Soil Pollution by Petroleum-Derived Substances and its Bioremediation: The Effect on Aphis fabae Scop. Infestation and Antioxidant Response in Vicia faba L.



Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Setup
2.2. Plants
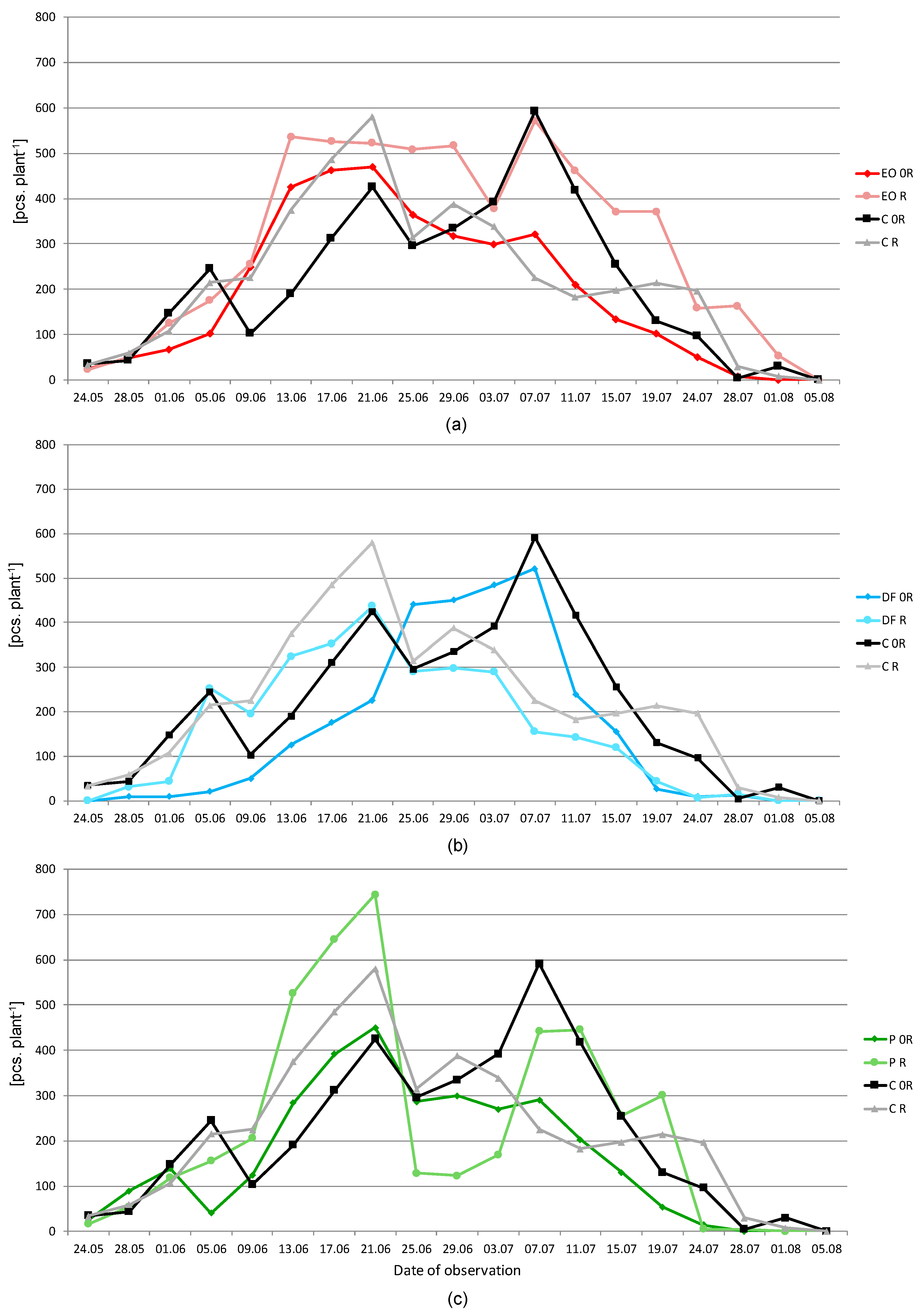
2.3. Aphis fabae Scop
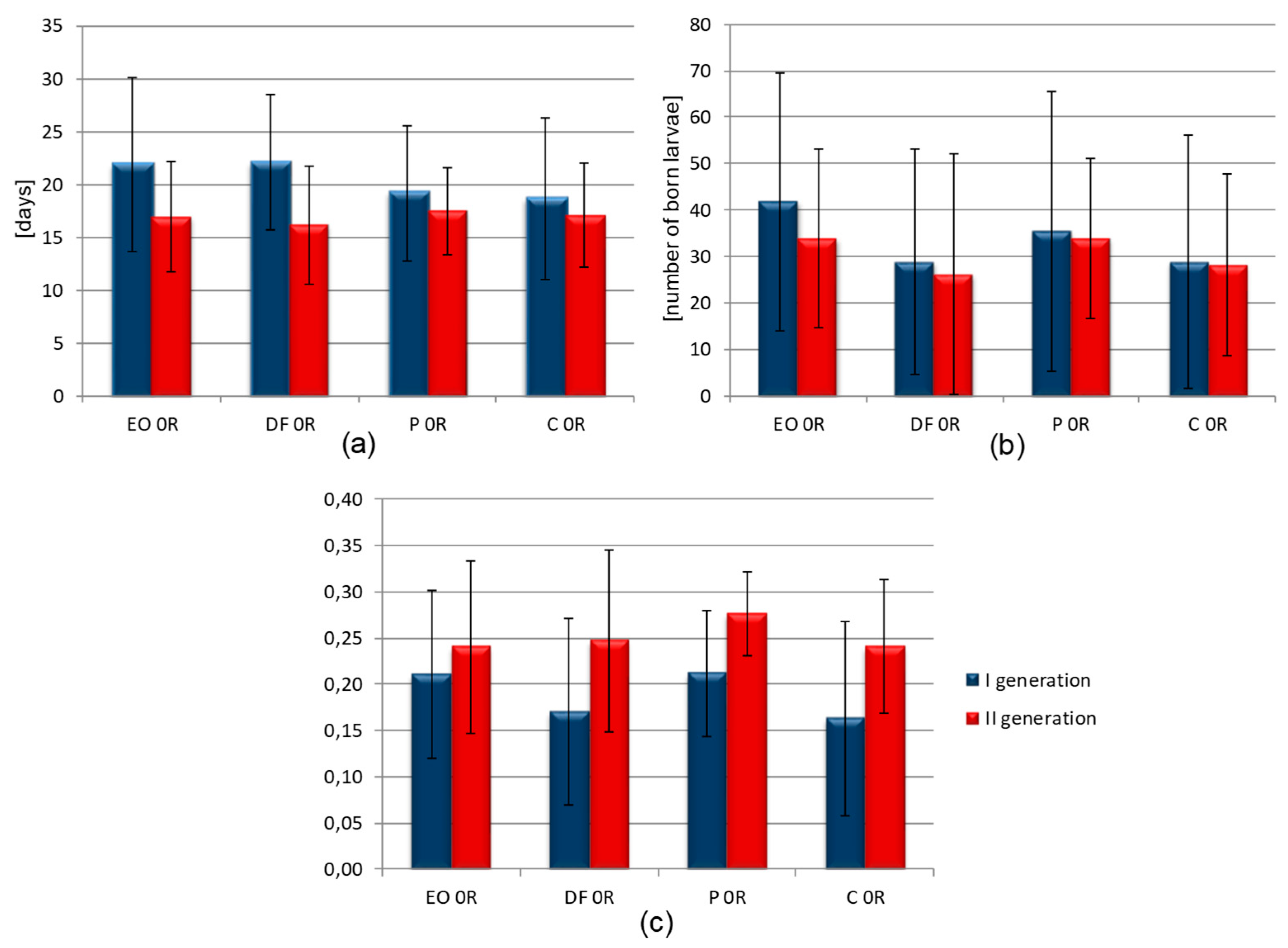
- rm is the population intrinsic growth rate,
- d is the duration of pre-reproductive period (from birth to producing the first offspring),
- Md is the mean number of larvae born in the period from d to 2d days from birth.
2.4. Analysis of the Biochemical Parameters of the Plants
2.5. Statistical Analysis
3. Results
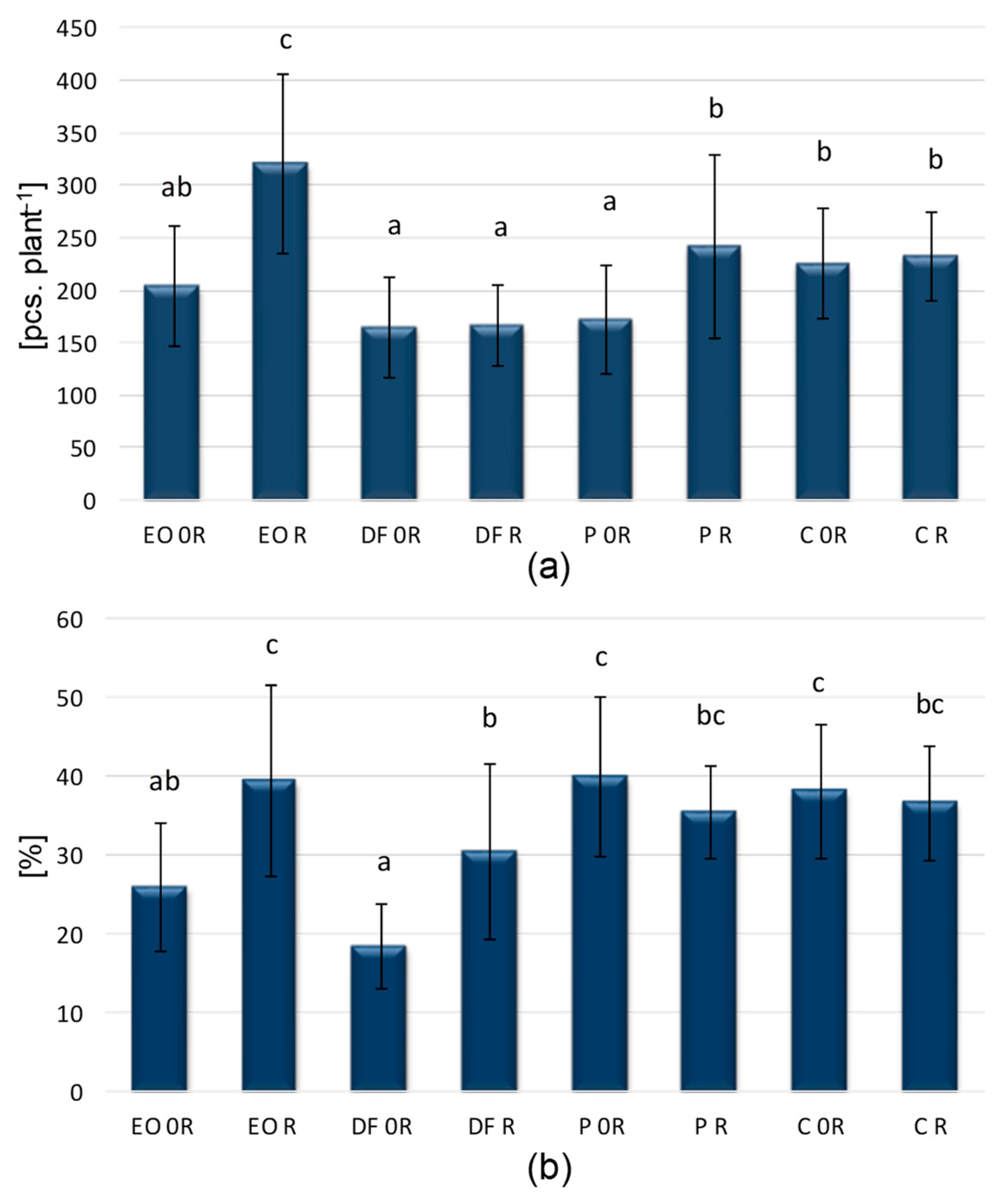
3.1. Aphids
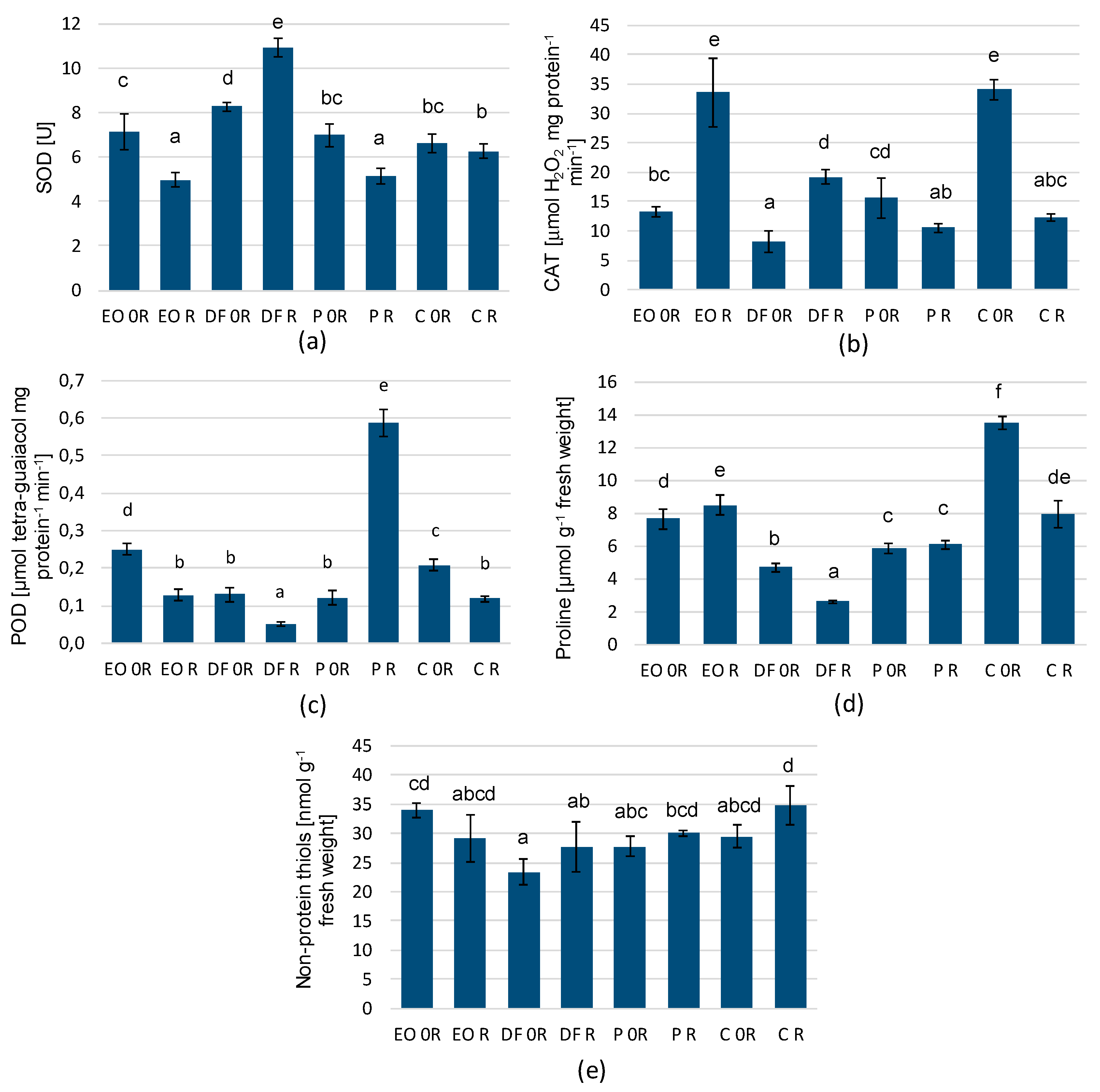
3.2. Analysis of the Biochemical Parameters of the Plants
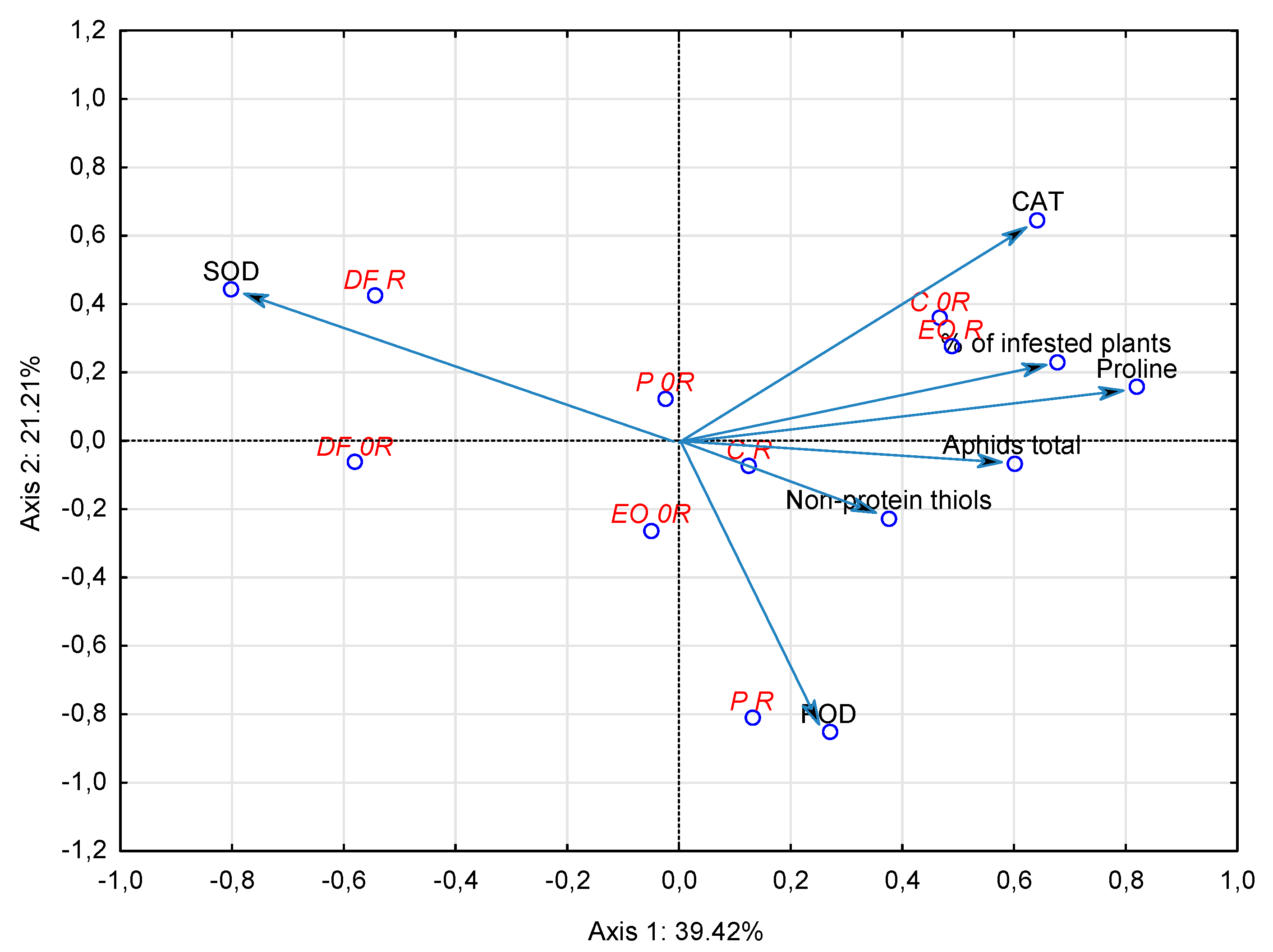
3.3. Relationships between Aphis fabae Occurrence, Biochemical Parameters and Soil Contamination with PDSs
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Abdel-Shafy, H.I.; Mansour, M.S.M. A review on polycyclic aromatic hydrocarbons: Source, environmental impact, effect on human health and remediation. Egypt. J. Pet. 2016, 25, 107–123. [Google Scholar] [CrossRef] [Green Version]
- Schaefer, M.; Filser, J. The influence of earthworms and organic additives on the biodegradation of oil contaminated soil. Appl. Soil Ecol. 2007, 36, 53–62. [Google Scholar] [CrossRef]
- Gkorezis, P.; Daghio, M.; Franzetti, A.; Van Hamme, J.D.; Sillen, W.; Vangronsveld, J. The interaction between plants and bacteria in the remediation of petroleum hydrocarbons, an environmental perspective. Front. Microbiol. 2016, 21, 18–36. [Google Scholar] [CrossRef]
- Truskewycz, A.; Gundry, T.D.; Khudur, L.S.; Kolobaric, A.; Taha, M.; Aburto-Medina, A.; Ball, A.S.; Shahsavari, E. Petroleum Hydrocarbon Contamination in Terrestrial Ecosystems-Fate and Microbial Responses. Molecules 2019, 24, 3400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wyszkowski, M.; Sivitskaya, V. Effect of sorbents on the content of trace elements in maize cultivated on soil contaminated with heating oil. Int. Agrophys. 2019, 33, 437–444. [Google Scholar] [CrossRef]
- Jyothi, K.; Babu, K.S.; Clara, N.K.; Kashyap, A. Identification and isolation of hydrocarbon degrading bacteria by molecular characterization. Helix 2012, 2, 105–111. [Google Scholar]
- Langenbach, T. Persistence and bioaccumulation of persistent organic pollutants (POPs), applied bioremediation: Active and passive approaches. Appl. Biorem. 2013, 394, 307–332. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.Y.; Kuo, Y.C.; Hong, A.; Chang, Y.M.; Kao, C.M. Bioremediation of diesel and lubricant oil-contaminated soils using enhanced landfarming system. Chemosphere 2016, 164, 558–567. [Google Scholar] [CrossRef]
- Azubuike, C.C.; Chikere, C.B.; Okpokwasili, G.C. Bioremediation techniques–classification based on site of application: Principles, advantages, limitations and prospects. World J. Microbiol. Biotechnol. 2016, 32, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Varjami, S.J. Microbial degradation of petroleum hydrocarbons. Bioresour. Technol. 2017, 223, 277–286. [Google Scholar] [CrossRef]
- Gbadebo, A.M.; Adenuga, M.D. Effect of crude oil on the emergence and growth of cowpea in two contrasting soil types from Abeokuta, Southwestern Nigeria. Asian J. Appl. Sci. 2012, 5, 232–239. [Google Scholar] [CrossRef] [Green Version]
- Rusin, M.; Gospodarek, J.; Nadgórska-Socha, A. The effect of petroleum-derived substances on the growth and chemical composition of Vicia faba L. Pol. J. Environ. Stud. 2015, 24, 2157–2166. [Google Scholar] [CrossRef]
- Pawlak-Sprada, S.; Arasimowicz-Jelonek, M.; Podgórska, M.; Deckert, J. Activation of phenylpropanoid pathway in legume plants exposed to heavy metals. Part I. Effects of cadmium and lead on phenylalanine ammonia-lyase gene expression, enzyme activity and lignin content. Acta Biochim. Pol. 2011, 58, 211–216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rusin, M.; Gospodarek, J.; Barczyk, G.; Nagórska-Socha, A. Antioxidant responses of Triticum aestivum plants to petroleum-derived substances. Ecotoxicology 2018, 27, 1353–1367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marti, M.C.; Camejo, D.; Fernández-García, N.; Rellán-Álvarez, R.; Marques, S.; Sevilla, F.; Jiménez, A. Effect of oil refinery sludges on the growth and antioxidant system of alfalfa plants. J. Hazard. Mater. 2009, 171, 879–885. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muszyńska, E.; Labudda, M.; Różańska, E.; Hanus-Fajerska, E.; Znojeka, E. Heavy metal tolerance in contrasting ecotypes of Alyssum montanum. Ecotox. Environ. Safe. 2018, 161, 305–317. [Google Scholar] [CrossRef] [PubMed]
- Jiang, K.; Wu, B.; Wanga, C.; Ran, Q. Ecotoxicological effects of metals with different concentrations and types on the morphological and physiological performance of wheat. Ecotox. Environ. Safe. 2019, 167, 345–353. [Google Scholar] [CrossRef]
- Rusin, M.; Gospodarek, J.; Nadgórska-Socha, A. Effect of petroleum-derived substances on life history traits of black bean aphid (Aphis fabae Scop.) and on the growth and chemical composition of broad bean. Ecotoxicology 2017, 26, 308–319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rusin, M.; Gospodarek, J.; Nadgórska-Socha, A.; Barczyk, G.; Boligłowa, E.; Dabioch, M. Effect of petroleum-derived substances on life history traits of bird cherry-oat aphid (Rhopalosiphum padi L.) and on the growth and chemical composition of winter wheat. Environ. Sci. Pollut. Res. 2018, 25, 27000–27012. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harrington, R.; Stork, N.E. Insect in a changing Environment. In Proceedings of the 17th Symposium of the Royal Entomological Society, Harpenden, UK, 7–10 September 1993; Academic Press: London, UK, 1995; p. 535. [Google Scholar]
- He, J.; Chen, F.; Chen, S.; Lv, G.; Deng, Y.; Fang, W.; Liu, Z.; Guan, Z.; He, C. Chrysanthemum leaf epidermal surface morphology and antioxidant and defense enzyme activity in response to aphid infestation. J. Plant Physiol. 2011, 168, 687–693. [Google Scholar] [CrossRef]
- Zhao, H.; Sun, X.; Xue, M.; Zhang, X.; Li, Q. Antioxidant Enzyme Responses Induced by Whiteflies in Tobacco Plants in Defense against Aphids, Catalase May Play a Dominant Role. PLoS ONE 2016, 11. [Google Scholar] [CrossRef] [Green Version]
- Malallah, G.; Afzal, M.; Gulshan, S.; Abraham, D.; Kurian, M.; Dhami, M.S.I. Vicia faba as a bioindicator of oil pollution. Environ. Pollut. 1996, 92, 213–217. [Google Scholar] [CrossRef]
- Grant, W.F.; Lee, H.G.; Logan, D.M.; Salamone, M.F. The use of Tradescantia and Vicia faba bioassays for the in situ detection of mutagens in an aquatic environment. Mutat. Res. 1992, 270, 53–64. [Google Scholar] [CrossRef]
- Kanaya, N.; Gill, B.S.; Grover, I.S.; Murin, A.; Osiecka, R.; Sandhu, S.S.; Anderson, H.C. Vicia faba chromosomal aberation assay. Mutat. Res. 1994, 310, 231–247. [Google Scholar] [CrossRef]
- Nadgórska-Socha, A.; Kafel, A.; Kandziora-Ciupa, M.; Gospodarek, J.; Zawisza-Raszka, A. Accumulation of heavy metals and antioxidant responses in Vicia faba plants grown on monometallic contaminated soil. Environ. Sci. Pollut. Res. 2013, 20, 1124–1134. [Google Scholar] [CrossRef]
- Gospodarek, J.; Petryszak, P.; Koloczek, H. The effect of the bioremediation of soil contaminated with petroleum derivatives on the occurrence of epigeic and edaphic fauna. Bioremediat. J. 2016, 20, 38–53. [Google Scholar] [CrossRef]
- Wyatt, I.J.; White, P.F. Simple estimation of intrinsic increase rates for aphids and tetranychid mites. J. Appl. Ecol. 1977, 14, 757–766. [Google Scholar] [CrossRef]
- Dazy, M.; Jung, V.; Férard, J.F.; Masfaraud, J.F. Ecological recovery of vegetation on a coke-factory soil, role of plant antioxidant enzymes and possible implications in site restoration. Chemosphere 2008, 74, 57–63. [Google Scholar] [CrossRef]
- Nadgórska-Socha, A.; Ptasiński, B.; Kita, A. Heavy metal bioaccumulation and antioxidative responses in Cardaminopsis arenosa and Plantago lanceolata leaves from metalliferous and non-metalliferous sites: A field study. Ecotoxicology 2013, 22, 1422–1434. [Google Scholar] [CrossRef] [Green Version]
- Fang, W.C.; Kao, C. Enhanced peroxidase activity in rice leaves in response to excess iron, copper and zinc. Plant Sci. 2000, 158, 71–76. [Google Scholar] [CrossRef]
- Aebi, H. Catalase in vitro. Methods Enzymol. 1984, 105, 121–126. [Google Scholar] [PubMed]
- Beauchamp, C.; Fridovich, I. Superoxide dismutase: Improved assays and an assay applicable to acrylamide gels. Anal. Biochem. 1971, 44, 276–287. [Google Scholar] [CrossRef]
- Bradford, M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein dye binding. Anal. Biochem. 1976, 44, 276–287. [Google Scholar] [CrossRef]
- Mass, F.; De Kok, L.; Peters, J.; Kuiper, P.A. Comparative study on the effects of H2S and SO2 fumigation on the growth and accumulation of sulfate and sulfhydryl compounds in Trifolium pratense L., Glycine max Merr., Phaseolus vulgaris L. J. Exp. Bot. 1987, 38, 1459–1469. [Google Scholar] [CrossRef]
- Bates, L.; Waldren, R.; Teare, D. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Cichocka, E.; Leszczyński, B.; Ciepiela, A.P.; Goszczyński, W. Response of Aphis fabae Scop. to different broad bean cultivars. EJPAU Horticulture 2002, 5. [Google Scholar]
- Wojciechowicz-Żytko, E. The effect of broad bean plants density on the occurrence of Aphis fabae Scop. (Homoptera, Aphidodea) and predatory syrphids (Diptera, Syrphidae). Veget. Crops Res. Bull. 2006, 64, 77–84. [Google Scholar]
- Gospodarek, J. Occurrence of black bean aphid (Aphis fabae Scop.) and its natural predators on broad bean under the conditions of soil pollution with heavy metals. Zesz. Nauk. UR w Krakowie 2012, 480, 207. (In Polish) [Google Scholar]
- Gospodarek, J.; Nadgorska-Socha, A. Chemical composition of broad beans (Vicia faba L.) and development parameters of black bean aphid (Aphis fabae Scop.) under conditions of soil contamination with oil derivatives. J. Elementol. 2016, 21, 1359–1376. [Google Scholar] [CrossRef]
- Davies, M.T.; Port, G.R.; Davison, A.W. Effects of dietary and gaseous fluoride on the aphid Aphis fabae. Environ. Pollut. 1998, 99, 405–409. [Google Scholar] [CrossRef]
- Gospodarek, J. The effect of zinc on development of Aphis fabae Scop. Post. Ochr. Roś./Progr. Plant Protect. 2005, 45, 680–682. (in Polish). [Google Scholar]
- Douglas, A.E. Provenance, experience and plant utilisation by the polyphagous aphid, Aphis fabae. Entomol. Exp. Appl. 1997, 83, 161–170. [Google Scholar] [CrossRef]
- Nwaichi, E.O.; Wegwu, M.O.; Nwosu, U.L. Distribution of selected carcinogenic hydrocarbon and heavy metals in an oil-polluted agriculture zone. Environ. Monit. Assess. 2014, 186, 8697–8706. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gospodarek, J.; Rusin, M.; Nadgórska-Socha, A. Effect of petroleum-derived substances and their bioremediation on Triticum aestivum L. growth and chemical composition. Pol. J. Environ. Stud. 2019, 28, 2131–2137. [Google Scholar] [CrossRef]
- Gospodarek, J.; Rusin, M. Residual effect of soil pollution with oil derivatives on the occurrence of Acarina. Proc. ECOpole 2015, 9, 71–77. [Google Scholar]
- Rusin, M.; Gospodarek, J. The occurrence of springtails (Collembola) andspiders (Araneae) as an effectiveness indicator of bioremediation of soil contaminated by petroleum-derived substances. Int. J. Environ. Res. 2016, 10, 449–458. [Google Scholar]
- Tabassum, S.; Shahid, N.; Wang, J.; Shafiq, M.; Mumtaz, M.; Arslan, M. The oxidative stress response of Mirabilis jalapa to exhausted engine oil (EEO) during phytoremediation. Pol. J. Environ. Stud. 2016, 25, 2581–2587. [Google Scholar] [CrossRef]
- Achuba, F.I. Petroleum products in soil mediated oxidative stress in cowpea (Vigna unguiculata) and maize (Zea mays) seedlings. Open J. Soil Sci. 2014, 4, 417–435. [Google Scholar] [CrossRef] [Green Version]
- Dazy, M.; Béraud, E.; Cotelle, S.; Grévilliot, F.; Férard, J.F.; Masfaraud, J.F. Changes in plant communities along soil pollution gradients: Responses of leaf antioxidant enzyme activities and phytochemlatin contents. Chemosphere 2009, 77, 376–383. [Google Scholar] [CrossRef]
- Kafel, A.; Nadgórska-Socha, A.; Gospodarek, J.; Babczyńska, A.; Skowronek, M.; Kandziora, M.; Rozpendek, K. The effects of Aphis fabae infestation on the antioxidant response and heavy metal content in field grown Philadelphus coronarius plants. Sci. Total Environ. 2010, 408, 1111–1119. [Google Scholar] [CrossRef]
- Tang, M.; Chen, H.; Huang, J.; Tian, Z. AM fungi effects on the growth and physiology of Zea mays seedlings under diesel stress. Soil Biol. Biochem. 2009, 41, 936–940. [Google Scholar] [CrossRef]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef] [PubMed]
- Farzana, S.; Fung Yee Tam, N. A combined effect of polybrominated diphenyl ether and aquaculture effluent on growth and antioxidative response of mangrove plants. Chemosphere 2018, 201, 483–491. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.; Li, J.; Gu, R.; Yu, L.; Wang, H.; Zhan, X.; Xing, B. Carotenoid and superoxide dismutase are the most effective antioxidants participating in ROS scavenging in phenanthrene accumulated wheat leaf. Chemosphere 2018, 197, 513–525. [Google Scholar] [CrossRef] [PubMed]
- Teklić, T.; Hancock, J.T.; Engler, M.; Paradicović, N.; Cesar, V.; Lepeduš, H.; Štolfa, I.; Bešlo, D. Antioxidative responses in radish (Raphanus sativus L.) plants stressed by copper and lead in nutrient solution and soil. Acta Biol. Crac. 2008, 50, 79–86. [Google Scholar]
- John, R.; Ahmad, P.; Gadgil, K.; Sharma, S. Heavy metal toxicity: Effect on plant growth, biochemical parameters and metal accumulation by Brassica juncea L. Int. J. Plant Prod. 2009, 3, 65–75. [Google Scholar]
- Mishra, S.; Tripathi, R.D.; Srivastava, S.; Dwivedi, S.; Trivedi, P.K.; Dhankher, O.P.; Khare, A. Thiol metabolism play significant role during cadmium detoxification by Ceratophyllum demersum L. Bioresour. Technol. 2009, 100, 2155–2161. [Google Scholar] [CrossRef]
- Kerchev, P.I.; Fenton, B.; Foyer, C.H.; Hancock, R.D. Infestation of potato (Solanum tuberosum L.) by the peach-potato aphid (Myzus persicae Sulzer) alters cellular redox status and is influenced by ascorbate. Plant Cell Environ. 2012, 35, 430–440. [Google Scholar] [CrossRef]
- Van-Chung, M.; Ba-Hoanh, N.; Duc-Dien, N.; Le-Ai-Vinh, N. Nostoc calcicola extract improved the antioxidative response of soybean to cowpea aphid. Bot. Stud. 2017, 58. [Google Scholar] [CrossRef] [Green Version]
- Lukasik, I.; Kornacka, A.; Golawska, S.; Sytykiewicz, H.; Sprawka, I.; Wojcicka, A. Effects of Acyrtosiphon pisum (Harris) infestation on the hydrogen peroxide content and activity of antioxidant enzymes in Fabaceae plants. Allelopathy J. 2017, 40, 143–150. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Wingless Females | Winged Females | Larvae |
|---|---|---|---|
| EO 0R | 29.21 (±3.50) a,* | 1.19 (±0.75) abc | 69.60 (±12.58) a |
| EO R | 29.57 (±7.76) a | 0.89 (±0.33) abc | 69.54 (±10.80) a |
| DF 0R | 29.14 (±5.34) a | 1.85 (±0.50) c | 69.01 (±11.66) a |
| DF R | 30.00 (±3.88) a | 0.57 (±0.22) ab | 69.43 (±7.42) a |
| P 0R | 29.52 (±6.36) a | 0.83 (±0.57) ab | 69.65 (±9.34) a |
| P R | 29.17 (±4.14) a | 0.53 (±0.22) a | 70.30 (±3.74) a |
| C 0R | 29.25 (±2.33) a | 1.58 (±0.76) bc | 69.17 (±5.60) a |
| C R | 27.94 (±1.92) a | 1.24 (±0.58) abc | 70.82 (±4.18) a |
| Treatment | Apexes of Plants | Shoots | Leaves | Flowers | Pods |
|---|---|---|---|---|---|
| EO 0R | 25.94 (±6.32) a,* | 52.37 (±12.57) ab | 12.78 (±3.22) ab | 4.99 (±1.38) a | 3.92 (±2.38) a |
| EO R | 25.39 (±4.49) a | 47.45 (±10.77) a | 20.57 (±6.14) b | 3.28 (±0.94) a | 3.31 (±1.76) a |
| DF 0R | 22.69 (±7.13) a | 44.87 (±9.54) a | 19.84 (±5.76) b | 3.04 (±1.20) a | 9.57 (±3.20) a |
| DF R | 34.04 (±4.64) b | 54.20 (±7.81) ab | 5.66 (±1.27) a | 0.87 (±0.62) a | 5.22 (±2.62) a |
| P 0R | 33.86 (±5.67) ab | 47.05 (±11.26) a | 9.56 (±2.77) a | 7.00 (±2.27) a | 2.52 (±1.68) a |
| P R | 35.92 (±8.79) b | 56.44 (±5.39) b | 3.74 (±1.89) a | 3.37 (±1.50) a | 0.52 (±0.50) a |
| C 0R | 30.32 (±6.43) ab | 47.62 (±8.26) a | 14.92 (±6.83) ab | 0.62 (±0.39) a | 6.51 (±3.50) a |
| C R | 35.87 (±7.53) b | 45.60 (±5.65) a | 11.61 (±4.50) ab | 4.53 (±1.71) a | 2.38 (±1.57) a |
| Details | d | Md | ||
|---|---|---|---|---|
| Generation | I | II | I | II |
| EO | 11.44 (±0.81) | 8.69 (±0.59) | 41.78 (±27.68) | 28.31 (±19.34) |
| DF | 11.75 (±4.42) | 8.88 (±0.74) | 28.80 (±24.30) | 34.00 (±25.77) |
| P | 11.19 (±0.98) | 9.06 (±0.25) | 35.40 (±30.22) | 33.93 (±17.25) |
| C | 11.13 (±1.41) | 9.25 (±0.68) | 26.66 (±27.17) | 26.19 (±19.57) |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rusin, M.; Gospodarek, J.; Nadgórska-Socha, A. Soil Pollution by Petroleum-Derived Substances and its Bioremediation: The Effect on Aphis fabae Scop. Infestation and Antioxidant Response in Vicia faba L. Agronomy 2020, 10, 147. https://doi.org/10.3390/agronomy10010147
Rusin M, Gospodarek J, Nadgórska-Socha A. Soil Pollution by Petroleum-Derived Substances and its Bioremediation: The Effect on Aphis fabae Scop. Infestation and Antioxidant Response in Vicia faba L. Agronomy. 2020; 10(1):147. https://doi.org/10.3390/agronomy10010147
Chicago/Turabian StyleRusin, Milena, Janina Gospodarek, and Aleksandra Nadgórska-Socha. 2020. "Soil Pollution by Petroleum-Derived Substances and its Bioremediation: The Effect on Aphis fabae Scop. Infestation and Antioxidant Response in Vicia faba L." Agronomy 10, no. 1: 147. https://doi.org/10.3390/agronomy10010147
APA StyleRusin, M., Gospodarek, J., & Nadgórska-Socha, A. (2020). Soil Pollution by Petroleum-Derived Substances and its Bioremediation: The Effect on Aphis fabae Scop. Infestation and Antioxidant Response in Vicia faba L. Agronomy, 10(1), 147. https://doi.org/10.3390/agronomy10010147