
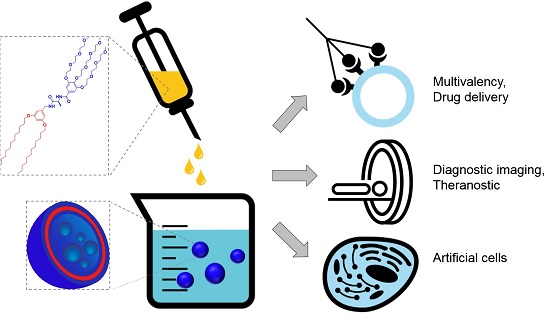
Vesicles from Amphiphilic Dumbbells and Janus Dendrimers: Bioinspired Self-Assembled Structures for Biomedical Applications


Abstract
:
1. Introduction
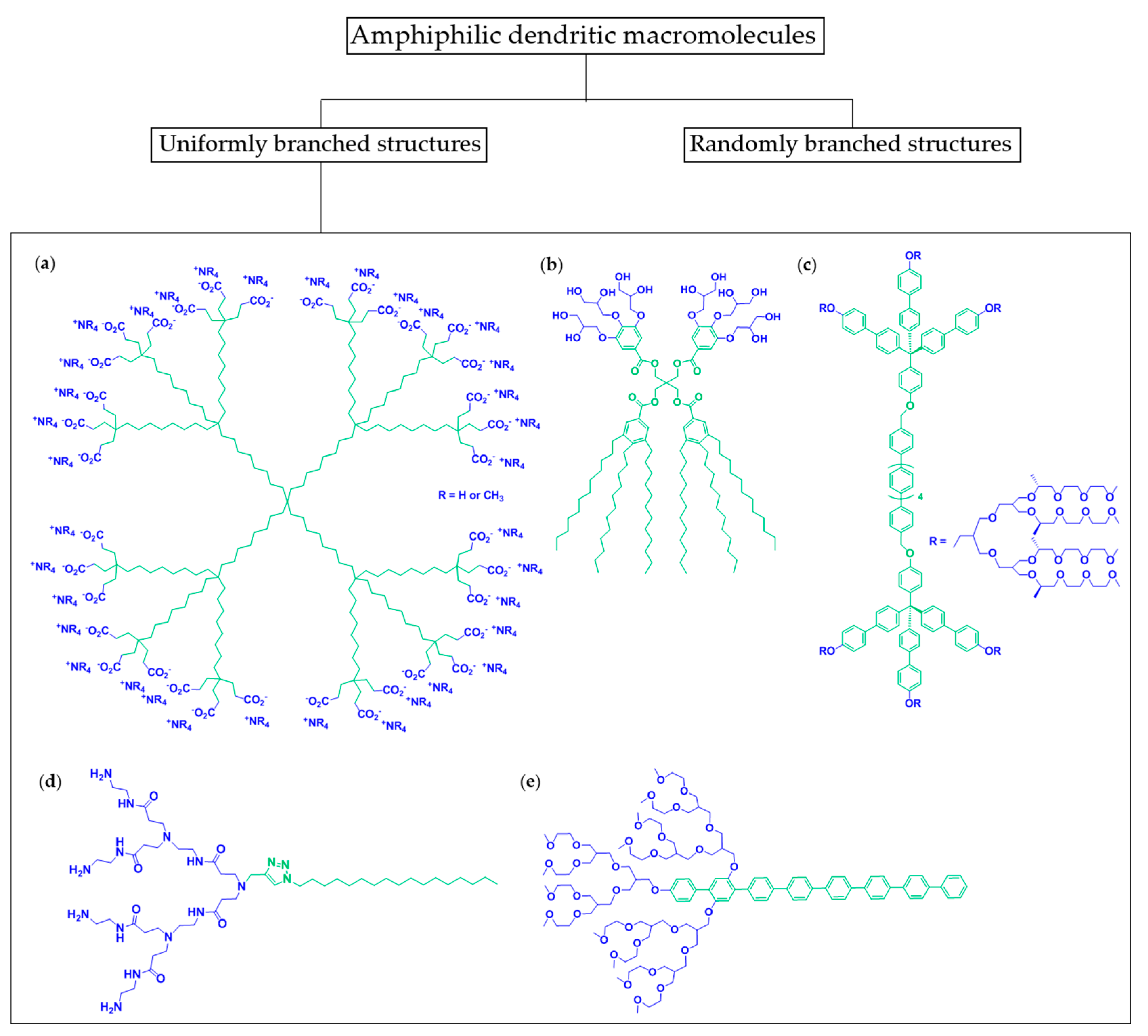
2. Amphiphiles of the Dendritic Family
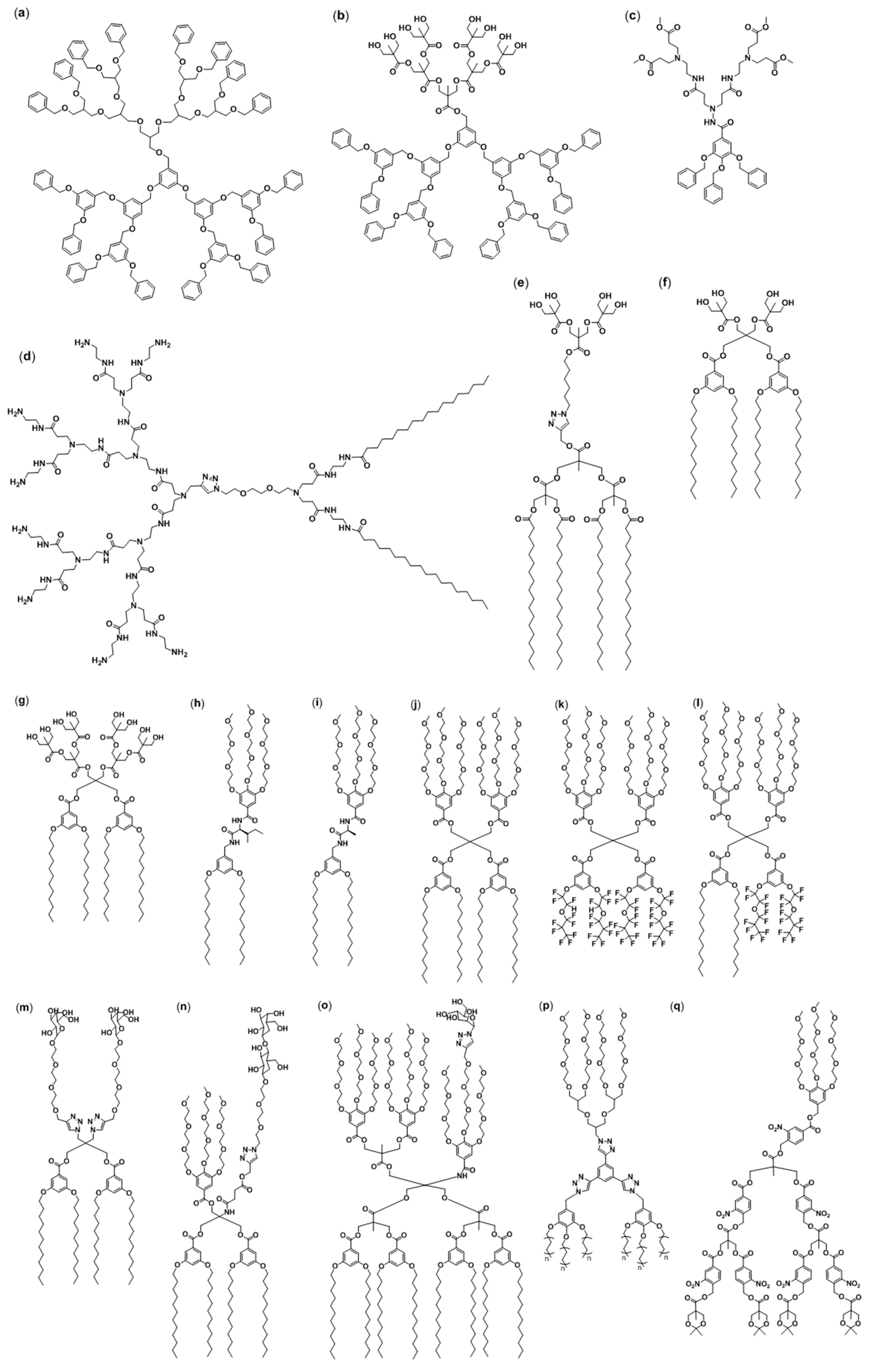
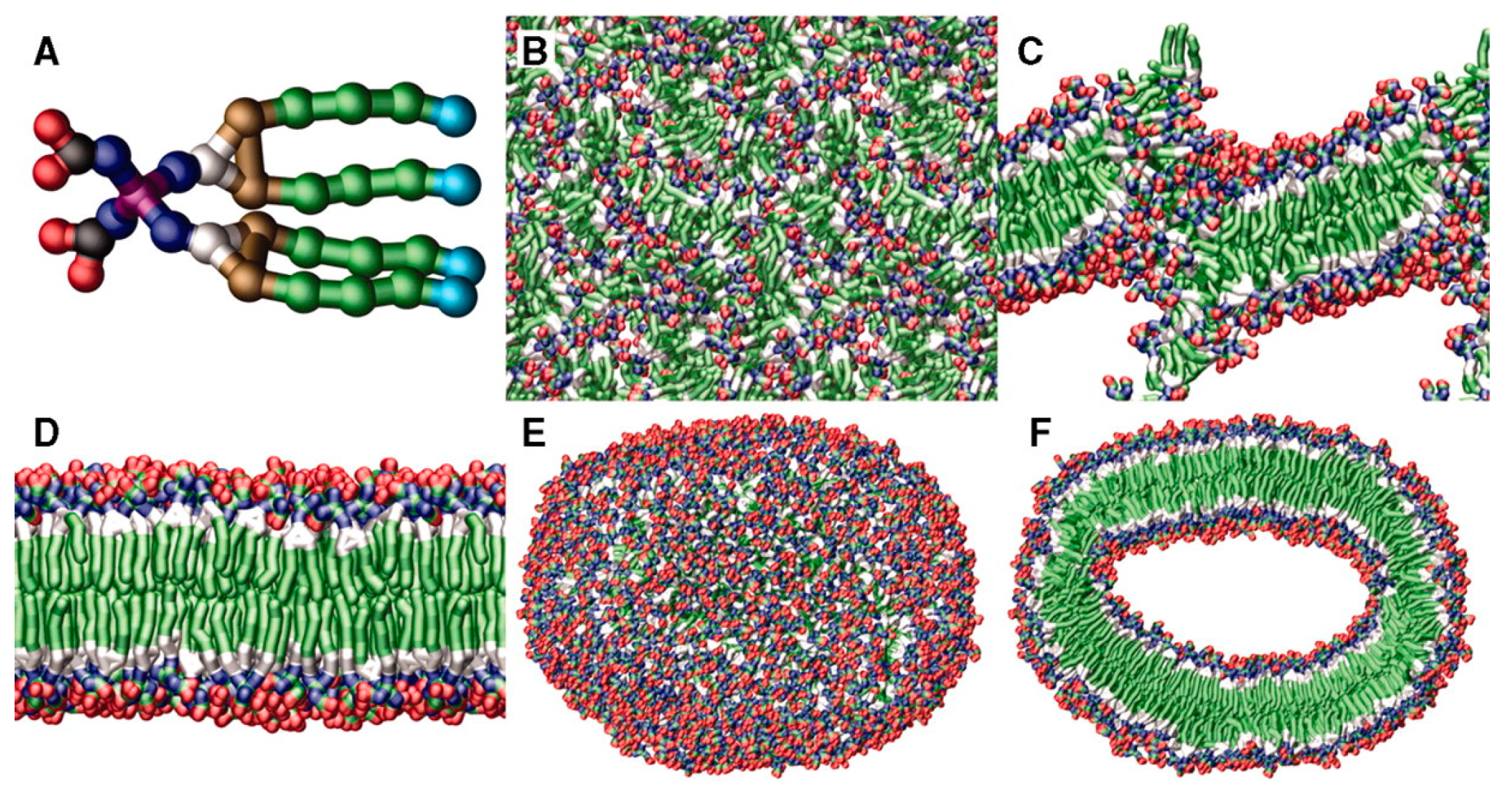
2.1. Janus Dendrimers
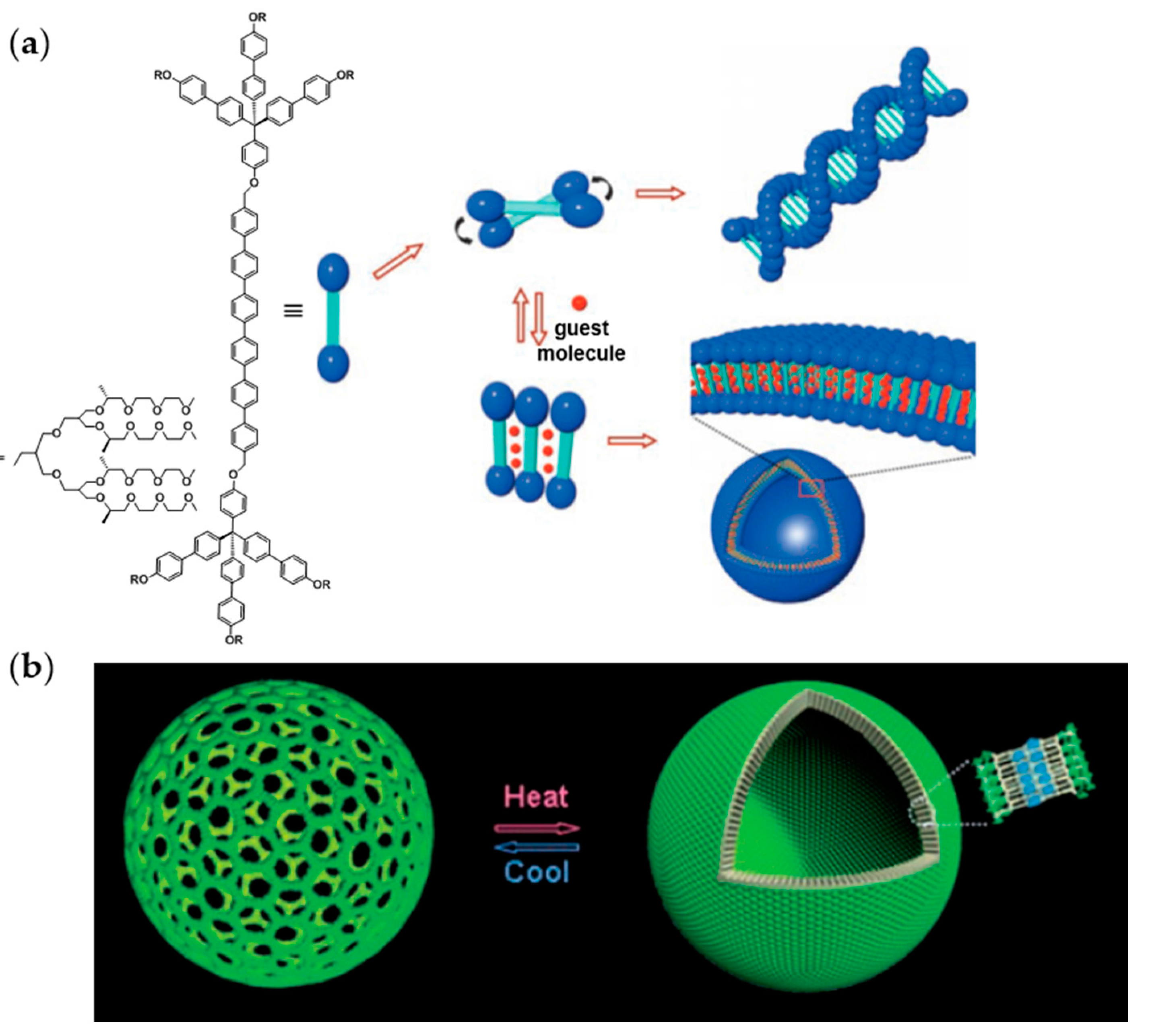
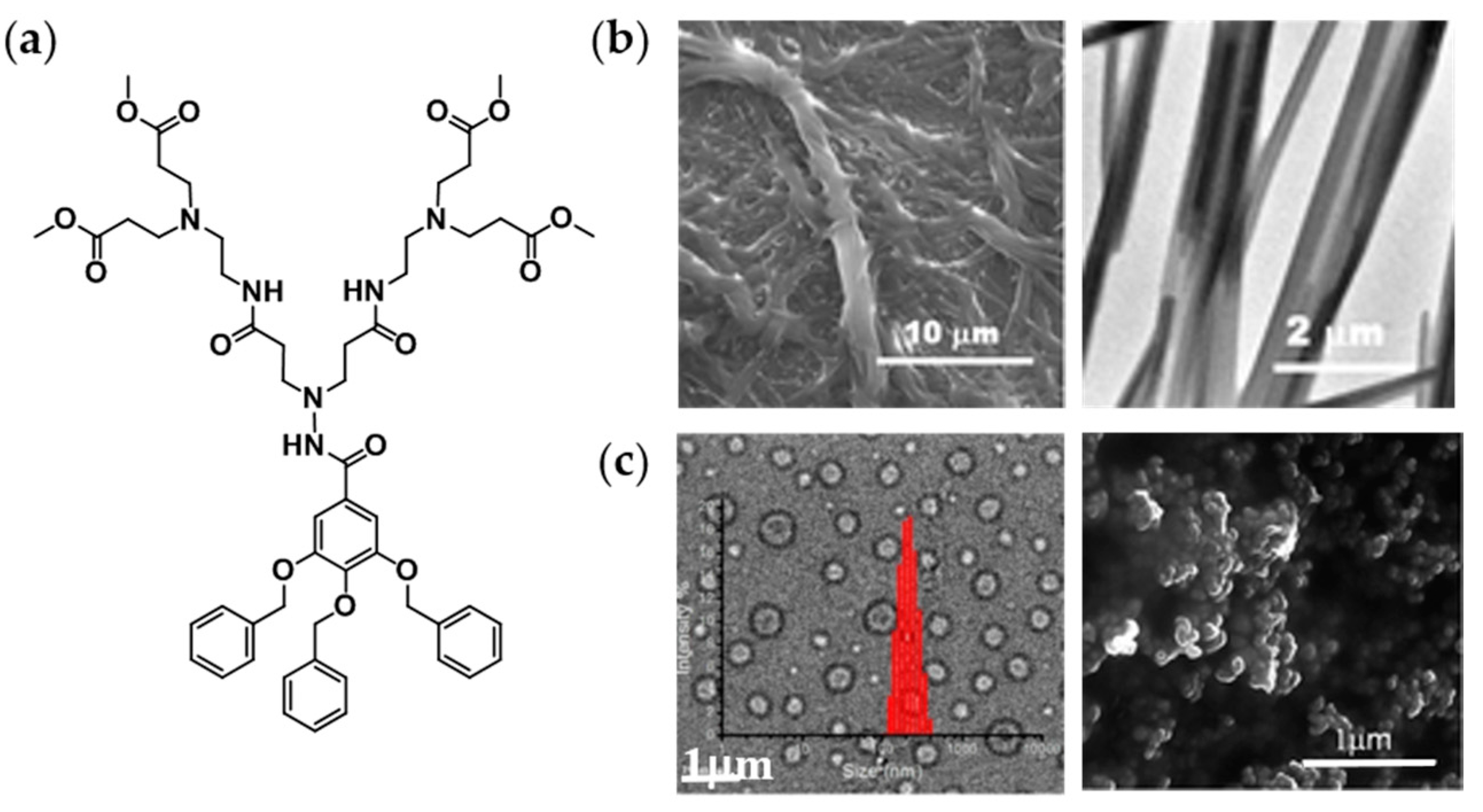
2.2. Amphiphilic Dumbbells
3. Vesicles Formation
3.1. Non-Covalent Interactions and Geometrical Considerations
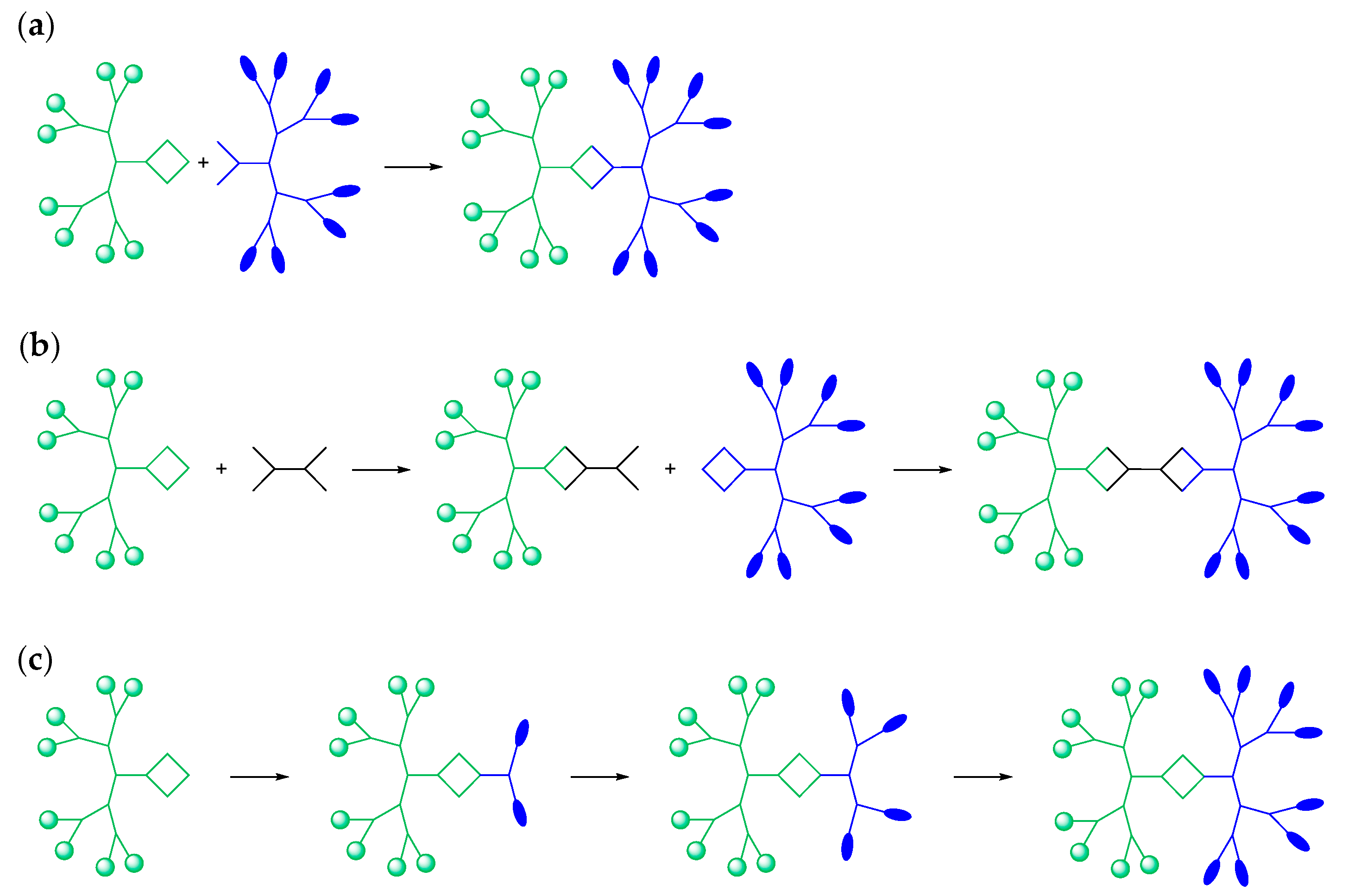
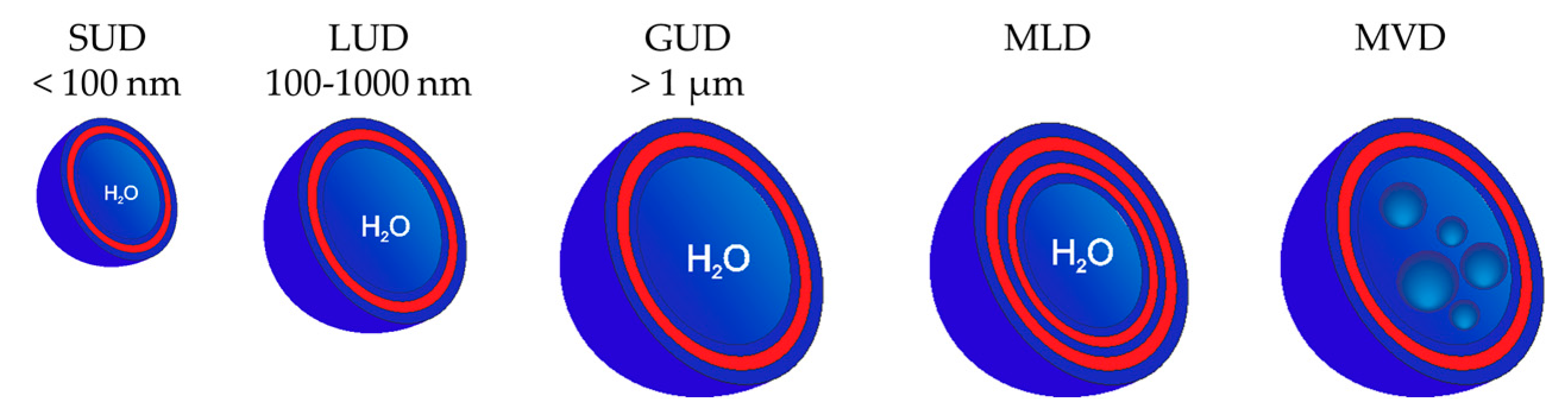
3.2. Classification of Dendrimersomes
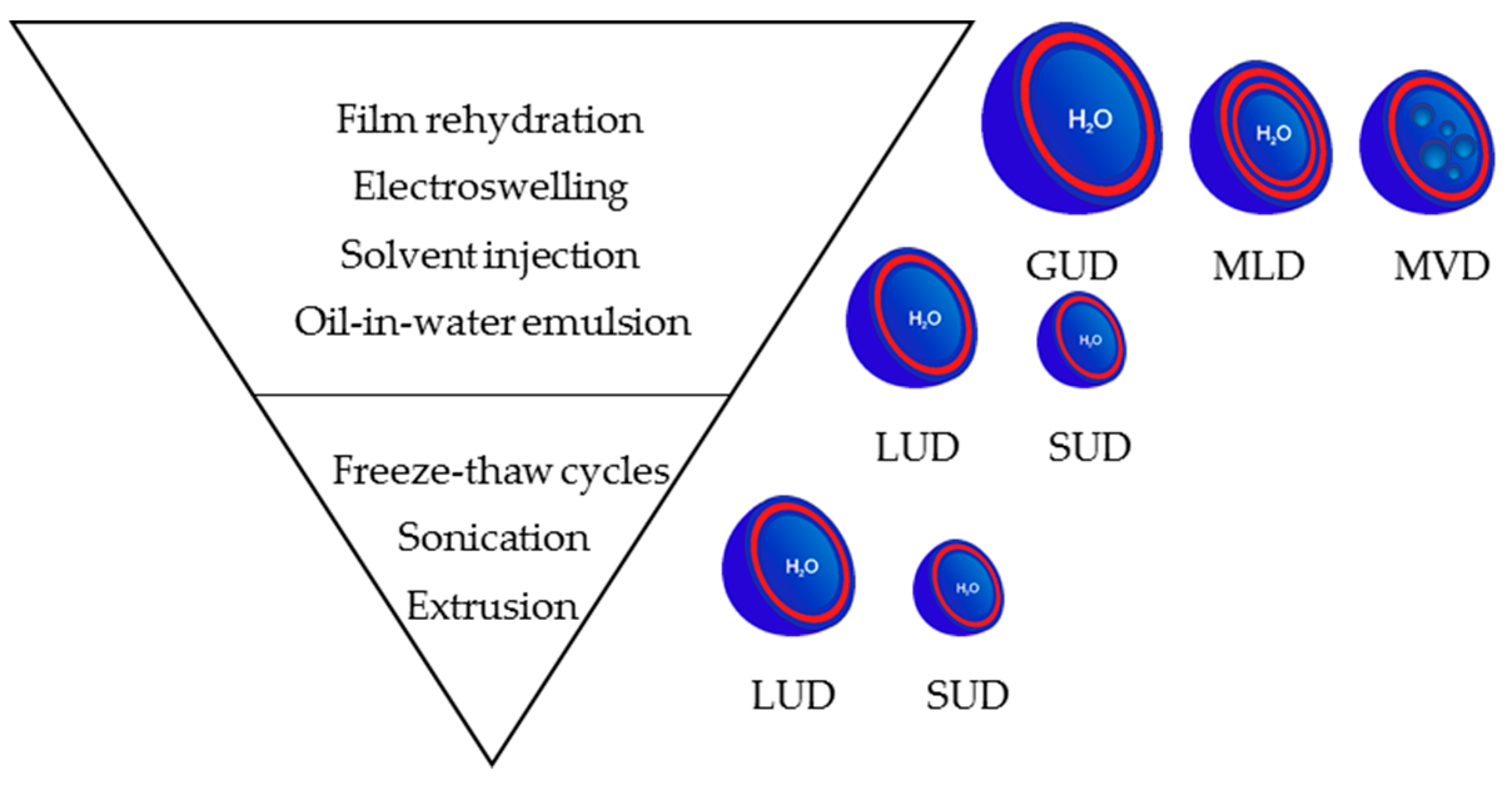
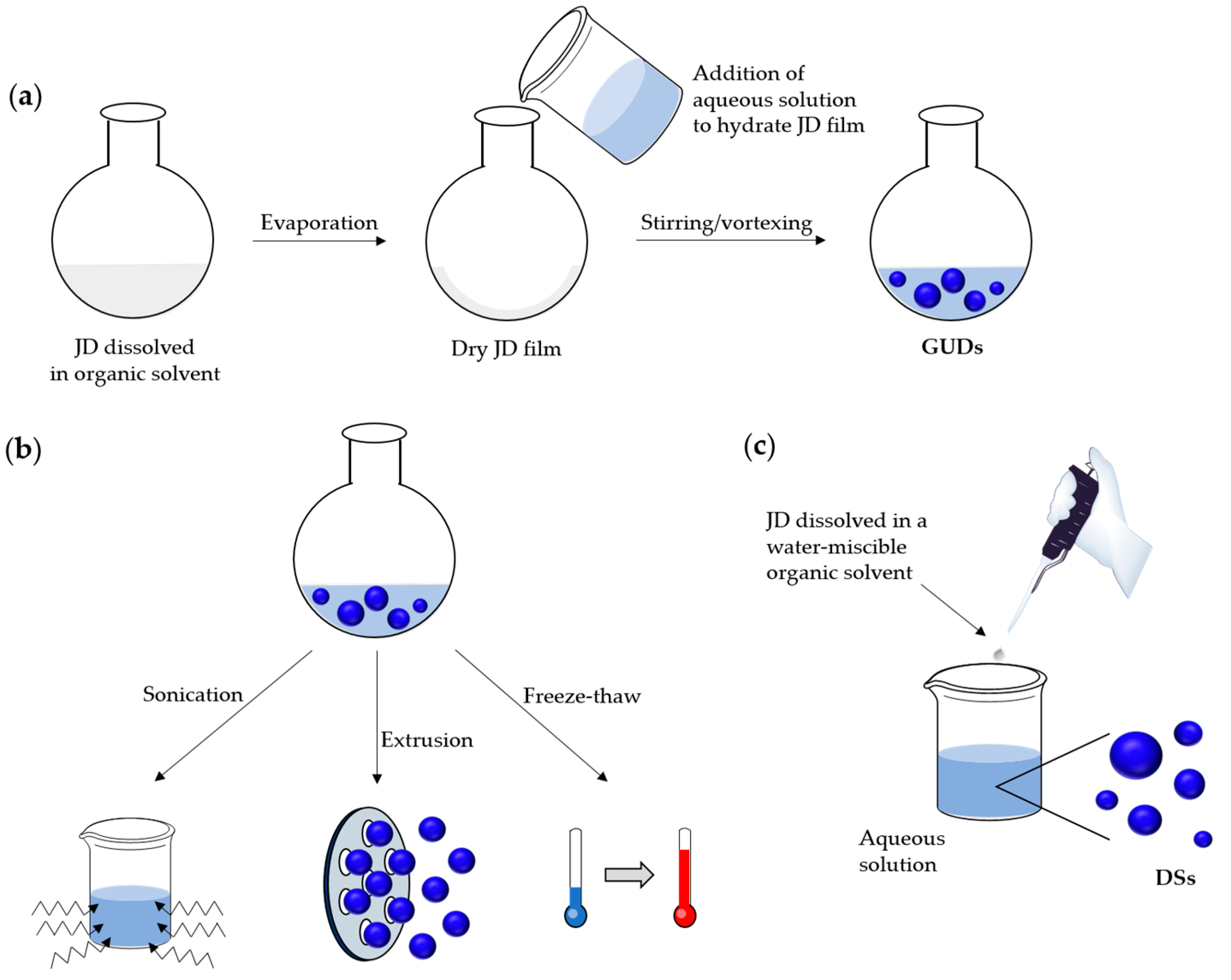
3.3. Preparation Methods for Dendrimersomes
3.3.1. At the Microscale: Giant Vesicles
Preparation by Film Rehydration Method
Preparation by Electroswelling Method
3.3.2. At the Nanoscale
Preparation by Film Rehydration Method Followed by Post-Preparation Steps
Solvent Injection Method
Emulsion
3.4. Parameters Influencing the Formation, Structure, Stability and Biocompatibility of Vesicles
3.4.1. Formation of Vesicles
Influence of JD Structure
Solvent Influence
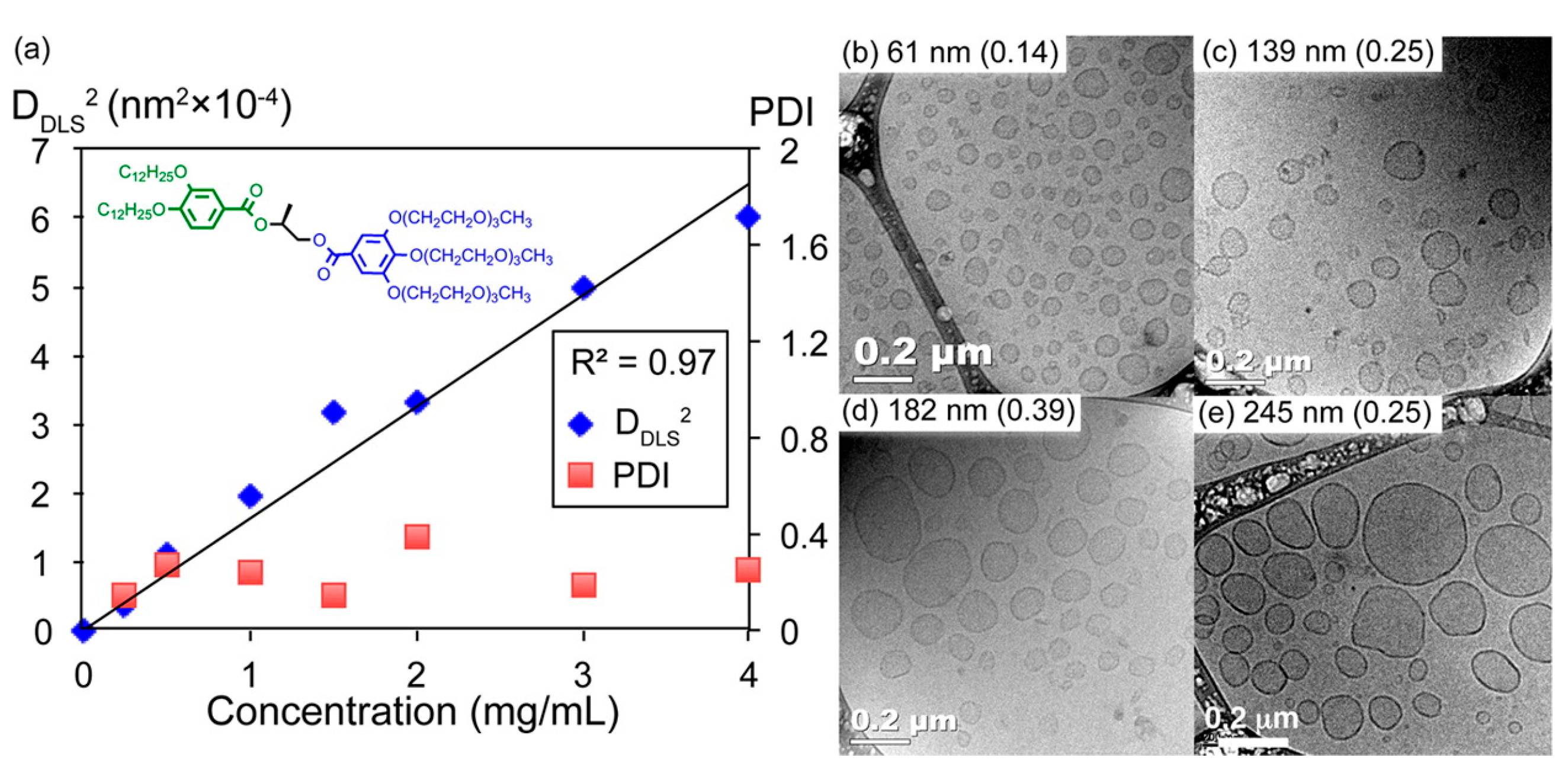
Critical Aggregation Concentration
3.4.2. Size of Vesicles and Membrane Characteristics
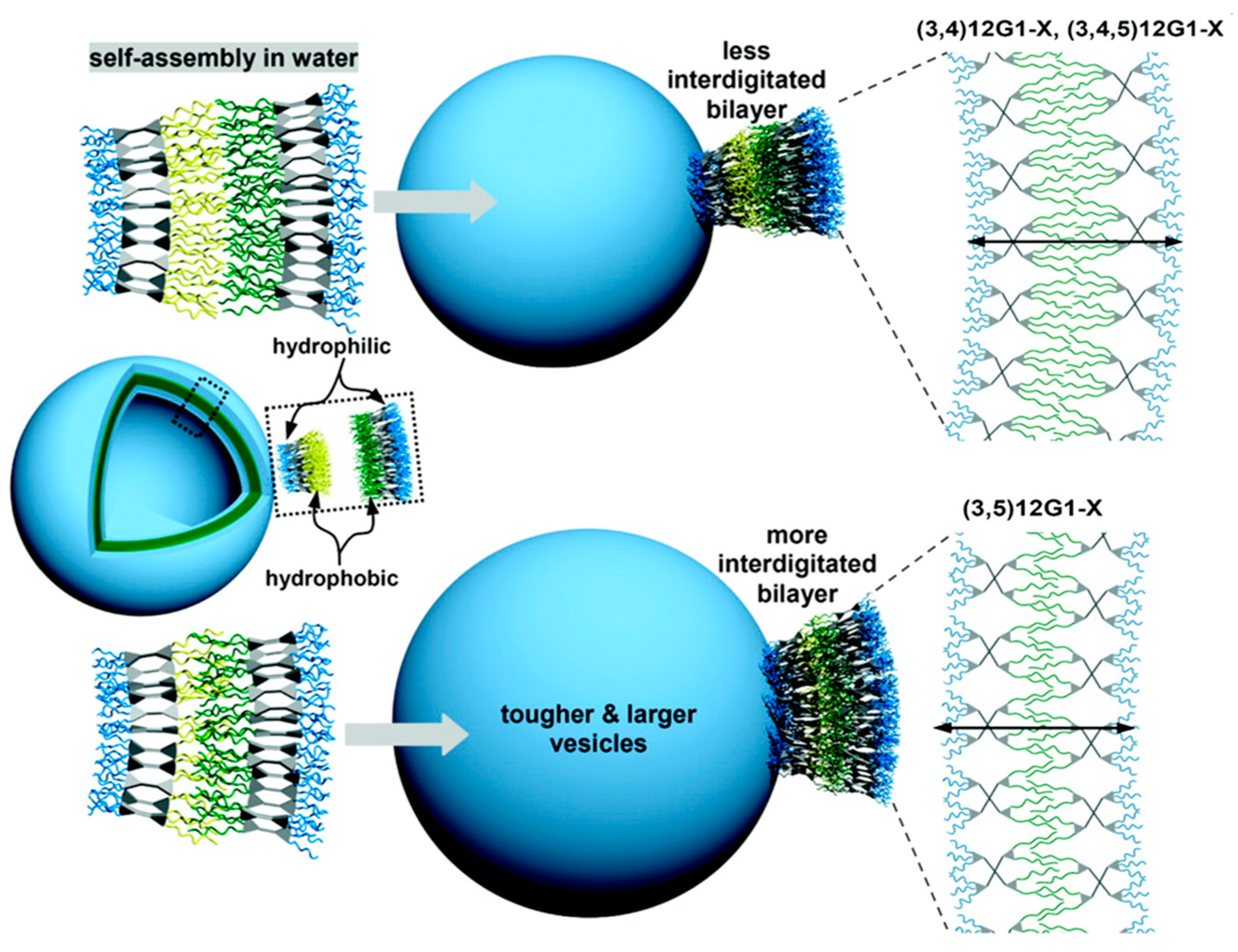
Membrane Thickness: Influence of JD Structure and Interdigitation of Its Hydrophobic Part
Curvature/Size: Influence of Hydrophilic Part, Core of the JDs and Concentration of the Solution
Membrane Fluidity: Influence of Hydrophobic Part
Lamellarity: Influence of H-Bonding Moieties
3.4.3. Biocompatibility and Stability of Vesicles
Biocompatibility
Stability Over Time and as a Function of Temperature, pH and Presence of Electrolyte
4. Biomedical Applications of Vesicles from Amphiphilic Dumbbells and Janus Dendrimers
4.1. At the Nanoscale
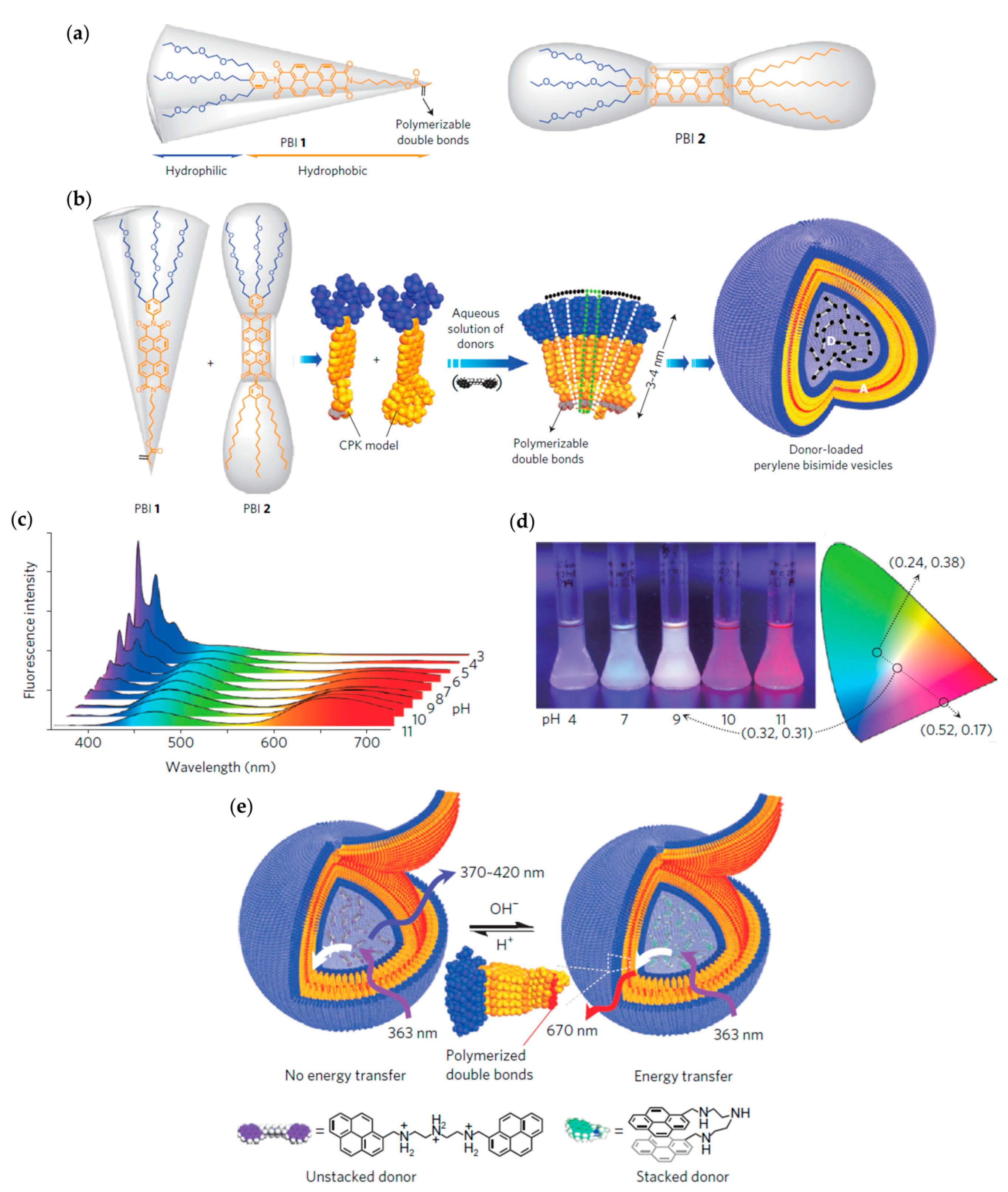
4.1.1. Vesicles from Amphiphilic Dumbbells as Fluorescent pH Sensor
4.1.2. Dendrimersomes as Nanocarriers
Drug Delivery: Model Drugs and Anticancer Drugs
Gene Delivery
4.1.3. Dendrimersomes in Diagnostic Imaging
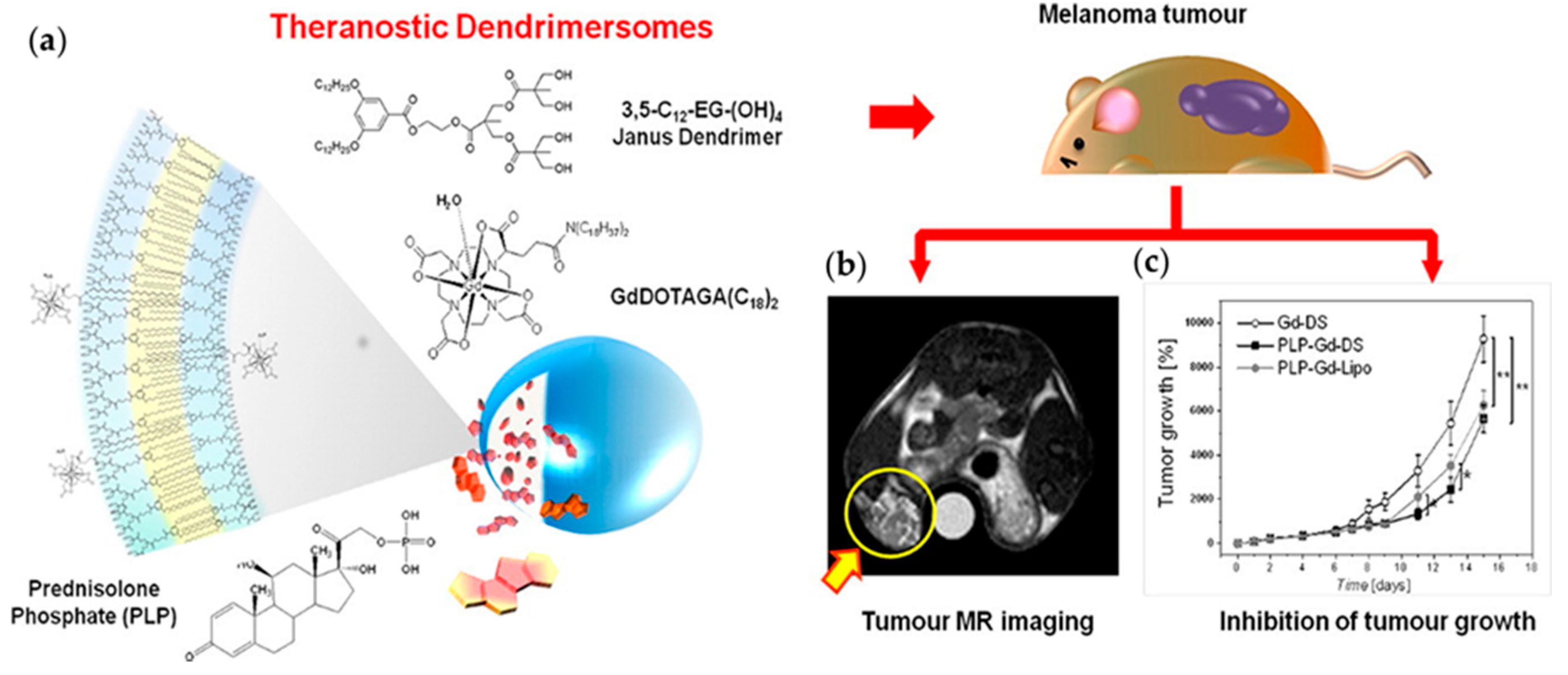
4.1.4. Dendrimersomes as Theranostics
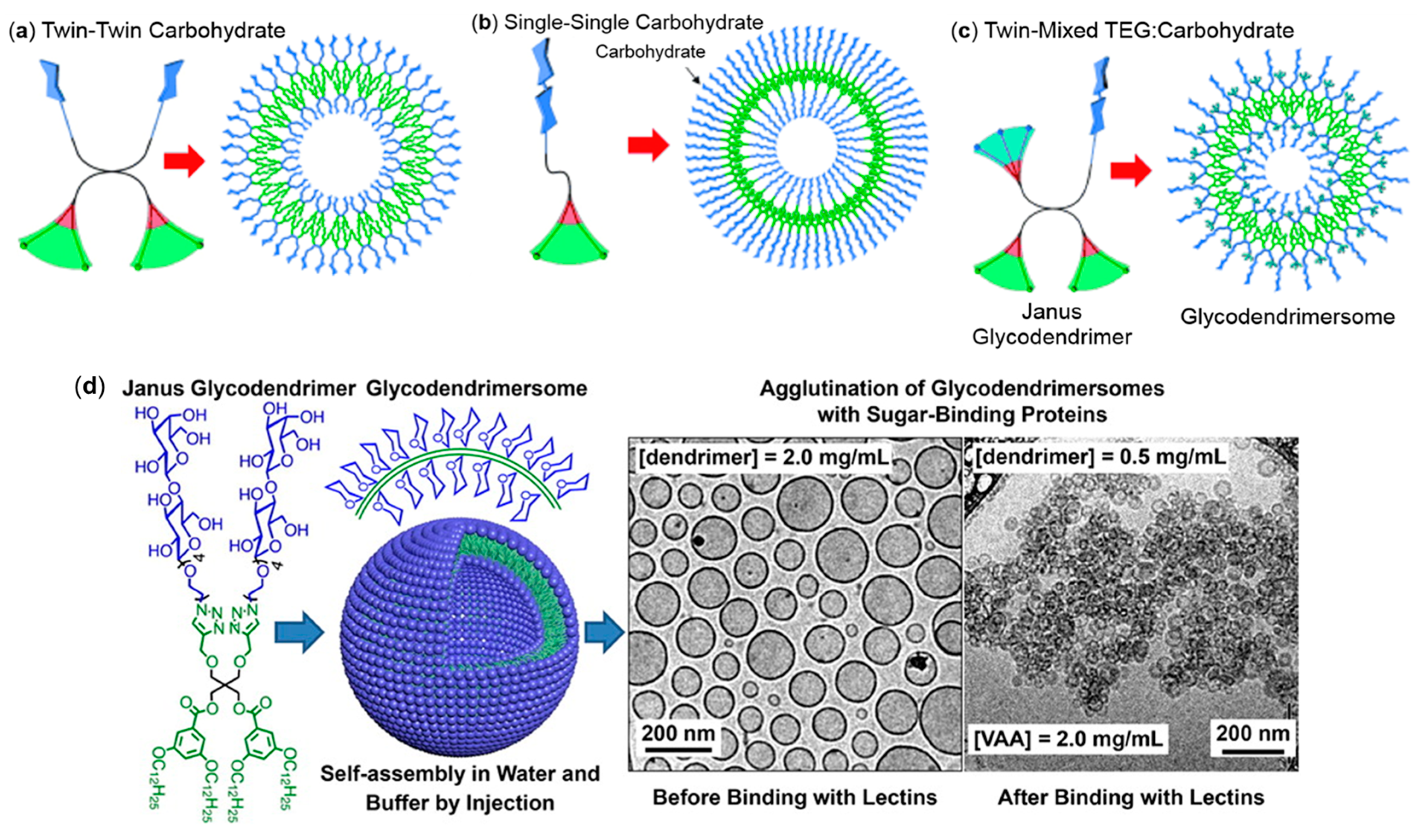
4.1.5. Dendrimersomes as Tools for Understanding Lectin Recognition on Cells
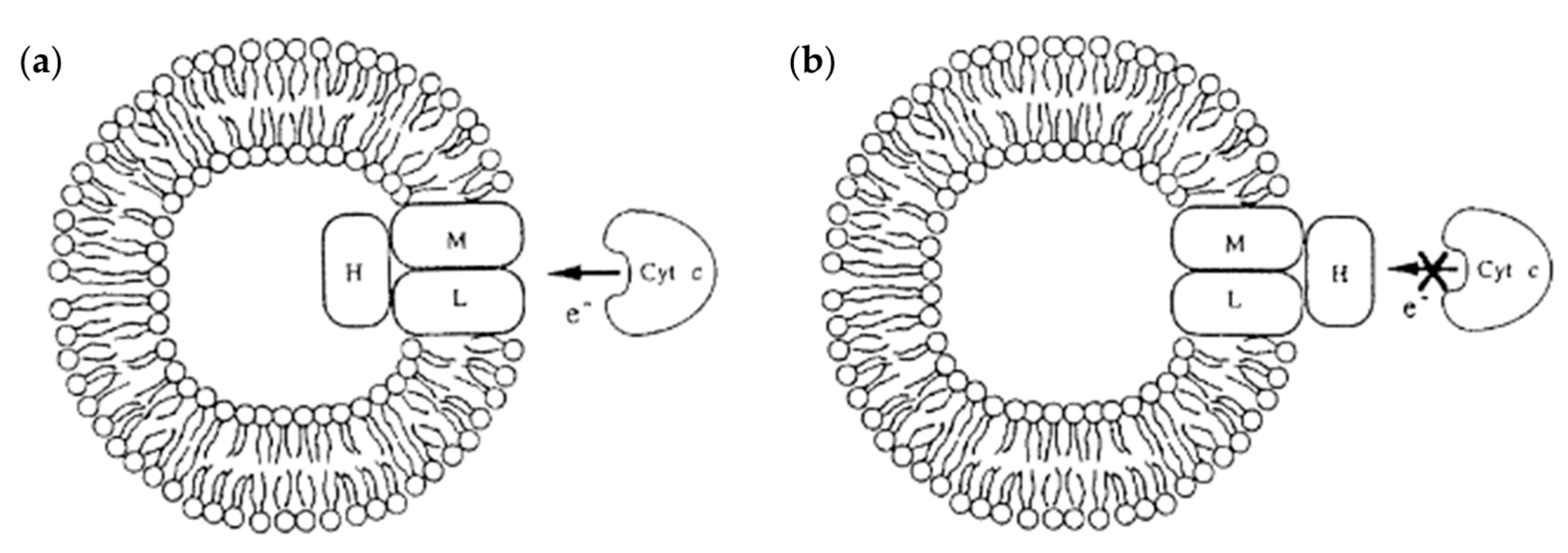
4.1.6. Dendrimersomes as Tools for Understanding the Role of Membrane Proteins
4.2. At the Microscale
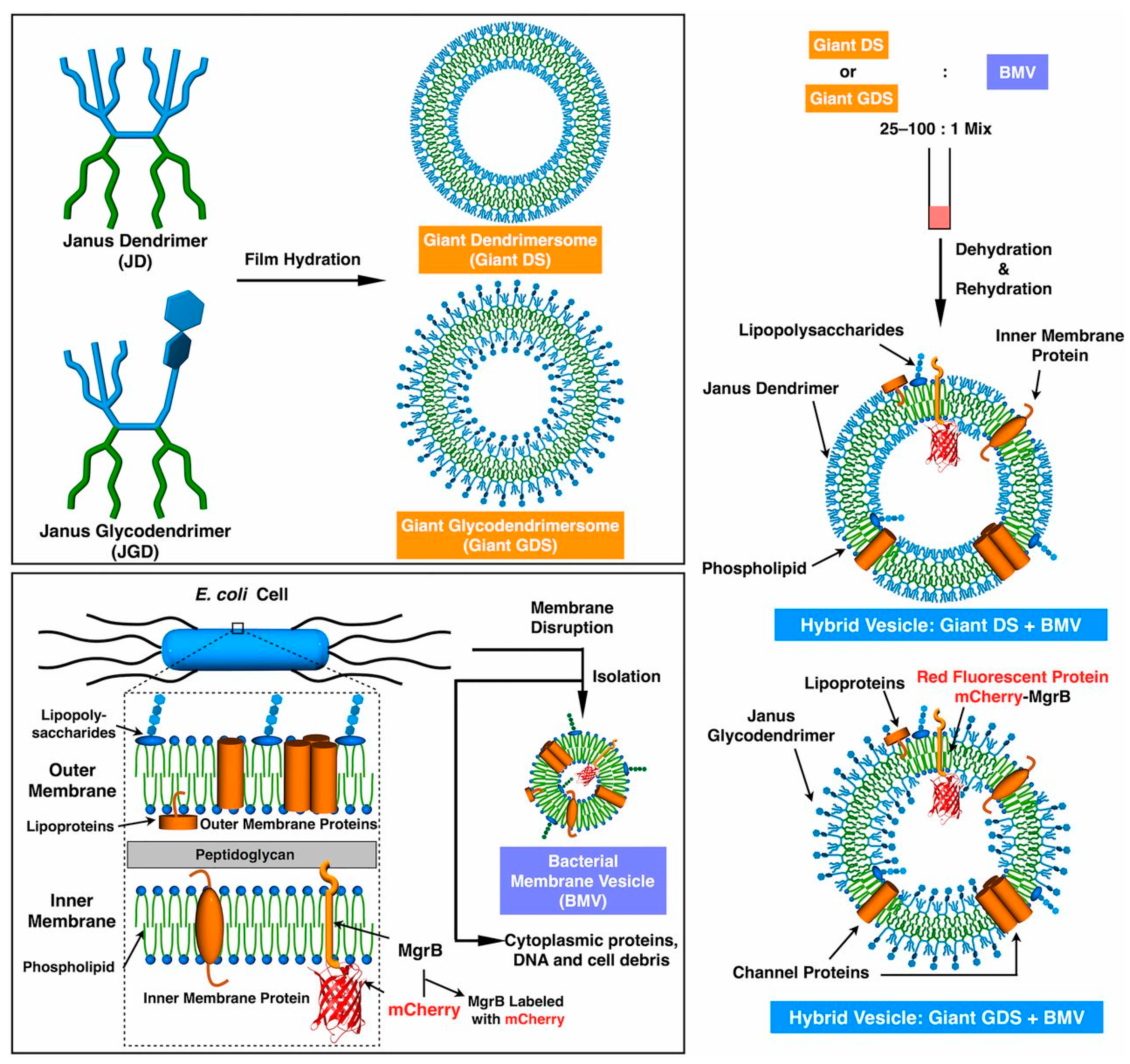
4.2.1. Dendrimersomes as Artificial Cells: Integration of Membrane Proteins
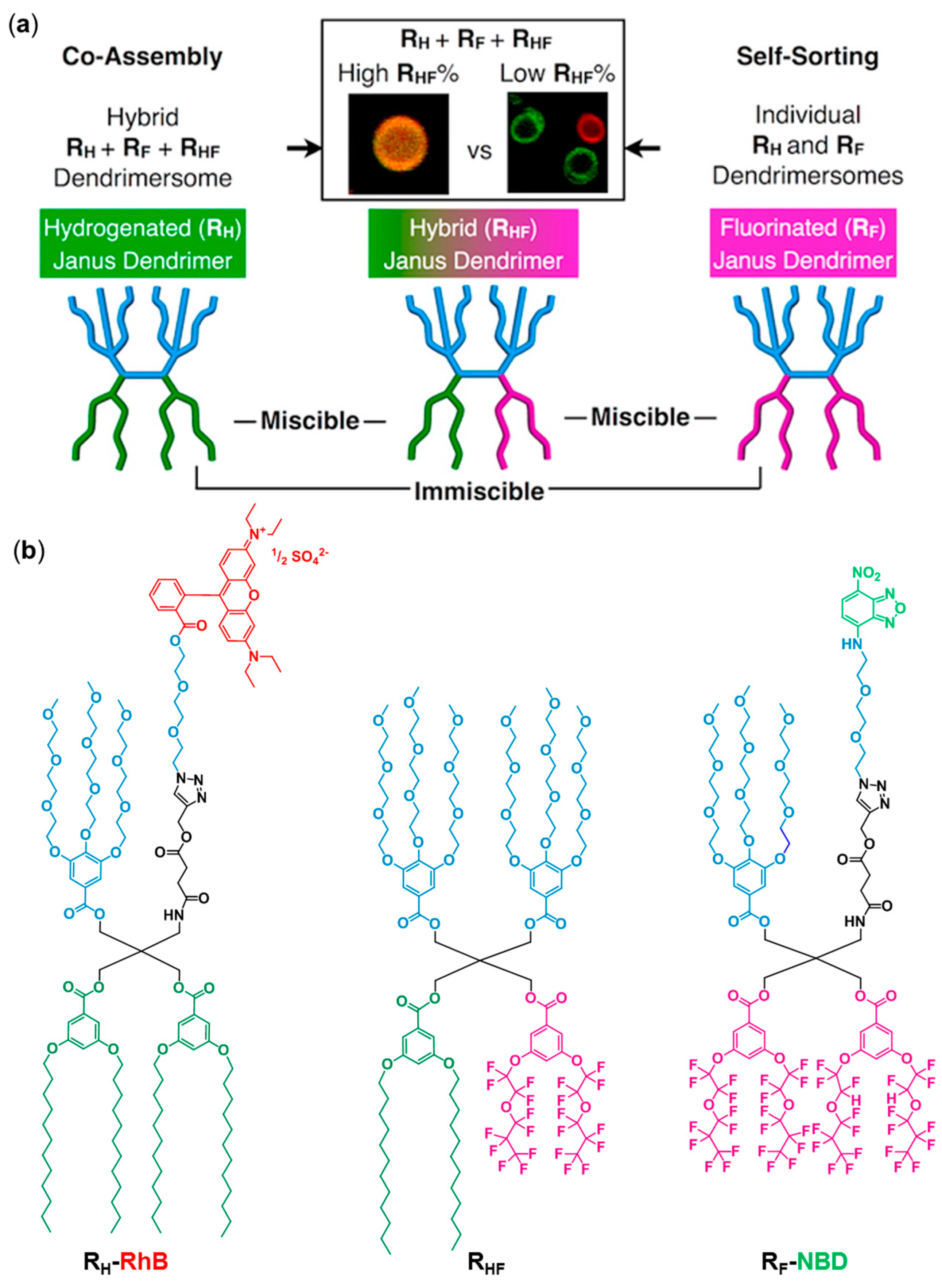
4.2.2. Self-Sorting Dendrimersomes
5. Conclusions
Acknowledgments
Conflicts of Interest
References
- Akbarzadeh, A.; Rezaei-Sadabady, R.; Davaran, S.; Joo, S.W.; Zarghami, N.; Hanifehpour, Y.; Samiei, M.; Kouhi, M.; Nejati-Koshki, K. Liposome: Classification, preparation, and applications. Nanoscale Res. Lett. 2013, 8, 102. [Google Scholar] [CrossRef] [PubMed]
- Antonietti, M.; Forster, S. Vesicles and liposomes: A self-assembly principle beyond lipids. Adv. Mater. 2003, 15, 1323–1333. [Google Scholar] [CrossRef]
- Discher, B.M.; Won, Y.-Y.; Ege, D.S.; Lee, J.C.-M.; Bates, F.S.; Discher, D.E.; Hammer, D.A. Polymersomes: Tough vesicles made from diblock copolymers. Science 1999, 284, 1143–1146. [Google Scholar] [CrossRef] [PubMed]
- Bertin, A.; Hermes, F.; Schlaad, H. Biohybrid and peptide-based polymer vesicles. Adv. Polym. Sci. 2010, 224, 167–195. [Google Scholar]
- Selin, M.; Peltonen, L.; Hirvonen, J.; Bimbo, L.M. Dendrimers and their supramolecular nanostructures for biomedical applications. J. Drug Deliv. Sci. Technol. 2016, 34, 10–20. [Google Scholar] [CrossRef]
- Chen, S.; Cheng, S.-X.; Zhuo, R.-X. Self-assembly strategy for the preparation of polymer-based nanoparticles for drug and gene delivery. Macromol. Biosci. 2011, 11, 576–589. [Google Scholar] [CrossRef] [PubMed]
- Branco, M.C.; Schneider, J.P. Self-assembling materials for therapeutic delivery. Acta Biomater. 2009, 5, 817–831. [Google Scholar] [CrossRef] [PubMed]
- Mintzer, M.A.; Simanek, E.E. Nonviral vectors for gene delivery. Chem. Rev. 2009, 109, 259–302. [Google Scholar] [CrossRef] [PubMed]
- Villaraza, A.J.L.; Bumb, A.; Brechbiel, M.W. Macromolecules, dendrimers, and nanomaterials in magnetic resonance imaging: The interplay between size, function, and pharmacokinetics. Chem. Rev. 2010, 110, 2921–2959. [Google Scholar] [CrossRef] [PubMed]
- Krasia-Christoforou, T.; Georgiou, T.K. Polymeric theranostics: Using polymer-based systems for simultaneous imaging and therapy. J. Mater. Chem. B 2013, 1, 3002–3025. [Google Scholar] [CrossRef]
- Noireaux, V.; Maeda, Y.T.; Libchaber, A. Development of an artificial cell, from self-organization to computation and self-reproduction. Proc. Natl. Acad. Sci. USA 2011, 108, 3473–3480. [Google Scholar] [CrossRef] [PubMed]
- Blain, J.C.; Szostak, J.W. Progress toward synthetic cells. Annu. Rev. Biochem. 2014, 83, 615–640. [Google Scholar] [CrossRef] [PubMed]
- Bangham, A.D.; Standish, M.M.; Watkins, J.C. Diffusion of univalent ions across the lamellae of swollen phospholipids. J. Mol. Biol. 1965, 13, 238–252. [Google Scholar] [CrossRef]
- Barenholz, Y. Liposome application: Problems and prospects. Curr. Opin. Colloid Interface Sci. 2001, 6, 66–77. [Google Scholar] [CrossRef]
- Moghimi, S.M.; Szebeni, J. Stealth liposomes and long circulating nanoparticles: Critical issues in pharmacokinetics, opsonization and protein-binding properties. Prog. Lipid Res. 2003, 42, 463–478. [Google Scholar] [CrossRef]
- Lasic, D.D.; Needham, D. The “stealth” liposome: A prototypical biomaterial. Chem. Rev. 1995, 95, 2601–2628. [Google Scholar] [CrossRef]
- LoPresti, C.; Lomas, H.; Massignani, M.; Smart, T.; Battaglia, G. Polymersomes: Nature inspired nanometer sized compartments. J. Mater. Chem. 2009, 19, 3576–3590. [Google Scholar] [CrossRef]
- Chang, H.-Y.; Sheng, Y.-J.; Tsao, H.-K. Structural and mechanical characteristics of polymersomes. Soft Matter 2014, 10, 6373–6381. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Kozlovskaya, V.; Medipelli, S.; Xue, B.; Ahmad, F.; Saeed, M.; Cropek, D.; Kharlampieva, E. Temperature-sensitive polymersomes for controlled delivery of anticancer drugs. Chem. Mater. 2015, 27, 7945–7956. [Google Scholar] [CrossRef]
- Xu, W.; Steinschulte, A.A.; Plamper, F.A.; Korolovych, V.F.; Tsukruk, V.V. Hierarchical assembly of star polymer polymersomes into responsive multicompartmental microcapsules. Chem. Mater. 2016, 28, 975–985. [Google Scholar] [CrossRef]
- Percec, V.; Wilson, D.A.; Leowanawat, P.; Wilson, C.J.; Hughes, A.D.; Kaucher, M.S.; Hammer, D.A.; Levine, D.H.; Kim, A.J.; Bates, F.S.; et al. Self-assembly of janus dendrimers into uniform dendrimersomes and other complex architectures. Science 2010, 328, 1009–1014. [Google Scholar] [CrossRef] [PubMed]
- Caminade, A.-M.; Laurent, R.; Delavaux-Nicot, B.; Majoral, J.-P. “Janus” dendrimers: Syntheses and properties. New J. Chem. 2012, 36, 217–226. [Google Scholar] [CrossRef]
- Sikwal, D.R.; Kalhapure, R.S.; Govender, T. An emerging class of amphiphilic dendrimers for pharmaceutical and biomedical applications: Janus amphiphilic dendrimers. Eur. J. Pharm. Sci. 2017, 97, 113–134. [Google Scholar] [CrossRef] [PubMed]
- Sowinska, M.; Urbanczyk-Lipkowska, Z. Advances in the chemistry of dendrimers. New J. Chem. 2014, 38, 2168–2203. [Google Scholar] [CrossRef]
- Grayson, S.M.; Fréchet, J.M.J. Convergent dendrons and dendrimers: From synthesis to applications. Chem. Rev. 2001, 101, 3819–3868. [Google Scholar] [CrossRef] [PubMed]
- Carlmark, A.; Hawker, C.; Hult, A.; Malkoch, M. New methodologies in the construction of dendritic materials. Chem. Soc. Rev. 2009, 38, 352–362. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Li, S.; Weng, Z.; Gao, C. Hyperbranched polymers: Advances from synthesis to applications. Chem. Soc. Rev. 2015, 44, 4091–4130. [Google Scholar] [CrossRef] [PubMed]
- Newkome, G.R.; Moorefield, C.N.; Baker, G.R.; Saunders, M.J.; Grossman, S.H. Unimolecular micelles. Angew. Chem. Int. Ed. 1991, 30, 1178–1180. [Google Scholar] [CrossRef]
- Newkome, G.R.; Moorefield, C.N.; Baker, G.R.; Johnson, A.L.; Behera, R.K. Alkane cascade polymers possessing micellar topology: Micellanoic acid derivatives. Angew. Chem. Int. Ed. 1991, 30, 1176–1178. [Google Scholar] [CrossRef]
- Kannaiyan, D.; Imae, T. pH-dependent encapsulation of pyrene in PPI-core: PAMAM-shell dendrimers. Langmuir 2009, 25, 5282–5285. [Google Scholar] [CrossRef] [PubMed]
- Hawker, C.J.; Wooley, K.L.; Frechet, J.M.J. Unimolecular micelles and globular amphiphiles: Dendritic macromolecules as novel recyclable solubilization agents. J. Chem. Soc. Perkin Trans. 1 1993, 1287–1297. [Google Scholar] [CrossRef]
- Liu, X.; Zhou, J.; Yu, T.; Chen, C.; Cheng, Q.; Sengupta, K.; Huang, Y.; Li, H.; Liu, C.; Wang, Y.; et al. Adaptive amphiphilic dendrimer-based nanoassemblies as robust and versatile sirna delivery systems. Angew. Chem. Int. Ed. 2014, 53, 11822–11827. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Liu, C.; Zhou, J.; Chen, C.; Qu, F.; Rossi, J.J.; Rocchi, P.; Peng, L. Promoting siRNA delivery via enhanced cellular uptake using an arginine-decorated amphiphilic dendrimer. Nanoscale 2015, 7, 3867–3875. [Google Scholar] [CrossRef] [PubMed]
- Ryu, J.-H.; Kim, H.-J.; Huang, Z.; Lee, E.; Lee, M. Self-assembling molecular dumbbells: From nanohelices to nanocapsules triggered by guest intercalation. Angew. Chem. Int. Ed. 2006, 45, 5304–5307. [Google Scholar] [CrossRef] [PubMed]
- Lee, E.; Kim, J.-K.; Lee, M. Interconversion of planar networks and vesicles triggered by temperature. Macromol. Rap. Commun. 2010, 31, 975–979. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.; Jeong, Y.-S.; Cho, B.-K.; Oh, N.-K.; Zin, W.-C. Self-assembly of molecular dumbbells into organized bundles with tunable size. Chem. A Eur. J. 2002, 8, 876–883. [Google Scholar] [CrossRef]
- Lee, E.; Jeong, Y.-H.; Kim, J.-K.; Lee, M. Controlled self-assembly of asymmetric dumbbell-shaped rod amphiphiles: Transition from toroids to planar nets. Macromolecules 2007, 40, 8355–8360. [Google Scholar] [CrossRef]
- Kim, J.-K.; Lee, E.; Huang, Z.; Lee, M. Nanorings from the self-assembly of amphiphilic molecular dumbbells. J. Am. Chem. Soc. 2006, 128, 14022–14023. [Google Scholar] [CrossRef] [PubMed]
- Bae, J.; Choi, J.-H.; Yoo, Y.-S.; Oh, N.-K.; Kim, B.-S.; Lee, M. Helical nanofibers from aqueous self-assembly of an oligo(p-phenylene)-based molecular dumbbell. J. Am. Chem. Soc. 2005, 127, 9668–9669. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.K.; Lee, E.; Lim, Y.B.; Lee, M. Supramolecular capsules with gated pores from an amphiphilic rod assembly. Angew. Chem. Int. Ed. 2008, 47, 4662–4666. [Google Scholar] [CrossRef] [PubMed]
- Trant, J.F.; Jain, N.; Mazzuca, D.M.; McIntosh, J.T.; Fan, B.; Haeryfar, S.M.M.; Lecommandoux, S.; Gillies, E.R. Synthesis, self-assembly, and immunological activity of α-galactose-functionalized dendron-lipid amphiphiles. Nanoscale 2016, 8, 17694–17704. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Ludwig, K.; Schade, B.; von Berlepsch, H.; Papp, I.; Tyagi, R.; Gulia, M.; Haag, R.; Böttcher, C. Introducing chirality into nonionic dendritic amphiphiles and studying their supramolecular assembly. Chem. A Eur. J. 2016, 22, 5629–5636. [Google Scholar] [CrossRef] [PubMed]
- De León, A.S.; Malhotra, S.; Molina, M.; Haag, R.; Calderón, M.; Rodríguez-Hernández, J.; Muñoz-Bonilla, A. Dendritic amphiphiles as additives for honeycomb-like patterned surfaces by breath figures: Role of the molecular characteristics on the pore morphology. J. Colloid Interface Sci. 2015, 440, 263–271. [Google Scholar] [CrossRef] [PubMed]
- Trappmann, B.; Ludwig, K.; Radowski, M.R.; Shukla, A.; Mohr, A.; Rehage, H.; Böttcher, C.; Haag, R. A new family of nonionic dendritic amphiphiles displaying unexpected packing parameters in micellar assemblies. J. Am. Chem. Soc. 2010, 132, 11119–11124. [Google Scholar] [CrossRef] [PubMed]
- Thota, B.N.S.; Berlepsch, H.V.; Bottcher, C.; Haag, R. Towards engineering of self-assembled nanostructures using non-ionic dendritic amphiphiles. Chem. Commun. 2015, 51, 8648–8651. [Google Scholar] [CrossRef] [PubMed]
- Yoo, Y.-S.; Choi, J.-H.; Song, J.-H.; Oh, N.-K.; Zin, W.-C.; Park, S.; Chang, T.; Lee, M. Self-assembling molecular trees containing octa-p-phenylene: From nanocrystals to nanocapsules. J. Am. Chem. Soc. 2004, 126, 6294–6300. [Google Scholar] [CrossRef] [PubMed]
- Rodrigo, A.C.; Malhotra, S.; Bottcher, C.; Adeli, M.; Haag, R. Dendritic polyglycerol cyclodextrin amphiphiles and their self-assembled architectures to transport hydrophobic guest molecules. RSC Adv. 2014, 4, 61656–61659. [Google Scholar] [CrossRef]
- Thota, B.N.S.; Urner, L.H.; Haag, R. Supramolecular architectures of dendritic amphiphiles in water. Chem. Rev. 2016, 116, 2079–2102. [Google Scholar] [CrossRef] [PubMed]
- Whitton, G.; Gillies, E.R. Functional aqueous assemblies of linear-dendron hybrids. J. Polym. Sci. Part A Polym. Chem. 2015, 53, 148–172. [Google Scholar] [CrossRef]
- Ryu, J.-H.; Hong, D.-J.; Lee, M. Aqueous self-assembly of aromatic rod building blocks. Chem. Commun. 2008, 1043–1054. [Google Scholar] [CrossRef] [PubMed]
- Gitsov, I. Hybrid linear dendritic macromolecules: From synthesis to applications. J. Polym. Sci. Part A Polym. Chem. 2008, 46, 5295–5314. [Google Scholar] [CrossRef]
- Dong, R.; Zhou, Y.; Zhu, X. Supramolecular dendritic polymers: From synthesis to applications. Acc. Chem. Res. 2014, 47, 2006–2016. [Google Scholar] [CrossRef] [PubMed]
- Park, C.; Lee, J.; Kim, C. Functional supramolecular assemblies derived from dendritic building blocks. Chem. Commun. 2011, 47, 12042–12056. [Google Scholar] [CrossRef] [PubMed]
- Al-Jamal, K.T.; Ramaswamy, C.; Florence, A.T. Supramolecular structures from dendrons and dendrimers. Adv. Drug Deliv. Rev. 2005, 57, 2238–2270. [Google Scholar] [CrossRef] [PubMed]
- Blasco, E.; Piñol, M.; Oriol, L. Responsive linear-dendritic block copolymers. Macromol. Rap. Commun. 2014, 35, 1090–1115. [Google Scholar] [CrossRef] [PubMed]
- Zeng, F.; Zimmerman, S.C. Dendrimers in supramolecular chemistry: From molecular recognition to self-assembly. Chem. Rev. 1997, 97, 1681–1712. [Google Scholar] [CrossRef] [PubMed]
- Yu, T.; Liu, X.; Bolcato-Bellemin, A.-L.; Wang, Y.; Liu, C.; Erbacher, P.; Qu, F.; Rocchi, P.; Behr, J.-P.; Peng, L. An amphiphilic dendrimer for effective delivery of small interfering RNA and gene silencing in vitro and in vivo. Angew. Chem. Int. Ed. 2012, 51, 8478–8484. [Google Scholar] [CrossRef] [PubMed]
- Fedeli, E.; Lancelot, A.; Serrano, J.L.; Calvo, P.; Sierra, T. Self-assembling amphiphilic janus dendrimers: Mesomorphic properties and aggregation in water. New J. Chem. 2015, 39, 1960–1967. [Google Scholar] [CrossRef]
- Pan, J.; Wen, M.; Yin, D.; Jiang, B.; He, D.; Guo, L. Design and synthesis of novel amphiphilic janus dendrimers for bone-targeted drug delivery. Tetrahedron 2012, 68, 2943–2949. [Google Scholar] [CrossRef]
- Pan, J.; Ma, L.; Zhao, Y.; Zhao, J.; Ouyang, L.; Guo, L. Design and synthesis of novel janus dendrimers as lipophilized antioxidants. Synlett 2013, 24, 1011–1015. [Google Scholar]
- Zhang, X.; Chen, Z.; Würthner, F. Morphology control of fluorescent nanoaggregates by co-self-assembly of wedge- and dumbbell-shaped amphiphilic perylene bisimides. J. Am. Chem. Soc. 2007, 129, 4886–4887. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Rehm, S.; Safont-Sempere, M.M.; Würthner, F. Vesicular perylene dye nanocapsules as supramolecular fluorescent ph sensor systems. Nat. Chem. 2009, 1, 623–629. [Google Scholar] [CrossRef] [PubMed]
- Nazemi, A.; Gillies, E.R. Dendrimersomes with photodegradable membranes for triggered release of hydrophilic and hydrophobic cargo. Chem. Commun. 2014, 50, 11122–11125. [Google Scholar] [CrossRef] [PubMed]
- Filippi, M.; Patrucco, D.; Martinelli, J.; Botta, M.; Castro-Hartmann, P.; Tei, L.; Terreno, E. Novel stable dendrimersome formulation for safe bioimaging applications. Nanoscale 2015, 7, 12943–12954. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Sun, H.-J.; Hughes, A.D.; Moussodia, R.-O.; Bertin, A.; Chen, Y.; Pochan, D.J.; Heiney, P.A.; Klein, M.L.; Percec, V. Self-assembly of amphiphilic janus dendrimers into uniform onion-like dendrimersomes with predictable size and number of bilayers. Proc. Natl. Acad. Sci. USA 2014, 111, 9058–9063. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Sun, H.-J.; Hughes, A.D.; Draghici, B.; Lejnieks, J.; Leowanawat, P.; Bertin, A.; Otero De Leon, L.; Kulikov, O.V.; Chen, Y.; et al. “Single-single” amphiphilic janus dendrimers self-assemble into uniform dendrimersomes with predictable size. ACS Nano 2014, 8, 1554–1565. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Q.; Rubien, J.D.; Wang, Z.; Reed, E.H.; Hammer, D.A.; Sahoo, D.; Heiney, P.A.; Yadavalli, S.S.; Goulian, M.; Wilner, S.E.; et al. Self-sorting and coassembly of fluorinated, hydrogenated, and hybrid janus dendrimers into dendrimersomes. J. Am. Chem. Soc. 2016, 138, 12655–12663. [Google Scholar] [CrossRef] [PubMed]
- Jang, J.; Kim, J.-K.; Choi, J.-W.; Hwang, T.-S.; Jo, M.; Kim, I.; Cho, B.-K.; Lee, E. Organic-inorganic vesicular hybrids driven by assembly of dendritic amphiphiles: Site-selective encapsulation of nanoparticles. Chem. Commun. 2013, 49, 8003–8005. [Google Scholar] [CrossRef] [PubMed]
- Prabakaran, P.; Prasad, E. Janus dendrimer from poly(aryl ether) linked PAMAM for supergelation and guest release. Chem. Sel. 2016, 1, 5561–5568. [Google Scholar] [CrossRef]
- Giustini, M.; Bellinazzo, C.; Galantini, L.; Mallardi, A.; Palazzo, G.; Sennato, S.; Bordi, F.; Rissanen, K. Incorporation of the bacterial reaction centre into dendrimersomes. Colloids Surf. A Physicochem. Eng. Asp. 2012, 413, 38–43. [Google Scholar] [CrossRef]
- Filippi, M.; Martinelli, J.; Mulas, G.; Ferraretto, M.; Teirlinck, E.; Botta, M.; Tei, L.; Terreno, E. Dendrimersomes: A new vesicular nano-platform for MR-molecular imaging applications. Chem. Commun. 2014, 50, 3453–3456. [Google Scholar] [CrossRef] [PubMed]
- Percec, V.; Leowanawat, P.; Sun, H.-J.; Kulikov, O.; Nusbaum, C.D.; Tran, T.M.; Bertin, A.; Wilson, D.A.; Peterca, M.; Zhang, S.; et al. Modular synthesis of amphiphilic janus glycodendrimers and their self-assembly into glycodendrimersomes and other complex architectures with bioactivity to biomedically relevant lectins. J. Am. Chem. Soc. 2013, 135, 9055–9077. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Xiao, Q.; Sherman, S.E.; Muncan, A.; Ramos Vicente, A.D.M.; Wang, Z.; Hammer, D.A.; Williams, D.; Chen, Y.; Pochan, D.J.; et al. Glycodendrimersomes from sequence-defined janus glycodendrimers reveal high activity and sensor capacity for the agglutination by natural variants of human lectins. J. Am. Chem. Soc. 2015, 137, 13334–13344. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Q.; Zhang, S.; Wang, Z.; Sherman, S.E.; Moussodia, R.-O.; Peterca, M.; Muncan, A.; Williams, D.R.; Hammer, D.A.; Vértesy, S.; et al. Onion-like glycodendrimersomes from sequence-defined janus glycodendrimers and influence of architecture on reactivity to a lectin. Proc. Natl. Acad. Sci. USA 2016, 113, 1162–1167. [Google Scholar] [CrossRef] [PubMed]
- Yuan, F.; Zhang, X.; Yang, M.; Wang, W.; Minch, B.; Lieser, G.; Wegner, G. Topological transformation of aggregates formed by an amphiphilic and truncated-cone-shaped codendrimer. Soft Matter 2007, 3, 1372–1376. [Google Scholar] [CrossRef]
- Xiao, Q.; Yadavalli, S.S.; Zhang, S.; Sherman, S.E.; Fiorin, E.; da Silva, L.; Wilson, D.A.; Hammer, D.A.; André, S.; Gabius, H.-J.; et al. Bioactive cell-like hybrids coassembled from (glyco)dendrimersomes with bacterial membranes. Proc. Natl. Acad. Sci. USA 2016, 113, E1134–E1141. [Google Scholar] [CrossRef] [PubMed]
- Peterca, M.; Percec, V.; Leowanawat, P.; Bertin, A. Predicting the size and properties of dendrimersomes from the lamellar structure of their amphiphilic janus dendrimers. J. Am. Chem. Soc. 2011, 133, 20507–20520. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Moussodia, R.-O.; Sun, H.-J.; Leowanawat, P.; Muncan, A.; Nusbaum, C.D.; Chelling, K.M.; Heiney, P.A.; Klein, M.L.; André, S.; et al. Mimicking biological membranes with programmable glycan ligands self-assembled from amphiphilic janus glycodendrimers. Angew. Chem. Int. Ed. 2014, 53, 10899–10903. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Moussodia, R.-O.; Vértesy, S.; André, S.; Klein, M.L.; Gabius, H.-J.; Percec, V. Unraveling functional significance of natural variations of a human galectin by glycodendrimersomes with programmable glycan surface. Proc. Natl. Acad. Sci. USA 2015, 112, 5585–5590. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Moussodia, R.-O.; Murzeau, C.; Sun, H.-J.; Klein, M.L.; Vértesy, S.; André, S.; Roy, R.; Gabius, H.-J.; Percec, V. Dissecting molecular aspects of cell interactions using glycodendrimersomes with programmable glycan presentation and engineered human lectins. Angew. Chem. Int. Ed. 2015, 54, 4036–4040. [Google Scholar] [CrossRef] [PubMed]
- Filippi, M.; Catanzaro, V.; Patrucco, D.; Botta, M.; Tei, L.; Terreno, E. First in vivo MRI study on theranostic dendrimersomes. J. Control. Release 2017, 248, 45–52. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Kim, Y.; Lee, M. Intelligent supramolecular assembly of aromatic block molecules in aqueous solution. Nanoscale 2013, 5, 7711–7723. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.-J.; Kim, T.; Lee, M. Responsive nanostructures from aqueous assembly of rigid-flexible block molecules. Acc. Chem. Res. 2011, 44, 72–82. [Google Scholar] [CrossRef] [PubMed]
- Stupp, S.I. Self-assembly of rodcoil molecules. Curr. Opin. Colloid Interface Sci. 1998, 3, 20–26. [Google Scholar] [CrossRef]
- Jang, C.-J.; Ryu, J.-H.; Lee, J.-D.; Sohn, D.; Lee, M. Synthesis and supramolecular nanostructure of amphiphilic rigid aromatic-flexible dendritic block molecules. Chem. Mater. 2004, 16, 4226–4231. [Google Scholar] [CrossRef]
- Maeda, H.; Wu, J.; Sawa, T.; Matsumura, Y.; Hori, K. Tumor vascular permeability and the EPR effect in macromolecular therapeutics: A review. J. Control. Release 2000, 65, 271–284. [Google Scholar] [CrossRef]
- Rao, D.A.; Robinson, J.R. Effect of size and surface properties of biodegradable PLGA-PMA: PLA:PEG nanoparticles on lymphatic uptake and retention in rats. J. Control. Release 2008, 132, e45–e47. [Google Scholar] [CrossRef]
- Sarin, H. Physiologic upper limits of pore size of different blood capillary types and another perspective on the dual pore theory of microvascular permeability. J. Angiogenesis Res. 2010, 2, 14. [Google Scholar] [CrossRef] [PubMed]
- Cabral, H.; Matsumoto, Y.; Mizuno, K.; Chen, Q.; Murakami, M.; Kimura, M.; Terada, Y.; Kano, M.R.; Miyazono, K.; Uesaka, M.; et al. Accumulation of sub-100 nm polymeric micelles in poorly permeable tumours depends on size. Nat. Nanotechnol. 2011, 6, 815–823. [Google Scholar] [CrossRef] [PubMed]
- Longmire, M.; Choyke, P.L.; Kobayashi, H. Clearance properties of nano-sized particles and molecules as imaging agents: Considerations and caveats. Nanomedicine 2008, 3, 703–717. [Google Scholar] [CrossRef] [PubMed]
- Litzinger, D.C.; Buiting, A.M.J.; van Rooijen, N.; Huang, L. Effect of liposome size on the circulation time and intraorgan distribution of amphipathic poly(ethylene glycol)-containing liposomes. Biochim. Biophys. Acta 1994, 1190, 99–107. [Google Scholar] [CrossRef]
- Nagayasu, A.; Uchiyama, K.; Kiwada, H. The size of liposomes: A factor which affects their targeting efficiency to tumors and therapeutic activity of liposomal antitumor drugs. Adv. Drug Deliv. Rev. 1999, 40, 75–87. [Google Scholar] [CrossRef]
- Nummelin, S.; Liljeström, V.; Saarikoski, E.; Ropponen, J.; Nykänen, A.; Linko, V.; Seppälä, J.; Hirvonen, J.; Ikkala, O.; Bimbo, L.M.; et al. Self-assembly of amphiphilic janus dendrimers into mechanically robust supramolecular hydrogels for sustained drug release. Chem. A Eur. J. 2015, 21, 14433–14439. [Google Scholar] [CrossRef] [PubMed]
- Israelachvili, J.N.; Mitchell, D.J.; Ninham, B.W. Theory of self-assembly of hydrocarbon amphiphiles into micelles and bilayers. J. Chem. Soc. Faraday Trans. 2 1976, 72, 1525–1568. [Google Scholar] [CrossRef]
- Shinoda, W.; DeVane, R.; Klein, M.L. Computer simulation studies of self-assembling macromolecules. Curr. Opin. Struct. Biol. 2012, 22, 175–186. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Sharma, U.S. Liposomes in drug delivery: Progress and limitations. Int. J. Pharm. 1997, 154, 123–140. [Google Scholar] [CrossRef]
- Laouini, A.; Jaafar-Maalej, C.; Limayem-Blouza, I.; Sfar, S.; Charcosset, C.; Fessi, H. Preparation, characterization and applications of liposomes: State of the art. J. Colloid Sci. Biotechnol. 2012, 1, 147–168. [Google Scholar] [CrossRef]
- Patil, Y.P.; Jadhav, S. Novel methods for liposome preparation. Chem. Phys. Lipids 2014, 177, 8–18. [Google Scholar] [CrossRef] [PubMed]
- Reeves, J.P.; Dowben, R.M. Formation and properties of thin-walled phospholipid vesicles. J. Cell. Physiol. 1969, 73, 49–60. [Google Scholar] [CrossRef] [PubMed]
- Walde, P.; Cosentino, K.; Engel, H.; Stano, P. Giant vesicles: Preparations and applications. ChemBioChem 2010, 11, 848–865. [Google Scholar] [CrossRef] [PubMed]
- Hishida, M.; Seto, H.; Yamada, N.L.; Yoshikawa, K. Hydration process of multi-stacked phospholipid bilayers to form giant vesicles. Chem. Phys. Lett. 2008, 455, 297–302. [Google Scholar] [CrossRef]
- Hishida, M.; Seto, H.; Yoshikawa, K. Smooth/rough layering in liquid-crystalline/gel state of dry phospholipid film, in relation to its ability to generate giant vesicles. Chem. Phys. Lett. 2005, 411, 267–272. [Google Scholar] [CrossRef]
- Rodriguez, N.; Pincet, F.; Cribier, S. Giant vesicles formed by gentle hydration and electroformation: A comparison by fluorescence microscopy. Colloids Surf. B Biointerfaces 2005, 42, 125–130. [Google Scholar] [CrossRef] [PubMed]
- Angelova, M.; Dimitrov, D.S. A mechanism of liposome electroformation. In Trends in Colloid and Interface Science II; Degiorgio, V., Ed.; Steinkopff: Darmstadt, Germany, 1988; pp. 59–67. [Google Scholar]
- Angelova, M.I.; Dimitrov, D.S. Liposome electroformation. Faraday Discuss. Chem. Soc. 1986, 81, 303–311. [Google Scholar] [CrossRef]
- Angelova, M.I.; Soléau, S.; Méléard, P.; Faucon, F.; Bothorel, P. Preparation of giant vesicles by external ac electric fields. Kinetics and applications. In Trends in Colloid and Interface Science VI; Helm, C., Lösche, M., Möhwald, H., Eds.; Steinkopff: Darmstadt, Germany, 1992; pp. 127–131. [Google Scholar]
- Shimanouchi, T.; Umakoshi, H.; Kuboi, R. Kinetic study on giant vesicle formation with electroformation method. Langmuir 2009, 25, 4835–4840. [Google Scholar] [CrossRef] [PubMed]
- Silva, R.; Ferreira, H.; Little, C.; Cavaco-Paulo, A. Effect of ultrasound parameters for unilamellar liposome preparation. Ultrason. Sonochem. 2010, 17, 628–632. [Google Scholar] [CrossRef] [PubMed]
- Bruinsma, R. Rheology and shape transitions of vesicles under capillary flow. Physica A 1996, 234, 249–270. [Google Scholar] [CrossRef]
- Frisken, B.J.; Asman, C.; Patty, P.J. Studies of vesicle extrusion. Langmuir 2000, 16, 928–933. [Google Scholar] [CrossRef]
- Hunter, D.G.; Frisken, B.J. Effect of extrusion pressure and lipid properties on the size and polydispersity of lipid vesicles. Biophys. J. 1998, 74, 2996–3002. [Google Scholar] [CrossRef]
- Patty, P.J.; Frisken, B.J. The pressure-dependence of the size of extruded vesicles. Biophys. J. 2003, 85, 996–1004. [Google Scholar] [CrossRef]
- Mayer, L.D.; Hope, M.J.; Cullis, P.R. Vesicles of variable sizes produced by a rapid extrusion procedure. Biochim. Biophys. Acta 1986, 858, 161–168. [Google Scholar] [CrossRef]
- Clerc, S.G.; Thompson, T.E. A possible mechanism for vesicle formation by extrusion. Biophys. J. 1994, 67, 475–476. [Google Scholar] [CrossRef]
- Patil, Y.P.; Kumbhalkar, M.D.; Jadhav, S. Extrusion of electroformed giant unilamellar vesicles through track-etched membranes. Chem. Phys. Lipids 2012, 165, 475–481. [Google Scholar] [CrossRef] [PubMed]
- Castile, J.D.; Taylor, K.M.G. Factors affecting the size distribution of liposomes produced by freeze–thaw extrusion. Int. J. Pharm. 1999, 188, 87–95. [Google Scholar] [CrossRef]
- Oku, N.; MacDonald, R.C. Differential effects of alkali metal chlorides on formation of giant liposomes by freezing and thawing and by dialysis. Biochemistry 1983, 22, 855–863. [Google Scholar] [CrossRef] [PubMed]
- Lasic, D.D. The mechanism of vesicle formation. Biochem. J. 1988, 256, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Lasic, D.D. Mechanisms of liposome formation. J. Liposome Res. 1995, 5, 431–441. [Google Scholar] [CrossRef]
- Marsh, D. Thermodynamics of phospholipid self-assembly. Biophys. J. 2012, 102, 1079–1087. [Google Scholar] [CrossRef] [PubMed]
- Cortesi, R.; Esposito, E.; Gambarin, S.; Telloli, P.; Menegatti, E.; Nastruzzi, C. Preparation of liposomes by reverse-phase evaporation using alternative organic solvents. J. Microencapsul. 1999, 16, 251–256. [Google Scholar] [CrossRef] [PubMed]
- Vemuri, S.; Rhodes, C.T. Preparation and characterization of liposomes as therapeutic delivery systems: A review. Pharm. Acta Helv. 1995, 70, 95–111. [Google Scholar] [CrossRef]
- Parker, W.A. Alcohol-containing pharmaceuticals. Am. J. Drug Alcohol Abus. 1982, 9, 195–209. [Google Scholar] [CrossRef]
- Wesselman, H.J. Quantitative determination of ethanol in pharmaceutical products by gas chromatography. J. Am. Pharm. Assoc. 1960, 49, 320–322. [Google Scholar] [CrossRef]
- Bleul, R.; Thiermann, R.; Maskos, M. Techniques to control polymersome size. Macromolecules 2015, 48, 7396–7409. [Google Scholar] [CrossRef]
- Sanson, C.; Schatz, C.; Le Meins, J.-F.; Brûlet, A.; Soum, A.; Lecommandoux, S. Biocompatible and biodegradable poly(trimethylene carbonate)-b-poly(l-glutamic acid) polymersomes: Size control and stability. Langmuir 2010, 26, 2751–2760. [Google Scholar] [CrossRef] [PubMed]
- Zhigaltsev, I.V.; Belliveau, N.; Hafez, I.; Leung, A.K.K.; Huft, J.; Hansen, C.; Cullis, P.R. Bottom-up design and synthesis of limit size lipid nanoparticle systems with aqueous and triglyceride cores using millisecond microfluidic mixing. Langmuir 2012, 28, 3633–3640. [Google Scholar] [CrossRef] [PubMed]
- Kastner, E.; Kaur, R.; Lowry, D.; Moghaddam, B.; Wilkinson, A.; Perrie, Y. High-throughput manufacturing of size-tuned liposomes by a new microfluidics method using enhanced statistical tools for characterization. Int. J. Pharm. 2014, 477, 361–368. [Google Scholar] [CrossRef] [PubMed]
- Thiermann, R.; Mueller, W.; Montesinos-Castellanos, A.; Metzke, D.; Löb, P.; Hessel, V.; Maskos, M. Size controlled polymersomes by continuous self-assembly in micromixers. Polymer 2012, 53, 2205–2210. [Google Scholar] [CrossRef]
- Bleul, R.; Thiermann, R.; Marten, G.U.; House, M.J.; Pierre, T.G.S.; Hafeli, U.O.; Maskos, M. Continuously manufactured magnetic polymersomes—A versatile tool (not only) for targeted cancer therapy. Nanoscale 2013, 5, 11385–11393. [Google Scholar] [CrossRef] [PubMed]
- Messager, L.; Gaitzsch, J.; Chierico, L.; Battaglia, G. Novel aspects of encapsulation and delivery using polymersomes. Curr. Opin. Pharm. 2014, 18, 104–111. [Google Scholar] [CrossRef] [PubMed]
- Won, Y.-Y.; Davis, H.T.; Bates, F.S. Molecular exchange in PEO−PB micelles in water. Macromolecules 2003, 36, 953–955. [Google Scholar] [CrossRef]
- Narrainen, A.P.; Pascual, S.; Haddleton, D.M. Amphiphilic diblock, triblock, and star block copolymers by living radical polymerization: Synthesis and aggregation behavior. J. Polym. Sci. Part A Polym. Chem. 2002, 40, 439–450. [Google Scholar] [CrossRef]
- Anraku, Y.; Kishimura, A.; Oba, M.; Yamasaki, Y.; Kataoka, K. Spontaneous formation of nanosized unilamellar polyion complex vesicles with tunable size and properties. J. Am. Chem. Soc. 2010, 132, 1631–1636. [Google Scholar] [CrossRef] [PubMed]
- Kishimura, A. Development of polyion complex vesicles (picsomes) from block copolymers for biomedical applications. Polym. J. 2013, 45, 892–897. [Google Scholar] [CrossRef]
- Liu, X.; Wu, J.; Kim, J.-S.; Eisenberg, A. Self-assembly of mixtures of block copolymers of poly(styrene-b-acrylic acid) with random copolymers of poly(styrene-co-methacrylic acid). Langmuir 2006, 22, 419–424. [Google Scholar] [CrossRef] [PubMed]
- Claessens, M.M.A.E.; Leermakers, F.A.M.; Hoekstra, F.A.; Cohen Stuart, M.A. Entropic stabilization and equilibrium size of lipid vesicles. Langmuir 2007, 23, 6315–6320. [Google Scholar] [CrossRef] [PubMed]
- Arai, N.; Yasuoka, K.; Zeng, X.C. Self-assembly of janus oligomers into onion-like vesicles with layer-by-layer water discharging capability: A minimalist model. ACS Nano 2016, 10, 8026–8037. [Google Scholar] [CrossRef] [PubMed]
- Wurthner, F. Perylene bisimide dyes as versatile building blocks for functional supramolecular architectures. Chem. Commun. 2004, 14, 1564–1579. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Dierschke, F.; Wu, J.; Grimsdale, A.C.; Mullen, K. Poly(2,7-carbazole) and perylene tetracarboxydiimide: A promising donor/acceptor pair for polymer solar cells. J. Mater. Chem. 2006, 16, 96–100. [Google Scholar] [CrossRef]
- Schmidt-Mende, L.; Fechtenkotter, A.; Mullen, K.; Moons, E.; Friend, R.H.; MacKenzie, J.D. Self-organized discotic liquid crystals for high-efficiency organic photovoltaics. Science 2001, 293, 1119–1122. [Google Scholar] [CrossRef] [PubMed]
- He, X.; Liu, H.; Li, Y.; Wang, S.; Li, Y.; Wang, N.; Xiao, J.; Xu, X.; Zhu, D. Gold nanoparticle-based fluorometric and colorimetric sensing of copper(II) ions. Adv. Mater. 2005, 17, 2811–2815. [Google Scholar] [CrossRef]
- Wang, B.; Yu, C. Fluorescence turn-on detection of a protein through the reduced aggregation of a perylene probe. Angew. Chem. Int. Ed. 2010, 49, 1485–1488. [Google Scholar] [CrossRef] [PubMed]
- Che, Y.; Yang, X.; Liu, G.; Yu, C.; Ji, H.; Zuo, J.; Zhao, J.; Zang, L. Ultrathin n-type organic nanoribbons with high photoconductivity and application in optoelectronic vapor sensing of explosives. J. Am. Chem. Soc. 2010, 132, 5743–5750. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Fu, L.; Zou, W.; Zhang, F.; Wei, Z. Ammonia sensory properties based on single-crystalline micro/nanostructures of perylenediimide derivatives: Core-substituted effect. J. Phys. Chem. C 2011, 115, 10399–10404. [Google Scholar] [CrossRef]
- Würthner, F.; Saha-Möller, C.R.; Fimmel, B.; Ogi, S.; Leowanawat, P.; Schmidt, D. Perylene bisimide dye assemblies as archetype functional supramolecular materials. Chem. Rev. 2016, 116, 962–1052. [Google Scholar] [CrossRef] [PubMed]
- Fedele, S. Diagnostic aids in the screening of oral cancer. Head Neck Oncol. 2009, 1, 5. [Google Scholar] [CrossRef] [PubMed]
- Vauthier, C.; Dubernet, C.; Chauvierre, C.; Brigger, I.; Couvreur, P. Drug delivery to resistant tumors: The potential of poly(alkyl cyanoacrylate) nanoparticles. J. Control. Release 2003, 93, 151–160. [Google Scholar] [CrossRef] [PubMed]
- Liang, X.-J.; Chen, C.; Zhao, Y.; Wang, P.C. Circumventing tumor resistance to chemotherapy by nanotechnology. Methods Mol. Biol. 2010, 596, 467–488. [Google Scholar] [PubMed]
- Bock, C.; Lengauer, T. Managing drug resistance in cancer: Lessons from HIV therapy. Nat. Rev. Cancer 2012, 12, 494–501. [Google Scholar] [CrossRef] [PubMed]
- Yin, Q.; Shen, J.; Zhang, Z.; Yu, H.; Li, Y. Reversal of multidrug resistance by stimuli-responsive drug delivery systems for therapy of tumor. Adv. Drug Deliv. Rev. 2013, 65, 1699–1715. [Google Scholar] [CrossRef] [PubMed]
- Savjani, K.T.; Gajjar, A.K.; Savjani, J.K. Drug solubility: Importance and enhancement techniques. Int. Sch. Res. Not. Pharm. 2012, 2012, 195727. [Google Scholar] [CrossRef] [PubMed]
- Allen, T.M.; Cullis, P.R. Drug delivery systems: Entering the mainstream. Science 2004, 303, 1818–1822. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Wang, X.; Zhang, T.; Wang, C.; Huang, Z.; Luo, X.; Deng, Y. A review on phospholipids and their main applications in drug delivery systems. Asian J. Pharm. Sci. 2015, 10, 81–98. [Google Scholar] [CrossRef]
- Ta, T.; Convertine, A.J.; Reyes, C.R.; Stayton, P.S.; Porter, T.M. Thermosensitive liposomes modified with poly(N-isopropylacrylamide-co-propylacrylic acid) copolymers for triggered release of doxorubicin. Biomacromolecules 2010, 11, 1915–1920. [Google Scholar] [CrossRef] [PubMed]
- Kneidl, B.; Peller, M.; Winter, G.; Lindner, L.H.; Hossann, M. Thermosensitive liposomal drug delivery systems: State of the art review. Int. J. Nanomed. 2014, 9, 4387–4398. [Google Scholar]
- Chandra, B.; Subramaniam, R.; Mallik, S.; Srivastava, D.K. Formulation of photocleavable liposomes and the mechanism of their content release. Org. Biomol. Chem. 2006, 4, 1730–1740. [Google Scholar] [CrossRef] [PubMed]
- Bayer, A.M.; Alam, S.; Mattern-Schain, S.I.; Best, M.D. Triggered liposomal release through a synthetic phosphatidylcholine analogue bearing a photocleavable moiety embedded within the sn-2 acyl chain. Chem. A Eur. J. 2014, 20, 3350–3357. [Google Scholar] [CrossRef] [PubMed]
- Leung, S.J.; Romanowski, M. Light-activated content release from liposomes. Theranostics 2012, 2, 1020–1036. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, D.D.S.; Lopes, S.C.D.A.; Franco, M.S.; Oliveira, M.C. pH-sensitive liposomes for drug delivery in cancer treatment. Ther. Deliv. 2013, 4, 1099–1123. [Google Scholar] [CrossRef] [PubMed]
- Kato, Y.; Ozawa, S.; Miyamoto, C.; Maehata, Y.; Suzuki, A.; Maeda, T.; Baba, Y. Acidic extracellular microenvironment and cancer. Cancer Cell Int. 2013, 13, 89. [Google Scholar] [CrossRef] [PubMed]
- Park, M.-K.; Jun, S.; Kim, I.; Jin, S.-M.; Kim, J.-G.; Shin, T.J.; Lee, E. Stepwise drug-release behavior of onion-like vesicles generated from emulsification-induced assembly of semicrystalline polymer amphiphiles. Adv. Funct. Mater. 2015, 25, 4570–4579. [Google Scholar] [CrossRef]
- Whitehead, K.A.; Langer, R.; Anderson, D.G. Knocking down barriers: Advances in siRNA delivery. Nat. Rev. Drug Discov. 2009, 8, 129–138. [Google Scholar] [CrossRef] [PubMed]
- Kolitz, M.; Cohen-Arazi, N.; Hagag, I.; Katzhendler, J.; Domb, A.J. Biodegradable polyesters derived from amino acids. Macromolecules 2009, 42, 4520–4530. [Google Scholar] [CrossRef]
- Li, N.; Zhao, L.; Qi, L.; Li, Z.; Luan, Y. Polymer assembly: Promising carriers as co-delivery systems for cancer therapy. Prog. Polym. Sci. 2016, 58, 1–26. [Google Scholar] [CrossRef]
- Tavares, A.J.; Chong, L.; Petryayeva, E.; Algar, W.R.; Krull, U.J. Quantum dots as contrast agents for in vivo tumor imaging: Progress and issues. Anal. Bioanal. Chem. 2011, 399, 2331–2342. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Z.; Thorek, D.L.J.; Tsourkas, A. Porous polymersomes with encapsulated Gd-labeled dendrimers as highly efficient MRI contrast agents. Adv. Funct. Mater. 2009, 19, 3753–3759. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Z.; Tsourkas, A. Paramagnetic porous polymersomes. Langmuir 2008, 24, 8169–8173. [Google Scholar] [CrossRef] [PubMed]
- Kamaly, N.; Miller, A.D. Paramagnetic liposome nanoparticles for cellular and tumour imaging. Int. J. Mol. Sci. 2010, 11, 1759–1776. [Google Scholar] [CrossRef] [PubMed]
- Langereis, S.; Geelen, T.; Grüll, H.; Strijkers, G.J.; Nicolay, K. Paramagnetic liposomes for molecular MRI and MRI-guided drug delivery. NMR Biomed. 2013, 26, 1195. [Google Scholar] [CrossRef]
- Chen, N.; He, Y.; Su, Y.; Li, X.; Huang, Q.; Wang, H.; Zhang, X.; Tai, R.; Fan, C. The cytotoxicity of cadmium-based quantum dots. Biomaterials 2012, 33, 1238–1244. [Google Scholar] [CrossRef] [PubMed]
- Immordino, M.L.; Dosio, F.; Cattel, L. Stealth liposomes: Review of the basic science, rationale, and clinical applications, existing and potential. Int. J. Nanomed. 2006, 1, 297–315. [Google Scholar]
- Huang, C.-H.; Tsourkas, A. Gd-based macromolecules and nanoparticles as magnetic resonance contrast agents for molecular imaging. Curr. Top. Med. Chem. 2013, 13, 411–421. [Google Scholar] [CrossRef] [PubMed]
- Mohr, K.; Müller, S.S.; Müller, L.K.; Rusitzka, K.; Gietzen, S.; Frey, H.; Schmidt, M. Evaluation of multifunctional liposomes in human blood serum by light scattering. Langmuir 2014, 30, 14954–14962. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, H.; Kawamoto, S.; Jo, S.-K.; Bryant, H.L.; Brechbiel, M.W.; Star, R.A. Macromolecular MRI contrast agents with small dendrimers: Pharmacokinetic differences between sizes and cores. Bioconjug. Chem. 2003, 14, 388–394. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Peng, J.; Han, M.; Omar, M.; Hu, D.; Ke, X.; Lu, N. A low-molecular-weight heparin-coated doxorubicin-liposome for the prevention of melanoma metastasis. J. Drug Target. 2015, 23, 335–346. [Google Scholar] [CrossRef] [PubMed]
- Kluza, E.; Yeo, S.Y.; Schmid, S.; van der Schaft, D.W.J.; Boekhoven, R.W.; Schiffelers, R.M.; Storm, G.; Strijkers, G.J.; Nicolay, K. Anti-tumor activity of liposomal glucocorticoids: The relevance of liposome-mediated drug delivery, intratumoral localization and systemic activity. J. Control. Release 2011, 151, 10–17. [Google Scholar] [CrossRef] [PubMed]
- Kramer, J.R.; Rodriguez, A.R.; Choe, U.-J.; Kamei, D.T.; Deming, T.J. Glycopolypeptide conformations in bioactive block copolymer assemblies influence their nanoscale morphology. Soft Matter 2013, 9, 3389–3395. [Google Scholar] [CrossRef]
- Voskuhl, J.; Stuart, M.C.A.; Ravoo, B.J. Sugar-decorated sugar vesicles: Lectin–carbohydrate recognition at the surface of cyclodextrin vesicles. Chem. A Eur. J. 2010, 16, 2790–2796. [Google Scholar] [CrossRef] [PubMed]
- Jayaraman, N.; Maiti, K.; Naresh, K. Multivalent glycoliposomes and micelles to study carbohydrate-protein and carbohydrate-carbohydrate interactions. Chem. Soc. Rev. 2013, 42, 4640–4656. [Google Scholar] [CrossRef] [PubMed]
- Rigaud, J.-L. Membrane proteins: Functional and structural studies using reconstituted proteoliposomes and 2-D crystals. Braz. J. Med. Biol. Res. 2002, 35, 753–766. [Google Scholar] [CrossRef] [PubMed]
- Hara, M.; Ueno, T.; Fujii, T.; Yang, Q.; Asada, Y.; Miyake, J. Orientation of photosynthetic reaction center reconstituted in neutral and charged liposomes. Biosci. Biotechnol. Biochem. 1997, 61, 1577–1579. [Google Scholar] [CrossRef] [PubMed]
- Safont-Sempere, M.M.; Fernández, G.; Würthner, F. Self-sorting phenomena in complex supramolecular systems. Chem. Rev. 2011, 111, 5784–5814. [Google Scholar] [CrossRef] [PubMed]
- Blin, J.L.; Henzel, N.; Stébé, M.J. Mixed fluorinated–hydrogenated surfactant-based system: Preparation of ordered mesoporous materials. J. Colloid Interface Sci. 2006, 302, 643–650. [Google Scholar] [CrossRef] [PubMed]
- Peyre, V.; Patil, S.; Durand, G.; Pucci, B. Mixtures of hydrogenated and fluorinated lactobionamide surfactants with cationic surfactants: Study of hydrogenated and fluorinated chains miscibility through potentiometric techniques. Langmuir 2007, 23, 11465–11474. [Google Scholar] [CrossRef] [PubMed]



















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Topology | Core | Hydrophobic dendron | Hydrophilic dendron | Solvent | Ref. |
|---|---|---|---|---|---|
| Single–single JDs | Ether | G3 Poly(benzyl ether) dendron | Benzyl-terminated aliphatic polyether dendron | Tetrahydrofuran/diisopropylether mixture | [75] |
| Ester | G3 Poly(benzyl ether) dendron | G3-dimethylol propionic acid (bis-MPA) | Chloroform | [76] | |
| Azide | Y-shaped dendron, including two triazoles, central benzene unit and arylalkyl ether with C5 alkyl chains | Aliphatic ether dendron with oligo(ethylene oxide) chains | n-Hexane | [68] |
| Hydrophilic dendron | Ref. |
|---|---|
| G1-G3 hydroxy-terminated bis-MPA dendron | [58,74,76] |
| G2 bis-MPA dendron terminated with tri(ethylene glycol) monomethyl ether chains | [21] |
| (3,4)/(3,5)/(3,4,5)G1 Percec-type dendron with tri(ethylene glycol) monomethyl ether chains | [21,63,65,66,77] |
| (3,4)/(3,5)/(3,4,5)G1 Percec-type dendron with tri(ethylene glycol) chains | [21,74] |
| (3,4,5)G1 Percec-type dendron with glycerol | [21] |
| G1-G3 thioglycerol dendron | [21] |
| Saccharide, such as d-mannose, d-galactose, d-lactose | [72,73,74,78,79,80] |
| G1 Percec-type dendron substituted with tri(ethylene glycol) monomethyl ether chains and saccharides | [73,74,78,79,80] |
| G1.5/G3 PAMAM-dendron | [32,69] |
| (3,4,5)G1 Percec-type dendron with quaternary ammonium salts end groups | [21] |
| (3,4,5)G1 Percec-type dendron with t-butylcarbamate end groups | [21] |
| Aliphatic ether dendron with oligo(ethylene oxide) chains | [68] |
| Hydrophobic dendron | Ref. |
|---|---|
| (3,4)/(3,5)/(3,4,5)G1 Percec-type dendron with C6-C8/C12 alkyl chains or with fluorinated chains | [21,64,65,66,70,71,72,73,74,77,78,79,80,81] |
| (3,5)G1 Percec-type dendron with C9-C11 alkyl chains | [21] |
| (3,4,5)G1 Percec-type dendron with C4 alkyl chains | [21] |
| (3,4)G1 Percec-type dendron with branched hexyl groups | [21,72,78] |
| G1 bis-MPA dendron terminated with (3,5)-Percec-type dendron with C12 alkyl chains | [73,74] |
| G2 bis-MPA dendron terminated with C12 alkyl chains | [21] |
| G3 bis-MPA dendron terminated with C17 alkyl chains | [58] |
| Arylether dendron | [69,76] |
| PAMAM dendron terminated with C18 alkyl chains | [32] |
| Methacryloyl-terminated C6 alkyl chain | [61,62] |
| (3,4,5)G1 aniline dendron with C12 alkyl chains | [21] |
| Y-shaped dendron, including two triazoles, central benzene unit and arylalkyl ether with C5 alkyl chains | [68] |
| Photodegradable G3-polyester dendron with o-nitrobenzyl units | [63] |
| Topology | Core | Remarks | Ref. |
|---|---|---|---|
| Twin–twin JD | Pentaerythritol | - | [21,64,67,70,71,72,73,74,77,78,79,80] |
| Pentaerythritol monosubstituted with fluorescent dye coumarin | To demonstrate coassembly of DSs with bacterial membrane vesicles (BMVs) | [74] | |
| Single–single JD | Amino acids, such as l-alanine, l-phenylalanine, glycine, l-isoleucine, l-valine | For the construction of constitutional isomeric libraries | [65,66] |
| H-bonding-motif: formation of onion-like DSs | |||
| 1,2-Propanediol | For the construction of constitutional isomeric libraries | [66] | |
| Ethylene glycol core | Vesicles for MR-molecular imaging | [64,81] | |
| Amine/amide-/ester-/ether-/azide-core | - | [21,32,58,65,68,69,73,74,75,76,78,79,80] | |
| Perylene bisimide | Perylene vesicles as fluorescent pH sensor | [61,62] |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Taabache, S.; Bertin, A. Vesicles from Amphiphilic Dumbbells and Janus Dendrimers: Bioinspired Self-Assembled Structures for Biomedical Applications. Polymers 2017, 9, 280. https://doi.org/10.3390/polym9070280
Taabache S, Bertin A. Vesicles from Amphiphilic Dumbbells and Janus Dendrimers: Bioinspired Self-Assembled Structures for Biomedical Applications. Polymers. 2017; 9(7):280. https://doi.org/10.3390/polym9070280
Chicago/Turabian StyleTaabache, Soraya, and Annabelle Bertin. 2017. "Vesicles from Amphiphilic Dumbbells and Janus Dendrimers: Bioinspired Self-Assembled Structures for Biomedical Applications" Polymers 9, no. 7: 280. https://doi.org/10.3390/polym9070280
APA StyleTaabache, S., & Bertin, A. (2017). Vesicles from Amphiphilic Dumbbells and Janus Dendrimers: Bioinspired Self-Assembled Structures for Biomedical Applications. Polymers, 9(7), 280. https://doi.org/10.3390/polym9070280