Investigation of Heat-Induced Changes in the Grain Yield and Grains Metabolites, with Molecular Insights on the Candidate Genes in Barley

 ,
,  , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experiment Layout and Planting Conditions
2.2. Meteorological Data
2.3. Yield Attributes and Physiological Traits
2.3.1. The Yield-Related Traits
2.3.2. The Physiological Traits
Chlorophyll Content (CCF)
Leaf Water Content (WC)
Grain Reserved Metabolites
Physiological Changes Due to Heat Stress
2.4. Principal Component Analysis (PCA)
2.5. Statistical Analysis
2.6. DNA Extraction and Genotyping-By-Sequencing (GBS)
2.7. Candidate Gene Identification
3. Results
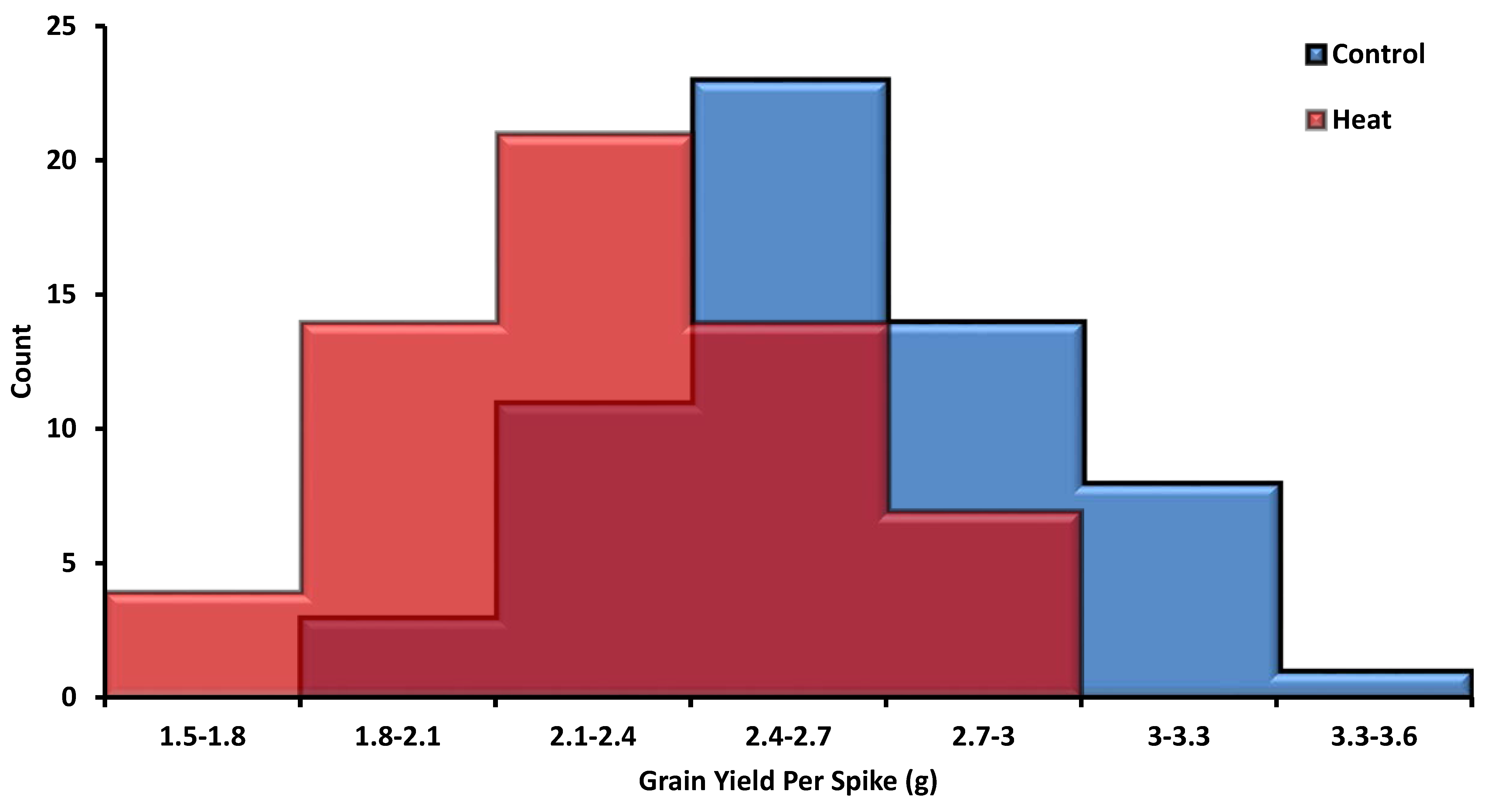
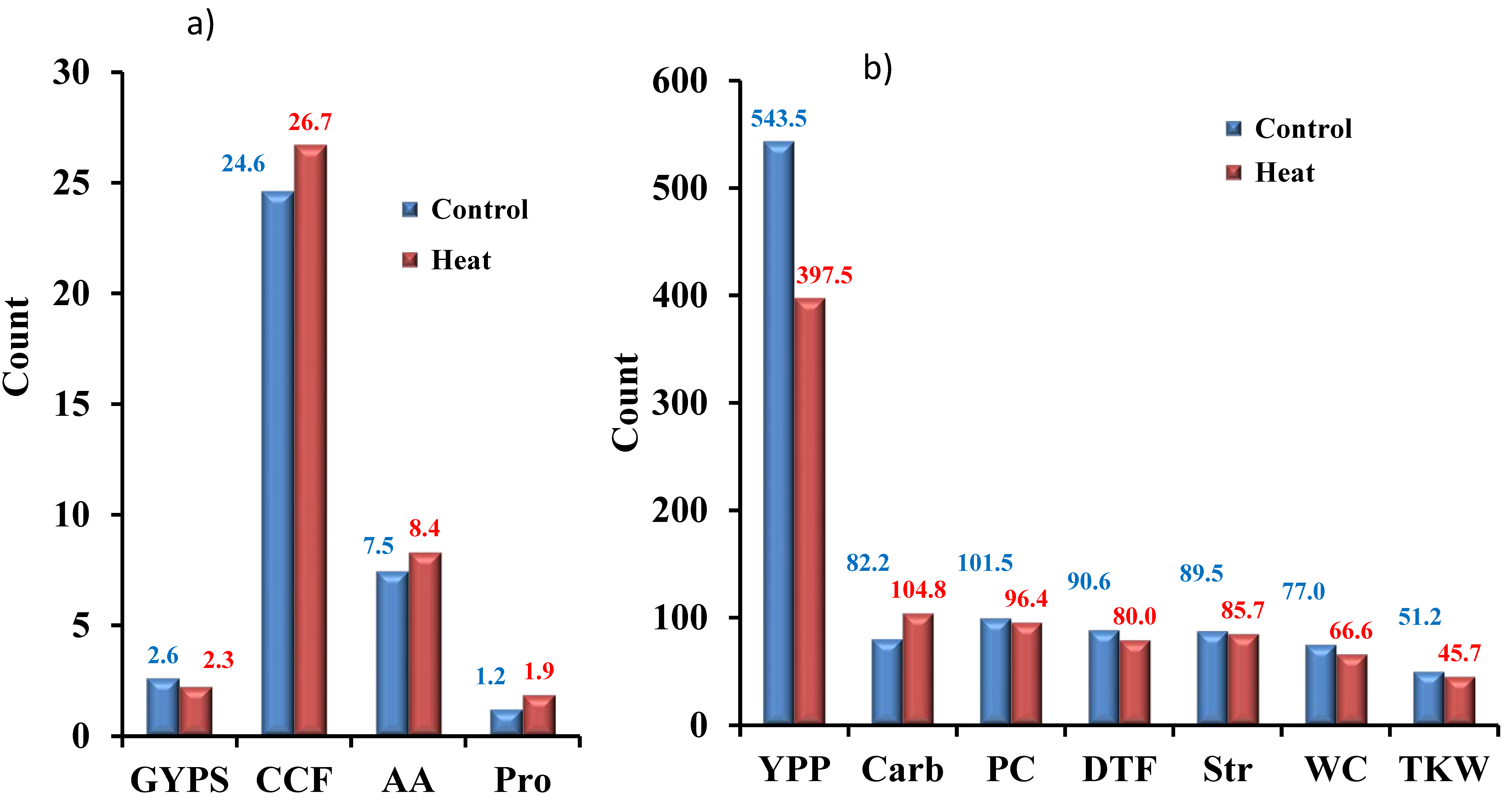
3.1. Yield-Related Traits
3.2. Physiological Traits
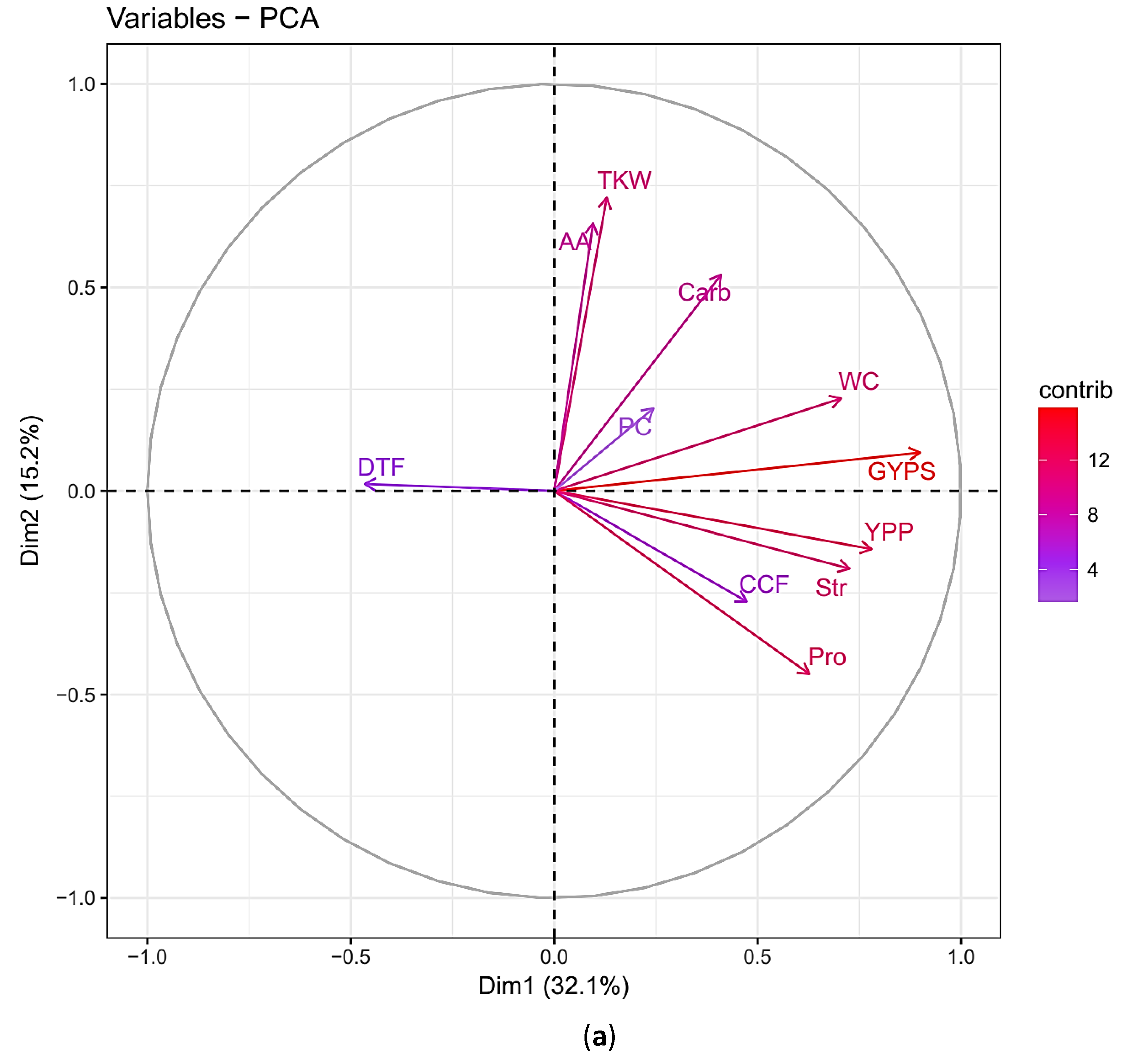
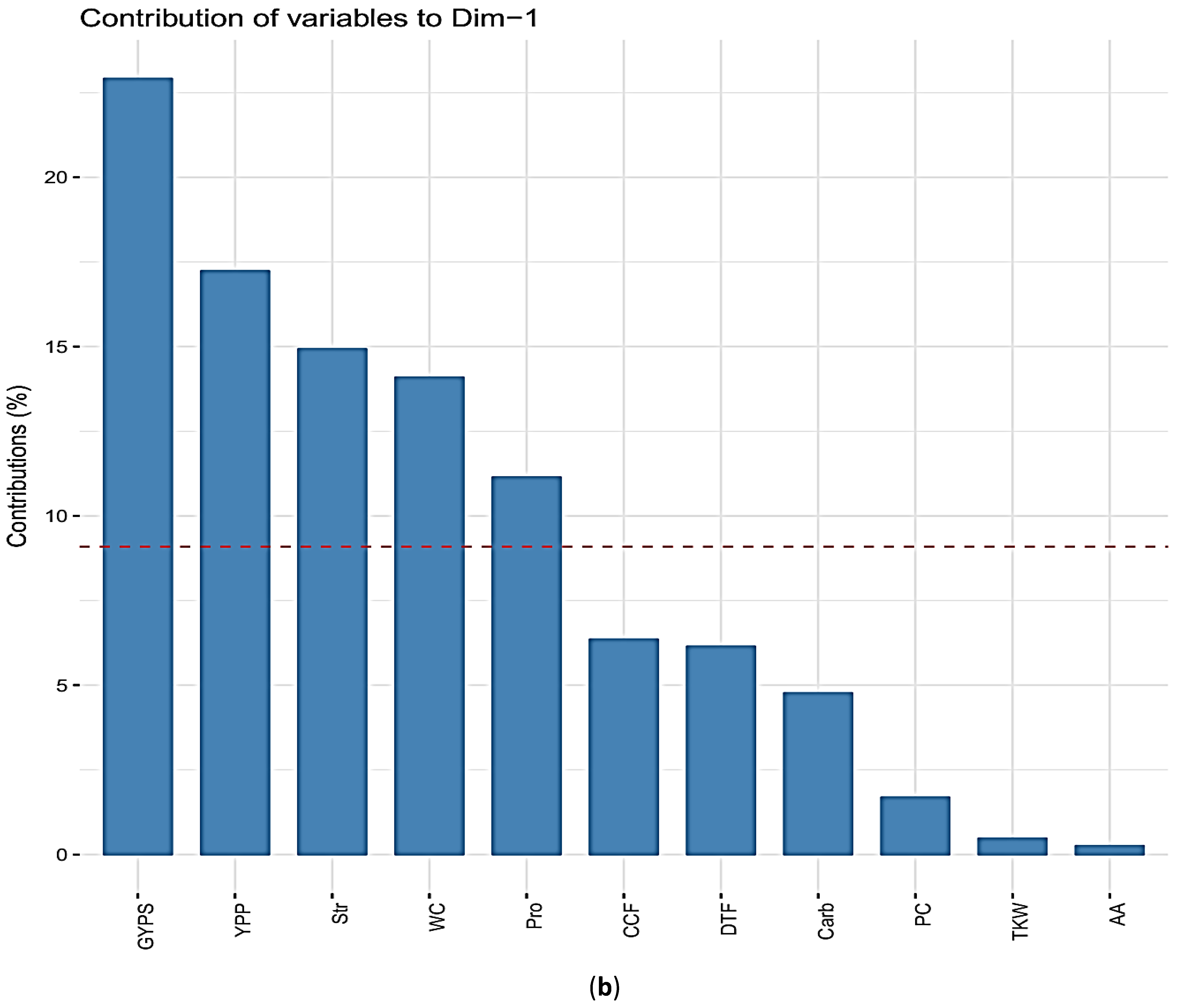
3.3. The Principal Component Analysis (PCA)
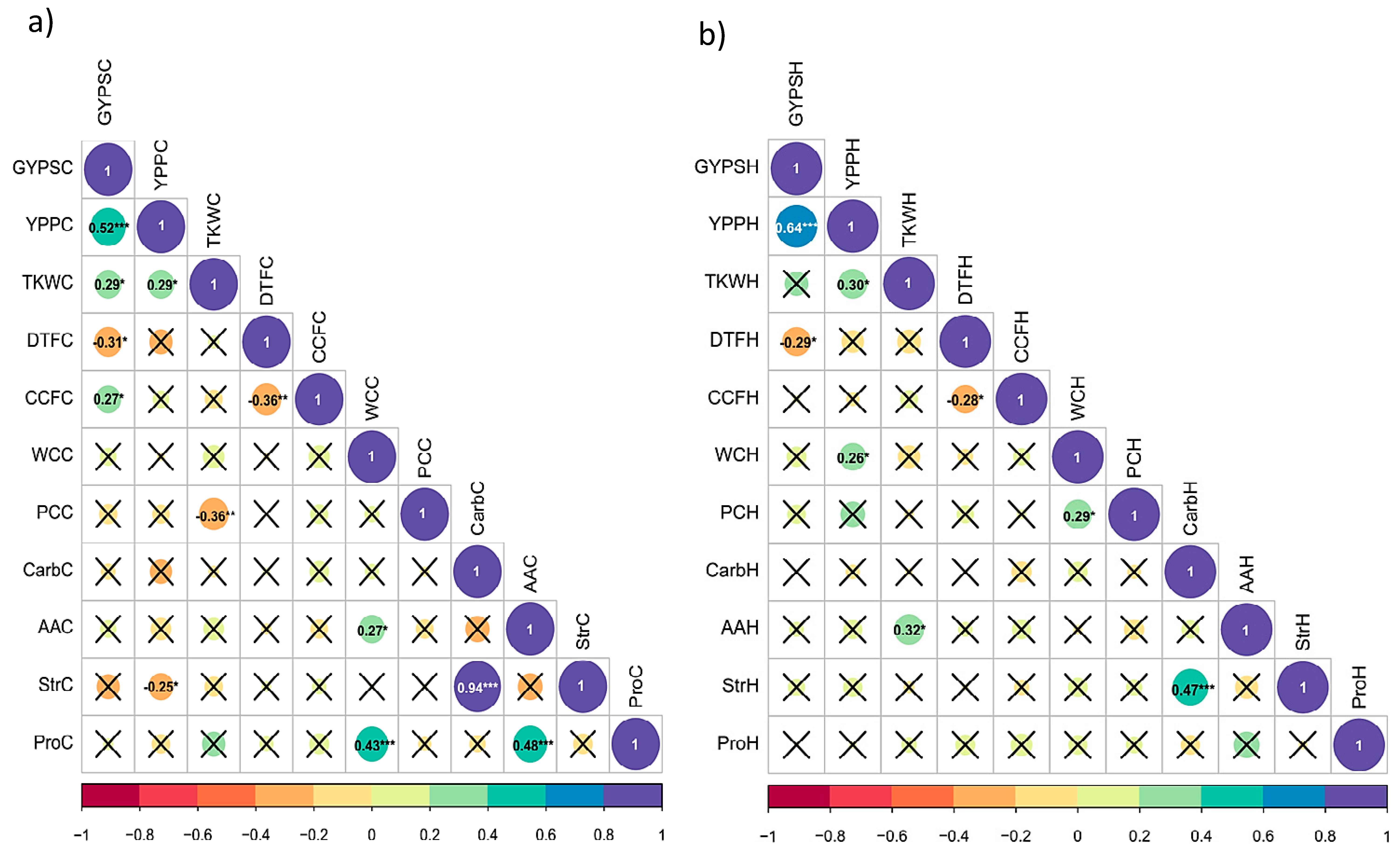
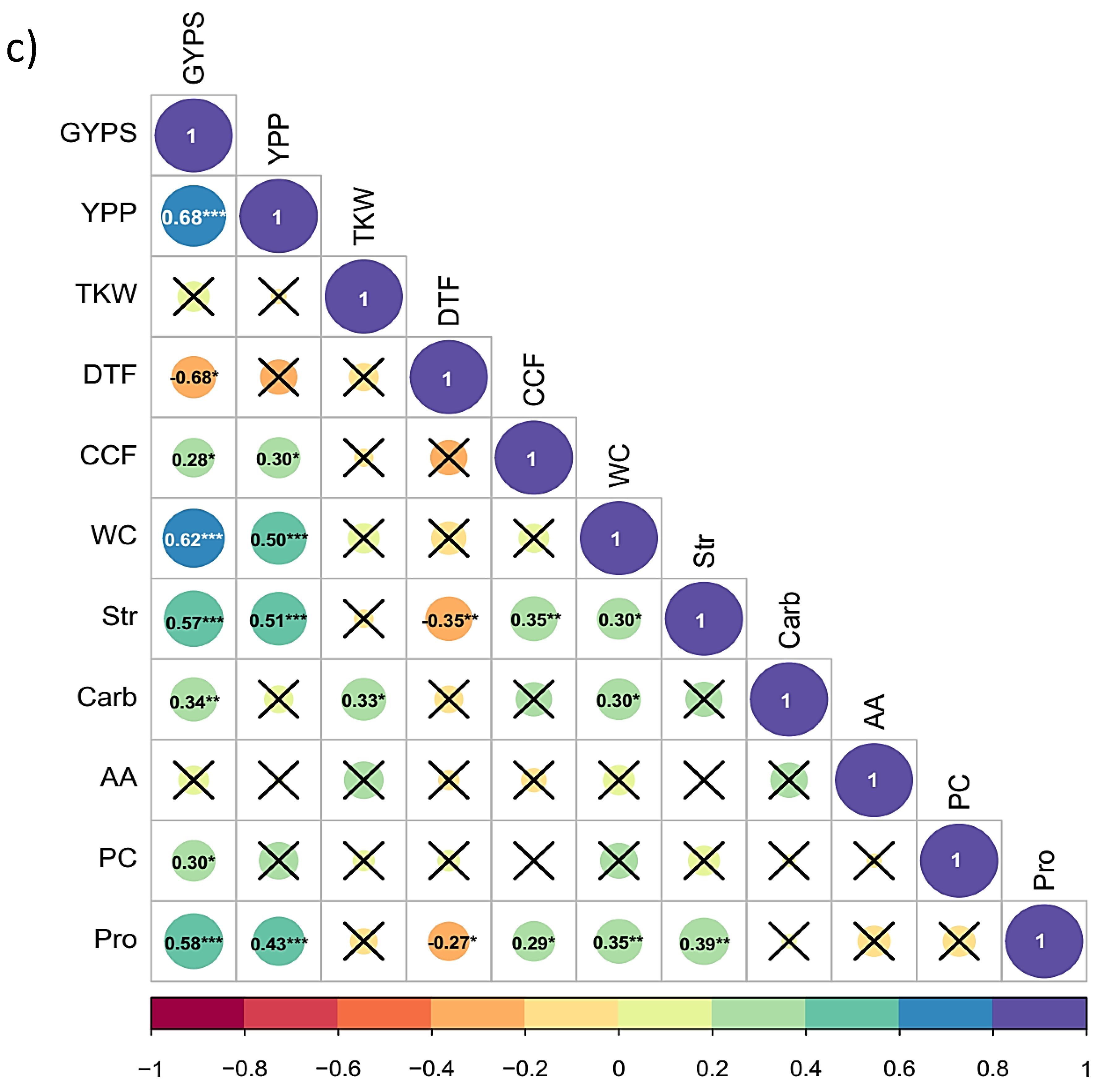
3.4. Correlation Analysis
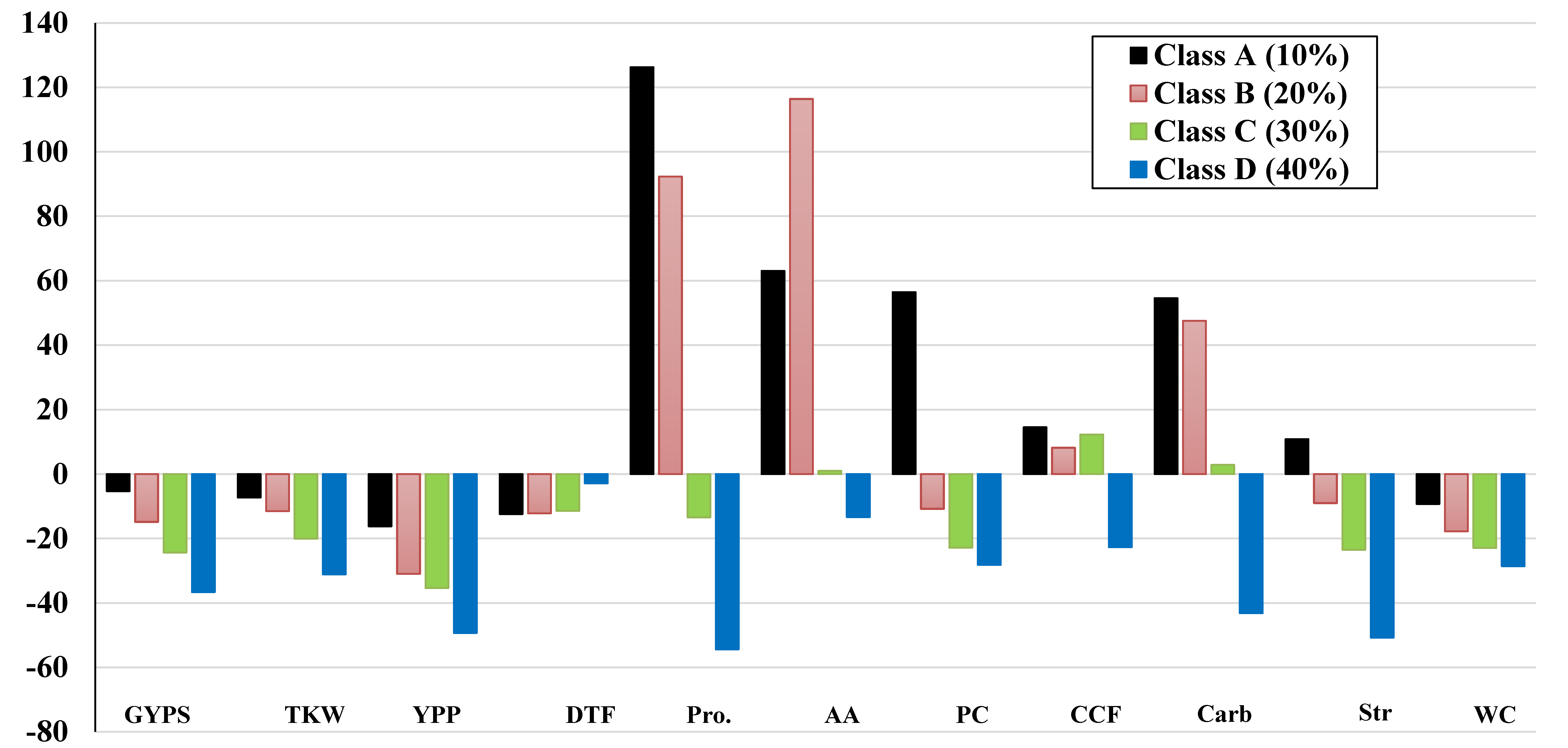
3.5. Percentage Changes and Classification of the Genotypes into Four Classes
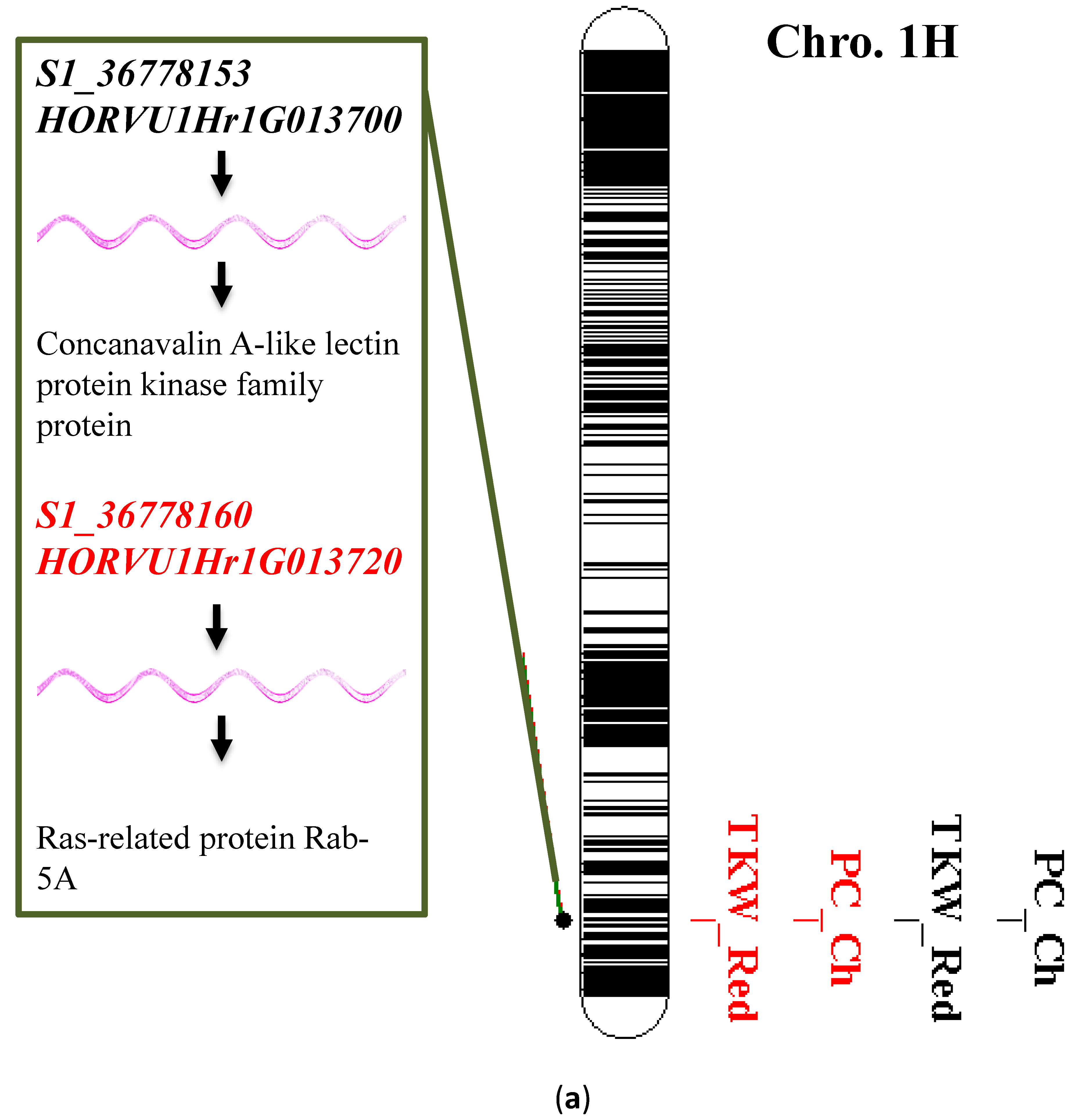
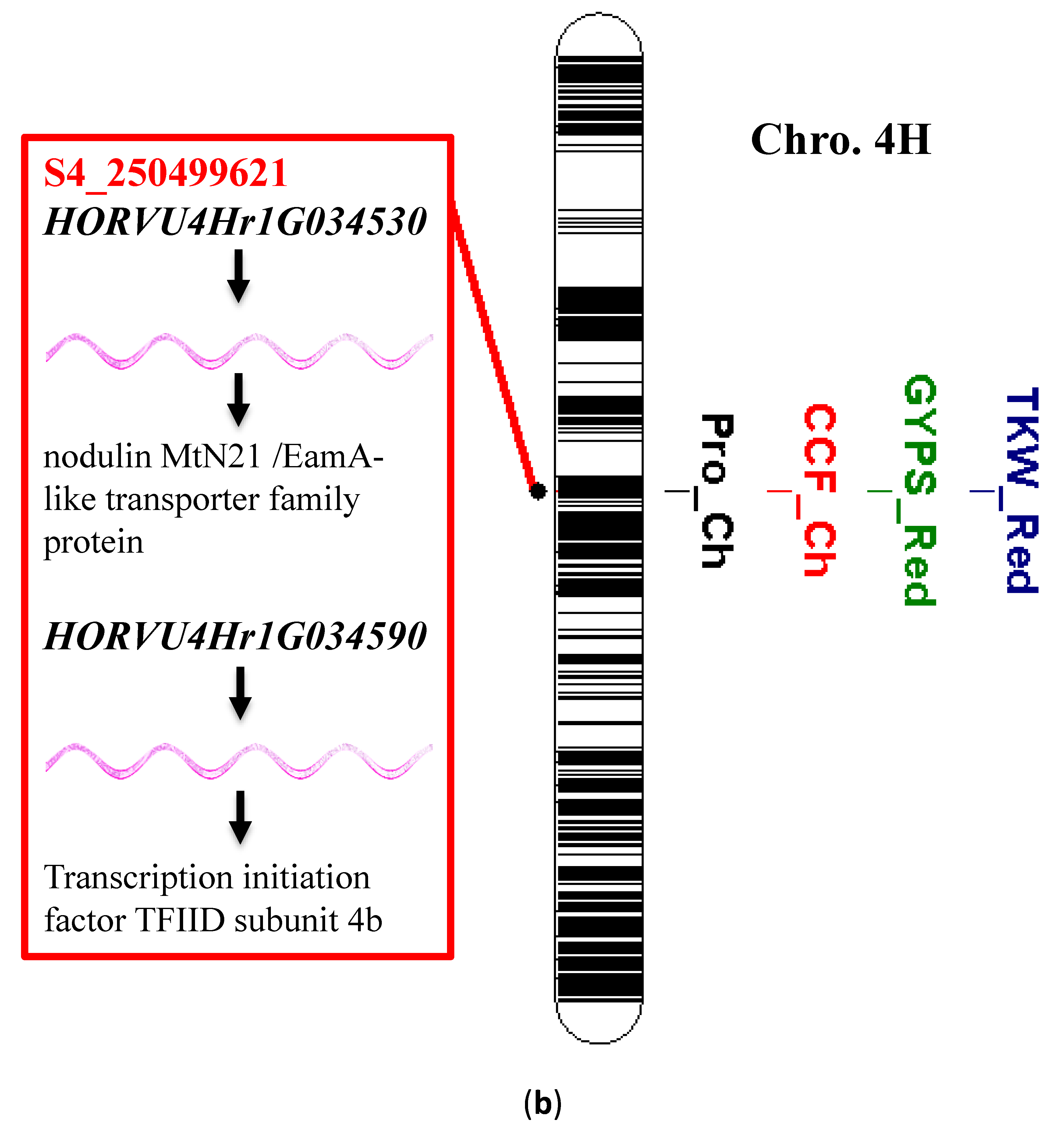
3.6. Single Marker Analysis and Gene Annotation
4. Discussion
4.1. Yield-Related and Physiological Traits Variations
4.2. Single Marker Analysis and Candidate Genes
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hasanuzzaman, M.; Nahar, K.; Alam, M.M.; Roychowdhury, R.; Fujita, M. Physiological, biochemical, and molecular mechanisms of heat stress tolerance in plants. Int. J. Mol. Sci. 2013, 14, 9643–9684. [Google Scholar] [CrossRef] [PubMed]
- Lobell, D.B.; Schlenker, W.; Costa-Roberts, J. Climate trends and global crop production since 1980. Science 2011, 333, 616–620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ortiz, R.; Braun, H.J.; Crossa, J.; Crouch, J.H.; Davenport, G.; Dixon, J.; Dreisigacker, S.; Duveiller, E.; He, Z.; Huerta, J.; et al. Wheat genetic resources enhancement by the International Maize and Wheat Improvement Center (CIMMYT). Genet. Resour. Crop Evol. 2008, 55, 1095–1140. [Google Scholar] [CrossRef]
- Weichert, H.; Högy, P.; Mora-Ramirez, I.; Fuchs, J.; Eggert, K.; Koehler, P.; Weschke, W.; Fangmeier, A.; Weber, H. Grain yield and quality responses of wheat expressing a barley sucrose transporter to combined climate change factors. J. Exp. Bot. 2017, 68, 5511–5525. [Google Scholar] [CrossRef] [Green Version]
- Christensen, J.H.; Christensen, O.B. A summary of the PRUDENCE model projections of changes in European climate by the end of this century. Springer 2007, 81, 7–30. [Google Scholar] [CrossRef]
- Fang, S.; Cammarano, D.; Zhou, G.; Tan, K.; Ren, S. Effects of increased day and night temperature with supplemental infrared heating on winter wheat growth in North China. Eur. J. Agron. 2015, 64, 67–77. [Google Scholar] [CrossRef]
- Fahad, S.; Hussain, S.; Saud, S.; Khan, F.; Hassan, S.; Amanullah Nasim, W.; Arif, M.; Wang, F.; Huang, J. Exogenously Applied Plant Growth Regulators Affect Heat-Stressed Rice Pollens. J. Agron. Crop Sci. 2016, 202, 139–150. [Google Scholar] [CrossRef]
- Wahid, A.; Gelani, S.; Ashraf, M.; Foolad, M.R. Heat tolerance in plants: An overview. Environ. Exp. Bot. 2007, 61, 199–223. [Google Scholar] [CrossRef]
- Bita, C.E.; Gerats, T. Plant tolerance to high temperature in a changing environment: Scientific fundamentals and production of heat stress-tolerant crops. Front. Plant Sci. 2013, 4, 273. [Google Scholar] [CrossRef] [Green Version]
- Sekhon, H.S.; Singh, G.; Sharma, P.; Bains, T.S. Water Use Efficiency Under Stress Environments. In Climate Change and Management of Cool Season Grain Legume Crops; Yadav, S.S., Redden, R., Eds.; Springer: Dordrecht, The Netherlands, 2010; pp. 207–227. ISBN 978-90-481-3709-1. [Google Scholar]
- Li, H.; Wang, X.M.; Chen, L.; Ahammed, G.J.; Xia, X.J.; Shi, K.; Considine, M.J.; Yu, J.Q.; Zhou, Y.H. Growth temperature-induced changes in biomass accumulation, photosynthesis and glutathione redox homeostasis as influenced by hydrogen peroxide in cucumber. Plant Physiol. Biochem. 2013, 71, 1–10. [Google Scholar] [CrossRef]
- Mathur, S.; Agrawal, D.; Jajoo, A. Photosynthesis: Response to high temperature stress. J. Photochem. Photobiol. B Biol. 2014, 137, 116–126. [Google Scholar] [CrossRef]
- Howarth, C.J. Genetic improvements of tolerance to high temperature. In Abiotic Stresses: Plant Resistance through Breeding and Molecular Approaches; Ashraf, M., Harris, P.J.C., Eds.; Haworth Press Inc.: New York, NY, USA, 2005; pp. 277–300. [Google Scholar]
- Wilhelm, E.P.; Mullen, R.E.; Keeling, P.L.; Singletary, G.W. Heat Stress during Grain Filling in Maize: Effects on Kernel Growth and Metabolism. Crop Sci. 1999, 39, 1733–1741. [Google Scholar] [CrossRef]
- Maestri, E.; Klueva, N.; Perrotta, C.; Gulli, M.; Nguyen, H.T.; Marmiroli, N. Molecular genetics of heat tolerance and heat shock proteins in cereals. Plant Mol. Biol. 2002, 48, 667–681. [Google Scholar] [CrossRef]
- Kishor, P.K.; Sangam, S.; Amrutha, R.N.; Laxmi, P.S.; Naidu, K.R.; Rao, K.S.; Sreenivasulu, N. Regulation of proline biosynthesis, degradation, uptake and transport in higher plants: Its implications in plant growth and abiotic stress tolerance. Curr. Sci. 2005, 88, 424–438. [Google Scholar] [CrossRef]
- Zhang, J.; Li, D.M.; Gao, Y.; Yu, B.; Xia, C.X.; Bai, J.G. Pretreatment with 5-aminolevulinic acid mitigates heat stress of cucumber leaves. Biol. Plant. 2012, 56, 780–784. [Google Scholar] [CrossRef]
- Ehlers, J.D.; Hall, A.E. Heat tolerance of contrasting cowpea lines in short and long days. Field Crop. Res. 1998, 55, 11–21. [Google Scholar] [CrossRef]
- Gous, P.W.; Hickey, L.; Christopher, J.T.; Franckowiak, J.; Fox, G.P. Discovery of QTL for stay-green and heat-stress in barley (Hordeum vulgare) grown under simulated abiotic stress conditions. Euphytica 2016, 207, 305–317. [Google Scholar] [CrossRef]
- Mangelsen, E.; Kilian, J.; Harter, K.; Jansson, C.; Wanke, D.; Sundberg, E. Transcriptome analysis of high-temperature stress in developing barley caryopses: Early stress responses and effects on storage compound biosynthesis. Mol. Plant 2011, 4, 97–115. [Google Scholar] [CrossRef]
- Templer, S.E.; Ammon, A.; Pscheidt, D.; Ciobotea, O.; Schuy, C.; McCollum, C.; Sonnewald, U.; Hanemann, A.; Förster, J.; Ordon, F.; et al. Metabolite profiling of barley flag leaves under drought and combined heat and drought stress reveals metabolic QTLs for metabolites associated with antioxidant defense. J. Exp. Bot. 2017, 68, 1697–1713. [Google Scholar] [CrossRef] [Green Version]
- Elakhdar, A.; EL-Sattar, M.A.; Amer, K.; Rady, A.; Kumamaru, T. Population structure and marker–trait association of salt tolerance in barley (Hordeum vulgare L.). Comptes Rendus Biol. 2016, 339, 454–461. [Google Scholar] [CrossRef]
- Sallam, A.; Amro, A.; EL-Akhdar, A.; Dawood, M.F.A.; Kumamaru, T.; Stephen Baenziger, P. Genetic diversity and genetic variation in morpho-physiological traits to improve heat tolerance in Spring barley. Mol. Biol. Rep. 2018, 45, 2441–2453. [Google Scholar] [CrossRef]
- LOWRY, O.H.; ROSEBROUGH, N.J.; FARR, A.L.; RANDALL, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef] [PubMed]
- MOORE, S.; STEIN, W.H. Photometric ninhydrin method for use in the chromatography of amino acids. J. Biol. Chem. 1948, 176, 367–388. [Google Scholar]
- FALES, F.W. The assimilation and degradation of carbohydrates by yeast cells. J. Biol. Chem. 1951, 193, 113–124. [Google Scholar] [CrossRef] [PubMed]
- Schlegel, H.-G. Die Verwertung organischer Säuren durch Chlorella im Licht. Planta 1956, 47, 510–526. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Julkowska, M.M.; Saade, S.; Agarwal, G.; Gao, G.; Pailles, Y.; Morton, M.; Awlia, M.; Tester, M. MV app-multivariate analysis application for streamlined data analysis and curation. Plant Physiol. 2019, 180, 1261–1276. [Google Scholar] [CrossRef] [Green Version]
- Utz, H. PLABSTAT: A Computer Program for the Statistical Analysis of Plant Breeding Experiments; Institute of Plant Breeding, Seed Science and Population Genetics: Stuttgart, Germany, 2011. [Google Scholar]
- 3.5.1., R.D.C.T. A Language and Environment for Statistical Computing. R Found. Stat. Comput. 2018, 2. Available online: https://www.R-project.org (accessed on 5 November 2020).
- Poland, J.A.; Rife, T.W. Genotyping-by-Sequencing for Plant Breeding and Genetics. Plant Genome 2012, 5, 92–102. [Google Scholar] [CrossRef] [Green Version]
- Bradbury, P.J.; Zhang, Z.; Kroon, D.E.; Casstevens, T.M.; Ramdoss, Y.; Buckler, E.S. TASSEL: Software for association mapping of complex traits in diverse samples. Bioinformatics 2007, 23, 2633–2635. [Google Scholar] [CrossRef]
- Francis, D.; Merk, H.; Namuth-Covert, D. Introduction to Single Marker Analysis (SMA). 3–5. 2011. Available online: https://plant-breeding-genomics.extension.org/introduction-to-single-marker-analysis-sma/ (accessed on 30 August 2020).
- Mascher, M.; Gundlach, H.; Himmelbach, A.; Beier, S.; Twardziok, S.O.; Wicker, T.; Radchuk, V.; Dockter, C.; Hedley, P.E.; Russell, J.; et al. A chromosome conformation capture ordered sequence of the barley genome. Nature 2017, 544, 427–433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bayer, M.M.; Rapazote-Flores, P.; Ganal, M.; Hedley, P.E.; Macaulay, M.; Plieske, J.; Ramsay, L.; Russell, J.; Shaw, P.D.; Thomas, W.; et al. Development and evaluation of a barley 50k iSelect SNP array. Front. Plant Sci. 2017, 8. [Google Scholar] [CrossRef] [Green Version]
- Kinsella, R.J.; Kähäri, A.; Haider, S.; Zamora, J.; Proctor, G.; Spudich, G.; Almeida-King, J.; Staines, D.; Derwent, P.; Kerhornou, A.; et al. Ensembl BioMarts: A hub for data retrieval across taxonomic space. Database 2011, 2011. [Google Scholar] [CrossRef]
- Pathak, S.; Poudyal, C.; Ojha, B.R.; Marahatta, S. Evaluation of the effects of terminal heat stress on grain traits of barley (Hordeum vulgare L.) IN CHITWAN. Int. J. Agric. Environ. Res. 2017, 3, 2856–2869. [Google Scholar]
- Sita, K.; Sehgal, A.; Bhandari, K.; Kumar, J.; Kumar, S.; Singh, S.; Siddique, K.H.M.; Nayyar, H. Impact of heat stress during seed filling on seed quality and seed yield in lentil (Lens culinaris Medikus) genotypes. J. Sci. Food Agric. 2018, 98, 5134–5141. [Google Scholar] [CrossRef]
- Sinsawat, V.; Leipner, J.; Stamp, P.; Fracheboud, Y. Effect of heat stress on the photosynthetic apparatus in maize (Zea mays L.) grown at control or high temperature. Environ. Exp. Bot. 2004, 52, 123–129. [Google Scholar] [CrossRef]
- Camejo, D.; Rodríguez, P.; Morales, M.A.; Dell’Amico, J.M.; Torrecillas, A.; Alarcón, J.J. High temperature effects on photosynthetic activity of two tomato cultivars with different heat susceptibility. J. Plant Physiol. 2005, 162, 281–289. [Google Scholar] [CrossRef]
- Fahad, S.; Bajwa, A.A.; Nazir, U.; Anjum, S.A.; Farooq, A.; Zohaib, A.; Sadia, S.; Nasim, W.; Adkins, S.; Saud, S.; et al. Crop production under drought and heat stress: Plant responses and management options. Front. Plant Sci. 2017, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zinn, K.E.; Tunc-Ozdemir, M.; Harper, J.F. Temperature stress and plant sexual reproduction: Uncovering the weakest links. J. Exp. Bot. 2010, 61, 1959–1968. [Google Scholar] [CrossRef] [Green Version]
- Rollins, J.A.; Habte, E.; Templer, S.E.; Colby, T.; Schmidt, J.; Von Korff, M. Leaf proteome alterations in the context of physiological and morphological responses to drought and heat stress in barley (Hordeum vulgare L.). J. Exp. Bot. 2013, 64, 3201–3212. [Google Scholar] [CrossRef] [Green Version]
- Makonya, G.M.; Ogola, J.B.O.; Muthama Muasya, A.; Crespo, O.; Maseko, S.; Valentine, A.J.; Ottosen, C.O.; Rosenqvist, E.; Chimphango, S.B.M. Chlorophyll fluorescence and carbohydrate concentration as field selection traits for heat tolerant chickpea genotypes. Plant Physiol. Biochem. 2019, 141, 172–182. [Google Scholar] [CrossRef]
- Tewolde, H.; Fernandez, C.J.; Erickson, C.A. Wheat cultivars adapted to post-heading high temperature stress. J. Agron. Crop Sci. 2006, 192, 111–120. [Google Scholar] [CrossRef]
- Vadez, V.; Kholova, J.; Choudhary, S.; Zindy, P.; Terrier, M.; Krishnamurthy, L.; Kumar, P.R.; Turner, N.C. Responses to Increased Moisture Stress and Extremes: Whole Plant Response to Drought under Climate Change. In Crop Adaptation to Climate Change; Yadav, S.S., Redden, R., Hatfield, J.L., Lotze-Campen, H., Hall, A.E., Eds.; Wiley/Blackwell: Chichester, UK, 2011; pp. 186–197. ISBN 9780813820163. [Google Scholar]
- Lin, W.; Guo, X.; Pan, X.; Li, Z. Chlorophyll composition, chlorophyll fluorescence, and grain yield change in esl mutant rice. Int. J. Mol. Sci. 2018, 19, 2945. [Google Scholar] [CrossRef] [Green Version]
- Sharkey, T.D.; Zhang, R. High Temperature Effects on Electron and Proton Circuits of Photosynthesis. J. Integr. Plant Biol. 2010, 52, 712–722. [Google Scholar] [CrossRef] [PubMed]
- Mirzaei, M.; Pascovici, D.; Atwell, B.J.; Haynes, P.A. Differential regulation of aquaporins, small GTPases and V-ATPases proteins in rice leaves subjected to drought stress and recovery. Proteomics 2012, 12, 864–877. [Google Scholar] [CrossRef]
- Hussain, H.A.; Men, S.; Hussain, S.; Chen, Y.; Ali, S.; Zhang, S.; Zhang, K.; Li, Y.; Xu, Q.; Liao, C.; et al. Interactive effects of drought and heat stresses on morpho-physiological attributes, yield, nutrient uptake and oxidative status in maize hybrids. Sci. Rep. 2019, 9. [Google Scholar] [CrossRef] [Green Version]
- Triboï, E.; Martre, P.; Triboï-Blondel, A.M. Environmentally-induced changes in protein composition in developing grains of wheat are related to changes in total protein content. J. Exp. Bot. 2003, 54, 1731–1742. [Google Scholar] [CrossRef] [PubMed]
- Nagarajan, S.; Jagadish, S.V.K.; Prasad, A.S.H.; Thomar, A.K.; Anand, A.; Pal, M.; Agarwal, P.K. Local climate affects growth, yield and grain quality of aromatic and non-aromatic rice in northwestern India. Agric. Ecosyst. Environ. 2010, 138, 274–281. [Google Scholar] [CrossRef]
- Dupont, F.M.; Hurkman, W.J.; Vensel, W.H.; Tanaka, C.; Kothari, K.M.; Chung, O.K.; Altenbach, S.B. Protein accumulation and composition in wheat grains: Effects of mineral nutrients and high temperature. Eur. J. Agron. 2006, 25, 96–107. [Google Scholar] [CrossRef]
- Ahmed, M.; Hassan, F.U.; Qadir, G.; Shaheen, F.A.; Aslam, M.A. Response of proline accumulation in bread wheat (Triticum aestivum L.) under rainfed conditions. J. Agric. Meteorol. 2017, 73, 147–155. [Google Scholar] [CrossRef] [Green Version]
- Khan, A.; Khan, A.L.; Imran, M.; Asaf, S.; Kim, Y.H.; Bilal, S.; Numan, M.; Al-Harrasi, A.; Al-Rawahi, A.; Lee, I.J. Silicon-induced thermotolerance in Solanum lycopersicum L. via activation of antioxidant system, heat shock proteins, and endogenous phytohormones. BMC Plant Biol. 2020, 20. [Google Scholar] [CrossRef]
- Sattar, A.; Sher, A.; Ijaz, M.; Ul-Allah, S.; Rizwan, M.S.; Hussain, M.; Jabran, K.; Cheema, M.A. Terminal drought and heat stress alter physiological and biochemical attributes in flag leaf of bread wheat. PLoS ONE 2020, 15. [Google Scholar] [CrossRef] [PubMed]
- Zargar, S.M.; Nagar, P.; Deshmukh, R.; Nazir, M.; Wani, A.A.; Masoodi, K.Z.; Agrawal, G.K.; Rakwal, R. Aquaporins as potential drought tolerance inducing proteins: Towards instigating stress tolerance. J. Proteom. 2017, 169, 233–238. [Google Scholar] [CrossRef]
- Larkindale, J.; Knight, M.R. Protection against heat stress-induced oxidative damage in Arabidopsis involves calcium, abscisic acid, ethylene, and salicylic acid. Plant Physiol. 2002, 128, 682–695. [Google Scholar] [CrossRef]
- Garg, A.K.; Kim, J.K.; Owens, T.G.; Ranwala, A.P.; Do Choi, Y.; Kochian, L.V.; Wu, R.J. Trehalose accumulation in rice plants confers high tolerance levels to different abiotic stresses. Proc. Natl. Acad. Sci. USA 2002, 99, 15898–15903. [Google Scholar] [CrossRef] [Green Version]
- Gómez, L.D.; Baud, S.; Gilday, A.; Li, Y.; Graham, I.A. Delayed embryo development in the ARABIDOPSIS TREHALOSE-6-PHOSPHATE SYNTHASE 1 mutant is associated with altered cell wall structure, decreased cell division and starch accumulation. Plant J. 2006, 46, 69–84. [Google Scholar] [CrossRef]
- Lyu, J.I.; Park, J.H.; Kim, J.K.; Bae, C.H.; Jeong, W.J.; Min, S.R.; Liu, J.R. Enhanced tolerance to heat stress in transgenic tomato seeds and seedlings overexpressing a trehalose-6-phosphate synthase/phosphatase fusion gene. Plant Biotechnol. Rep. 2018, 12, 399–408. [Google Scholar] [CrossRef]
- Nuccio, M.L.; Wu, J.; Mowers, R.; Zhou, H.P.; Meghji, M.; Primavesi, L.F.; Paul, M.J.; Chen, X.; Gao, Y.; Haque, E.; et al. Expression of trehalose-6-phosphate phosphatase in maize ears improves yield in well-watered and drought conditions. Nat. Biotechnol. 2015, 33, 862–869. [Google Scholar] [CrossRef]
- Killiny, N.; Hijaz, F.; Nehela, Y.; Hajeri, S.; Gowda, S. Effects of δ-aminolevulinic acid dehydratase silencing on the primary and secondary metabolisms of citrus. Plant Direct 2018, 2. [Google Scholar] [CrossRef] [Green Version]
- Han, M.J.; Jung, K.H.; Yi, G.; An, G. Rice Importin β1 gene affects pollen tube elongation. Mol. Cells 2011, 31, 523–530. [Google Scholar] [CrossRef] [Green Version]
- Sato, S.; Kamiyama, M.; Iwata, T.; Makita, N.; Furukawa, H.; Ikeda, H. Moderate increase of mean daily temperature adversely affects fruit set of Lycopersicon esculentum by disrupting specific physiological processes in male reproductive development. Ann. Bot. 2006, 97, 731–738. [Google Scholar] [CrossRef]
- Li, L.; Zheng, W.; Zhu, Y.; Ye, H.; Tang, B.; Arendseea, Z.W.; Jones, D.; Li, R.; Ortiz, D.; Zhao, X.; et al. QQS orphan gene regulates carbon and nitrogen partitioning across species via NF-YC interactions. Proc. Natl. Acad. Sci. USA 2015, 112, 14734–14739. [Google Scholar] [CrossRef] [Green Version]
- Xu, Z.; Dooner, H.K. The maize aberrant pollen transmission 1 gene is a SABRE/KIP homolog required for pollen tube growth. Genetics 2006, 172, 1251–1261. [Google Scholar] [CrossRef] [PubMed] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Trait | Abbreviation | Description of Measurement |
|---|---|---|
| Grain Yield per Spike | GYPS | Grain weight in gram per spike |
| Yield per Plot | YPP | Grain weight per area |
| Days to Flowering | DTF | Days from sowing to 50% flowering |
| Chlorophyll at Flowering | CCF | Measured by SPAD |
| Leaves Water content | WC | |
| Starch | Str | Fales (1951) and Schlegel (1956); mg/g DW |
| Carbohydrates | Carb | Fales (1951) and Schlegel (1956); mg/g DW |
| Protein content | PC | Lowery et al. (1951); mg/g DW |
| Amino acids | AA | Moore and Stein (1948); mg/g DW |
| Proline | Pro | (Bates et al. 1973); mg/g DW |
| Trait | Control | Heat Stress |
|---|---|---|
| Starch (Str) | 62.42 ** | 138.80 ** |
| Carbohydrates (Carb) | 101.95 ** | 161.96 ** |
| Amino Acid (AA) | 862.45 ** | 494.39 ** |
| Protein content (PC) | 22.42 ** | 56.16 ** |
| Proline content (Pro) | 816.03 ** | 979.71 ** |
| Trait | Class A (≤10%) | Class B (≤20%) | Class C (≤30%) | Class D (≤40%) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Min | Max | Mean | Min | Max | Mean | Min | Max | Mean | Min | Max | Mean | |
| Yield-Related Traits | ||||||||||||
| GYPS | −0.35 | −9.97 | −5.27 | −10.12 | −17.96 | −13.88 | −20.33 | −29.10 | −24.07 | −33.66 | −39.87 | −36.63 |
| YPP | −0.61 | −28.25 | −16.21 | −9.92 | −50.57 | −30.96 | −7.68 | −56.61 | −35.02 | −32.59 | −61.84 | −49.34 |
| TKW | −1.39 | −24.22 | −10.91 | −3.07 | −15.07 | −9.90 | −0.36 | −18.57 | −7.95 | −22.22 | −29.67 | −25.81 |
| DTF | −6.52 | −19.47 | −12.41 | −8.45 | −16.62 | −12.18 | −8.49 | −14.36 | −11.56 | 10.37 | −9.94 | −2.87 |
| Physiological Traits | ||||||||||||
| CCF | −22.94 | 69.96 | 14.54 | −22.59 | 30.17 | 8.16 | −15.61 | 35.18 | 13.94 | −17.13 | −27.03 | −22.68 |
| WC | −21.17 | 9.54 | −9.25 | −26.80 | 6.19 | −10.60 | −13.53 | −37.94 | −21.32 | −24.83 | −33.60 | −28.58 |
| Str | −13.68 | 109.31 | 10.84 | −43.64 | 45.78 | −8.99 | −4.76 | −41.27 | −22.15 | −45.88 | −53.63 | −50.76 |
| Car | −51.39 | 248.65 | 54.56 | −23.67 | 169.90 | 47.52 | −48.81 | 120.82 | 14.07 | −34.54 | −55.42 | −43.20 |
| AA | −88.13 | 561.62 | 63.04 | −74.39 | 463.27 | 116.44 | −83.58 | 355.75 | 12.78 | −50.99 | 60.98 | −13.35 |
| PCC | −36.68 | 325.63 | 56.70 | −90.85 | 371.32 | −10.16 | −84.65 | 141.29 | −22.10 | 0.00 | −52.97 | −28.20 |
| Pro | −1.27 | 356.09 | 126.29 | −4.15 | 239.84 | 92.31 | −50.60 | 210.73 | 2.57 | −47.60 | −64.13 | −54.44 |
| Trait | Chr | Gene Name | Gene Start | Gene End | Gene Length | Gene Annotation | Orthologs |
|---|---|---|---|---|---|---|---|
| PC, TKWH | 1 | HORVU1Hr1G013210 | 34083562 | 34088731 | 5170 | IQ-domain 2 | - |
| PC, TKWH | 1 | HORVU1Hr1G013360 | 35213246 | 35218550 | 5305 | Hexosyltransferase | AT3G25140 |
| PC, TKWH | 1 | HORVU1Hr1G013450 | 35606596 | 35616067 | 9472 | Alpha, alpha-trehalose-phosphate synthase [UDP-forming] 1 | AT1G68020 |
| PC, TKWH | 1 | HORVU1Hr1G013470 | 35676705 | 35678280 | 1576 | Delta-aminolevulinic acid dehydratase | AT1G44318 |
| PC, TKW | 1 | HORVU1Hr1G013480 | 35679610 | 35683434 | 3825 | sugar transporter 1 | AT1G08890 |
| PC, TKWH | 1 | HORVU1Hr1G013520 | 35975927 | 35977108 | 1182 | Pectin lyase-like superfamily protein | At3g16850 |
| PC, TKWH | 1 | HORVU1Hr1G013580 | 36169093 | 36172878 | 3786 | Importin subunit beta-1 | AT5G53480 |
| PC, TKWH | 1 | HORVU1Hr1G013600 | 36438691 | 36461615 | 22925 | E3 SUMO-protein ligase SIZ1 | AT5G60410 |
| PC, GYPS | 4 | HORVU4Hr1G034360 | 247366727 | 247371722 | 4996 | Nuclear transcription factor Y subunit C-3 | AT1G54830 |
| CCFH | 4 | HORVU4Hr1G034650 | 251600275 | 251637083 | 36809 | Protein ABERRANT POLLEN TRANSMISSION 1 | - |
| TKWH | 4 | HORVU4Hr1G034750 | 252632653 | 252637368 | 4716 | U4/U6 small nuclear ribonucleoprotein PRP4-like protein | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dawood, M.F.A.; Moursi, Y.S.; Amro, A.; Baenziger, P.S.; Sallam, A. Investigation of Heat-Induced Changes in the Grain Yield and Grains Metabolites, with Molecular Insights on the Candidate Genes in Barley. Agronomy 2020, 10, 1730. https://doi.org/10.3390/agronomy10111730
Dawood MFA, Moursi YS, Amro A, Baenziger PS, Sallam A. Investigation of Heat-Induced Changes in the Grain Yield and Grains Metabolites, with Molecular Insights on the Candidate Genes in Barley. Agronomy. 2020; 10(11):1730. https://doi.org/10.3390/agronomy10111730
Chicago/Turabian StyleDawood, Mona F. A., Yasser S. Moursi, Ahmed Amro, P. Stephen Baenziger, and Ahmed Sallam. 2020. "Investigation of Heat-Induced Changes in the Grain Yield and Grains Metabolites, with Molecular Insights on the Candidate Genes in Barley" Agronomy 10, no. 11: 1730. https://doi.org/10.3390/agronomy10111730
APA StyleDawood, M. F. A., Moursi, Y. S., Amro, A., Baenziger, P. S., & Sallam, A. (2020). Investigation of Heat-Induced Changes in the Grain Yield and Grains Metabolites, with Molecular Insights on the Candidate Genes in Barley. Agronomy, 10(11), 1730. https://doi.org/10.3390/agronomy10111730