Analysis of the Similarity between in Silico Ideotypes and Phenotypic Profiles to Support Cultivar Recommendation—A Case Study on Phaseolus vulgaris L.

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Characterization of the Study Area
2.2. Calibration of the Crop Model
2.3. Determination of Functional Trait Values
2.4. Ideotype Design and Cultivar Recommendation
3. Results
3.1. Model Evaluation
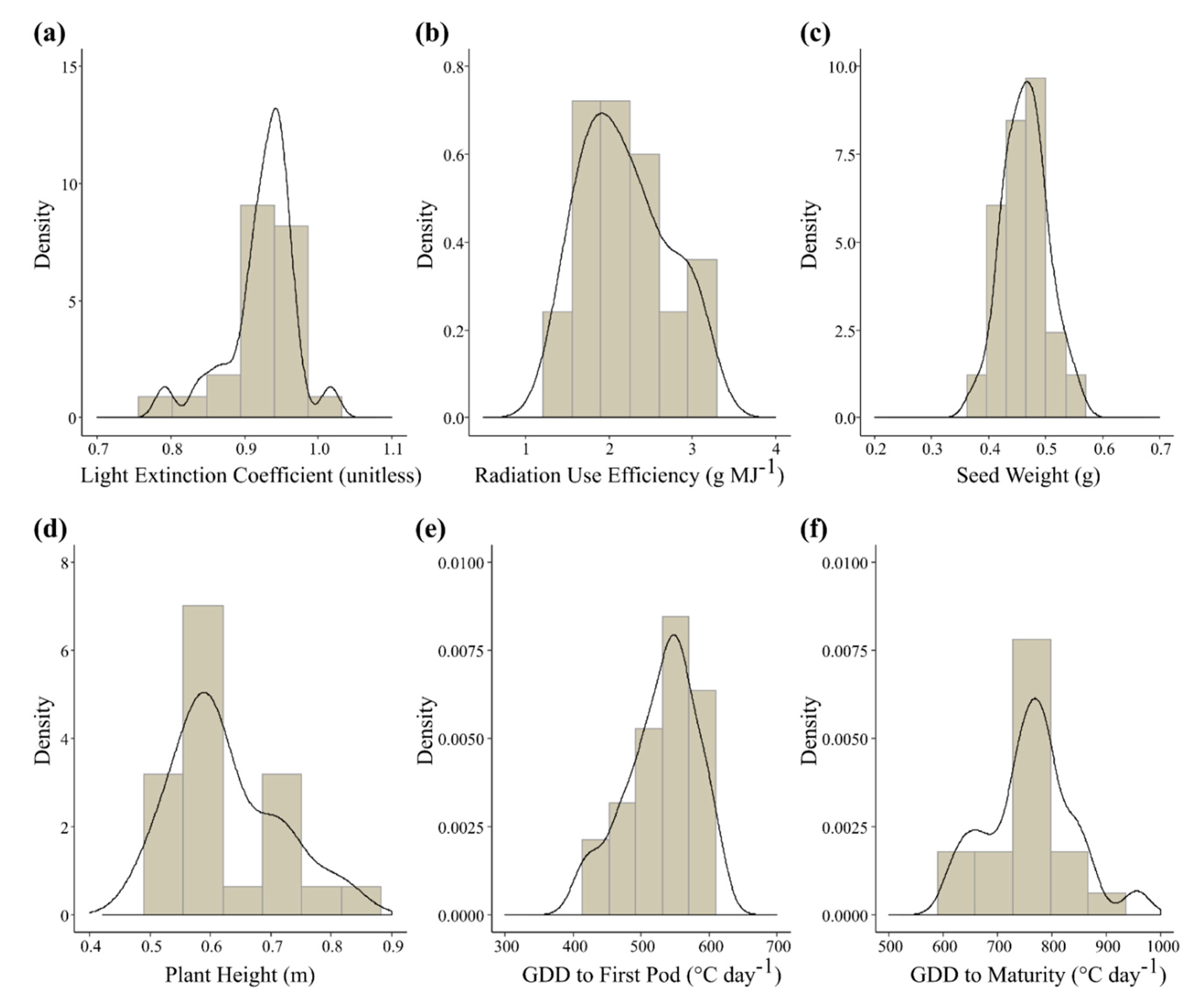
3.2. Variability of Functional Trait Values among Varieties
3.3. Ideotypes Derived for the Agro-Climatic Contexts
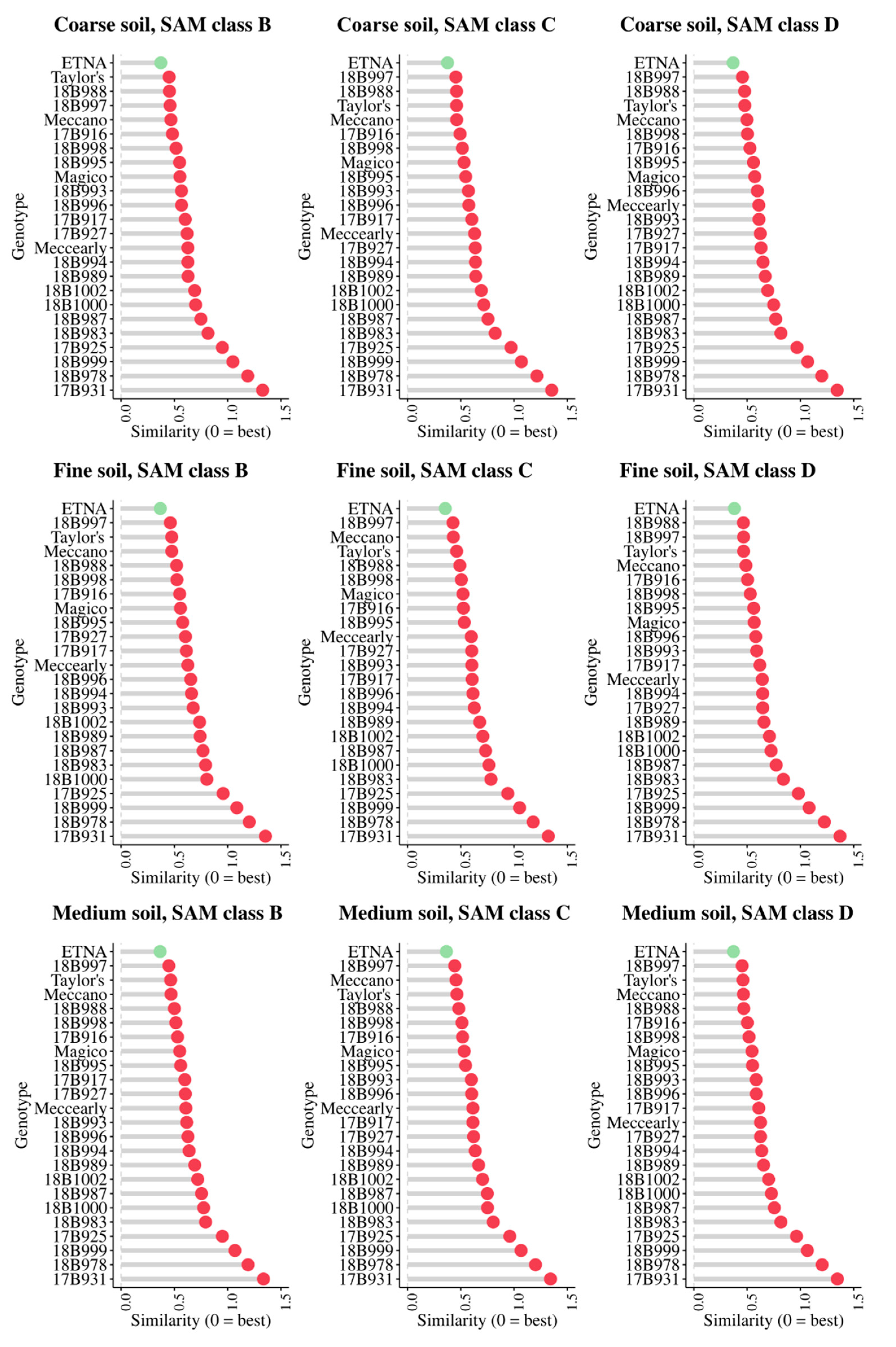
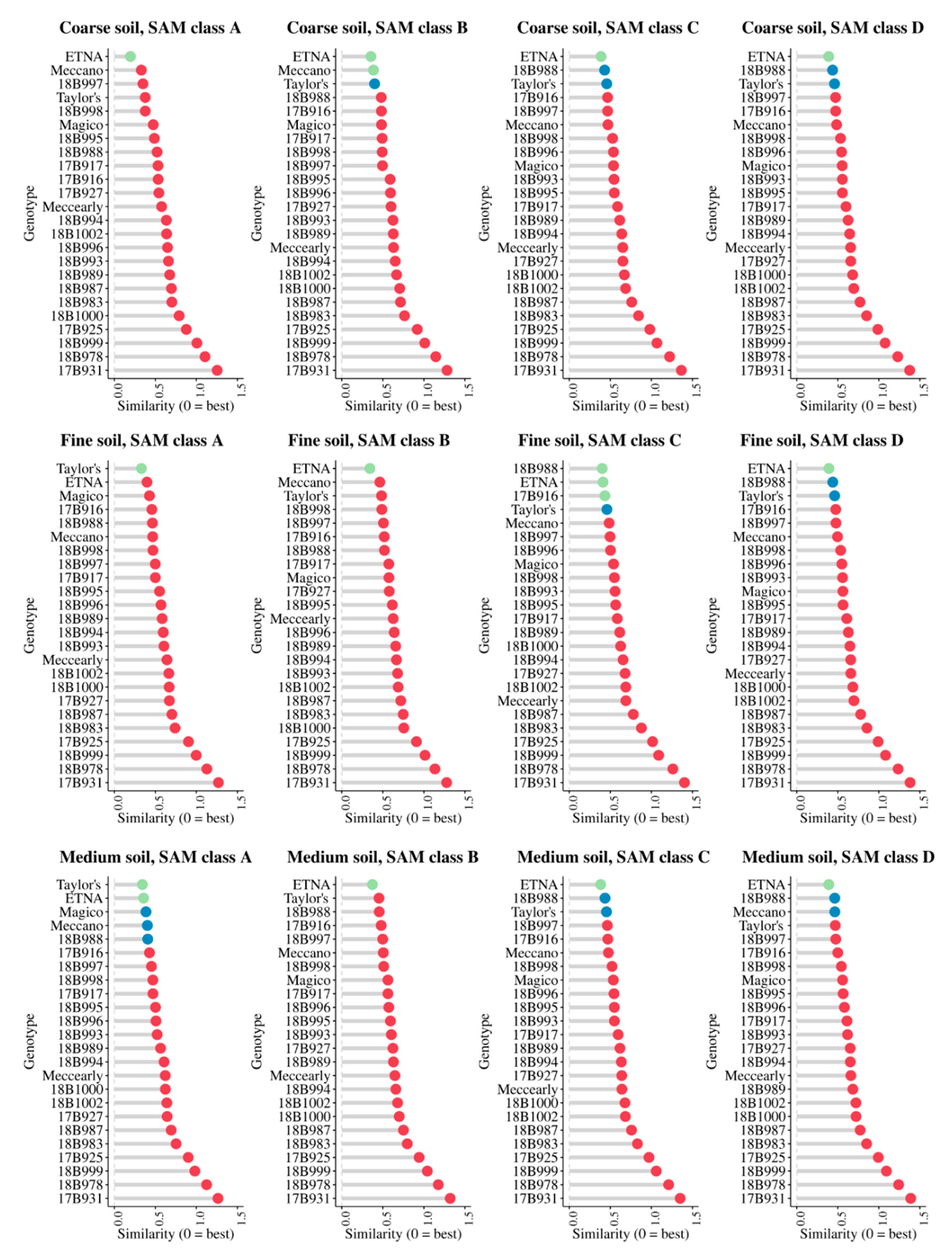
3.4. Cultivar Recommendation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Messina, C.; Hammer, G.; Dong, Z.; Podlich, D.; Cooper, M. Modelling crop improvement in a G×E×M framework via gene–trait–phenotype relationships. In Crop Physiology: Application for Genetic Improvement and Agronomy; Sadras, V., Calderini, D., Eds.; Academic press: London, UK, 2009; pp. 235–265. ISBN 978-0-12-374431-9. [Google Scholar]
- Annicchiarico, P. Genotype X Environment Interactions: Challenges and Opportunities for Plant Breeding and Cultivar Recommendations; FAO: Rome, Italy, 2002; ISBN 978-92-5-104870-2. [Google Scholar]
- DeLacy, I.H.; Basford, K.E.; Cooper, M.; Bull, J.K.; McLaren, C.G. Analysis of multi-environment trials—An historical perspective. In Plant Adaptation and Crop Improvement; Cooper, M., Hammer, G.L., Eds.; CAB International: Wallingford, UK, 1996; Volume 39124, pp. 39–124. [Google Scholar]
- Casadebaig, P.; Mestries, E.; Debaeke, P. A model-based approach to assist variety evaluation in sunflower crop. Eur. J. Agron. 2016, 81, 92–105. [Google Scholar] [CrossRef] [Green Version]
- Comstock, R.E. Quantitative Genetics and the Design of Breeding Programs. In Proceedings of the International Conference Quantitative Genetics, Ames, IA, USA, 16–21 August 1976; Iowa State University Press: Ames, IA, USA, 1977; pp. 1705–1718. [Google Scholar]
- Lúquez, J.E.; Aguirrezábal, L.A.N.; Agüero, M.E.; Pereyra, V.R. Stability and adaptability of cultivars in non-balanced yield trials. Comparison of methods for selecting ‘High Oleic’ sunflower hybrids for grain yield and quality. J. Agron. Crop Sci. 2002, 188, 225–234. [Google Scholar] [CrossRef]
- Kang, M.S. Breeding: Genotype-by-environment interaction. In Encyclopedia of Plant and Crop Science; Goodman, R.M., Ed.; CRC Press: Boca Raton, FL, USA, 2004; pp. 218–221. [Google Scholar]
- Fan, X.-M.; Kang, M.S.; Chen, H.; Zhang, Y.; Tan, J.; Xu, C. Yield stability of maize hybrids evaluated in multi-environment trials in Yunnan, China. Agron. J. 2007, 99, 220–228. [Google Scholar] [CrossRef]
- Chenu, K.; Cooper, M.; Hammer, G.L.; Mathews, K.L.; Dreccer, M.F.; Chapman, S.C. Environment characterization as an aid to wheat improvement: Interpreting genotype–environment interactions by modelling water-deficit patterns in North-Eastern Australia. J. Exp. Bot. 2011, 62, 1743–1755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeuffroy, M.-H.; Casadebaig, P.; Debaeke, P.; Loyce, C.; Meynard, J.-M. Agronomic model uses to predict cultivar performance in various environments and cropping systems. A review. Agron. Sustain. Dev. 2014, 34, 121–137. [Google Scholar] [CrossRef]
- Chenu, K.; Porter, J.R.; Martre, P.; Basso, B.; Chapman, S.C.; Ewert, F.; Bindi, M.; Asseng, S. Contribution of crop models to adaptation in wheat. Trends Plant Sci. 2017, 22, 472–490. [Google Scholar] [CrossRef] [PubMed]
- Chapman, S.C.; Cooper, M.; Hammer, G.L.; Butler, D.G. Genotype by environment interactions affecting grain sorghum. II. Frequencies of different seasonal patterns of drought stress are related to location effects on hybrid yields. Aust. J. Agric. Res. 2000, 51, 209. [Google Scholar] [CrossRef]
- Hammer, G.L.; Jordan, D.R. An integrated systems approach to crop improvement. In Scale and Complexity in Plant Systems Research: Gene–Plant–Crop Relations; Wageningen UR: Frontis Series; Spiertz, J.H.J., Struik, P.C., van Laar, H.H., Eds.; Springer: Dordrecht, The Netherlands, 2007; pp. 45–61. [Google Scholar]
- Paleari, L.; Movedi, E.; Cappelli, G.; Wilson, L.T.; Confalonieri, R. Surfing parameter hyperspaces under climate change scenarios to design future rice ideotypes. Glob. Change Biol. 2017, 23, 4651–4662. [Google Scholar] [CrossRef]
- Tao, F.; Rötter, R.P.; Palosuo, T.; Díaz-Ambrona, C.G.H.; Mínguez, M.I.; Semenov, M.A.; Kersebaum, K.C.; Nendel, C.; Cammarano, D.; Hoffmann, H.; et al. Designing future barley ideotypes using a crop model ensemble. Eur. J. Agron. 2017, 82, 144–162. [Google Scholar] [CrossRef]
- Violle, C.; Navas, M.-L.; Vile, D.; Kazakou, E.; Fortunel, C.; Hummel, I.; Garnier, E. Let the concept of trait be functional! Oikos 2007, 116, 882–892. [Google Scholar] [CrossRef]
- Paleari, L.; Cappelli, G.; Bregaglio, S.; Acutis, M.; Donatelli, M.; Sacchi, G.A.; Lupotto, E.; Boschetti, M.; Manfron, G.; Confalonieri, R. District specific, in silico evaluation of rice ideotypes improved for resistance/tolerance traits to biotic and abiotic stressors under climate change scenarios. Clim. Change 2015, 132, 661–675. [Google Scholar] [CrossRef]
- Gauffreteau, A. Using ideotypes to support selection and recommendation of varieties. OCL 2018, 25, D602. [Google Scholar] [CrossRef]
- Broughton, W.J.; Hernández, G.; Blair, M.; Beebe, S.; Gepts, P.; Vanderleyden, J. Beans (Phaseolus spp.)—model food legumes. Plant Soil 2003, 252, 55–128. [Google Scholar] [CrossRef] [Green Version]
- Graham, P.H.; Ranalli, P. Common bean (Phaseolus vulgaris L.). Field Crop. Res. 1997, 53, 131–146. [Google Scholar] [CrossRef]
- Beebe, S.E.; Rao, I.M.; Blair, M.W.; Acosta-Gallegos, J.A. Phenotyping common beans for adaptation to drought. Front. Physiol. 2013, 4. [Google Scholar] [CrossRef] [Green Version]
- Mousavi-Derazmahalleh, M.; Bayer, P.E.; Hane, J.K.; Valliyodan, B.; Nguyen, H.T.; Nelson, M.N.; Erskine, W.; Varshney, R.K.; Papa, R.; Edwards, D. Adapting legume crops to climate change using genomic approaches. Plant Cell Environ. 2019, 42, 6–19. [Google Scholar] [CrossRef] [Green Version]
- Beebe, S.; Ramirez, J.; Jarvis, A.; Rao, I.M.; Mosquera, G.; Bueno, J.M.; Blair, M.W. Genetic improvement of common beans and the challenges of climate change. In Crop Adaptation to Climate Change; Yadav, S.S., Redden, R.J., Hatfield, J.L., Lotze-Campen, H., Hall, A.E., Eds.; Wiley-Blackwell: Oxford, UK, 2011; pp. 356–369. ISBN 978-0-470-96092-9. [Google Scholar]
- Brisson, N.; Mary, B.; Ripoche, D.; Jeuffroy, M.H.; Ruget, F.; Nicoullaud, B.; Gate, P.; Devienne-Barret, F.; Antonioletti, R.; Durr, C.; et al. STICS: A generic model for the simulation of crops and their water and nitrogen balances. I. Theory and parameterization applied to wheat and corn. Agronomie 1998, 18, 311–346. [Google Scholar] [CrossRef]
- Brisson, N.; Launay, M.; Mary, B.; Beaudoin, N. Conceptual Basis, Formalisations and Parameterization of the Stics Crop Model; Editions Quae: Paris, France, 2009; ISBN 978-2-7592-0290-4. [Google Scholar]
- Annicchiarico, P. Coping with and exploiting genotype-by-environment interactions. In Plant Breeding and Farmer Participation; Ceccarelli, S., Guimarães, E.P., Weltzien, E., Eds.; FAO: Rome, Italy, 2009; pp. 519–564. [Google Scholar]
- Al Majou, H.; Bruand, A.; Duval, O.; Le Bas, C.; Vautier, A. Prediction of soil water retention properties after stratification by combining texture, bulk density and the type of horizon. Soil Use Manag. 2008, 24, 383–391. [Google Scholar] [CrossRef] [Green Version]
- Antolini, G.; Auteri, L.; Pavan, V.; Tomei, F.; Tomozeiu, R.; Marletto, V. A daily high-resolution gridded climatic data set for Emilia-Romagna, Italy, during 1961–2010. Int. J. Climatol. 2016, 36, 1970–1986. [Google Scholar] [CrossRef] [Green Version]
- Hargreaves, G.H.; Samani, Z.A. Estimating potential evapotranspiration. J. Irrig. Drain. Div. 1982, 108, 225–230. [Google Scholar]
- Hargreaves, G.H. Reference crop evapotranspiration from temperature. Appl. Eng. Agric. 1985, 1, 96–99. [Google Scholar] [CrossRef]
- Confalonieri, R.; Bregaglio, S.; Acutis, M. A proposal of an indicator for quantifying model robustness based on the relationship between variability of errors and of explored conditions. Ecol. Model. 2010, 221, 960–964. [Google Scholar] [CrossRef]
- Brisson, N.; Gary, C.; Justes, E.; Roche, R.; Mary, B.; Ripoche, D.; Zimmer, D.; Sierra, J.; Bertuzzi, P.; Burger, P.; et al. An overview of the crop model stics. Eur. J. Agron. 2003, 18, 309–332. [Google Scholar] [CrossRef]
- Coucheney, E.; Buis, S.; Launay, M.; Constantin, J.; Mary, B.; García de Cortázar-Atauri, I.; Ripoche, D.; Beaudoin, N.; Ruget, F.; Andrianarisoa, K.S.; et al. Accuracy, robustness and behavior of the STICS soil–crop model for plant, water and nitrogen outputs: Evaluation over a wide range of agro-environmental conditions in France. Environ. Model. Softw. 2015, 64, 177–190. [Google Scholar] [CrossRef]
- Falconnier, G.N.; Journet, E.-P.; Bedoussac, L.; Vermue, A.; Chlébowski, F.; Beaudoin, N.; Justes, E. Calibration and evaluation of the STICS soil-crop model for faba bean to explain variability in yield and N2 fixation. Eur. J. Agron. 2019, 104, 63–77. [Google Scholar] [CrossRef]
- Ravasi, R.A.; Paleari, L.; Vesely, F.M.; Movedi, E.; Thoelke, W.; Confalonieri, R. Ideotype definition to adapt legumes to climate change: A case study for field pea in Northern Italy. Agric. For. Meteorol. 2020, 291, 108081. [Google Scholar] [CrossRef]
- Confalonieri, R.; Bregaglio, S.; Acutis, M. Quantifying plasticity in simulation models. Ecol. Model. 2012, 225, 159–166. [Google Scholar] [CrossRef]
- Meier, U. Growth Stages of Mono-And Dicotyledonous Plants, 2nd ed.; Federal Biological Research Centre for Agriculture and Forestry: Braunschweig, Germany, 2001. [Google Scholar]
- Confalonieri, R.; Foi, M.; Casa, R.; Aquaro, S.; Tona, E.; Peterle, M.; Boldini, A.; De Carli, G.; Ferrari, A.; Finotto, G.; et al. Development of an app for estimating leaf area index using a smartphone. Trueness and precision determination and comparison with other indirect methods. Comput. Electron. Agric. 2013, 96, 67–74. [Google Scholar] [CrossRef]
- Willmott, C.; Matsuura, K. Advantages of the mean absolute error (MAE) over the root mean square error (RMSE) in assessing average model performance. Clim. Res. 2005, 30, 79–82. [Google Scholar] [CrossRef]
- Jørgensen, S.E.; Kamp-Nielsen, L.; Christensen, T.; Windolf-Nielsen, J.; Westergaard, B. Validation of a prognosis based upon a eutrophication model. Ecol. Model. 1986, 32, 165–182. [Google Scholar] [CrossRef]
- Nash, J.E.; Sutcliffe, J.V. River flow forecasting through conceptual models part I—A discussion of principles. J. Hydrol. 1970, 10, 282–290. [Google Scholar] [CrossRef]
- Loague, K.; Green, R.E. Statistical and graphical methods for evaluating solute transport models: Overview and application. J. Contam. Hydrol. 1991, 7, 51–73. [Google Scholar] [CrossRef]
- Confalonieri, R.; Paleari, L.; Foi, M.; Movedi, E.; Vesely, F.M.; Thoelke, W.; Agape, C.; Borlini, G.; Ferri, I.; Massara, F.; et al. PocketPlant3D: Analysing canopy structure using a smartphone. Biosyst. Eng. 2017, 164, 1–12. [Google Scholar] [CrossRef]
- Campbell, G.S. Extinction coefficients for radiation in plant canopies calculated using an ellipsoidal inclination angle distribution. Agric. For. Meteorol. 1986, 36, 317–321. [Google Scholar] [CrossRef]
- Paleari, L.; Movedi, E.; Vesely, F.M.; Confalonieri, R. Tailoring parameter distributions to specific germplasm: Impact on crop model-based ideotyping. Sci. Rep. 2019, 9, 18309. [Google Scholar] [CrossRef] [PubMed]
- Grubbs, F.E. Procedures for Detecting Outlying Observations in Samples. Technometrics 1969, 11, 1–21. [Google Scholar] [CrossRef]
- Shapiro, S.S.; Wilk, M.B. An analysis of variance test for normality (Complete Samples). Biometrika 1965, 52, 591. [Google Scholar] [CrossRef]
- Saltelli, A.; Tarantola, S.; Chan, K.P.-S. A quantitative model-independent method for global sensitivity analysis of model output. Technometrics 1999, 41, 39–56. [Google Scholar] [CrossRef]
- Confalonieri, R.; Bellocchi, G.; Bregaglio, S.; Donatelli, M.; Acutis, M. Comparison of sensitivity analysis techniques: A case study with the rice model WARM. Ecol. Model. 2010, 221, 1897–1906. [Google Scholar] [CrossRef]
- Carvalho, C.G.P.; Cruz, C.D.; Viana, J.M.S.; Silva, D.J.H. Selection based on distances from ideotype. Crop Breed. Appl. Biotechnol. 2002, 2, 171–178. [Google Scholar] [CrossRef]
- Tesfaye, K.; Walker, S.; Tsubo, M. Radiation interception and radiation use efficiency of three grain legumes under water deficit conditions in a semi-arid environment. Eur. J. Agron. 2006, 25, 60–70. [Google Scholar] [CrossRef]
- Karimzadeh Soureshjani, H.; Nezami, A.; Kafi, M.; Tadayon, M. Responses of two common bean (Phaseolus vulgaris L.) genotypes to deficit irrigation. Agric. Water Manag. 2019, 213, 270–279. [Google Scholar] [CrossRef]
- Mercati, F.; Leone, M.; Lupini, A.; Sorgonà, A.; Bacchi, M.; Abenavoli, M.R.; Sunseri, F. Genetic diversity and population structure of a common bean (Phaseolus vulgaris L.) collection from Calabria (Italy). Genet. Resour. Crop Evol. 2013, 60, 839–852. [Google Scholar] [CrossRef]
- Campion, B.; Perrone, D.; Galasso, I.; Bollini, R. Common bean (Phaseolus vulgaris L.) lines devoid of major lectin proteins. Plant Breed. 2009, 128, 199–204. [Google Scholar] [CrossRef]
- Tardieu, F. Any trait or trait-related allele can confer drought tolerance: Just design the right drought scenario. J. Exp. Bot. 2012, 63, 25–31. [Google Scholar] [CrossRef] [Green Version]
- Farooq, M.; Nadeem, F.; Gogoi, N.; Ullah, A.; Alghamdi, S.S.; Nayyar, H.; Siddique, K.H.M. Heat stress in grain legumes during reproductive and grain-filling phases. Crop Pasture Sci. 2017, 68, 985–1005. [Google Scholar] [CrossRef]
- Barkley, A.; Peterson, H.H.; Shroyer, J. Wheat variety selection to maximize returns and minimize risk: An application of portfolio theory. J. Agric. Appl. Econ. 2010, 42, 39–55. [Google Scholar] [CrossRef] [Green Version]
- Berry, P.M.; Sylvester-Bradley, R.; Berry, S. Ideotype design for lodging-resistant wheat. Euphytica 2007, 154, 165–179. [Google Scholar] [CrossRef]
- Singh, P.; Nedumaran, S.; Traore, P.C.S.; Boote, K.J.; Rattunde, H.F.W.; Prasad, P.V.V.; Singh, N.P.; Srinivas, K.; Bantilan, M.C.S. Quantifying potential benefits of drought and heat tolerance in rainy season sorghum for adapting to climate change. Agric. For. Meteorol. 2014, 185, 37–48. [Google Scholar] [CrossRef] [Green Version]
- Boote, K.J.; Prasad, V.; Allen, L.H.; Singh, P.; Jones, J.W. Modeling sensitivity of grain yield to elevated temperature in the DSSAT crop models for peanut, soybean, dry bean, chickpea, sorghum, and millet. Eur. J. Agron. 2018, 100, 99–109. [Google Scholar] [CrossRef]
- Willocquet, L.; Savary, S.; McDonald, B.A.; Mikaberidze, A. A polyetic modelling framework for plant disease emergence. Plant Pathol. 2020, 69, 91045. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Process | Trait | Parameter (Acronym) | Unit | Distribution | Shapiro-Wilk Test |
|---|---|---|---|---|---|
| Photosynthesis | Radiation use efficiency | Maximum radiation use efficiency of the vegetative phase (efcroiveg) | g MJ−1 | Normal (2.2 ± 0.5) | W = 0.949, p-value = 0.256 |
| Yield formation | Seed weight | Maximum grain weight at 0% humidity (pgrainmaxi) | g | Normal (0.46 ± 0.04) | W = 0.987, p-value = 0.983 |
| Canopy structure | Plant height | Maximum plant height (hautmax) | m | Normal (0.6 ± 0.09) | W = 0.921, p-value = 0.062 |
| Light extinction coefficient | Extinction coefficient for solar radiation (extin) | - | Normal (0.9 ± 0.05) | W = 0.912, p-value = 0.039 a | |
| Phenological development | Thermal time to first pod | Cumulative thermal time from emergence to the onset of filling of harvested organs (stlevdrp) | °C-day | Normal (530 ± 52) | W = 0.955, p-value = 0.349 |
| Thermal time to maturity | Cumulative thermal time from beginning of harvested organs filling to maturity (stdrpmat) | °C-day | Normal (761 ± 80) | W = 0.973, p-value = 0.752 |
| Activity | Variable | MAE | RRMSE | EF | CRM | R2 |
|---|---|---|---|---|---|---|
| Calibration | Leaf area index (−) | 0.70 | 29.8 | 0.38 | 0.04 | 0.68 |
| Aboveground biomass (t ha−1) | 0.91 | 21.4 | 0.93 | 0.01 | 0.99 | |
| Pod biomass (t ha−1) | 0.73 | 42.6 | 0.77 | −0.35 | 0.94 | |
| Yield (t ha−1) | 0.40 | 12.9 | 0.10 | −0.10 | 0.64 | |
| Validation | Leaf area index (−) | 0.82 | 29.1 | 0.79 | 0.12 | 0.96 |
| Aboveground Biomass (t ha−1) | 0.40 | 13.0 | 0.96 | 0.03 | 0.98 | |
| Pod biomass (t ha−1) | 0.38 | 29.6 | 0.93 | −0.09 | 0.95 | |
| Yield (t ha−1) | 0.32 | 8.4 | 0.31 | −0.04 | 0.69 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Paleari, L.; Vesely, F.M.; Ravasi, R.A.; Movedi, E.; Tartarini, S.; Invernizzi, M.; Confalonieri, R. Analysis of the Similarity between in Silico Ideotypes and Phenotypic Profiles to Support Cultivar Recommendation—A Case Study on Phaseolus vulgaris L. Agronomy 2020, 10, 1733. https://doi.org/10.3390/agronomy10111733
Paleari L, Vesely FM, Ravasi RA, Movedi E, Tartarini S, Invernizzi M, Confalonieri R. Analysis of the Similarity between in Silico Ideotypes and Phenotypic Profiles to Support Cultivar Recommendation—A Case Study on Phaseolus vulgaris L. Agronomy. 2020; 10(11):1733. https://doi.org/10.3390/agronomy10111733
Chicago/Turabian StylePaleari, Livia, Fosco M. Vesely, Riccardo A. Ravasi, Ermes Movedi, Sofia Tartarini, Mattia Invernizzi, and Roberto Confalonieri. 2020. "Analysis of the Similarity between in Silico Ideotypes and Phenotypic Profiles to Support Cultivar Recommendation—A Case Study on Phaseolus vulgaris L." Agronomy 10, no. 11: 1733. https://doi.org/10.3390/agronomy10111733
APA StylePaleari, L., Vesely, F. M., Ravasi, R. A., Movedi, E., Tartarini, S., Invernizzi, M., & Confalonieri, R. (2020). Analysis of the Similarity between in Silico Ideotypes and Phenotypic Profiles to Support Cultivar Recommendation—A Case Study on Phaseolus vulgaris L. Agronomy, 10(11), 1733. https://doi.org/10.3390/agronomy10111733