In Vitro Rooting Response of Yellow-Flowered Magnolia in Relation to the Phenolic Acids Content




Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and In Vitro Culture
2.2. Effects of Auxins on In Vitro Rooting and Acclimatization
2.3. Qualitative and Quantitative Analyses of Phenolic Acids by HPLC Method
2.4. Effects of Exogenous Chlorogenic Acid on Rooting of Magnolia In Vitro
2.5. Analysis of Soluble Sugars
2.6. Statistical Analysis
3. Results
3.1. Rooting
3.2. Phenolic Acids Content
3.3. Effects of Exogenous Chlorogenic Acid on Rooting of Magnolia In Vitro
3.4. Effects of Exogenous Chlorogenic Acid on Soluble Sugars Production
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Camelbeke, K.; Foster, M. The best yellow magnolias. Plantsman 2015, 14, 78–85. [Google Scholar]
- Fare, D.C. Performance of yellow-flowering magnolias in zone 6b. Horttechnology 2017, 27, 291–295. [Google Scholar] [CrossRef] [Green Version]
- Sharma, J.; Knox, G.W.; Ishida, M.L. Adventitious rooting of stem cuttings of yellow-flowered Magnolia cultivars is influenced. by time after budbreak and indole-3-butyric acid. HortScience 2006, 41, 202–206. [Google Scholar] [CrossRef] [Green Version]
- Wojtania, A.; Skrzypek, E.; Marasek-Ciolakowska, A. Soluble sugar, starch and phenolic status during rooting of easy- and difficult-to-root magnolia cultivars. Plant Cell Tissue Organ Cult. 2019, 136, 499–510. [Google Scholar] [CrossRef]
- De Klerk, G.-J.; Guan, H.; Huisman, P.; Marinova, S. Effects of phenolic compounds on adventitious root formation and oxidative decarboxylation of applied indoleacetic acid in Malus ‘Jork 9’. Plant Growth Regul. 2011, 63, 175–185. [Google Scholar] [CrossRef] [Green Version]
- De Klerk, G.J.; Van der Krieken, W.; De Jong, J.C. Review-The formation of adventitious roots: New concepts, new possibilities. Vitr. Cell. Dev. Biol. Plant 1999, 35, 189–199. [Google Scholar] [CrossRef]
- Trobec, M.; Stampar, F.; Veberic, R.; Osterc, G. Fluctuations of different endogenous phenolic compounds and cinnamic acid in the first days of the rooting process of cherry rootstock ‘GiSelA 5’ leafy cuttings. J. Plant Physiol. 2005, 162, 589–597. [Google Scholar] [CrossRef]
- Osterc, G.; Stefancic, M.; Solar, A.; Stampar, F. Phenolic content in cuttings of two clones of hybrid chestnut (Castanea crenata × Castanea sativa) in the first days after cutting severance. Acta Agric. Scand. B Soil Plant Sci. 2008, 58, 162–168. [Google Scholar] [CrossRef]
- Izadi, M.; Shahsavar, A.R.; Mirsoleimani, A. Relation between leaf and stem biochemical constituents and rooting ability of olive cuttings. Int. J. Hortic. Sci. 2016, 3, 231–242. [Google Scholar]
- Singh, H.P.; Kaur, S.; Batish, D.R.; Kohli, R.K. Ferulic acid impairs rhizogenesis and root growth, and alters associated biochemical changes in mung bean (Vigna radiata) hypocotyls. J. Plant Interact. 2014, 9, 267–274. [Google Scholar] [CrossRef]
- Shang, W.; Wang, Z.; He, S.; He, D.; Liu, Y.; Fu, Z. Research on the relationship between phenolic acids and rooting of tree peony (Paeonia suffruticosa) plantlets in vitro. Sci. Hortic. 2017, 224, 53–60. [Google Scholar] [CrossRef]
- Bandurski, R.S.; Cohen, J.D.; Slovin, J.P.; Reinecke, D.M. Auxin biosynthesis and metabolism. In Plant Hormones; Davies, P.J., Ed.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1995; pp. 39–65. [Google Scholar]
- Denaxa, N.-K.; Vemmos, S.N.; Roussos, P.A. The role of endogenous carbohydrates and seasonal variation in rooting ability of cuttings of an easy and a hard to root olive cultivars (Olea europaea L.). Sci. Hortic. 2012, 143, 19–28. [Google Scholar] [CrossRef]
- Otiende, M.A.; Nyabundi, J.O.; Ngamau, K.; Opala, P. Effects of cutting position of rose rootstock cultivars on rooting and its relationship with mineral nutrient content and endogenous carbohydrates. Sci. Hortic. 2017, 225, 204–212. [Google Scholar] [CrossRef]
- Osterc, G.; Stefancic, M.; Solar, A.; Stampar, F. The effect of severance date on rooting ability of chestnut cuttings and associated changes in phenolic content during adventitious root formation. Phyton 2007, 46, 285–294. [Google Scholar]
- Denaxa, N.-K.; Roussos, P.A.; Vemmos, S.N. Assigning a role to the endogenous phenolic compounds on adventitious root formation of olive stem cuttings. J. Plant Growth Regul. 2020, 39, 411–421. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Hura, T.; Dziurka, M.; Hura, K.; Ostrowska, A.; Dziurka, K. Different allocation of carbohydrates and phenolics in dehydrated leaves of triticale. J. Plant Physiol. 2016, 202, 1–9. [Google Scholar] [CrossRef]
- Golebiowska-Pikania, G.; Dziurka, M.; Wasek, I.; Wajdzik, K.; Dyda, M.; Wedzony, M. Changes in phenolic acid abundance involved in low temperature and Microdochium nivale (Samuels and Hallett) cross-tolerance in winter triticale (x Triticosecale Wittmack). Acta Physiol. Plant. 2019, 41, 38. [Google Scholar] [CrossRef] [Green Version]
- Dubois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Parris, J.K.; Touchell, D.H.; Ranney, T.G.; Adelberg, J. Basal salt composition, cytokinins, and phenolic binding agents influence in vitro growth and ex vitro establishment of Magnolia ‘Ann’. HortScience 2012, 47, 1625–1629. [Google Scholar] [CrossRef] [Green Version]
- Sokolov, R.S.; Atanassova, B.Y.; Iakimova, E.T. Physiological response of in vitro cultured Magnolia sp. to nutrient medium composition. J. Hortic. Res. 2014, 22, 49–61. [Google Scholar] [CrossRef] [Green Version]
- Wojtania, A.; Skrzypek, E.; Gabryszewska, E. Effect of cytokinin, sucrose and nitrogen salts concentrations on the growth and development and phenolics content in Magnolia × soulangiana ‘Coates’ shoots in vitro. Acta Sci. Pol. Hortorum Cultus 2015, 14, 51–62. [Google Scholar]
- Wojtania, A.; Skrzypek, E.; Gabryszewska, E. Morphological and biochemical responses to gibberellic acid in Magnolia × ‘Spectrum’ in vitro. Acta Biol. Crac. Ser. Bot. 2016, 58, 103–111. [Google Scholar] [CrossRef]
- Kamenicka, A.; Lanakova, M. Effect of medium composition and type of vessel closure on axillary shoot production of magnolia in vitro. Acta Physiol. Plant. 2000, 22, 129–134. [Google Scholar] [CrossRef]
- Radomir, A.-M. Comparative study on the in vitro multiplication potential of Magnolia stellata and Magnolia x soulangiana species. J. Hortic. For. Biotechnol. 2012, 16, 39–44. [Google Scholar]
- Gercheva, P.; Nacheva, L.; Ivanova, V.; Ibrakhim, O. Rooting of in vitro-raised microcuttings of Magnolia grandiflora L., and Magnolia × soulangiana Soul.-Bod. J. Mt. Agric. Balk. 2011, 4, 854–868. [Google Scholar]
- Podwyszynska, M.; Cieslinska, M. Rooting shoots of apple varieties and their tetraploids obtained by the in vitro technique. Acta Sci. Pol. Hortorum Cultus 2018, 17, 49–62. [Google Scholar] [CrossRef]
- Strader, L.C.; Bartel, B. Transport and metabolism of the endogenous auxin precursor indole-3-butyric acid. Mol. Plant 2011, 4, 477–486. [Google Scholar] [CrossRef] [Green Version]
- Biedermann, I.E. Factors affecting establishment and development of Magnolia hybrids in vitro. Acta Hortic. 1985, 212, 625–629. [Google Scholar] [CrossRef]
- Kamenicka, A. Influence of selected carbohydrates on rhizogenesis of shoots saucer magnolia in vitro. Acta Physiol. Plant. 1998, 20, 425–429. [Google Scholar] [CrossRef]
- Zanardo, D.I.L.; Lima, R.B.; Ferrarese, M.d.L.L.; Bubna, G.A.; Ferrarese-Filho, O. Soybean root growth inhibition and lignification induced by p-coumaric acid. Environ. Exp. Bot. 2009, 66, 25–30. [Google Scholar] [CrossRef]
- Zohra, K.D.; Asma, Z.; Kamel, M.; Helmi, H.; Bechir, E. Changes of phenolic compounds in Carignan merithallus (Vitis vinifera L.) during bud dormancy and end of dormancy phase: Correlation with rhizogenesis. Agric. Sci. 2011, 2, 498–504. [Google Scholar] [CrossRef] [Green Version]
- Franklin, G.; Dias, A.C.P. Chlorogenic acid participates in the regulation of shoot, root and root hair development in Hypericum perforatum . Plant Physiol. Biochem. 2011, 49, 835–842. [Google Scholar] [CrossRef]
- Dixon, R.A.; Paiva, N.L. Stress-induced phenylpropanoid metabolism. Plant Cell 1995, 7, 1085–1097. [Google Scholar] [CrossRef]
- Shiozaki, S.; Makibuchi, M.; Ogata, T. Indole-3-acetic acid, polyamines, and phenols in hardwood cuttings of recalcitrant-to-root wild grapes native to East Asia: Vitis davidii and Vitis kiusiana . J. Bot. 2013, 2013, 819531. [Google Scholar] [CrossRef] [Green Version]
- Warchol, M.; Skrzypek, E.; Kusibab, T.; Dubert, F. Induction of somatic embryogenesis and biochemical characterization of Cordyline australis (G. Forst.) Endl. ’Red Star’ callus. Sci. Hortic. 2015, 192, 338–345. [Google Scholar] [CrossRef]
- Bouza, L.; Sotta, B.; Bonnet, M.; Jacques, M.; Arnaud, Y. Hormone content and meristematic activity of Paeonia suffruticosa ANDR. cv. Madame de Vatry vitroplants during in vitro rooting. Acta Hortic. 1992, 320, 213–216. [Google Scholar] [CrossRef]
- Cheynier, V.; Comte, G.; Davies, K.M.; Lattanzio, V.; Martens, S. Plant phenolics: Recent advances on their biosynthesis, genetics, and ecophysiology. Plant Physiol. Biochem. 2013, 72, 1–20. [Google Scholar] [CrossRef]
- Sharma, E.; Sharma, R.; Borah, P.; Jain, M.; Khurana, J.P. Emerging roles of auxin in abiotic stress responses. In Elucidation of Abiotic Stress Signaling in Plants; Pandey, G.K., Ed.; Springer: New York, NY, USA, 2015; pp. 299–328. [Google Scholar]
- Liu, J.; Sherif, S.M. Hormonal orchestration of bud dormancy cycle in deciduous woody perennials. Front. Plant Sci. 2019, 10, 1136. [Google Scholar] [CrossRef]
- Li, Z.; Zhang, J.; Liu, Y.; Zhao, J.; Fu, J.; Ren, X.; Wang, G.; Wang, J. Exogenous auxin regulates multi-metabolic network and embryo development, controlling seed secondary dormancy and germination in Nicotiana tabacum L. BMC Plant Biol. 2016, 16, 41. [Google Scholar] [CrossRef] [Green Version]
- Shuai, H.; Meng, Y.; Luo, X.; Chen, F.; Zhou, W.; Dai, Y.; Qi, Y.; Du, J.; Yang, F.; Liu, J.; et al. Exogenous auxin represses soybean seed germination through decreasing the gibberellin/abscisic acid (GA/ABA) ratio. Sci. Rep. 2017, 7, 12620. [Google Scholar] [CrossRef] [Green Version]
- Inacio, M.C.; Moraes, R.M.; Mendonca, P.C.; Morel, L.J.; Franca, S.C.; Bertoni, B.W.; Pereira, A.M. Phenolic compounds influence seed dormancy of Palicourea rigida HBK (Rubiaceae), a medicinal plant of the Brazilian Savannah. J. Plant Sci. 2013, 4, 129–133. [Google Scholar] [CrossRef] [Green Version]
- Michailidis, M.; Karagiannis, E.; Tanou, G.; Sarrou, E.; Adamakis, I.-D.; Karamanoli, K.; Martens, S.; Molassiotis, A. Metabolic mechanisms underpinning vegetative bud dormancy release and shoot development in sweet cherry. Environ. Exp. Bot. 2018, 155, 1–11. [Google Scholar] [CrossRef]




{kind=link}
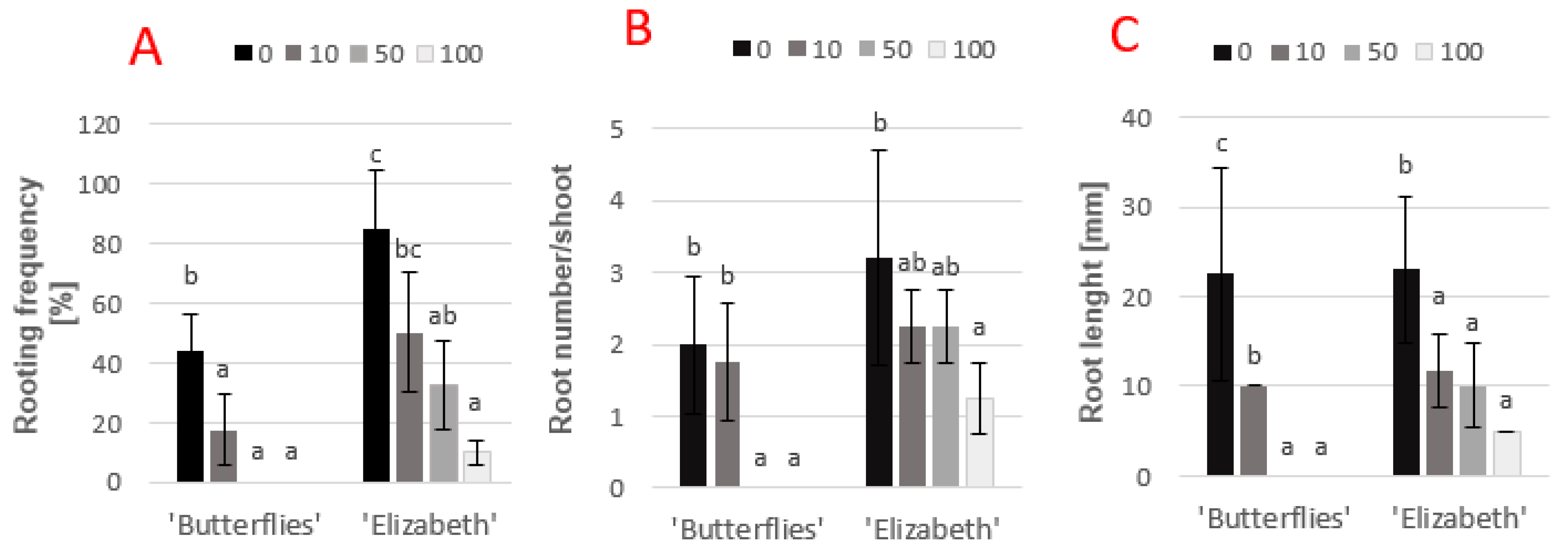{kind=link}
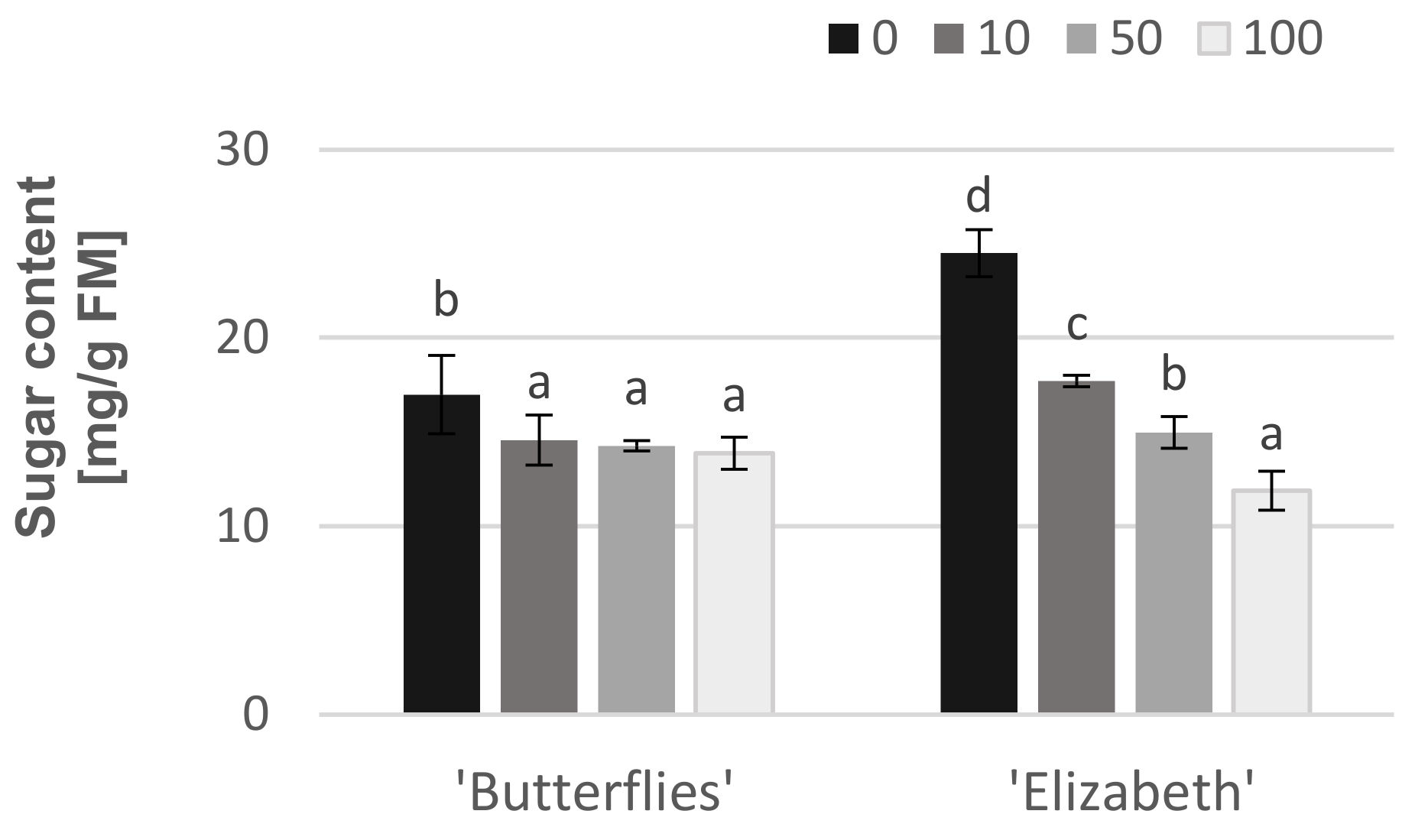{kind=link}
| Cultivar | Auxin Treatment (mg·L−1) | Rooting Frequency (%) | Root Number/Explant | Root Length (mm) | Shoot Length (mm) | Yellow Leaves (%) |
|---|---|---|---|---|---|---|
| ‘Yellow Bird’ | Control | 0.0 a | 0.0 a | 0.0 a | 27.5 a | 9.6 c |
| 6 IBA | 11.6 b | 1.0 b | 55.8 f | 37.1 b | 32.3 d | |
| 4 IBA + 2 NAA | 21.4 c | 1.3 b | 42.5 de | 44.6 c | 41.5 e | |
| ‘Butterflies’ | Control | 0.0 a | 0.0 a | 0.0 a | 35.8 b | 7.3 ab |
| 6 IBA | 13.9 b | 1.3 b | 44.6 e | 37.1 b | 13.0 c | |
| 4 IBA + 2 NAA | 46.1 d | 1.4 b | 33.1 c | 32.1 ab | 7.6 ab | |
| ‘Elizabeth’ | Control | 18.3 bc | 1.1 b | 23.0 b | 56.0 c | 0.0 a |
| 6 IBA | 93.3 e | 3.0 c | 34.2 cd | 61.3 c | 0.0 a | |
| IBA + 2 NAA | 95.8 e | 2.9 c | 26.2 bc | 62.5 c | 0.0 a |
| Phenolic Acids | Auxin | ‘Yellow Bird’ | ‘Butterflies’ | ‘Elizabeth’ |
|---|---|---|---|---|
| benzoic | - | 67.2 ± 30.8 a | 142.4 ± 7.9 b | 137.1 ± 18.8 b |
| + | 153.3 ± 49.4 b | 198.7 ± 23.0 c | 156.9 ± 16.7 b | |
| gallic | - | 31,960 ± 2243 cd | 26,255 ± 1891 a | 24,745 ± 2587 a |
| + | 34,946 ± 2210 d | 31,029 ± 3281 bc | 27,589 ± 3281 ab | |
| 3,4-dihydroxybenzoic | - | 83.5 ± 19.3 a | 108.5 ± 10.7 b | 80.8 ± 11.9 a |
| + | 115.2 ± 4.4 b | 77.2 ± 26.0 a | 113.7 ± 6.0 b | |
| p-hydrobenzoic | - | 13.6 ± 4.9 a | 26.7 ± 6.8 b | 12.3 ± 6.4 a |
| + | 41.8 ± 7.9 c | 41.9 ± 6.8 c | 24.7 ± 6.4 ab | |
| gentisic | - | 174.4 ± 21.3 c | 111.4 ± 12.1 b | 86.4 ± 14.4 a |
| + | 156.5 ± 12.2 c | 125.2 ± 12.5 b | 103.1 ± 16.6 bc | |
| syringic | - | 189.2 ± 66.0 a | 344.2 ± 14.2 b | 293.5 ± 15.7 ab |
| + | 554.3 ± 185.2 c | 772.0 ± 30.1 d | 425.5 ± 124.3 ab | |
| vanillic | - | 71.1 ± 15.1 a | 136.1 ± 7.7 b | 100.0 ± 10.2 ab |
| + | 222.0 ± 36.5 b | 587.5 ± 38.4 e | 318.4 ± 64.7 c | |
| homovanillic | - | 413.7 ± 149.0 b | 560.4 ± 51.6 c | 364.4 ± 54.7 b |
| + | 354.9 ± 70.8 b | 324.9 ± 46.7 b | 155.0 ± 44.2 a | |
| salicylic | - | 84.8 ± 9.7 bc | 102.5 ± 28.9 c | 87.1 ± 12.7 bc |
| + | 51.7 ± 9.9 a | 61.3 ± 16.1 ab | 59.0 ± 26.6 ab | |
| chlorogenic | - | 3181 ± 1263 a | 3445 ± 311 a | 2920 ± 451 a |
| + | 8961 ± 1263 c | 11,485 ± 1285 d | 5008 ± 1142 b | |
| caffeic | - | 2657 ± 702 d | 833.7 ± 169 ab | 1467 ± 277 c |
| + | 1619 ± 72 c | 581.3 ± 144 a | 1312 ± 463 bc | |
| coumaric | - | 64.4 ± 25.5 ab | 100.2 ± 29.9 b | 21.2 ± 8.3 a |
| + | 178.9 ± 46.6 c | 234.2 ± 60.9 d | 53.8 ± 21.9 ab | |
| ferulic | - | 4407 ± 256 d | 2830 ± 80 bc | 3173 ± 149 c |
| + | 2220 ± 311 a | 3264 ± 275 c | 2582 ± 506 ab | |
| sinapic | - | 26.6 ± 4.3 ab | 88.0 ± 17.5 c | 17.7 ± 4.8 a |
| + | 57.9 ± 13.8 bc | 179.4 ± 45.8 d | 29.5 ± 7.8 ab | |
| rosmarinic | - | 22.8 ± 8.0 ab | 33.0 ± 10.5 b | 8.2 ± 1.3 a |
| + | 36.6 ± 7.0 b | 127.9 ± 27.3 c | 19.0 ± 5.5 ab | |
| cinnamic | - | 5.7 ± 2.7 a | 4.2 ± 1.2 a | 5.9 ± 2.6 a |
| + | 4.2 ± 0.7 a | 3.4 ± 0.3 a | 3.3 ± 0.8 a | |
| Total | - | 43,423 ± 4821 | 35,121 ± 2651 | 33,520 ± 3625 |
| + | 49,673 ± 300 | 49,094 ± 5319 | 37,953 ± 5733 |
| Trait/Interaction | Phenolic Acids Content | Chlorogenic Acid Content | Coumaric Acid Content | Rooting Frequency | Root Number/Explant | Root Length | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| F | p | F | p | F | p | F | p | F | p | F | p | |
| Genotype | 132.3 | <0.000 *** | 84.8 | <0.000 *** | 95.6 | <0.000 *** | 118.1 | <0.000 *** | 23.1 | <0.000 *** | 5.27 | <0.000 ** |
| Treatment | 210.0 | <0.000 *** | 43.0 | <0.000 *** | 145.4 | <0.000 *** | 381.5 | <0.000 *** | 70.4 | <0.000 *** | 179.2 | <0.000 *** |
| Genotype*Treatment | 31.14 | <0.000 *** | 327.9 | <0.000 *** | 16.0 | <0.000 *** | 44.6 | <0.000 *** | 0.7 | 0.49 ns | 35.1 | <0.000 *** |
| Feature | Phenolic Acids | Chlorogenic Acid | Coumaric Acid | Rooting Frequency | Root Number | Root Length |
|---|---|---|---|---|---|---|
| Phenolic acids | 1 | |||||
| Chlorogenic acid | 0.552 | 1 | ||||
| Coumaric acid | 0.719 | 0.420 | 1 | |||
| Rooting frequency | 0.037 | 0.116 | 0.021 | 1 | ||
| Root number | 0.021 | −0.053 | 0.040 | 0.759 | 1 | |
| Root length | 0.420 | −0.060 | 0.448 | 0.419 | 0.405 | 1 |
| Genotype | Survival Frequency (%) | Plantlet Length (cm) | Leaf Number/Plantlet | Leaf Necrosis (%) |
|---|---|---|---|---|
| ‘Yellow Bird’ | 24.6 a | 44.0 a | 5.9 a | 54.3 c |
| ‘Butterflies’ | 49.9 b | 65.0 b | 7.7 b | 16.1 b |
| ‘Elizabeth’ | 97.5 c | 135.0 c | 10.3 c | 0.0 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wojtania, A.; Dziurka, M.; Skrzypek, E. In Vitro Rooting Response of Yellow-Flowered Magnolia in Relation to the Phenolic Acids Content. Agronomy 2020, 10, 1880. https://doi.org/10.3390/agronomy10121880
Wojtania A, Dziurka M, Skrzypek E. In Vitro Rooting Response of Yellow-Flowered Magnolia in Relation to the Phenolic Acids Content. Agronomy. 2020; 10(12):1880. https://doi.org/10.3390/agronomy10121880
Chicago/Turabian StyleWojtania, Agnieszka, Michał Dziurka, and Edyta Skrzypek. 2020. "In Vitro Rooting Response of Yellow-Flowered Magnolia in Relation to the Phenolic Acids Content" Agronomy 10, no. 12: 1880. https://doi.org/10.3390/agronomy10121880
APA StyleWojtania, A., Dziurka, M., & Skrzypek, E. (2020). In Vitro Rooting Response of Yellow-Flowered Magnolia in Relation to the Phenolic Acids Content. Agronomy, 10(12), 1880. https://doi.org/10.3390/agronomy10121880