The Influence of Bio-Stimulants and Foliar Fertilizers on Yield, Plant Features, and the Level of Soil Biochemical Activity in White Lupine (Lupinus albus L.) Cultivation

 ,
, 

Abstract
:1. Introduction
2. Material and Methods
2.1. Experimental Design
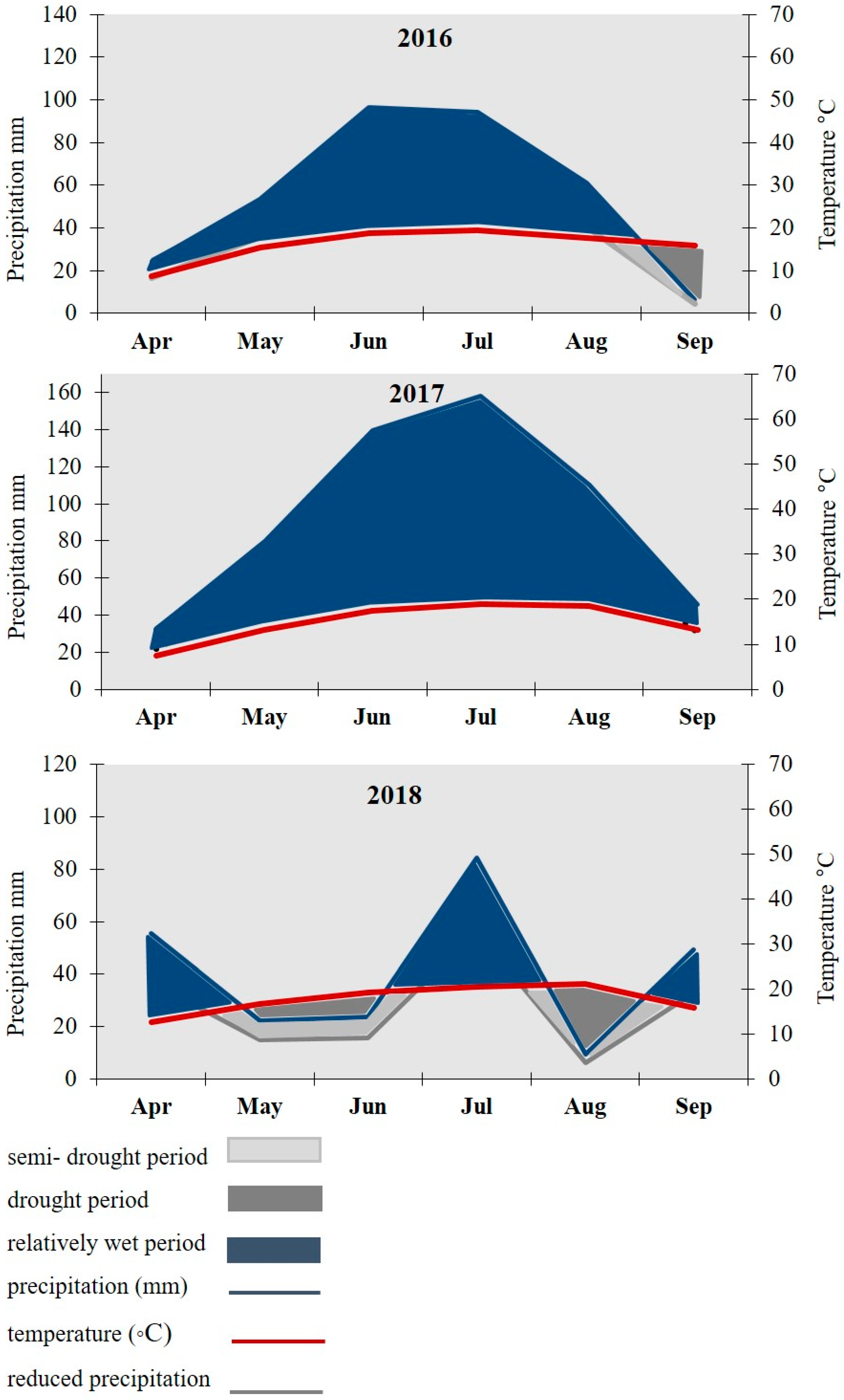
2.2. Weather Conditions
2.3. Influence of Fertilizers on Nitrogenase Activity (Diazotrophy)
2.4. Plant Biometric Assessment
2.5. Chlorophyll Fluorescence and Chlorophyll Content Measurements
2.6. Soil Sampling for Biochemical Analyzes
2.7. Soil Enzymatic Activity
2.8. Biological Index of Fertility
2.9. Statistical Analyses
3. Results
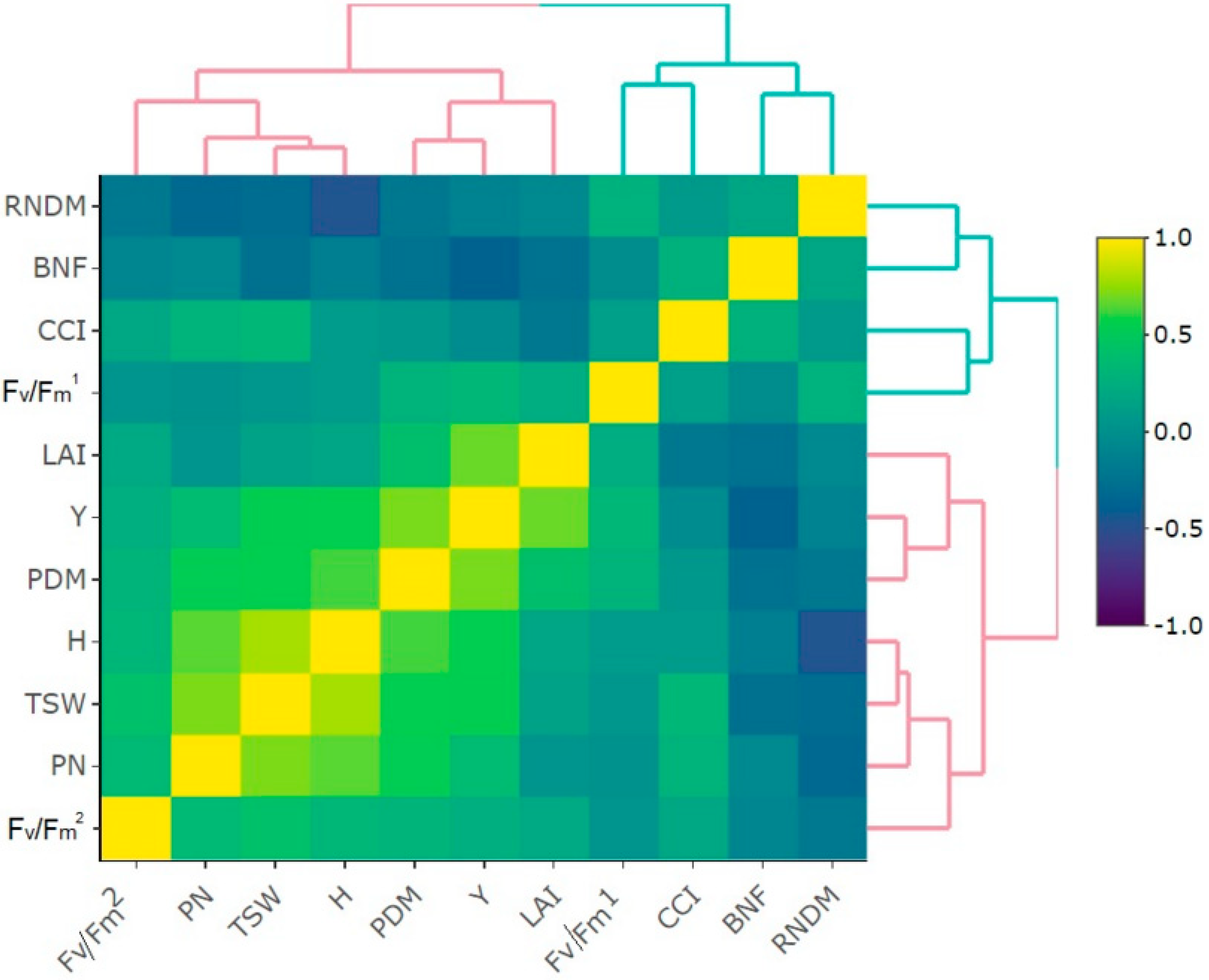
3.1. Yield, Biometric, and Physiological Traits of White Lupine Plants
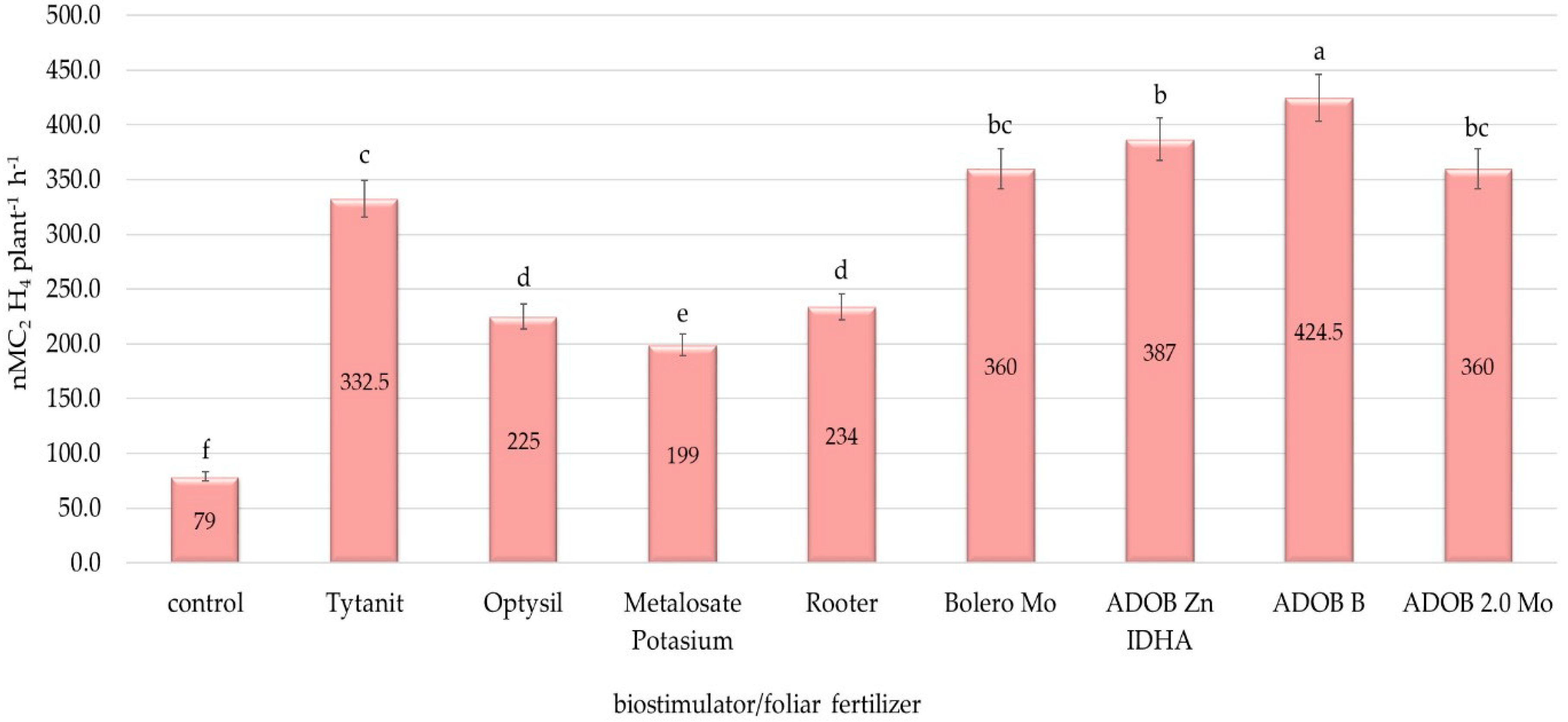
3.2. Biological Fixation of Nitrogen under Lupine Plantation
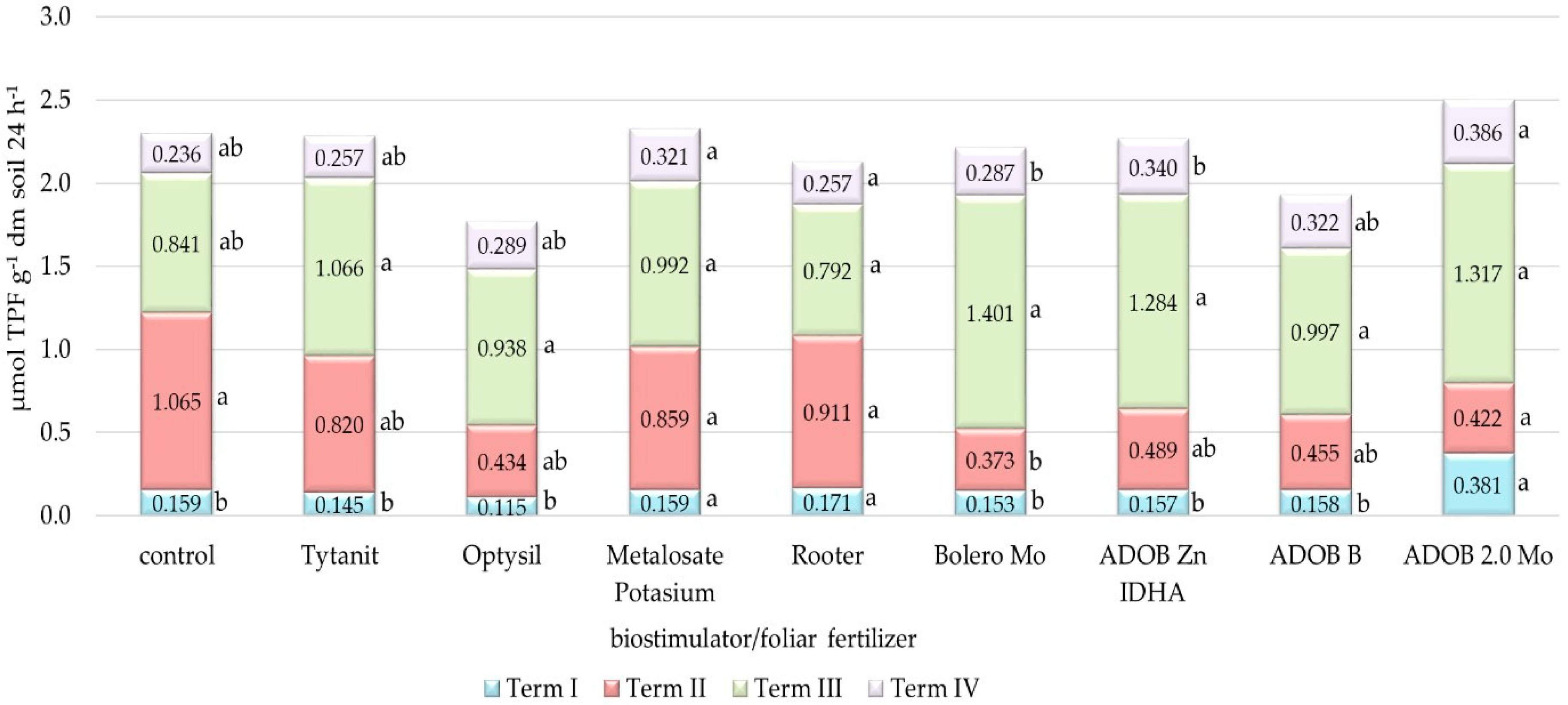
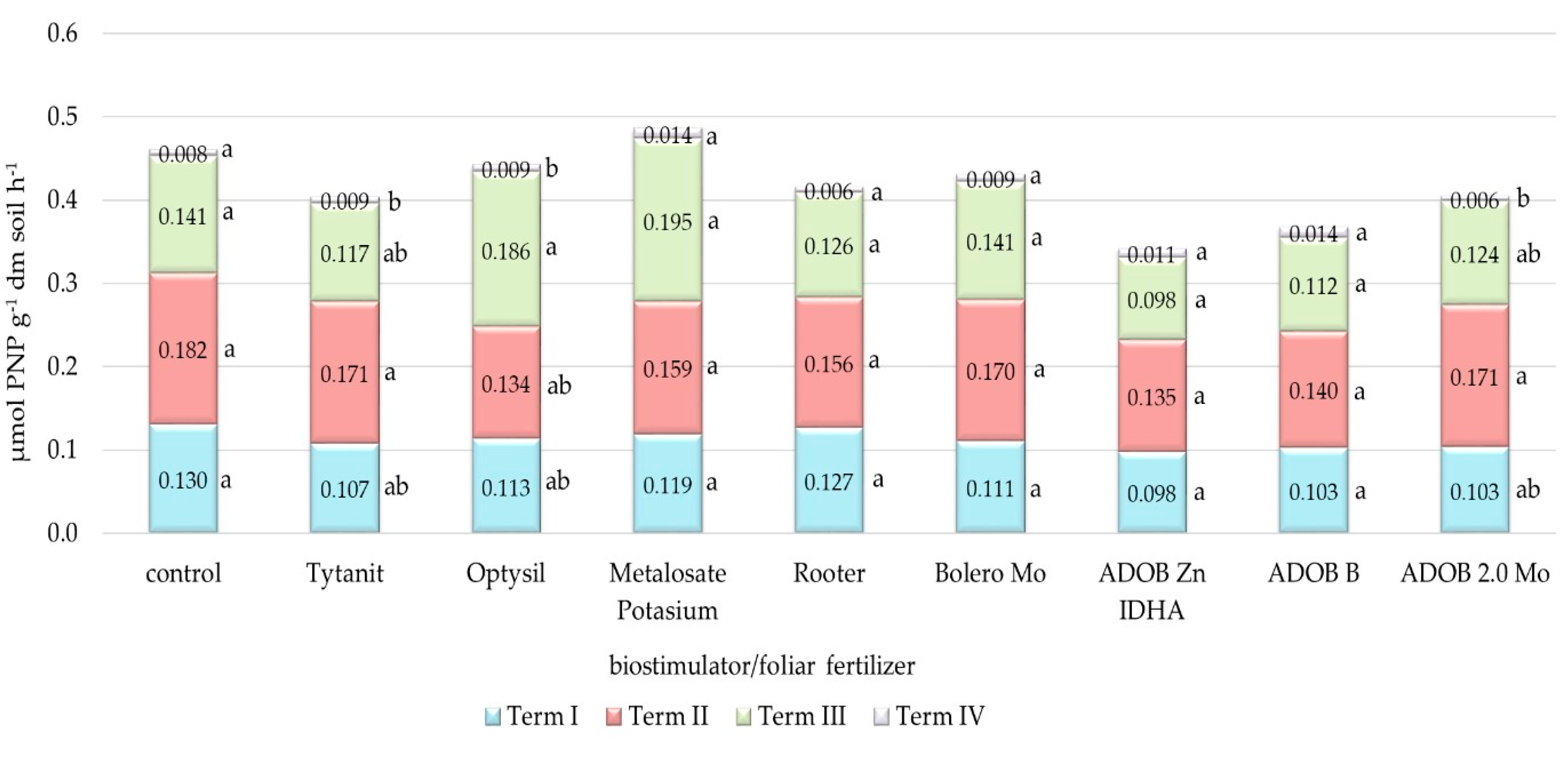
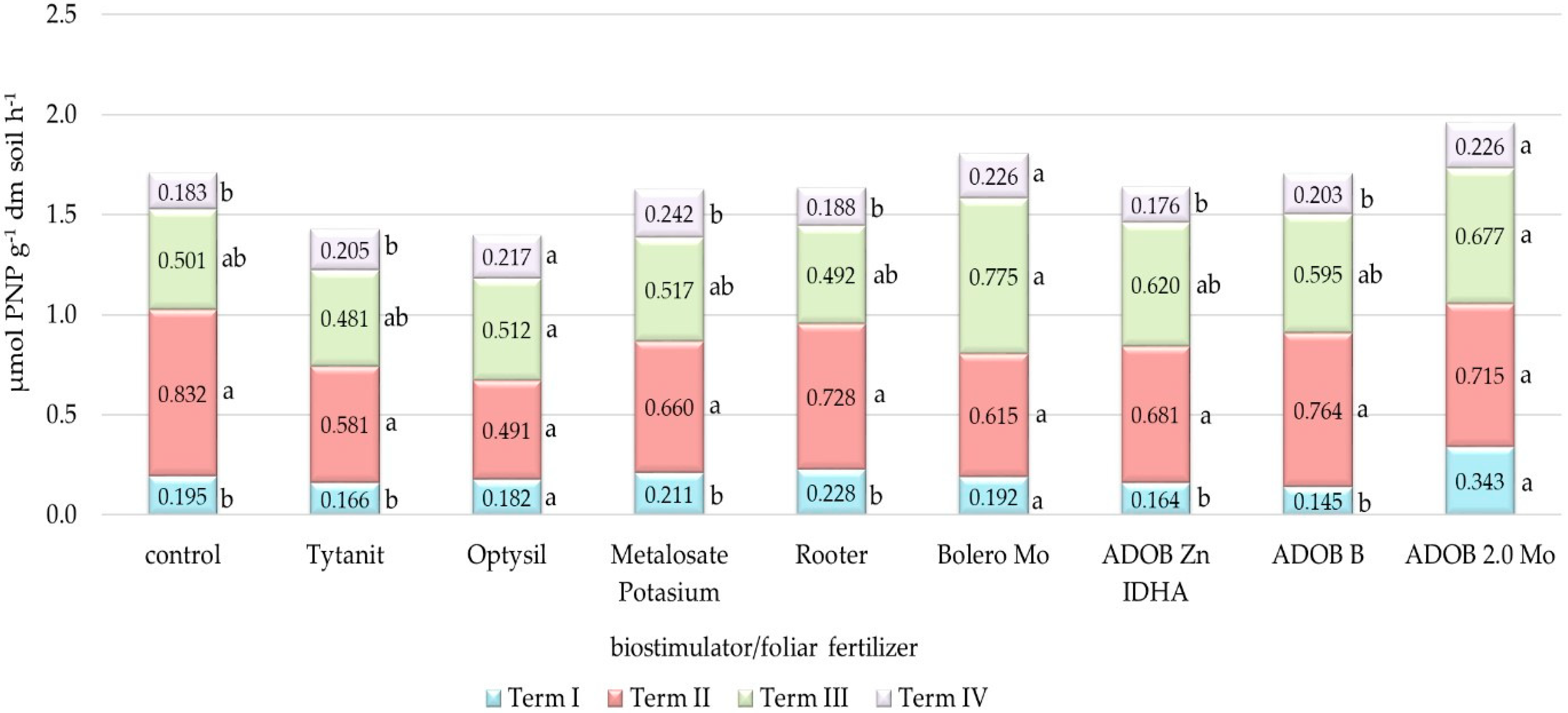
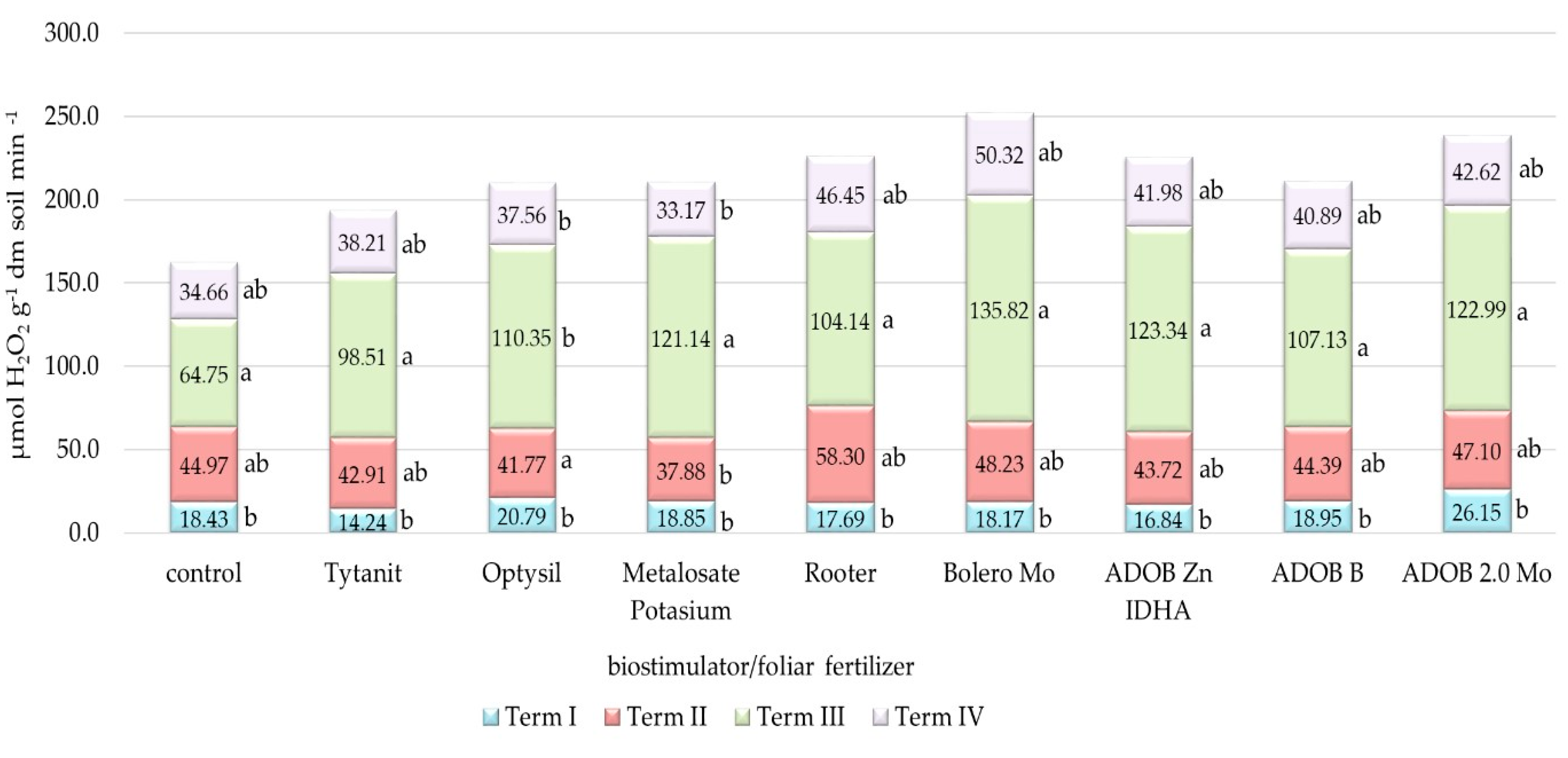
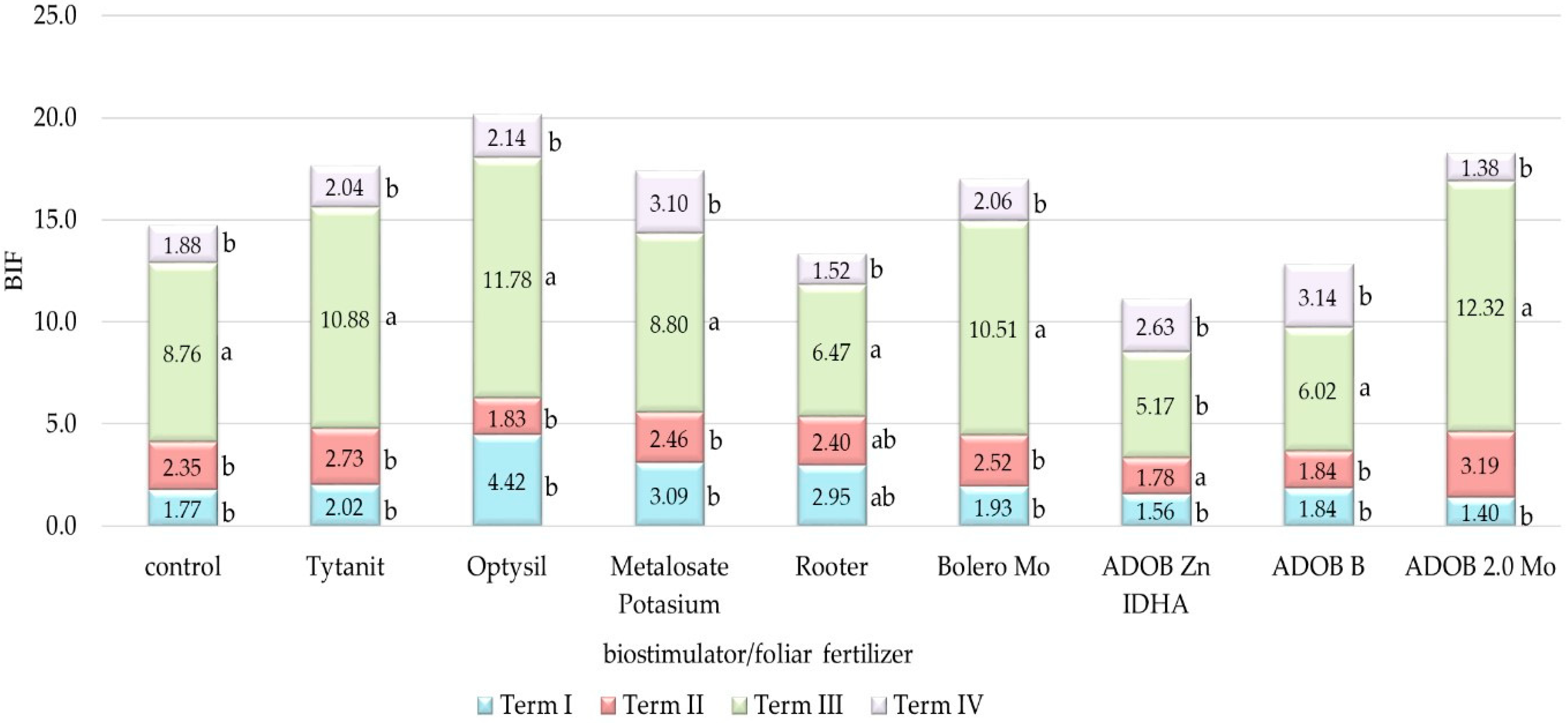
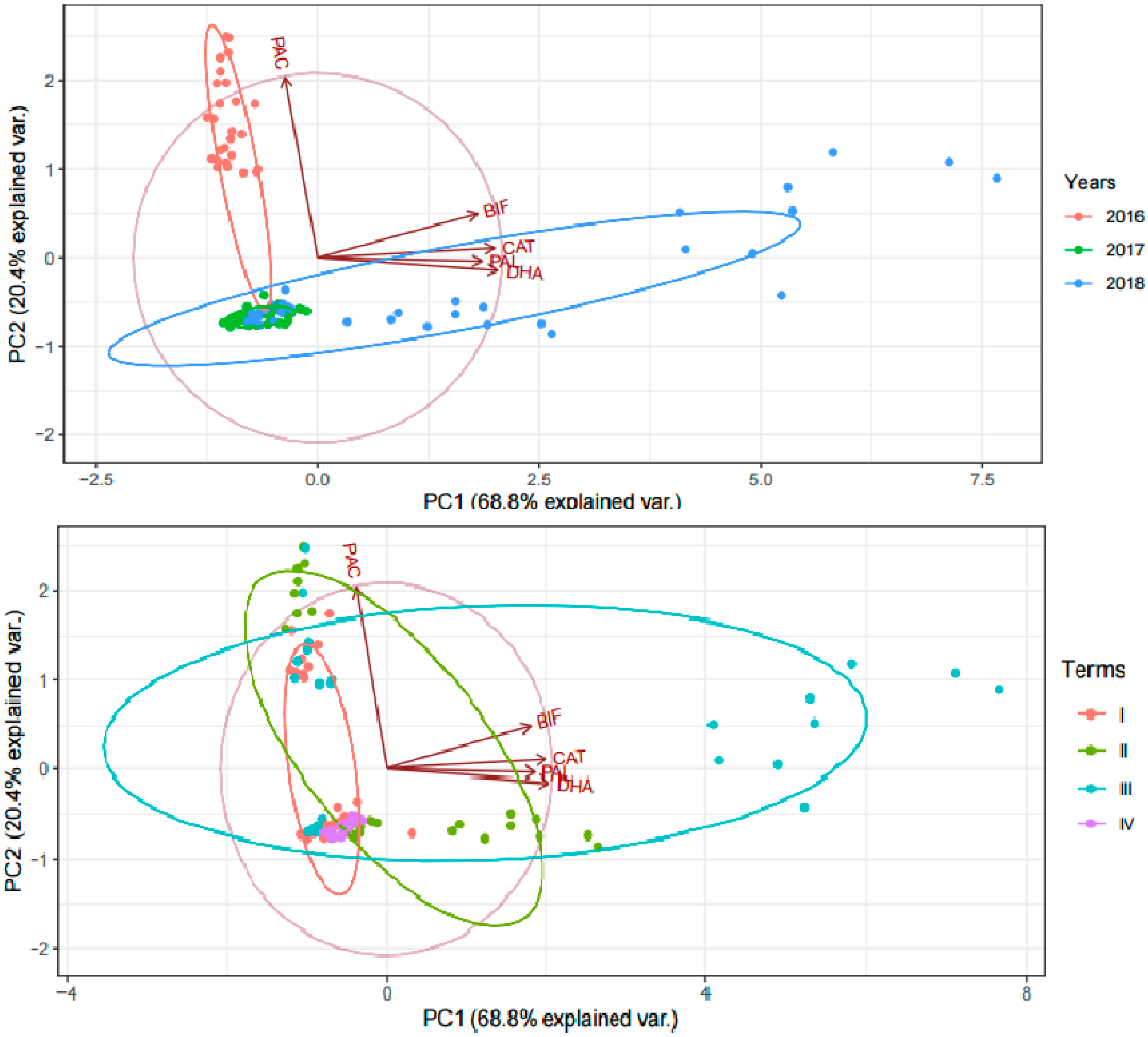
3.3. Analysis of Soil Biochemical Activity
4. Discussion
4.1. Yield, Biometric, and Physiological Traits
4.2. Biological Fixation of Nitrogen
4.3. Biochemical Activity
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Directive 2009/128/EC of the European Parliament and of the Council of 21 October 2009 Establishing a Framework for Community Action to Achieve the Sustainable Use of Pesticides:1-16. Available online: https://ec.europa.eu/food/plant/pesticides/sustainable_use_pesticides_en (accessed on 9 October 2019).
- Małecka-Jankowiak, I.; Blecharczyk, A.; Sawinska, Z.; Waniorek, W. Effect of lupins and pea on the subsequent winter wheat yield depending of tillage and nitrogen fertilization. Fragm. Agron. 2018, 35, 67–79. [Google Scholar]
- Prusinski, J. White lupin (Lupinus albus L.)—Nutritional and health values in human nutrition—A review. Czech J. Food Sci. 2017, 35, 95–105. [Google Scholar]
- GUS. Agriculture in 2015 Year; GUS: Warsaw, Poland, 2016; p. 177. [Google Scholar]
- Dobosz, R.; Krawczyk, R. Comparison of the development ability of the northern root-knot nematode Meloidogyne hapla Chitwood, 1949 on selected species and varieties of Fabaceae Lindl. Prog. Plant Prot. 2017, 57, 159–163. [Google Scholar]
- Calvo, P.; Nelson, L.; Kloepper, J.W. Agricultural uses of plant biostimulants. Plant Soil 2014, 383, 3–41. [Google Scholar] [CrossRef] [Green Version]
- Fageria, N.K.; Filho, M.B.; Moreira, A.; Guimarães, C.M. Foliar fertilization of crop plants. J. Plant Nutr. 2009, 32, 1044–1064. [Google Scholar] [CrossRef]
- Kocira, S.; Szparaga, A.; Kocira, A.; Czerwińska, E.; Wójtowicz, A.; Bronowicka-Mielniczuk, U.; Koszel, M.; Findura, P. Modeling biometric traits, yield and nutritional and antioxidant properties of seeds of three soybean cultivars through the application of biostimulant containing seaweed and amino acids. Front. Plant. Sci. 2018, 9, 388. [Google Scholar] [CrossRef] [Green Version]
- Przybysz, A.; Gawronska, H.; Gajc-Wolska, J. Biological mode of action of a nitrophenolates-based biostimulant: Case study. Front. Plant Sci. 2014, 5, 713. [Google Scholar] [CrossRef]
- Du Jardin, P. Plant biostimulants: Definition, concept, main categories and regulation. Sci. Hortic. 2015, 196, 3–14. [Google Scholar] [CrossRef] [Green Version]
- Artyszak, A. Effect of silicon fertilization on crop yield quantity and quality—A literature review in Europe. Plants 2018, 7, 54. [Google Scholar] [CrossRef] [Green Version]
- Raj, A.B.; Raj, S.K. Zinc and boron nutrition in pulses: A review. J. App. Nat. Sci. 2019, 11, 673–679. [Google Scholar] [CrossRef]
- Fageria, N.K. The Use of Nutrients in Crop Plants; CRC Press: Boca Raton, FL, USA, 2009. [Google Scholar]
- Sulewska, H.; Ratajczak, K. Chemical composition of selected preparations supporting plant development and evaluation of their activity in soybean cultivation. Chem. Ind. 2017, 96, 1352–1355. [Google Scholar]
- Karaca, A.; Cetin, S.C.; Turgay, O.C.; Kizilkaya, R. Soil enzymes as indication of soil quality. In Soil Enzymology; Springer: Berlin/Heidelberg, Germany, 2010; pp. 119–148. [Google Scholar]
- Nannipieri, P.; Kandeler, E.; Ruggiero, P. Enzyme activities and microbiological and biochemical processes in soil. In Enzymes in the Environment. Activity, Ecology and Applications; Marcel Dekker Inc.: New York, NY, USA, 2002; pp. 1–33. [Google Scholar]
- Saviozzi, A.; Levi-Minzi, R.; Cardelli, R.; Riffaldi, R. A comparison of soil quality in adjacent cultivated, forest and native grassland soils. Plant Soil 2001, 233, 251–259. [Google Scholar] [CrossRef]
- IUSS Working Group WRB. World Reference Base for Soil Resources 2006, First Update 2007; World Soil Resources Reports No.103; FAO: Rome, Italy, 2007. [Google Scholar]
- Van Reeuwijk, L.P. Procedures for Soil Analysis, 6th ed.; ISRIC: Wageningen, The Netherlands, 2002. [Google Scholar]
- Krawczyk, R.; Mrówczyński, M. (Eds.) Metodyka Integrowanej Ochrony Łubinu Wąskolistnego, Żółtego i Białego dla Producentów; Instytut Ochrony Roślin-Państwowy Instytut Badawczy: Poznan, Poland, 2012. [Google Scholar]
- Walter, H. Vegetation Zones and Climate; PWRiL: Warsaw, Poland, 1976; pp. 26–31. [Google Scholar]
- Sawicka, A. The Ecological Aspects of Dinitrogen Fixation; Dissertations Scientific: 134; University of Agriculture in Poznan: Poznan, Poland, 1983. [Google Scholar]
- Kalaji, M.H.; Guo, P. Chlorophyll fluorescence: A useful tool in barley plant breeding programs. In Photochemistry Research Progress; Sánchez, A., Gutierrez, S.J., Eds.; Nova Science Publishers: Hauppauge, NY, USA, 2008; pp. 447–471. [Google Scholar]
- Thalmann, A. Zur Methodik der Bestimmung der Dehydrogenase aktivität im Boden mittels triphenytetrazoliumchlorid (TTC). Landwirtsch Forsch. 1968, 21, 249–258. [Google Scholar]
- Tabatabai, M.A.; Bremner, J.M. Use of p-nitrophenyl phosphate for assay of soil phosphatase activity. Soil Biol. Biochem. 1969, 1, 301–307. [Google Scholar] [CrossRef]
- Johnson, J.L.; Temple, K.L. Some variables affecting the measurement of “catalase activity” in soil 1. Soil Sci. Soc. Am. 1964, 28, 207–209. [Google Scholar] [CrossRef]
- Stefanic, F.; Ellade, G.; Chirnageanu, J. Researches concerning a biological index of soil fertility. In Proceeding of the 5th Symposium of Soil Biology; Nemes, M.P., Kiss, S., Papacostea, P., Stefanic, C., Rusan, M., Eds.; Romanian National Society of Soil Science: Bucharest, Romania, 1984; pp. 35–45. [Google Scholar]
- Abdi, H.; Lynne, J.W. Principal component analysis. Wiley Interdiscip. Rev. Comput. Stat. 2010, 2, 433–459. [Google Scholar] [CrossRef]
- Murtagh, F.; Legendre, P. Ward’s hierarchical clustering method: Clustering criterion and agglomerative algorithm. arXiv 2011, arXiv:1111.6285. [Google Scholar]
- Sacała, E. Role of silicon in plant resistance to water stress. J. Elementol. 2009, 14, 619–630. [Google Scholar] [CrossRef]
- Artyszak, A.; Gozdowski, D.; Kuchcińska, K. The effect of silicon foliar fertilization in sugar beet—Beta vulgaris (L.) ssp. vulgaris conv. crassa (Alef.) prov. altissima (Döll). Turk. J. Field Crop. 2015, 20, 115–119. [Google Scholar] [CrossRef]
- Fageria, N.K.; Baligar, V.C. Growth components and zinc recovery efficiency of upland rice genotypes. Pesqui. Agropecu. Bras. 2005, 40, 1211–1215. [Google Scholar] [CrossRef] [Green Version]
- Duffy, B. Zinc and plant disease. In Mineral Nutrition and Plant Disease; Datnoff, L.E., Elmer, W.H., Huber, D.M., Eds.; APS Press: St. Paul, MN, USA, 2007; pp. 155–175. [Google Scholar]
- Kaya, M.U.; Atak, M.E.; Khawar, K.M.; Ciftci, C.Y.; Ozcan, S. Effect of pre-sowing seed treatment with zinc and foliar spray of humic acids on yield of common bean (Phaseolus vulgaris L.). Int. J. Agric. Biol. 2005, 7, 875–878. [Google Scholar]
- Omer, F.A.; Abbas, D.N.; Khalaf, A.S. Effect of molybdenum and potassium application on nodulation, growth and yield of lentil (Lens culinaris MEDIC). Pak. J. Bot. 2016, 48, 2255–2259. [Google Scholar]
- Rahman, M.M.; Bhuiyan, M.M.H.; Sutradhar, G.N.C.; Rahman, M.M.; Paul, A.K. Effect of phosphorus, molybdenum and Rhizobium inoculation on yield and yield attributes of mung bean. Int. J. Sustain. Crop Prod. 2008, 3, 26–33. [Google Scholar]
- Hassanein, M.S.; Shalaby, M.A.F.; Rashad, E.M. Improving growth and yield of some faba bean cultivars by using some plant growth promoters in newly cultivated land. Ann. Agric. Sci. 2000, 38, 2141–2155. [Google Scholar]
- Tobbal, Y.F.M. Physiological Studies on the Effect of Some Nutrients and Growth Regulators on Plant Growth and Metabolism. Ph.D. Thesis, Faculty of Science Al-Azhar University, Cairo, Egypt, 2006. [Google Scholar]
- Artyszak, A.; Gozdowski, D.; Kucińska, K. Effect of foliar fertilization with silicon on the chosen physiological features and yield of sugar beet. Fragm. Agron. 2016, 33, 7–14. [Google Scholar]
- Guntzer, F.; Keller, C.; Meunier, J.D. Benefits of plant silicon for crops: A review. Agron. Sustain. Dev. 2012, 32, 201–213. [Google Scholar] [CrossRef] [Green Version]
- Marschner, H. Mineral Nutrition of Higher Plants, 2nd ed.; Academic Press: London, UK, 2002. [Google Scholar]
- Weisany, W.; Raei, Y.; Allahverdipoor, K.H. Role of some of mineral nutrients in biological nitrogen fixation. Bull. Environ. Pharm. Life Sci. 2013, 2, 77–84. [Google Scholar]
- Vieira, R.F.; Cardoso, E.J.B.N.; Vieira, C.; Cassini, S.T.A. Foliar application of molybdenum in common beans. I. Nitrogenase and reductase activities in a soil of high fertility. J. Plant Nutr. 1998, 21, 169–180. [Google Scholar] [CrossRef]
- Brown, P.H.; Bellaloui, N.; Wimmer, M.A.; Bassi, E.S.; Ruiz, J.; Hu, H.; Pfeffer, H.; Dannel, F.; Römheld, V. Boron in plant biology. Plant Biol. 2002, 4, 205–223. [Google Scholar] [CrossRef]
- Wani, P.A.; Khan, S.; Zaidi, A. Impact of zinc-tolerant plant growth-promoting rhizobacteria on lentil grown in zinc-amended soil. Agron. Sustain. Dev. 2008, 28, 449–455. [Google Scholar] [CrossRef]
- Bucher, M. Functional biology of plant phosphate uptake at root and mycorrhiza interfaces. New Phytol. 2007, 173, 11–26. [Google Scholar] [CrossRef] [PubMed]
- Bonilla, I.; Bolaños, L. Mineral nutrition for legume-rhizobia symbiosis: B, Ca, N, P, S, K, Fe, Mo, Co, and Ni. A review. Sustain. Agric. Rev. 2009, 1, 253–274. [Google Scholar]
- Abusuwar, A.O.; Omer, E.A. Effect of intercropping, phosphorus fertilization and rhizobium inoculation on the growth and nodulation of some leguminous and cereal forages. Agric. Biol. J. N. Am. 2011, 2, 109–124. [Google Scholar] [CrossRef]
- Zahran, H.H. Rhizobium-legume symbiosis and nitrogen fixation under severe conditions and in arid climate. Microbiol. Mol. Biol. Rev. 1999, 63, 968–989. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karaca, A.; Cetin, S.C.; Turgay, O.C.; Kizilkaya, R. Effects of heavy metals on soil enzyme activities. In Soil Heavy Metals; Springer: Berlin/Heidelberg, Germany, 2010; pp. 237–262. [Google Scholar]
- Das, S.K.; Varma, A. Role of enzymes in maintaining soil health. In Soil Enzymology, Soil Biology; Springer-Verlag: Berlin/Heidelberg, Germany, 2011; pp. 25–42. [Google Scholar]
- Bielińska, E.; Mocek-Płóciniak, A. Impact of the tillage system on the soil enzymatic activity. Arch Environ. Prot. 2012, 38, 75–82. [Google Scholar] [CrossRef]
- Panasiewicz, K.; Niewiadomska, A.; Sulewska, H.; Wolna-Maruwka, A.; Borowiak, K.; Budka, A.; Ratajczak, K. The effect of sewage sludge and BAF inoculant on plant condition and yield as well as biochemical and microbial activity of soil in willow (Salix viminalis L.) culture as an energy crop. Peer J. 2019, 7, e6434. [Google Scholar] [CrossRef] [Green Version]
- De-la-Peña, C.; Badri, D.V.; Lei, Z.; Watson, B.S.; Brăndao, M.M.; Silva-Filho, M.C.; Sumner, L.W.; Vivanco, J.M. Root secretion of defense-related proteins is development-dependent and correlated with flowering time. J. Biol. Chem. 2010, 285, 30654–30665. [Google Scholar] [CrossRef] [Green Version]
- Badri, D.V.; De-la-Peña, C.; Lei, Z.; Manter, D.K.; Chaparro, J.M.; Guimarães, R.L.; Sumner, L.W.; Vivanco, J.M. Root secreted metabolites and proteins are involved in the early events of plant-plant recognition prior to competition. PLoS ONE 2012, 7, e46640. [Google Scholar] [CrossRef] [Green Version]
- Niewiadomska, A.; Sulewska, H.; Wolna-Maruwka, A.; Ratajczak, K.; Głuchowska, K.; Waraczewska, Z.; Budka, A. An assessment of the influence of co-inoculation with endophytic bacteria and rhizobia, and the influence of PRP SOL and PRP EBV fertilizers on the microbial parameters of soil and nitrogenase activity in yellow lupine (Lupinus luteus L.) cultivation. Polish J. Environ. Stud. 2018, 27, 2687–2702. [Google Scholar] [CrossRef]
- Bielińska, E.J.; Futa, B.; Bik-Mołodzińska, M.; Szewczuk, C.; Sugier, D. The impact of fertilizing agents on the enzymatic activity of soils. J. Res. Appl. Agric. Eng. 2013, 58, 15–19. [Google Scholar]
- Swędrzyńska, D.; Zielewicz, W.; Przybył, P.; Starzyk, J. Effect of bio soil conditioner on microbial state and enzymatic activity of soil under Lolium perenne cultivation. Grassl. Sci. 2013, 16, 111–128. [Google Scholar]
- Bilen, S.; Bilen, M.; Bardhan, S. The effects of boron management on soil microbial population and enzyme activities. Afri. J. Biotech. 2011, 10, 5311–5319. [Google Scholar]
- Taran, N.Y.; Gonchar, O.M.; Lopatko, K.G.; Batsmanova, L.M.; Patyka, M.V.; Volkogon, M.V. The effect of colloidal solution of molybdenum nanoparticles on the microbial composition in rhizosphere of Cicer arietinum L. Nanoscale Res. Lett. 2014, 9, 289–297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ciereszko, I.; Szczygła, A.; Żebrowska, E. Phosphate deficiency affects acid phosphatase activity and growth of two wheat varieties. J. Plant Nutr. 2011, 34, 815–829. [Google Scholar] [CrossRef]
- Lemanowicz, J.; Siwik-Ziomek, A.; Koper, J. Effect of spring barley nitrogen fertilization on the changes in the content of phosphorus and the activity of alkaline and acid phosphatase in soil. Ecol. Chem. Eng. A 2012, 19, 1497–1507. [Google Scholar]
- Bielińska, E.; Mocek-Płóciniak, A. Phosphatases in the Soil Environment; UP Pub.: Poznan, Poland, 2009; ISBN 978-83-7160-554-3. [Google Scholar]
- Wang, J.B.; Chen, Z.H.; Chen, L.J.; Zhu, A.N.; Wu, Z.J. Surface soil phosphorus and phosphatase activities affected by tillage and crop residue input amounts. Plant Soil Environ. 2011, 57, 251–257. [Google Scholar] [CrossRef] [Green Version]
- Waldrip, H.M.; He, Z.; Erich, M.S. Effects of poultry manure amendment on phosphorus uptake by ryegrass, soil phosphorus fractions and phosphatase activity. Biol. Fert. Soils 2011, 47, 407–418. [Google Scholar] [CrossRef]
- Koter, M. Research on the impact of boron on the growth and development of plants. Soil Sci. Ann. 1963, 13, 185–211. [Google Scholar]
- Hu, X.J.; Zhu, Z.J. Effect of silicon on resistance of powdery mildew and the activities of antioxidative enzymes in leaf apoplast of cucumber. Acta Agric. Zhejiang 2008, 20, 67–71. [Google Scholar]
- Wyszkowska, J.; Borowik, A.; Kucharski, M.A.; Kucharski, J. Effect of cadmium, copper and zinc on plants, soil microorganisms and soil enzymes. J. Elementol. 2013, 18, 2587–2602. [Google Scholar] [CrossRef]
- Siwik-Ziomek, A.; Szczepanek, M. Soil extracellular enzyme activities and uptake of N by oilseed rape depending on fertilization and seaweed biostimulant application. Agron. J. 2019, 9, 480. [Google Scholar] [CrossRef] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bio-Stimulants/Foliar Fertilizers | Term and Dose of Bio-Stimulant | Fertilizer Characteristics | |
|---|---|---|---|
| Bio-stimulants | Tytanit | I: leaf and shoot development (BBCH 13–29)—0.3 dm3 ha−1 II: inflorescence development (BBCH 51–59)—0.3 dm3 ha−1 III: beginning of pod development (BBCH 71)—0.3 dm3 ha−1 | Liquid, mineral stimulant containing titanium (Ti). It increases the yield volume and development of plants, improves yield quality parameters and increases plants’ natural resistance to stress factors. Composition: 8.5 g Ti (dm3)−1 |
| Rooter | BBCH 13–14—1 dm3 ha−1 | Bio-stimulant—it stimulates the growth of the root system, accelerates regeneration and improves the uptake of soil minerals. Composition: P2O5 13.0%; K2O 5.0% | |
| Foliar fertilizers | Optysil | I: leaf and shoot development (BBCH 15–29)—0.5 dm3 ha−1 II: inflorescence development (BBCH 51–55)—0.5 dm3 ha−1 III: beginning of pod development (BBCH 71–73)—0.5 dm3 ha−1 | Liquid, silicon antistressor stimulating the growth and development of plants, activating their natural immune systems and increasing tolerance to unfavorable cultivation conditions. Composition: 200 g SiO2 (dm3)−1 |
| Metalosate Potassium | 2–3 treatments every 10–14 days during intensive growth—3 dm3 ha−1 | Liquid foliar fertilizer containing an easily absorbable form of potassium, which supplements potassium deficit in plants with amino acids. Composition: K2O 24% | |
| Bolero Mo | Before florescence—1.5 dm3 ha−1 | Liquid foliar fertilizer containing boron and molybdenum to supplement the deficit of these elements in plants. Composition: B 8.2%; Mo 0.8% | |
| ADOB 2.0 Zn IDHA | Before florescence—1 dm3 ha−1 | Foliar fertilizer containing zinc (Zn) fully chelated by biodegradable chelating agent IDHA. Composition: Zn 100 g kg−1 (weight percentage content 10, chelated by IDHA) | |
| ADOB B | I: before florescence—2 dm3 ha−1 II: after florescence on pods—1 dm3 ha−1 | Liquid, highly concentrated foliar fertilizer containing boron that regulates auxin activity and participates in cell division. Composition: N 78 g kg−1; B 150 g kg−1 | |
| ADOB 2.0 Mo | early stages of development—0.15 dm3 ha−1 | Liquid, single-component fertilizer which increases the rate and efficiency of use of nitrogen by plants and improves interaction with iron. Composition: Mo 20% | |
| Percentage of Soil Fractions | Texture Class | |||
|---|---|---|---|---|
| Fraction [mm] | Sand 2–0.05 | Silt 0.05–0.002 | Clay <0.002 | |
| 78 | 18 | 4 | LS | |
| Soil Chemical Properties | ||||
| pH in 1 mol KCl | 6.0 | |||
| Phosphorus P (mg·kg−1) | 70.1 | |||
| Potassium K (mg·kg−1) | 99.3 | |||
| Magnesium Mg (mg·kg−1) | 56.7 | |||
| Manganese Mn (mg·kg−1) | 303.4 | |||
| Zinc Zn (mg·kg−1) | 10.9 | |||
| Copper Cu (mg·kg−1) | 2.6 | |||
| Iron Fe (mg·kg−1) | 1525.2 | |||
| Boron B (mg·kg−1) | >20 | |||
| Organic carbon (%) | 0.5 | |||
| Percent of caries | 0.8 | |||
| Objects | Seed Yield, dt·ha−1 | TSW, g | Height, cm | Number of Pods, pc.·m−2 | Plant Dry Mass, g | Root Nodules Dry Mass, g | Fv/Fm BBCH 69 | CCI | LAI |
|---|---|---|---|---|---|---|---|---|---|
| 1 | 12.06 bc | 302.4 bc | 34.3 e | 221.8 bc | 5.05 | 0.145 bc | 0.784 cd | 33.5 d | 1.41 g |
| 2 | 11.82 bc | 295.2 c | 37.8 bcd | 300.8 a | 5.72 | 0.147 bc | 0.796 b | 46.7 b | 1.73 de |
| 3 | 13.88 a | 301.7 bc | 39.8 ab | 250.5 b | 6.46 | 0.160 bc | 0.792 bc | 23.1 f | 1.81 c |
| 4 | 11.96 bc | 305.8 bc | 40.5 a | 189.1 c | 5.28 | 0.170 b | 0.813 a | 24.2 f | 1.70 de |
| 5 | 13.18 ab | 313.9 ab | 37.1 cd | 235.0 bc | 5.36 | 0.142 bc | 0.776 d | 25.6 f | 2.03 a |
| 6 | 12.76 abc | 306.1 bc | 38.6 abc | 250.0 b | 5.14 | 0.129 c | 0.774 d | 30.2 e | 1.88 b |
| 7 | 13.63 a | 322.7 a | 36.4 d | 318.4 a | 6.17 | 0.212 a | 0.815 a | 50.9 a | 1.61 f |
| 8 | 11.67 c | 310.4 b | 37.9 bcd | 273.4 ab | 5.12 | 0.169 bc | 0.779 d | 42.9 c | 1.68 e |
| 9 | 11.94 bc | 309.0 b | 38.9 abc | 240.7 b | 5.24 | 0.170 bc | 0.797 b | 40.7 c | 1.76 cd |
| p-value | 0.001 | 0.000 | 0.000 | 0.000 | 0.236 | 0.001 | 0.000 | 0.000 | 0.000 |
| Parameter | Fertilization | Development Phase | Interaction |
|---|---|---|---|
| White Lupine Butan | |||
| Dehydrogenase | 13.393 ns | 159.989 *** | 41.123 ns |
| Alkaline phosphatase | 7.036 ns | 51.672 *** | 5.37 ns |
| Acid phosphatase | 14.907 ns | 116.200 *** | 10.116 ns |
| Catalase | 192.47 ns | 1558.42 *** | 121.42 ns |
| BIF | 2.90 ns | 131.96 *** | 2.71 ns |
| Nitrogenase | 14.08 *** | - | - |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Niewiadomska, A.; Sulewska, H.; Wolna-Maruwka, A.; Ratajczak, K.; Waraczewska, Z.; Budka, A. The Influence of Bio-Stimulants and Foliar Fertilizers on Yield, Plant Features, and the Level of Soil Biochemical Activity in White Lupine (Lupinus albus L.) Cultivation. Agronomy 2020, 10, 150. https://doi.org/10.3390/agronomy10010150
Niewiadomska A, Sulewska H, Wolna-Maruwka A, Ratajczak K, Waraczewska Z, Budka A. The Influence of Bio-Stimulants and Foliar Fertilizers on Yield, Plant Features, and the Level of Soil Biochemical Activity in White Lupine (Lupinus albus L.) Cultivation. Agronomy. 2020; 10(1):150. https://doi.org/10.3390/agronomy10010150
Chicago/Turabian StyleNiewiadomska, Alicja, Hanna Sulewska, Agnieszka Wolna-Maruwka, Karolina Ratajczak, Zyta Waraczewska, and Anna Budka. 2020. "The Influence of Bio-Stimulants and Foliar Fertilizers on Yield, Plant Features, and the Level of Soil Biochemical Activity in White Lupine (Lupinus albus L.) Cultivation" Agronomy 10, no. 1: 150. https://doi.org/10.3390/agronomy10010150
APA StyleNiewiadomska, A., Sulewska, H., Wolna-Maruwka, A., Ratajczak, K., Waraczewska, Z., & Budka, A. (2020). The Influence of Bio-Stimulants and Foliar Fertilizers on Yield, Plant Features, and the Level of Soil Biochemical Activity in White Lupine (Lupinus albus L.) Cultivation. Agronomy, 10(1), 150. https://doi.org/10.3390/agronomy10010150