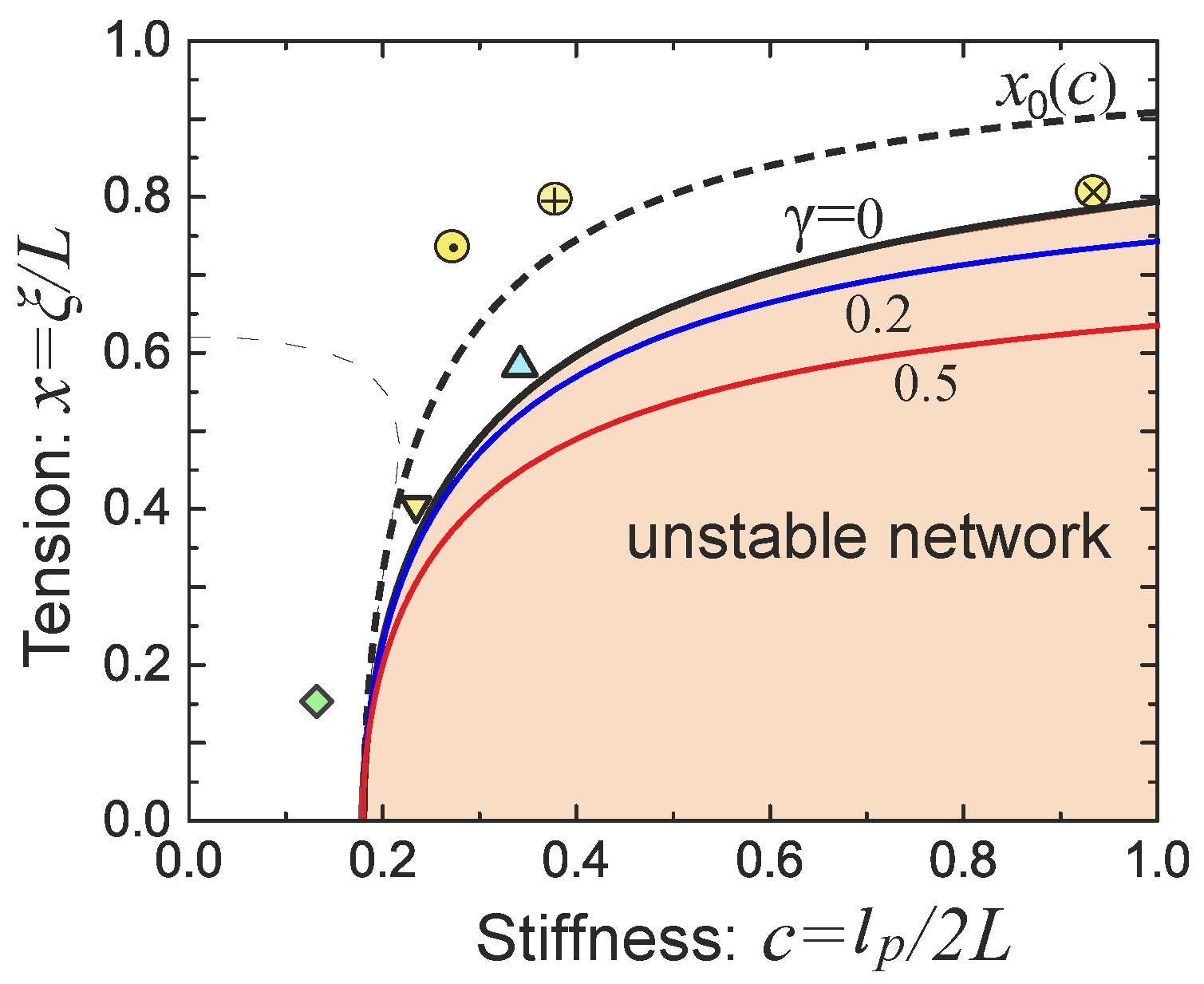
Blundell-Terentjev model [
14]. Equations (
7) and (
8) constitute the model of Blundell and Terentjev, which has an advantage of being fully analytical and capturing the right physics in both the low and the high temperature limits, as well as in tension and compression/buckling regimes up to the highly bent elastica limit [
21]. The force-extension relation is written as
where an internal elastic energy of a bent filament is obtained in the limit
for a fixed bending modulus
. As always for theories based on WLC, the finite extension limit gives the divergent force scaling:
. For a very short or stiff filament (
), the internal potential dominates over the entropic effects, and the filament can be treated as a rigid athermal rod. The zero-force average natural extension is
(e.g., at
we have
). As shown in
Figure 2b, if the ratio
is below the critical value of
, the minimum of the free energy will be located at
, i.e., the filament can be regarded as a flexible chain and reproduce a Gaussian limit upon first-order expansion (with the entropic spring constant
).
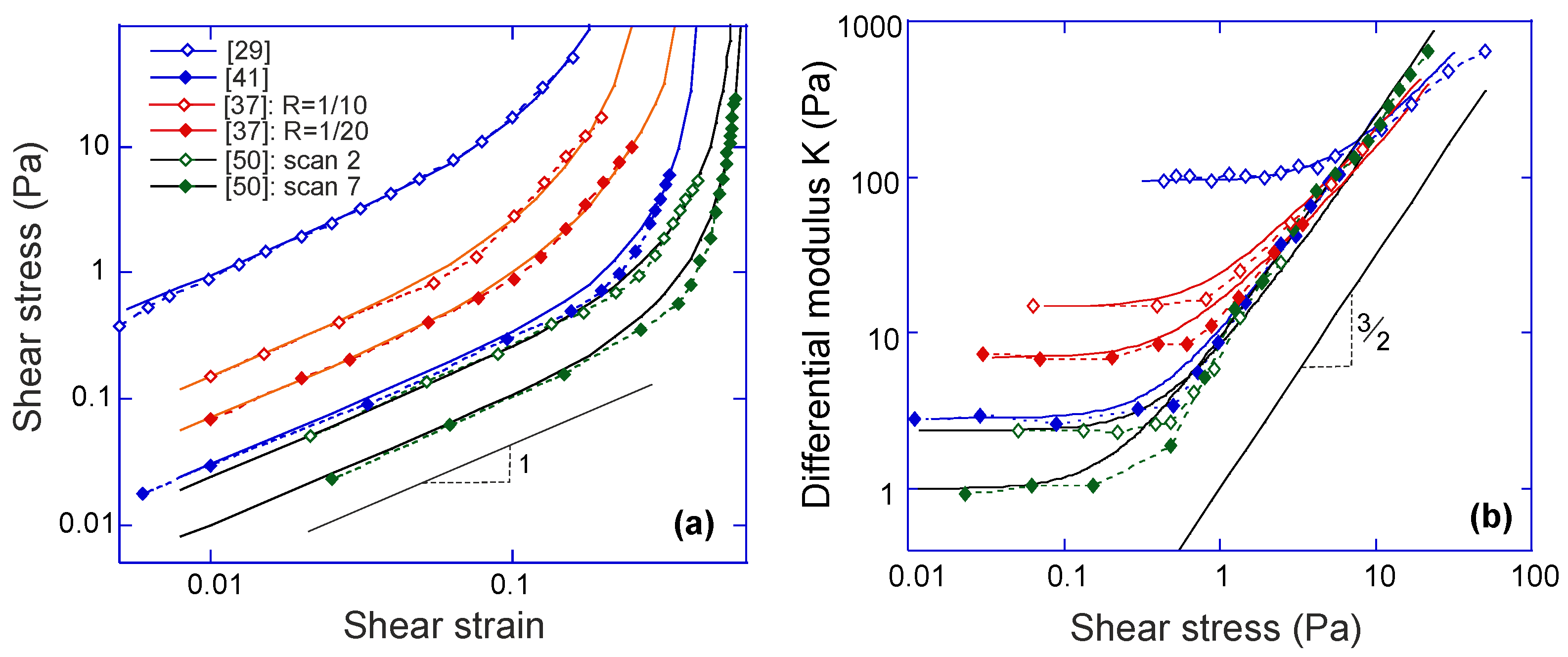
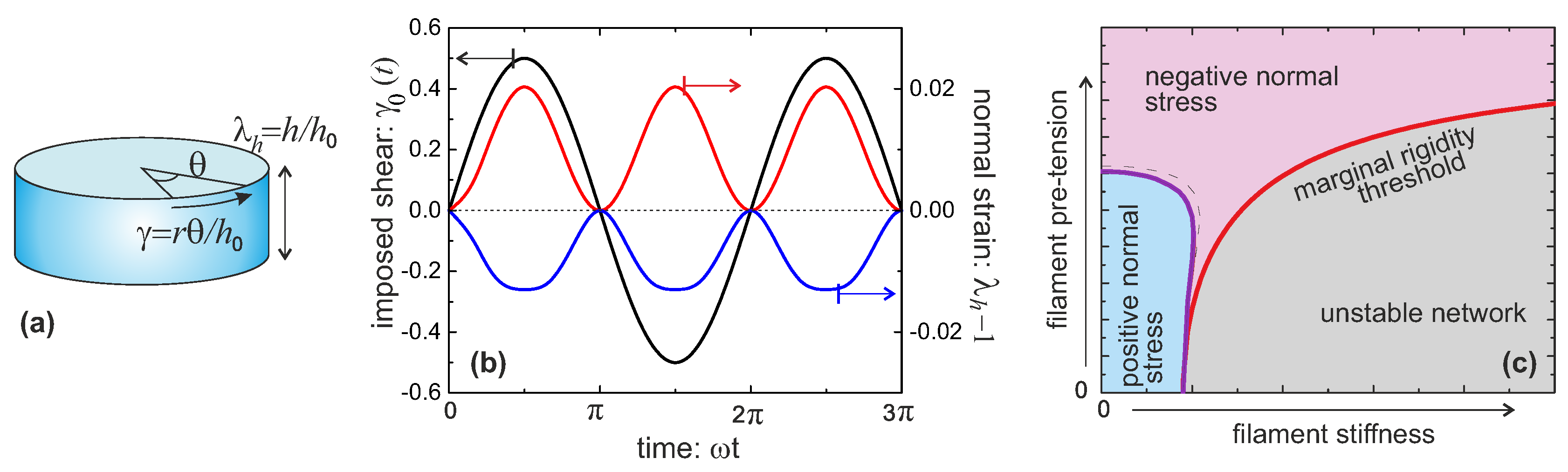
2.2. Mechanical Response of a Semiflexible Filament
The characteristic response of a chain or filament to a tensile force has been widely explored in the literature and remains the basis of determining the persistence length
, usually by fitting the M-S model. For the filaments that are stiff enough to have a non-zero average natural extension
, the response to a compression force also becomes relevant. When a rigid elastic rod is compressed, it will buckle if the compression force exceeds what is known as the Euler buckling threshold force
[
22]. Similar response also appears in the case of a semiflexible filament, although in this case the thermal fluctuations result in a finite range of buckling force, and the associated hysteresis. The free energy
given in Equation (
8) has the minimum located at
. The value of the free energy at
decreases with the increase of the compression force until it becomes a metastable state; in other words, the Euler buckling of a semiflexible filament is a typical first order phase transition [
19,
21]. The equilibrium threshold force for buckling is obtained as
where the bending modulus
κ is replaced by the persistence length
, and
and
are fitting (interpolation) parameters [
21]. The threshold force reduces to the classical Euler form for an athermal elastic rod with
.
So far we only discussed the physics of semiflexible filaments with a fixed contour length
L, which was the basis of the WLC statistical model. However, especially at large tension forces, the real filament may be elastically stretched by extending the bonds holding its monomers in a chain. In actin filaments, these bonds are the physical interactions (hydrophobic and hydrogen bonds) holding the G-actin units together, with a characteristic energy of ∼25–30
; a similar bonding energy is found to hold the
β-sheet peptides in an amyloid filament [
23,
24]. A stretched DNA double helix would elongate by modifying hydrogen-bond interactions between base pairs. In all of these cases, a finite stretching modulus was indeed detected by experiment.
When a semiflexible filament is highly stretched, its end-to-end factor will be close to unity, and an approximation can be implemented in Equation (
8):
, making the force-extension relation at
:
An effective spring constant in charge of the elasticity of a filament can be defined by
, producing
where the second version of the expression is obtained implementing the Equation (
14), with the shothand notation for the constant
, and the Euler critical force
. If in addition, the material making up the filament has its own intrinsic Young’s modulus
Y, an effective mechanical spring constant has to be added:
, with
a as radius of the filament. For actin filament of length
m, this spring constant is about 5 pN/nm, while a microtubule of the same length has a spring constant around 15 pN/nm. Considering the possibility of filament extension, the full length now becomes
under a large tension
f, and force-extension relation (with the approximation that
x is close to 1) can be expressed as
In other words, the mechanic response becomes softer than entropic one if the filament is experiencing a very large stretching force, while the entropic mode is softer if a filament is stretched by a smaller force. Similar expressions were obtained by Odijk [
17], Lipowsky et al. [
25], and Holzapfel and Ogden [
26].
There is a crossover region bridging the entropic stretching regime with the mechanical stretching regime, see
Figure 4. One can make an analogy of the extensible semiflexible filament to two elastic springs connected in series, giving the effective spring constant
as:
If the tensile stretching force is small, i.e.,
, the spring constant of the filament is approximately a constant
, which defines linear entropic response regime. If the tensile force is very large,
, the filament extension is dominated by the mechanical (enthalpic) stretching, while the entropic elasticity essentially diverges. If the width of ’nonlinear-entropic’ regime is sufficiently large, the entropic part in Equation (
17) scales as
; however, if the filament material is more easily stretched, the crossover could be narrow and much less distinct.
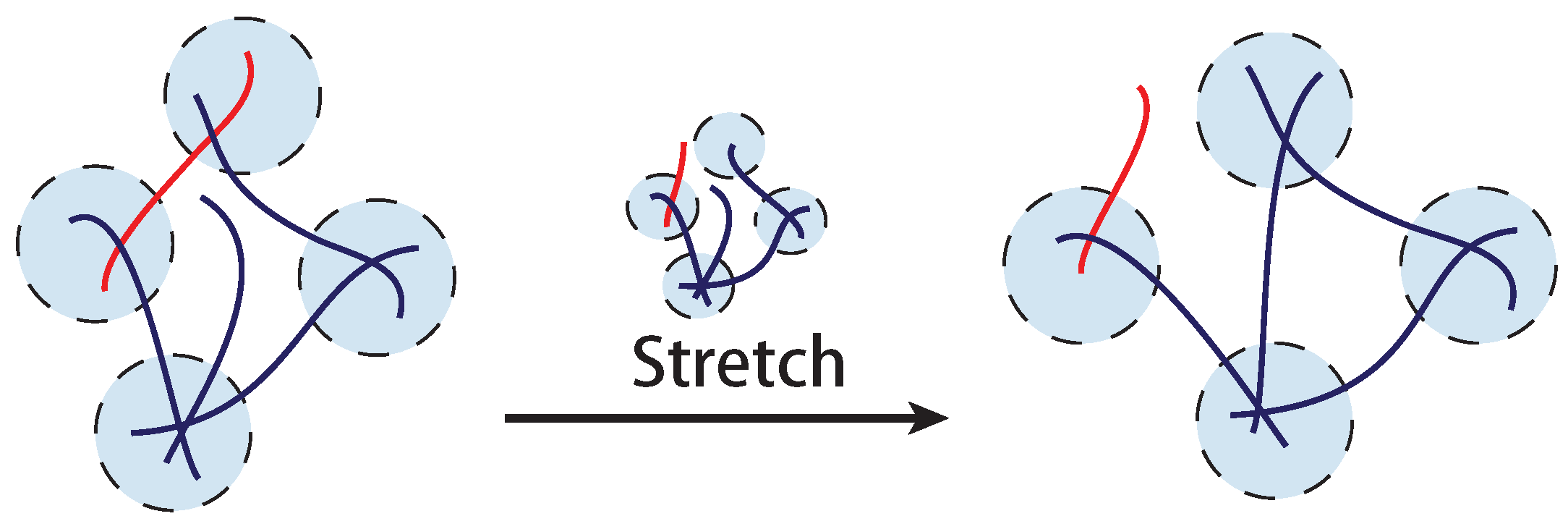
4. Transient Filament Network with Breakable Crosslinks
In contrast to a permanently crosslinked network such as classical rubber, biological crosslinks in a filament network can be dynamic. The classical collagen network is held together by hydrogen-bonded triple-helical segments that are known to dynamically respond to an applied tension. The F-actin network is often held together by molecular motors (such as myosins) or physically bonded proteins (such as filamins). Self-assembled biological filaments themselves can be broken from or re-crosslinked to the network. In synthetic networks, one frequently finds dynamic crosslinks formed by physical bonds (such as hydrophobic interactions or hydrogen bonds,
Figure 12). Actually, the concept of a transient crosslink here is very similar to the “trap” defined in soft glassy systems [
64,
65], where semiflexible chains are assumed to be trapped in a glassy environment, and also to the dynamic “entanglement” in concentrated polymer solutions [
1].
Due to the transient feature of such crosslinks, the resulting network shows characteristic rheological properties, such as a high stress relaxation at given constant strain, a yield stress upon ramp deformation, or the elastic-plastic transition. As transient filament network is a better analogy to the filament network in many biological systems, there are several relevant theories developed in the recent years from different viewpoints.
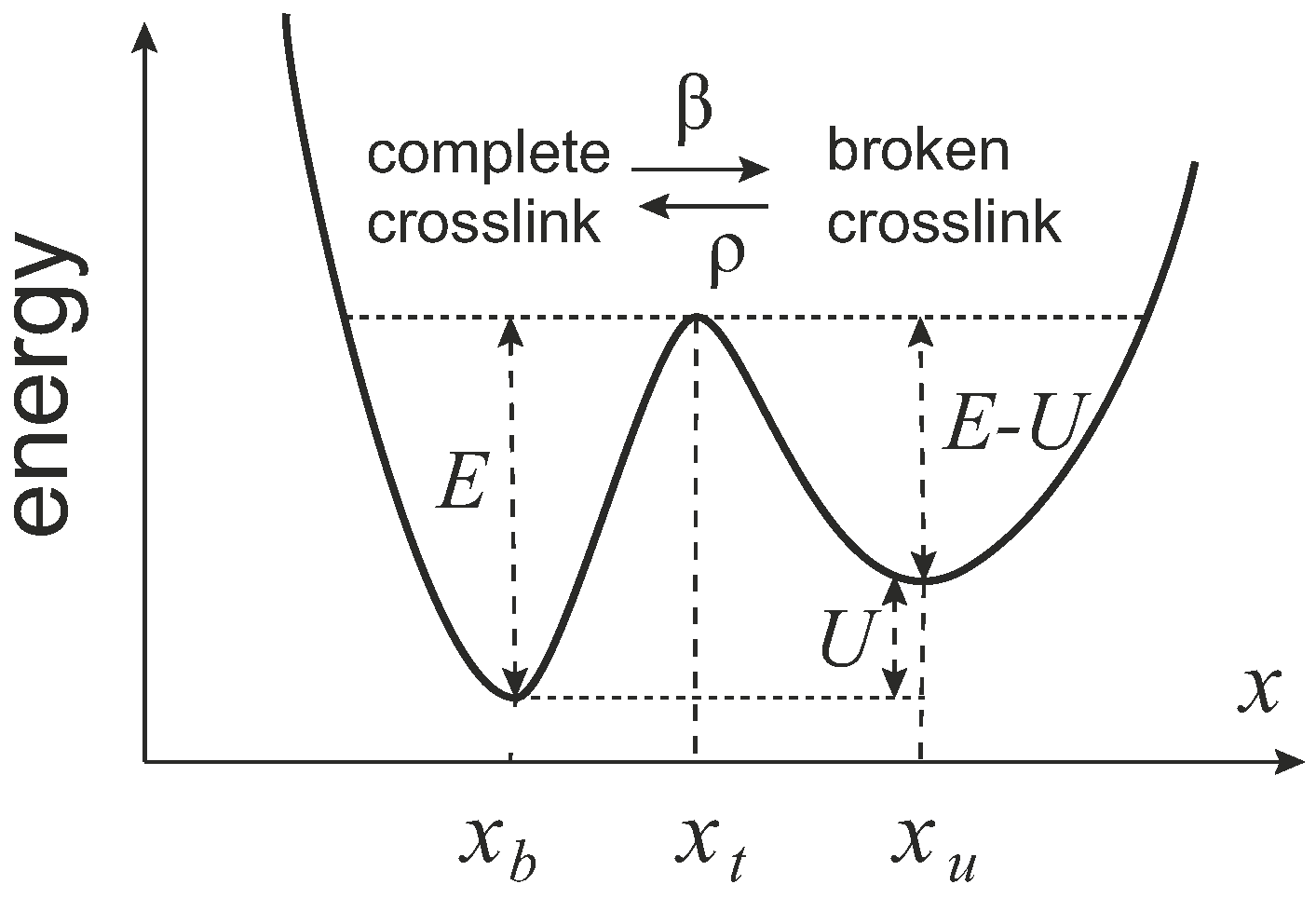
In 2007, Kroy et al. proposed the glassy wormlike chain model [
66,
67], and then used it to study the viscoelastic properties of “sticky filament network” [
68]. One assumes there is a certain crosslink breaking rate
β, and also a rate of crosslink re-connection,
ρ. In adiabatic approximation, the rates can be expressed by the activation laws (
Figure 13):
where
is an intrinsic time scale of attempts due to the thermal fluctuations,
E is the energy barrier for breaking the crosslink,
U is the energy penalty for the broken crosslink,
f is the pulling force,
is the coordinate of crosslinked state,
is the coordinate of the barrier, and
is the coordinate of the uncrosslinked state (crosslink broken). The fraction of the complete crosslinks is interpreted via the effective change in filament length:
(the fraction of broken crosslinks is
), where
is the minimum contour length of the filament between two crosslinks, and
is the contour length of the filament between crosslinks at time
t, which can change with the breakage and the formation of the crosslinks. Then the evolution equation of the fraction of the complete crosslinks is
Assuming that the linear shear modulus is proportional to the density of active crosslinks, the main difference in the complex modulus between the ordinary filament and the glassy semiflexible one in transient network is in the relaxation time
, which is a constant
in the first case, while for glassy semiflexible filament it depends on time via the
relaxation:
with
as wavelength of the
nth mode. Microscopic complex modulus for transverse point excitations of a generic filament element was defined in this model as
where
f is filament tension, and
the Euler buckling force for rigid rod as defined previously.
In 2008, Lieleg et al. [
69,
70] performed a rheological experiment on actin network crosslinked by rigor heavy meromyosins (HMM), which is yet another kind of transient crosslinks. They reported local stress relaxation and energy dissipation in an intermediate elasticity-dominated frequency regime. In order to explain this, they developed a semi-phenomenological model by introducing a breakage rate of the crosslinks,
, which is a simpler version of the glassy filament network model above. By assuming a breakage rate of crosslinks, the number of the crosslinks decays exponentially with time as
, which can be translated into the frequency space by Fourier transformation,
, where
ω is the oscillation frequency in the experiment. With the rubber-elasticity relation
, the storage and the loss modulus are written as functions of the oscillation frequency:
where the terms with fitting parameters
a and
c arise from the breakage of the crosslinks, via
, the
terms with parameters
b and
d arise from the fluctuation of single filaments [
30,
71], and the time scale of the relaxation mode is set by
which is dependent on the solvent viscosity. There is a characteristic frequency in experiments,
, at which the dissipation has a minimum. When the frequency
ω is larger than
, the storage modulus is almost a constant, while decreasing at lower frequencies due to the breakage of crosslinks at longer time scales. This semi-phenomenological model provides a basic understanding in the viscoelastic response of a transient network at different time scales.
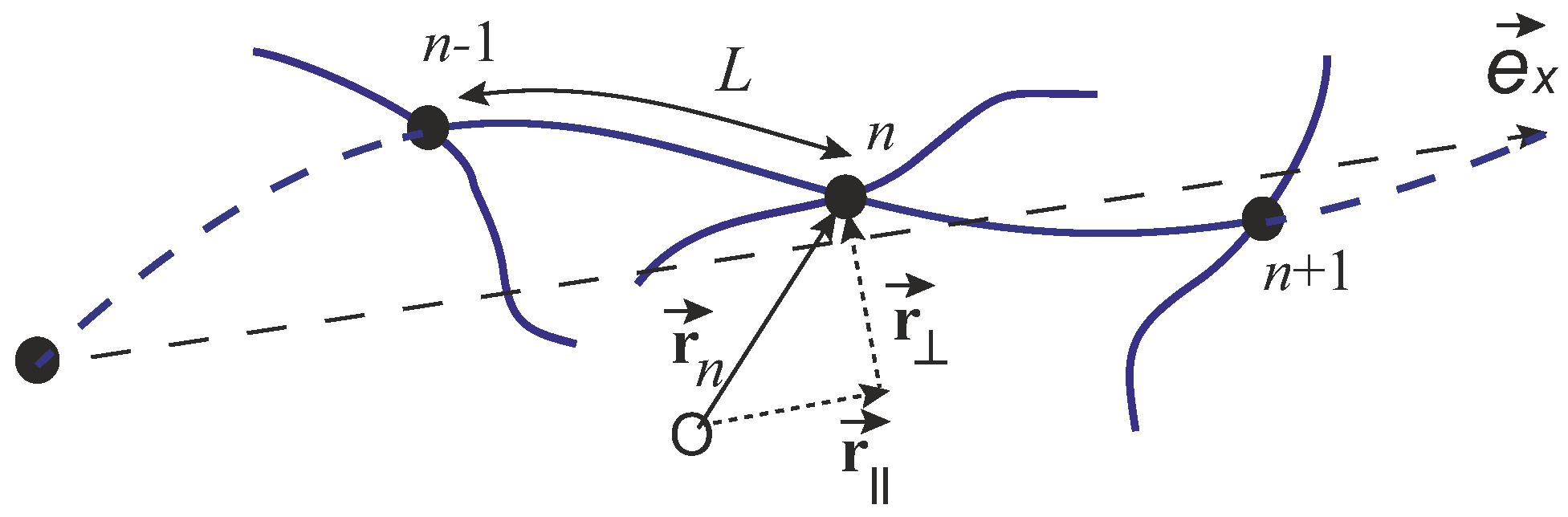
In 2010, Broedersz et al. [
72] proposed a model of transient filament networks based on crosslink-governed dynamics, where Monte Carlo simulations and an analytic approach are combined for explaining the rheological properties of the network. There are three time scales in the model: the thermal relaxation time for chains after breaking off a crosslink,
, the re-crosslink time,
, and the crosslink breakage time,
. In this model they are assumed to obey a relation:
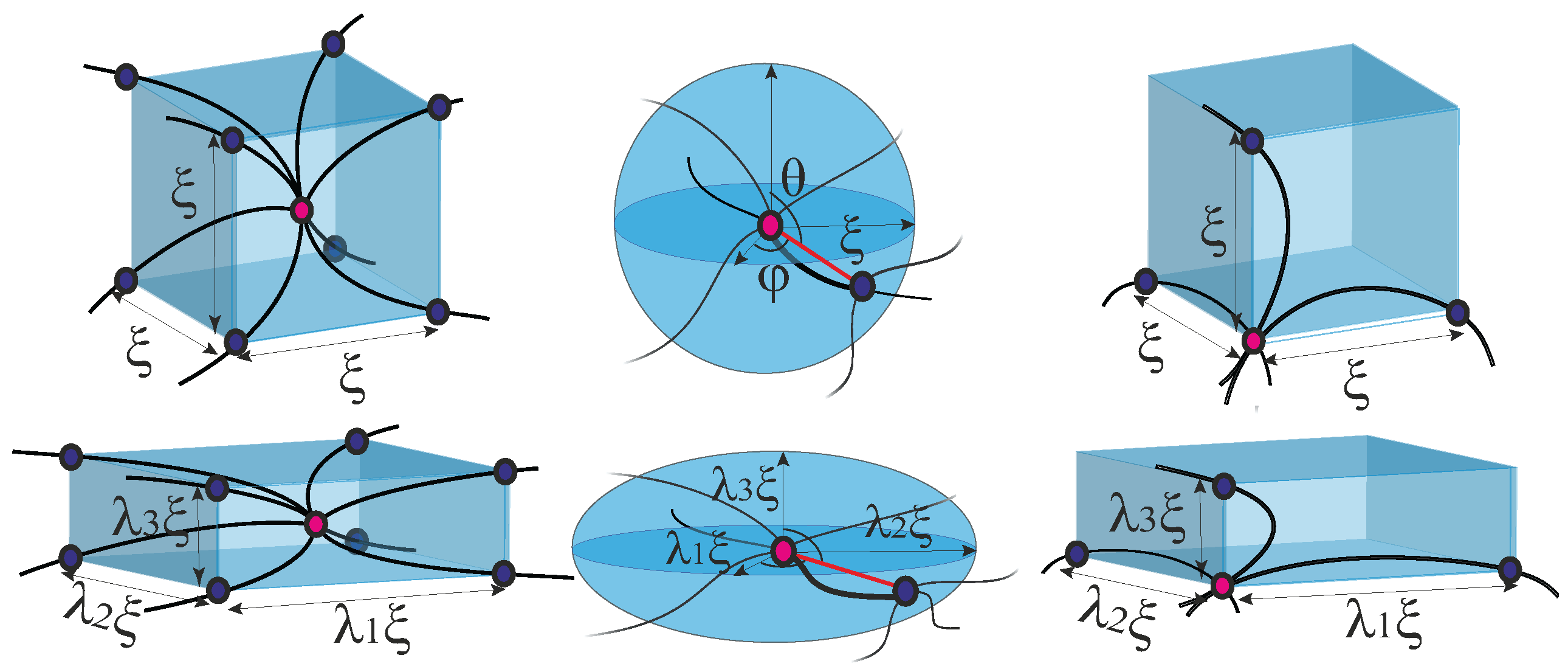
. The elastic energy of the network can be expressed as a sum of the bending energy and the stretching energy of all the subfilaments connecting the pairs of crosslinks:
where
L is the length of a strand between the two crosslinks, which is assumed to be the same in the whole network, the sum is taken for all the crosslink positions
(with the “mesh size”
), and
is unit tangent vector of n
th crosslink (sketched in
Figure 14. In this case, the n
th segment curvature is
, defining the first term (the bending energy). The second term in Equation (
42) denotes for the elastic energy of the spring, where the stretching modulus
is directly related to the discussion below Equation (
11) in
Section 2. In order to calculate the polymer displacement due to the unbinding and rebinding, they separate the local equilibration step into a move to the mechanical equilibrium position, with an added stochastic contribution around this equilibrium. Mechanical relaxation step from the initial position
to the local equilibrium position
is determined by
Actually, this numerical method can describe non-affine deformation in filament networks. By solving Equation (
43) discretely and taking the continuum long-wavelength limit, the leading order evolution equations with thermal fluctuations are
where
x denotes for the position of the cross-linker along the average direction
of the relaxed filament, and
and
are the longitudinal and transverse deflections of the filament with respect to its average direction. The noise term
possesses both thermal effects and also the local buckling contributions from thermal compression. Note that the noise actually depends on the local state of the polymer and couples the longitudinal and the transverse motion of a filament as described Equation (
44). In the limit of small stretch modulus,
, Equation (
44) decouple and the resulting transverse contribution to the power spectrum (
with
as extension of length) approaches
, in agreement with simulation. In this limit, the transverse bending dynamics are effectively those of a stiff filament fluctuating in a viscous solvent, which gives
[
30,
71]. Recently, this Langevin-type model was applied to active network systems [
73].
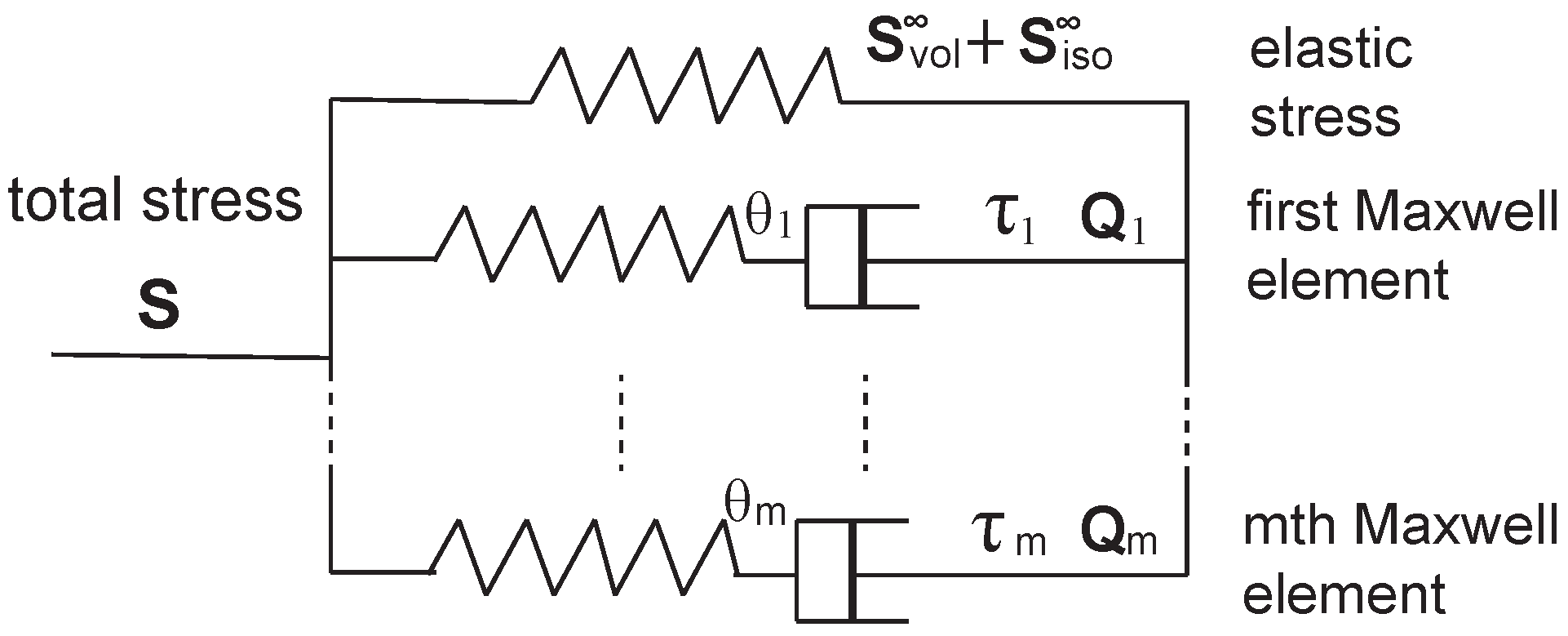
In 2013, Unterberger et al. [
74] applied a generalized Maxwell model to the transient filament network for describing its viscoelastic properties, as shown in
Figure 15. In addition to the pure elastic part of the material, the model adds another
m parallel Maxwell elements, each of which consists of an elastic spring and a viscous dashpots. In studies of viscoelasticity and relaxation, this is a common method of introducing the spectrum of relaxation times that is often necessary to describe complex relaxation processes.
Rather than using Cauchy stress
, as defined in Equation (
26), they applied the second Piola-Kirchhoff stress,
, which is related with Cauchy stress as:
. There are three distinct contributions to the total stress of the material, which is expressed as
where the first term denotes the equilibrium volumetric part, the second term denotes the equilibrium isochoric-elastic part, and the third term reflects the isochoric relaxation (non-equilibrium) part. The first two contributions are typical elastic ones. Here we will only focus on the third part, which is closely related to the “transient” characteristics of the filament network. The volume-preserving non-equilibrium stress tensors
, are defined as a function of internal variables,
, through the dissipative potential,
,
where
is the right Cauchy tensor
,
is the isochoric part of the right Cauchy tensor, and
is the isochoric part of the
Γ. A set of differential equations and initial conditions for the non-equilibrium part of the
nth Maxwell element,
is introduced to describe the transient property of the material, as
where
are non-dimensional free energy parameters, related to the relaxation times
. Convolution integrals provided a solution for Equation (
47) of the
nth Maxwell element as
which basically describes how the energy of the material dissipates with time due to the transient crosslinks, and the time scale for the
nth Maxwell element is given by its the relaxation time,
.
One could instead apply a dynamic theory of transient rubbery networks [
75,
76], which was based on classical Gaussian (neo-Hookean) chain response, to the case of filament network. This strategy is similar to the theory of equilibrium filament network, Equations (
25) and (
27), where the specific form of a non-linear elastic response of the filament strand was implemented in a continuum model. By combining the free energy form of a permanent filament network with the transient characteristics of its crosslinks, we could work out the time-dependent response to various modes of dynamic deformation.
We shall use the crosslink dynamics very similar with those in glassy wormlike filament network model, Equation (
37). The rate of a chain to break from a crosslink depends on the force
f acting on the chain as:
, where
is the breakage rate for a force-free chain, and
a is the monomer size and taken as a characteristic length of a bond. As in Equation (
37),
. The dangling chains are able to re-crosslinked into the network with a rate
ρ, determined by the barrier
(cf.
Figure 13), although here we will assume that the free filament would be able to relax and re-crosslink in the force-free state (which is the basis of the stress relaxation in the material).
Suppose the total number of filaments in the network is
, with
of them crosslinked at the time
;
chains are un-crosslinked. The breakage rate of the crosslinks is a function of the local force acting on the given chain, rather than as a constant,and thus can be only defined locally and dynamically: it depends on both the current state at time
t and the crosslinking state at time
that defines the reference state for the given chain. We denote the breakage rate of the crosslinks at time
t formed at time
as
. After an infinitesimal time interval,
, the number of the originally crosslinked chains, will decrease to
, where
denotes the breakage rate at time
. Meanwhile, there will be
new filaments re-crosslinked into the network in the same time. By combining these two parts, the total number of the crosslinked chains at time
then becomes:
. After another time interval, i.e., at
, the number of the crosslinked chains surviving from the beginning decreases further to
; the number of chains which were crosslinked during the first time interval decreases to
; finally, the number of newly crosslinked chain during the second time interval is
. The total number of filaments that are crosslinked (i.e., a part of the elastic network) at time
is
. By repeating this process, the total number of the crosslinked chains after
m time intervals will become
which can be expressed in continuous form as
where the first term on the right hand side represents the number of the originally crosslinked chains surviving from the beginning, and the second term represents those chains that were crosslinked between
and
t, and still remain crosslinked [
76]. For the first term, the elastic reference state remains the original undeformed configuration of the network at
. In contrast, the tension force that determines the rate
in the second term depends on the reference state at a time
when the particular filament was re-crosslinked into the network at its force-free conformation.
As the reference states of the filaments are defined according to when they are crosslinked, the elastic energy density of the system is
where the free energy form of the permanent network,
, can be chosen from the continuum models showed in the previous section, e.g., Equation (
25). As with the rate of breaking, the expression for
depends on the deformation at time
, measured with respect to the reference state (at
) when the fraction of the chains that is counted towards this contribution was re-crosslinked. The corresponding deformation tensor, measured at
with respect to the reference state at
is:
. With the formal expression for the dynamic free energy of the material, one can obtain the stress-strain relationship and other rheological characteristics of the transient network, once the regime of imposed deformation is specified. Two simple applications of the theory are explored below:
Stress relaxation. If the transient network is subjected to a constant uniaxial extension with stretch ratio
λ, imposed instantaneously at
(stretch ratios along the other two orthogonal directions are
from the incompressibility condition), then the tensile stress along the stretching direction decays with time purely due to the breakage of crosslinks: any crosslinks that are re-established during this process remain in the force-free state and the second term in Equation (
51) does not contribute to the response at all. The rate of this exponential decay depends on
λ but remains constant over the course of relaxation:
where
is the instant stress at
, which is equal to
. The breakage rate of the crosslink in a primitive cubic cell of a 3-chain model,
, can be calculated by taken the force in the equation as
, which is the maximum force acting on the three orthogonal chains of a crosslink. The explicit form of the force can be calculated by Equation (
12). Assuming relatively stiff filaments with
, originally crosslinked near their force-free point (as most biological filaments studied in
Figure 10 appear to be), the rate of relaxation takes the form
where the additional exponential factor arising from the tensile force on a filament is approximated for small deformations
to allow easy linearization. This characteristic exponential stress relaxation is a feature of all transient networks with a single value of the bare rate of crosslink breaking (a single value of the breaking barrier
E). This is often the case in biological networks, as well as in the new vitrimer materials [
77], while in physically crosslinked synthetic polymer networks one often finds a broad distribution of barriers, and the breaking rates. In such systems, the observed stress relaxation is usually the stretched exponential [
76].
Ramp deformation. This deformation regime occurs when the material undergoes a continuously increasing uniaxial stretch, i.e., the stretch ratio changes with time as
, with a constant rate
. Contributions from the re-crosslinks can be usually ignored, since this rate remains constant while the rate of breaking continuously increases with time [
78]. In this case the tensile engineering stress can be obtained as
The 3-chain model is chosen for
as an example. The network can respond in three distinct regimes: increase stress as an elastic solid, plastically flow as a fluid, or exhibit a necking instability when the two regimes coexist along the length of the sample, as illustrated in
Figure 16a. The transitions between these regimes depend on how the imposed strain rate
compares with the bare breaking rate
(at zero tensile force on a crosslink). These regimes arise from the competition between the intrinsic non-linear elasticity of the material and the plasticity due to the breakage of the crosslinks.
Figure 16b illustrates a typical predicted response. When the reduced strain rate is large, the material just responses as pure elastic one as the chains have no time to break from the crosslinks. Note that the stress rapidly falls to
zero when the strain approaches its critical value where the free energy of the network diverges, as the filaments experience infinitely large forces which break the filaments from the crosslinks. When the reduced strain rate is small, the stress of the material first increases due to elasticity of the network with its crosslinks still largely intact, and then reaches the yield point, following which the stress decreases (the network fluidises) due to an increasing number of broken crosslinks. When the reduced strain rate is moderate, the stress of the material first increases then decreases after a yield poin—however, the stress of the material will increase again due to a non-negligible number of the crosslinks surviving from the beginning and the increasingly high force some of these filaments exert. In this intermediate case the material shows the classical necking instability, where a strongly stretched region (plastic zone) and a weakly stretched region (elastic zone) coexist at the same level of stress across the sample.





















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}