Assessing the Effect of Rubber (Hevea brasiliensis (Willd. ex A. Juss.) Muell. Arg.) Leaf Chemical Composition on Some Soil Properties of Differently Aged Rubber Tree Plantations


 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site and Treatment
2.2. Plant and Soil Sampling
2.3. Chemical and Biological Analyses and Leaf Chemical Analysis
2.4. Synchrotron-Based Fourier Transform Infrared (FTIR) Microspectroscopy
2.4.1. Plant and Soil Sample Preparation for Fourier Transform Infrared (FTIR) Microspectroscopy Analysis
2.4.2. Fourier Transform Infrared (FTIR) Microspectroscopy Analysis
2.4.3. Data Processing and Image Analysis
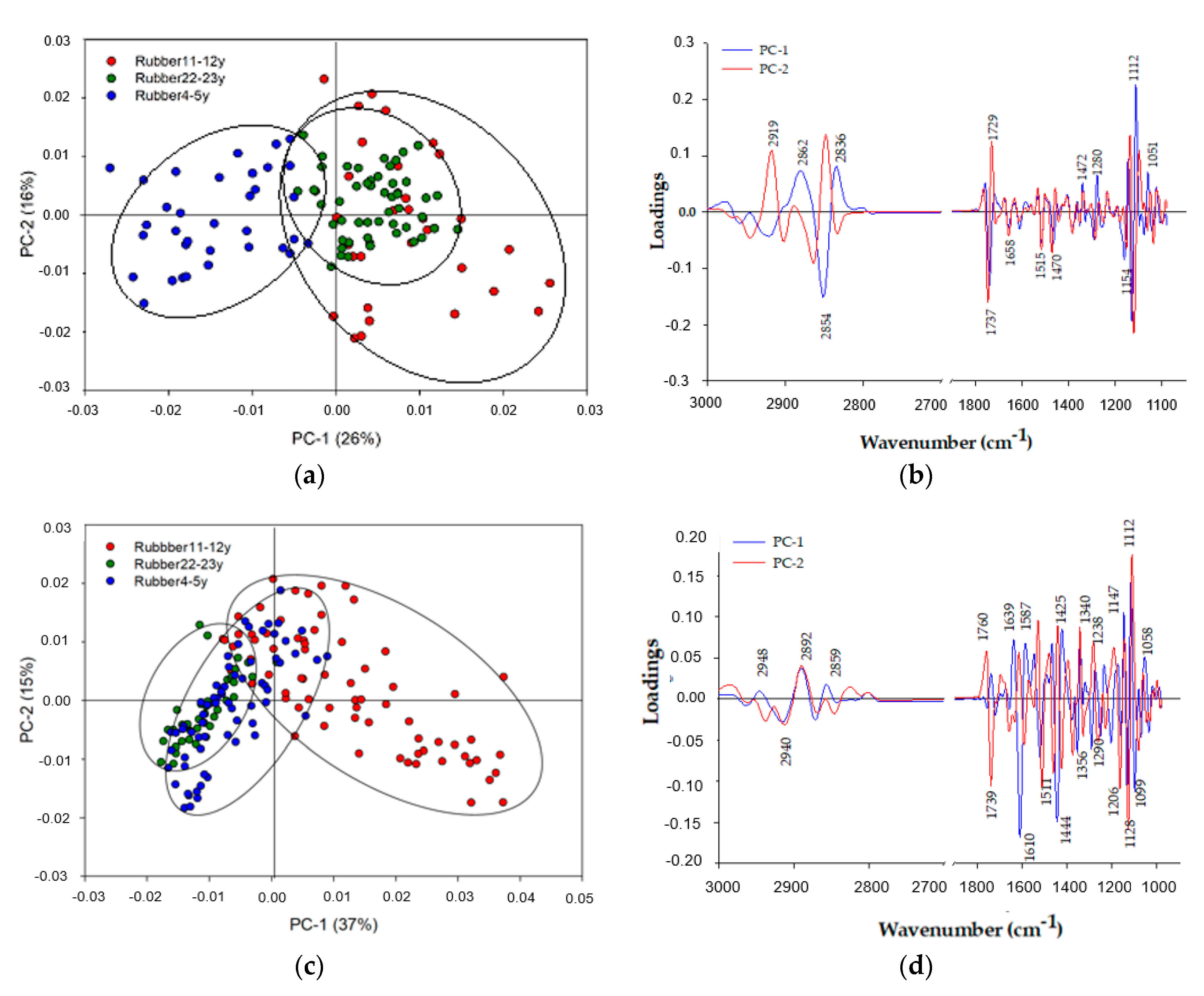
2.5. Multivariate Data Analysis
2.6. Statistical Analyses
3. Results
3.1. Soil Properties
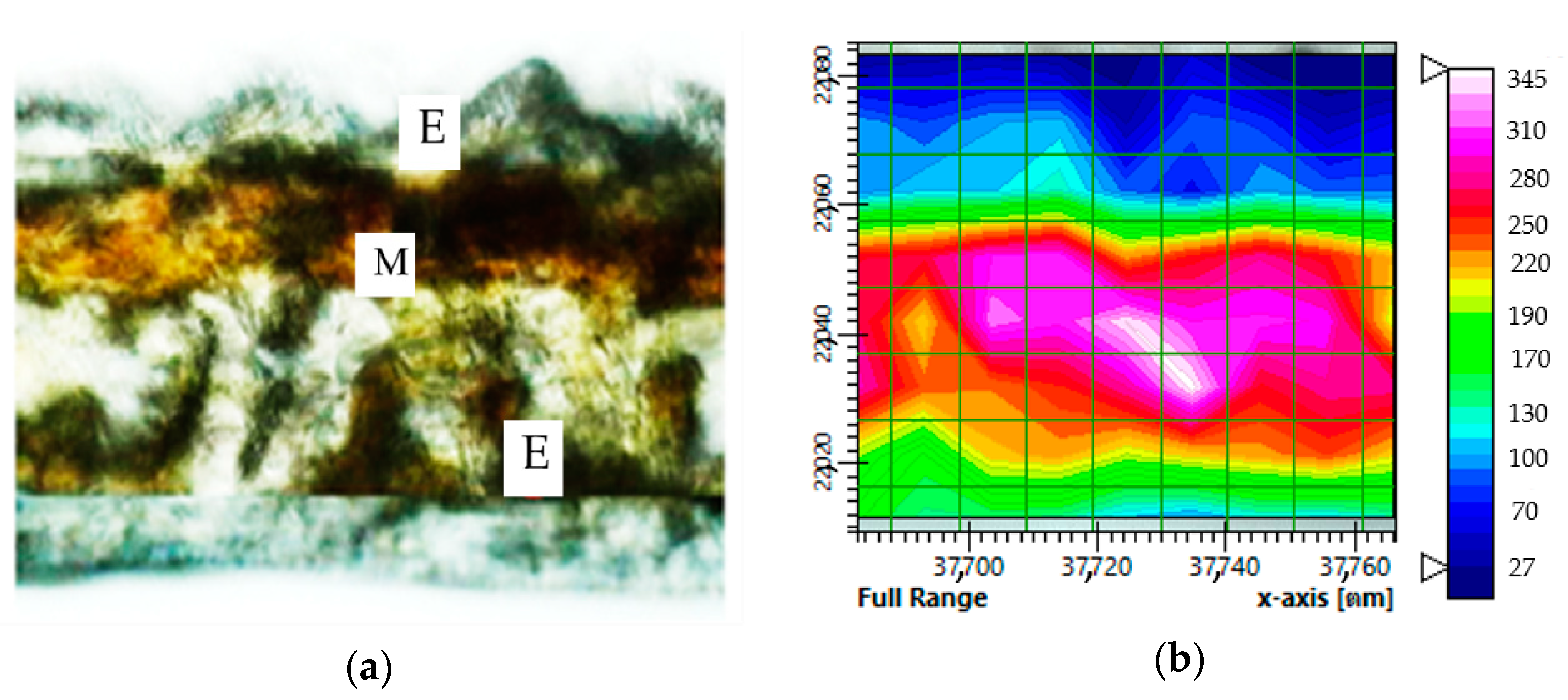
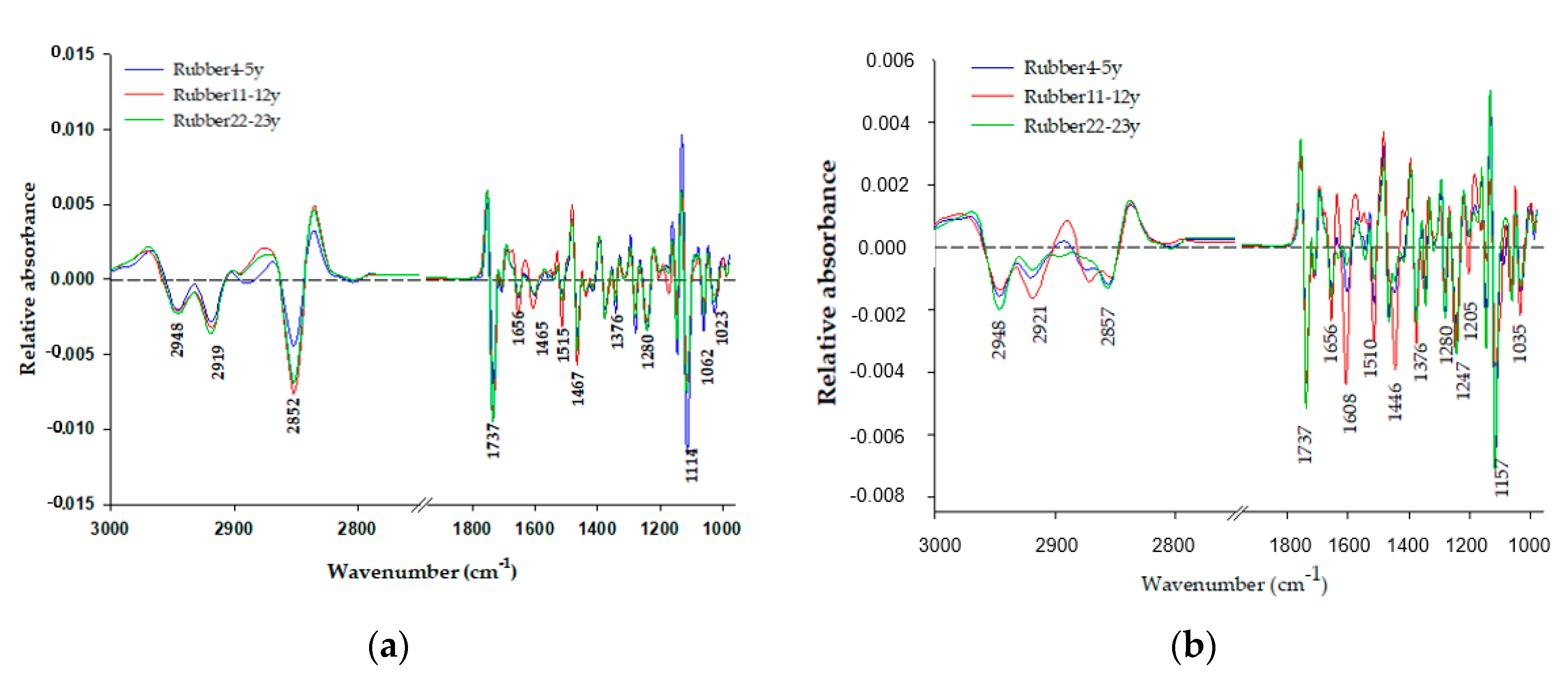
3.2. Synchrotron-Based Fourier Transform Infrared (Sr-FTIR) Microspectroscopy of Soil Organic Carbon
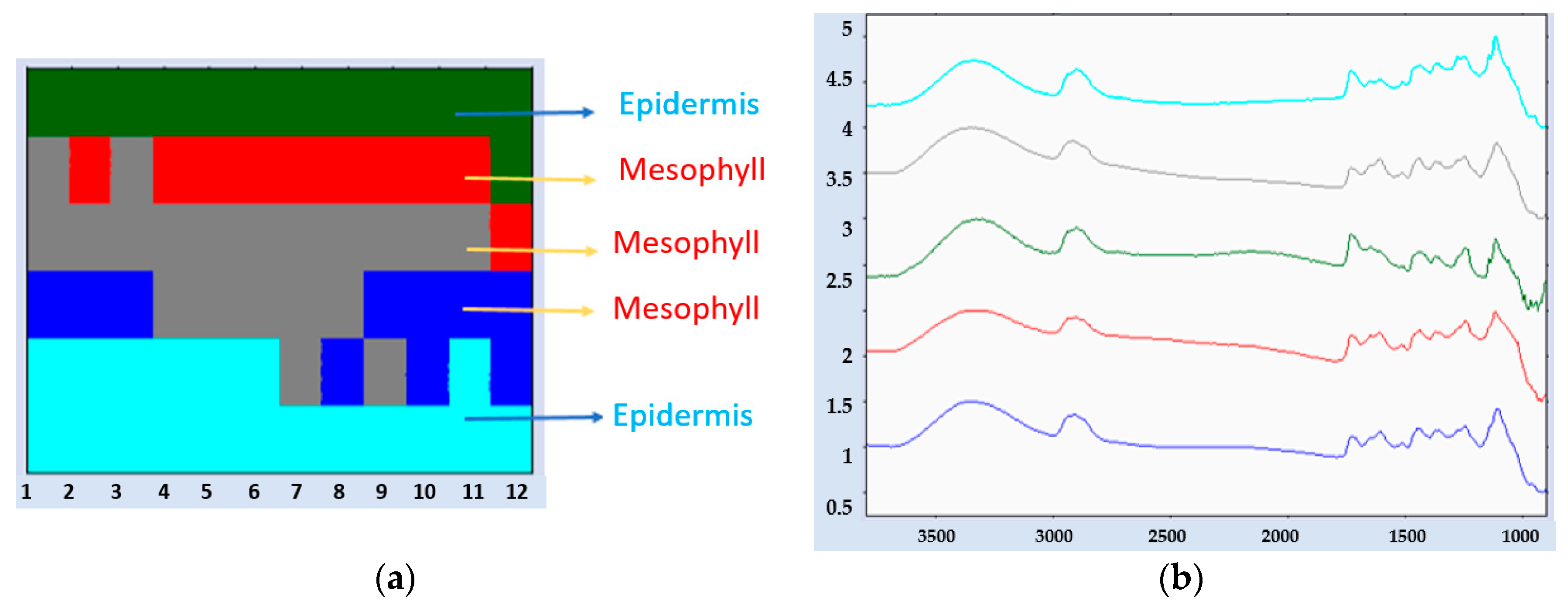
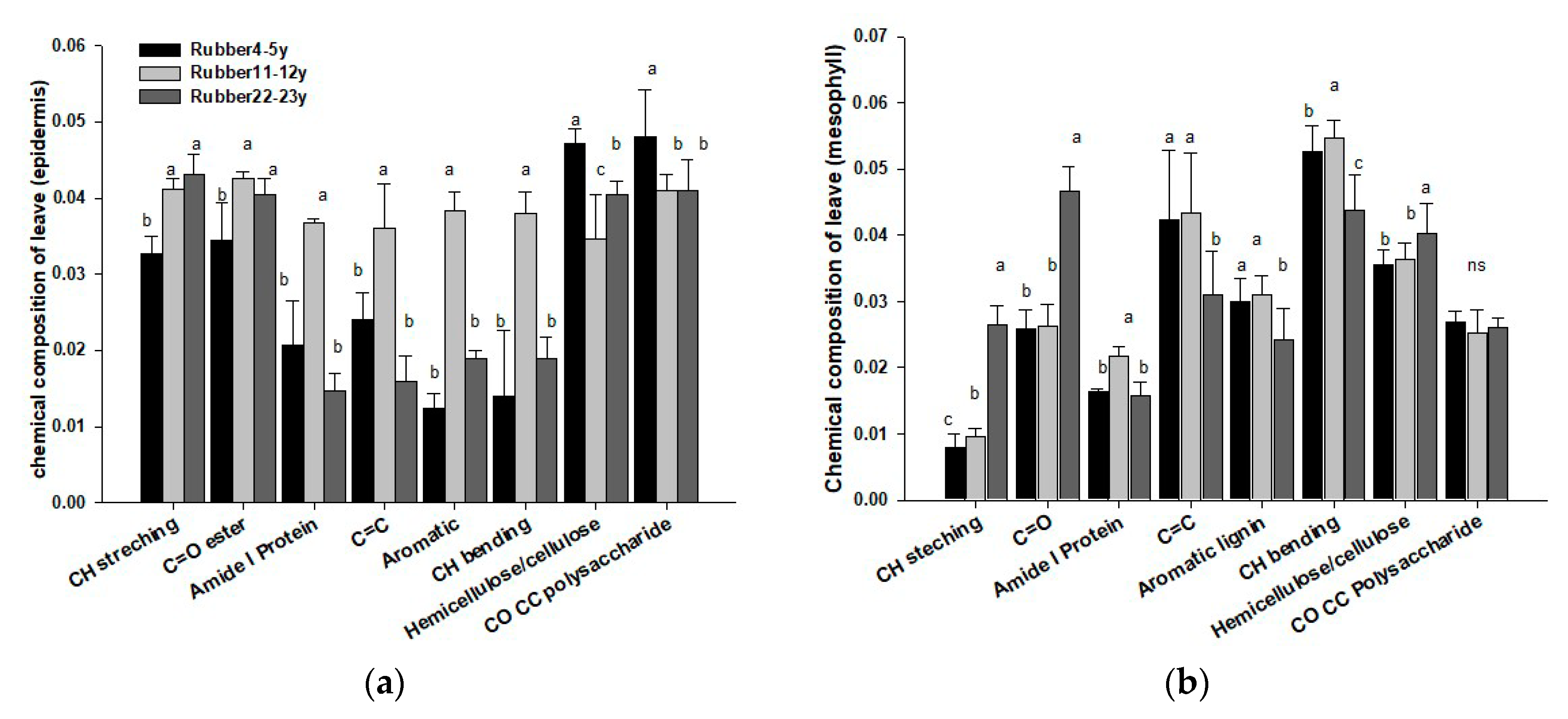
3.3. Distribution of Carbon Chemical Composition in Plant Leaves
4. Discussion
4.1. Relationships of Synchrotron-Based Fourier Transform Infrared (Sr-FTIR) Bands with Bio-Chemical Properties of Soil
4.2. Relationships of Synchrotron-Based Fourier Transform Infrared (Sr-FTIR) Peak Areas with the Biochemical Composition of the Rubber Leaf and Certain Soil Properties
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Li, Z.; Fox, J.M. Mapping rubber tree growth in mainland Southeast Asia using time-series MODIS 250 m NDVI and statistical data. Appl. Geogr. 2012, 32, 420–432. [Google Scholar] [CrossRef]
- Thaochan, N.; Pornsuriya, C.; Chairin, T.; Sunpapao, A. Role of systemic fungicide in antifungal activity and induced defense responses in rubber tree (Hevea brasiliensis) against leaf fall disease caused by Neopestalotiopsis cubana. Physiol. Mol. Plant. Pathol. 2020, 111, 101511. [Google Scholar] [CrossRef]
- Calderon, F.J.; Mikha, M.M.; Vigil, M.F.; Nielsen, D.C.; Benjamin, J.G.; Reeves, J.B. Diffuse-reflectance mid-infrared spectral properties of soil under alternative crop rotations in a semi-arid climate. Commun. Soil. Sci. Plant Anal. 2011, 42, 2143–2159. [Google Scholar] [CrossRef]
- Stohlgren, T.J. Litter dynamic in two Sierran mixed conifer forest. I. Litterfall and decomposition rates. Can. J. For. Res. 1988, 18, 1127–1135. [Google Scholar] [CrossRef]
- Baker, M.J.; Trevisan, J.; Bassan, P.; Bhargava, R.; Butler, H.J.; Dorling, K.M.; Fielden, P.R.; Fogarty, S.W.; Fullwood, N.J.; Heys, K.A.; et al. Using Fourier transform IR spectroscopy to analyze biological materials. Nat. Protoc. 2014, 9, 1771–1791. [Google Scholar] [CrossRef] [Green Version]
- Bouyanfif, A.; Liyanage, S.; Hequet, E.; Moustaid-Moussa, N.; Abidi, N. Review of FTIR microspectroscopy applications to investigate biochemical changes in C. elegans. Vib. Spectrosc. 2018, 96, 74–82. [Google Scholar] [CrossRef]
- Ribeiro da Luz, B. Attenuated total reflectance spectroscopy of plant leaves: A tool for ecological and botanical studies. New Phytol. 2006, 172, 305–318. [Google Scholar] [CrossRef]
- Schulz, H.; Baranska, M. Identification and quantification of valuable plant substances by IR and Raman spectroscopy. Vib. Spectrosc. 2007, 43, 13–25. [Google Scholar] [CrossRef]
- Talari, A.C.S.; Martinez, M.A.G.; Movasaghi, Z.; Rehman, S.; Rehman, I.U. Advances in Fourier transform infrared (FTIR) spectroscopy of biological tissues. Appl. Spectrosc. Rev. 2017, 52, 456–506. [Google Scholar] [CrossRef]
- Smith, B.C. Fundamentals of Fourier Transform Infrared Spectroscopy, 2nd ed.; CRC Press: Boca Raton, FL, USA, 2011; ISBN 978-1-4200-6929-7. [Google Scholar]
- Lawongsa, P.; Somboon, S.; Kamolmanit, B.; Namanusart, W.; Thumanu, K. The appearance of soil organic carbon compositions for short-term applications of biochemical contrasting organic residue using synchrotron-based fouier transform infrared (FTIR) microspectroscopy. Biosci. Res. 2020, 17, 257–266. [Google Scholar]
- Euring, D.; Löfke, C.; Teichmann, T.; Polle, A. Nitrogen fertilization has differential effects on N allocation and lignin in two Populus species with contrasting ecology. Trees 2012, 26, 1933–1942. [Google Scholar] [CrossRef] [Green Version]
- Usman, K.; Al-Ghouti, M.A.; Abu-Dieyeh, M.H. The assessment of cadmium, chromium, copper, and nickel tolerance and bioaccumulation by shrub plant Tetraena qataranse. Sci. Rep. 2019, 9, 5658. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McNear, D.H.; Chaney, R.L.; Sparks, D.L. The hyperaccumulator Alyssum murale uses complexation with nitrogen and oxygen donor ligands for Ni transport and storage. Phytochemistry 2010, 71, 188–200. [Google Scholar] [CrossRef] [PubMed]
- Xue, S.G.; Zhu, F.; Ye, C.; Wang, J.; Wu, X.E. Physiological response of Mirabilis jalapa Linn to lead stress by FTIR spectroscopy. Acta Ecol. Sin. 2011, 31, 6143–6148. [Google Scholar] [CrossRef]
- Wei, Z.L.; Dong, L.; Tian, Z.H. Fourier transform infrared spectrometry study on early stage of cadmium stress in clover leaves. Pak. J. Bot. 2009, 41, 1743–1750. [Google Scholar]
- Traoré, M.; Kaal, J.; Cortizas, A.M. Differentiation between pine woods according to species and growing location using FTIR-ATR. Wood Sci. Technol. 2018, 52, 487–504. [Google Scholar] [CrossRef] [Green Version]
- Rana, R.; Herz, K.; Bruelheide, H.; Dietz, S.; Haider, S.; Jandt, U.; Pena, R. Leaf attenuated total reflection Fourier transform infrared (ATR-FTIR) biochemical profile of grassland plant species related to land-use intensity. Ecol. Indic. 2018, 84, 803–810. [Google Scholar] [CrossRef]
- Yu, P. Molecular chemistry imaging to reveal structural features of various plant feed tissues. J. Struct. Biol. 2005, 150, 81–89. [Google Scholar] [CrossRef]
- Somboon, S.; Kamolmanit, B.; Namanusart, W.; Thammanu, K.; Lawongsa, P. Changes in soil organic carbon composite resulting from long-term application of biochemical contrasting organic residues monitoring by synchrotron-based FTIR microspectroscopy. Biosci. Res. 2018, 15, 2542–2550. [Google Scholar]
- Ashokkumar, R.; Ramaswamy, M. Phytochemical screening by FTIR spectroscopic analysis of leaf extracts of selected Indian medicinal plants. Int. J. Curr. Microbiol. Appl. Sci. 2014, 3, 395–406. [Google Scholar]
- Sankaran, S.; Maja, J.M.; Buchanon, S.; Ehsani, R. Huanglongbing (citrus greening) detection using visible, near infrared and thermal imaging techniques. Sensors 2013, 13, 2117–2130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thumanu, K.; Mathukorn, S.; Phansak, P.; Nontapot, K.; Buensanteai, N. Use of infrared microspectroscopy to determine leaf biochemical composition of cassava in response to Bacillus subtilis CaSUT007. J. Plant Interact. 2015, 10, 270–279. [Google Scholar] [CrossRef] [Green Version]
- Puttaso, P.; Lawongsa, P. Impact of Chronosequence of rubber tree plantation on soil microbial activity. J. Pure Appl. Microbiol. 2018, 12, 1085–1091. [Google Scholar] [CrossRef]
- Allison, L.E. Organic carbon. In Methods of Soil Analysis, Part 2, Chemical and Microbiological Properties; Black, C.A., Ed.; American Society of Agronomy: Madison, WI, USA, 1965; pp. 1367–1378. ISBN 978-0-8911-8374-7. [Google Scholar] [CrossRef]
- Culman, S.W.; Snapp, S.S.; Freeman, M.A.; Schipanski, M.E.; Beniston, J.; Lal, R.; Drinkwater, L.E.; Franzluebbers, A.J.; Glover, J.D.; Grandy, A.T.; et al. Permanganate oxidizable carbon reflects a processed soil fraction that is sensitive to management. Soil Sci. Soc. Am. J. 2012, 76, 494–504. [Google Scholar] [CrossRef] [Green Version]
- Amato, M.; Ladd, J.N. Assay for microbial biomass based on ninhydrin reactive nitrogen in extracts of fumigated soil. Soil. Biol. Biochem. 1988, 20, 107–114. [Google Scholar] [CrossRef]
- Anderson, J.P.E. Soil Respiration. In Methods of Soil Analysis, Part 2, 2nd ed.; Page, A.L., Miller, R.H., Keeney, D.R., Eds.; American Society of Agronomy: Madison, WI, USA, 1982; pp. 837–871. ISBN 978-0-8911-8072-2. [Google Scholar] [CrossRef]
- Kocsis, T.; Kotroczó, Z.; Kardos, L.; Biró, B. Optimization of increasing biochar doses with soil–plant–microbial functioning and nutrient uptake of maize. Environ. Technol. Innov. 2020, 101191. [Google Scholar] [CrossRef]
- Goering, H.K.; Van Soest, P.J. Forage Fiber Analyses (Apparatus, Reagents, Procedures, and Some Applications); United State, Agricultural Research Service: Washington, DC, USA, 1970; pp. 387–598.
- Tatzber, M.; Stemmer, M.; Spiegel, H.; Katzlberger, C.; Haberhauer, G.; Mentler, A.; Gerzabek, M.H. FTIR-spectroscopic characterization of humic acids and humin fractions obtained by advanced NaOH, Na4P2O7, and Na2CO3 extraction procedures. J. Plant Nutr. Soil Sci. 2007, 170, 522–529. [Google Scholar] [CrossRef]
- Parikh, S.J.; Goyne, K.W.; Margenot, A.J.; Mukome, F.N.; Calderón, F.J. Soil chemical insights provided through vibrational spectroscopy. In Advances in Agronomy; Academic Press: New York, NY, USA, 2014; Volume 126, pp. 1–148. [Google Scholar]
- Demyan, M.S.; Rasche, F.; Schütt, M.; Smirnova, N.; Schulz, E.; Cadisch, G. Combining a coupled FTIR-EGA system and in situ DRIFTS for studying soil organic matter in arable soils. Biogeosciences 2013, 10, 2897–2913. [Google Scholar] [CrossRef] [Green Version]
- Egli, M.; Mavris, C.; Mirabella, A.; Giaccai, D. Soil organic matter formation along a chronosequence in the Morteratsch proglacial area (Upper Engadine, Switzerland). Catena 2010, 82, 61–69. [Google Scholar] [CrossRef] [Green Version]
- Spaccini, R.; Piccolo, A. Molecular characterization of compost at increasing stag of maturity. 1. Chemical fractionation and infrared spectroscopy. J. Agric. Food Chem. 2007, 55, 2293–2302. [Google Scholar] [CrossRef]
- Yu, P. Application of advanced synchrotron-based Fourier transform infrared microspectroscopy (SR-FTIR) to animal nutrition and feed science: A novel approach. J. Nutr. 2004, 92, 869–885. [Google Scholar] [CrossRef] [PubMed]
- Skotti, E.; Kountouri, S.; Bouchagier, P.; Tsitsiannis, D.I.; Polissiou, M.; Tarantilis, P.A. FTIR spectroscopic evalution of change in the cellular biochemical composition of the phytopathogenic fugus Alternaria alternate induced by extracts of some Greek medicinal and aromatic plants. Spectrochim. Acta A Mol. Biomol. Spectrosc. 2014, 127, 463–472. [Google Scholar] [CrossRef] [PubMed]
- Kotroczó, Z.; Juhos, K.; Biró, B.; Kocsis, T.; Pabar, S.A.; Fekete, I.; Varga, C. Effect of Detritus Manipulation on Different Organic Matter Decompositions in Temperate Deciduous Forest Soils. Forests 2020, 11, 675. [Google Scholar] [CrossRef]
- Bowden, R.D.; Wurzbacher, S.J.; Washko, S.E.; Wind, L.; Rice, A.M.; Coble, A.E.; Lajtha, K. Long-term Nitrogen Addition Decreases Organic Matter Decomposition and Increases Forest Soil Carbon. Soil Sci. Soc. Am. J. 2019, 83, S82–S95. [Google Scholar] [CrossRef]
- Bailey, R.W.; Connor, H.E. Structural polysaccharides in leaf blades and sheaths in the Arundinoid grass Chionochloa. N. Z. J. Bot. 1972, 10, 533–544. [Google Scholar] [CrossRef] [Green Version]
- Thumanu, K.; Wongchalee, D.; Mathukorn, S.; Piyaporn, P.; Toah Le, T.; Namanusert, W.; Vechklang, K.; Kaewnum, S.; Buensanteai, N. Synchrotron-based FTIR microspectroscopy of chili resistance induced by Bacillus subtilis strain D604 against anthracnose disease. J. Plant Interact. 2017, 12, 255–263. [Google Scholar] [CrossRef] [Green Version]
- Wetzel, D.J.; LeVine, S.M. Biological applications of infrared microspectroscopy. In Infrared and Reman Spectroscopy of Biological Materials; Gremlich, H.U., Yan, B., Eds.; Marcel Dekker: New York, NY, USA, 2000; pp. 101–142. ISBN 978-3-5273-1993-0. [Google Scholar]
- Kenneth, M.D.; Lawrence, C.D. Use of infrared microspectroscopy in plant growth and development. Appl. Spectrosc. Rev. 2005, 40, 301–326. [Google Scholar] [CrossRef]
- Nelson, W.H. Modern Techniques for Rapid Microbiological Analysis, 1st ed.; Wiley-VCH: New York, NY, USA, 1991; ISBN 978-0-4711-8777-6. [Google Scholar]
- McCann, M.C.; Hammouri, M.; Wilson, R.; Belton, P.; Roberts, K. Fourier transform infrared microspectroscopy is a new way to look at plant cell walls. J. Plant Physiol. 1992, 100, 1940–1947. [Google Scholar] [CrossRef] [Green Version]
- Dokken, K.M.; Davis, L.C. Infrared imaging of sunflower and maize root anatomy. J. Agric. Food Chem. 2007, 55, 10517–10530. [Google Scholar] [CrossRef]
- Demyan, M.S.; Rasche, F.; Schulz, E.; Breulmann, M.; Müller, T.; Cadisch, G. Use of specific peaks obtained by diffuse reflectance Fourier transform mid-infrared spectroscopy to study the composition of organic matter in a Haplic Chernozem. Eur. J. Soil Sci. 2012, 63, 189–199. [Google Scholar] [CrossRef]
- Campbell, M.M.; Sederoff, R.R. Variation in lignin content and composition mechanisms of control and implications for the genetic improvement of plants. J. Plant Physiol. 1996, 110, 3–13. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Zhuac, J.; Huangac, R.Z.; Yang, Y. Investigation of cell wall composition related to stem lodging resistance in wheat (Triticum aestivum L.) by FTIR spectroscopy. Plant Signal. Behav. 2012, 7, 856–863. [Google Scholar] [CrossRef] [Green Version]
- Drinkwater, L.E.; Wagoner, P.; Sarrantonio, M. Legume–based cropping systems have reduced carbon and nitrogen losses. Nature 1998, 396, 262–265. [Google Scholar] [CrossRef]
- Hammel, K.E. Fungal degradation of lignin. In Driven by Nature: Plant Litter Quality and Decomposition; Cadisch, G., Giller, K.E., Eds.; CAB International: Wallingford, UK, 1997; pp. 33–46. [Google Scholar]
- Margenot, A.J.; Calderón, F.J.; Bowles, T.M.; Parikh, S.J.; Jackson, L.E. Soil organic matter functional group composition in relation to organic carbon, nitrogen, and phosphorus fractions in organically managed tomato fields. Soil Sci. Soc. Am. J. 2015, 79, 772–782. [Google Scholar] [CrossRef]
- Talbot, J.M.; Yelle, D.J.; Nowick, J.; Treseder, K.K. Litter decay rates are determined by lignin chemistry. Biogeochemistry 2012, 108, 279–295. [Google Scholar] [CrossRef]
- Talbot, J.M.; Treseder, K.K. Interactions among lignin, cellulose, and nitrogen drive litter chemistry-decay relationships. Ecology 2012, 93, 345–354. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tree Ages | SOC (%) | Microbial Respiration (gCO2·m−2·d−1) | MBC (mg·kg−1) | MBN (mg·kg−1) | qCO2 | POXC (mg·kg−1) |
|---|---|---|---|---|---|---|
| 4–5 years | 0.26b | 482.5b | 468.0a | 534.6a | 0.8b | 52.58c |
| 11–12 years | 0.30a | 607.7ab | 243.6b | 250.6b | 2.5a | 114.82b |
| 22–23 years | 0.31a | 685.6a | 260.6b | 453.6a | 2.2a | 169.66a |
| p-value | * | ** | * | * | * | * |
| Tree Age | Cellulose (g·kg−1) | Hemicellulose (g·kg−1) | Lignin (g·kg−1) |
|---|---|---|---|
| 4–5 years | 29.40 | 10.73 | 24.45 |
| 11–12 years | 23.69 | 11.21 | 13.36 |
| 22–23 years | 22.77 | 13.36 | 25.00 |
| Wavenumber (cm−1) | Chemical Composition | Tree Age | ||
|---|---|---|---|---|
| 4–5 Years | 11–12 Years | 22–23 Years | ||
| 1253 | C-O of phenolic groups [32] | - | - | 3.9 |
| 1206 | Phenol vas (C-O-,carboxylic acid v(C-O) [33] | 12.8a | 12.0a | 8.5b |
| 1137 | C-H of aliphatic OH and C-O bands of pholyalcoholic and ether, C-H of aliphatic OH [32,34,35] | 5.3a | 4.6a | 2.9b |
| 1119–1067 | Polysaccharide C-O [32,33] | 57.5b | 32.0c | 70.2a |
| Wavenumber (cm−1) | Chemical Composition | Reference |
|---|---|---|
| 2946–2850 | CH2 asymmetrical and symmetrical stretching | [24] |
| 1737 | C=O ester from lignin and hemicellulose/pectin | [3,24,41,42,43] |
| 1654 | Amide I | [24,41,44] |
| 1515 | C=C aromatic ring from lignin | [45] |
| 1444, 1370 | Symmetric CH2 and CH3 bending from lipids, protein, and lignin | [3] |
| 1278, 1241 | hemicellulose | [46] |
| 1145, 1114, 1060, 1031 | Mainly CO CC of polysaccharides, which are very complex and depend upon contributions from polysaccharides, cellulose, hemicellulose, and pectin | [40,47] |
| Biochemical Quality | SOC | POXC | Bulk Density |
|---|---|---|---|
| Cellulose | −0.74 ** | −0.78 ** | 0.69 * |
| Hemicellulose | 0.79 * | −0.77 * | |
| Lignin | 0.69 ** |
| Wavenumber (cm−1) | Chemical Composition | Soil Properties | Biochemical Quality | |||||
|---|---|---|---|---|---|---|---|---|
| SOC | POXC | MBC | Bulk Density | Cellulose | Hemicellulose | Lignin | ||
| Epidermis | ||||||||
| 2946–2850 | CH2 asymmetrical and symmetrical stretching | 0.68 * | 0.84 * | 0.8 * | −0.70 * | |||
| 1737 | C=O ester from lignin and hemicellulose/pectin | −0.80 * | ||||||
| 1444, 1370 | Symmetric CH2 and CH3 bending from lipids, protein, and lignin | 0.69 * | 0.70 * | |||||
| Mesophyll | ||||||||
| 2946–2850 | CH2 asymmetrical and symmetrical stretching | 0.68 * | 0.84 * | 0.8 * | ||||
| 1654 | Amide I | −0.93 ** | ||||||
| 1515 | C=C aromatic ring from lignin | −0.69 * | −0.70 * | 0.97 ** | ||||
| 1444, 1370 | Symmetric CH2 and CH3 bending from lipids, protein, and lignin | 0.70 * | ||||||
| 1145, 1114, 1060, 1031 | Mainly C-O-C of polysaccharides, which very complex and depend upon contributions from polysaccharides, cellulose, hemicellulose, and pectin | 0.94 ** | −0.90 * | |||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Puttaso, P.; Namanusart, W.; Thumanu, K.; Kamolmanit, B.; Brauman, A.; Lawongsa, P. Assessing the Effect of Rubber (Hevea brasiliensis (Willd. ex A. Juss.) Muell. Arg.) Leaf Chemical Composition on Some Soil Properties of Differently Aged Rubber Tree Plantations. Agronomy 2020, 10, 1871. https://doi.org/10.3390/agronomy10121871
Puttaso P, Namanusart W, Thumanu K, Kamolmanit B, Brauman A, Lawongsa P. Assessing the Effect of Rubber (Hevea brasiliensis (Willd. ex A. Juss.) Muell. Arg.) Leaf Chemical Composition on Some Soil Properties of Differently Aged Rubber Tree Plantations. Agronomy. 2020; 10(12):1871. https://doi.org/10.3390/agronomy10121871
Chicago/Turabian StylePuttaso, Porntip, Weravart Namanusart, Kanjana Thumanu, Bhanudacha Kamolmanit, Alain Brauman, and Phrueksa Lawongsa. 2020. "Assessing the Effect of Rubber (Hevea brasiliensis (Willd. ex A. Juss.) Muell. Arg.) Leaf Chemical Composition on Some Soil Properties of Differently Aged Rubber Tree Plantations" Agronomy 10, no. 12: 1871. https://doi.org/10.3390/agronomy10121871
APA StylePuttaso, P., Namanusart, W., Thumanu, K., Kamolmanit, B., Brauman, A., & Lawongsa, P. (2020). Assessing the Effect of Rubber (Hevea brasiliensis (Willd. ex A. Juss.) Muell. Arg.) Leaf Chemical Composition on Some Soil Properties of Differently Aged Rubber Tree Plantations. Agronomy, 10(12), 1871. https://doi.org/10.3390/agronomy10121871