Induced Mutagenesis Enhances Lodging Resistance and Photosynthetic Efficiency of Kodomillet (Paspalum Scrobiculatum)

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Methodology for Generating Mutant Population and Identifying the Desirable Mutants
2.3. Stress Treatment, RNA Isolation, cDNA Synthesis and Quantitative RealTime—Polymerase Chain Reaction (qRT-PCR) Analysis
3. Results and Discussion
3.1. Determination of Lethal Dose (LD50) of Mutagens
3.2. Identification of Photosynthetically Efficient Mutant Lines
3.3. Identification of Non-Lodging Mutant Lines
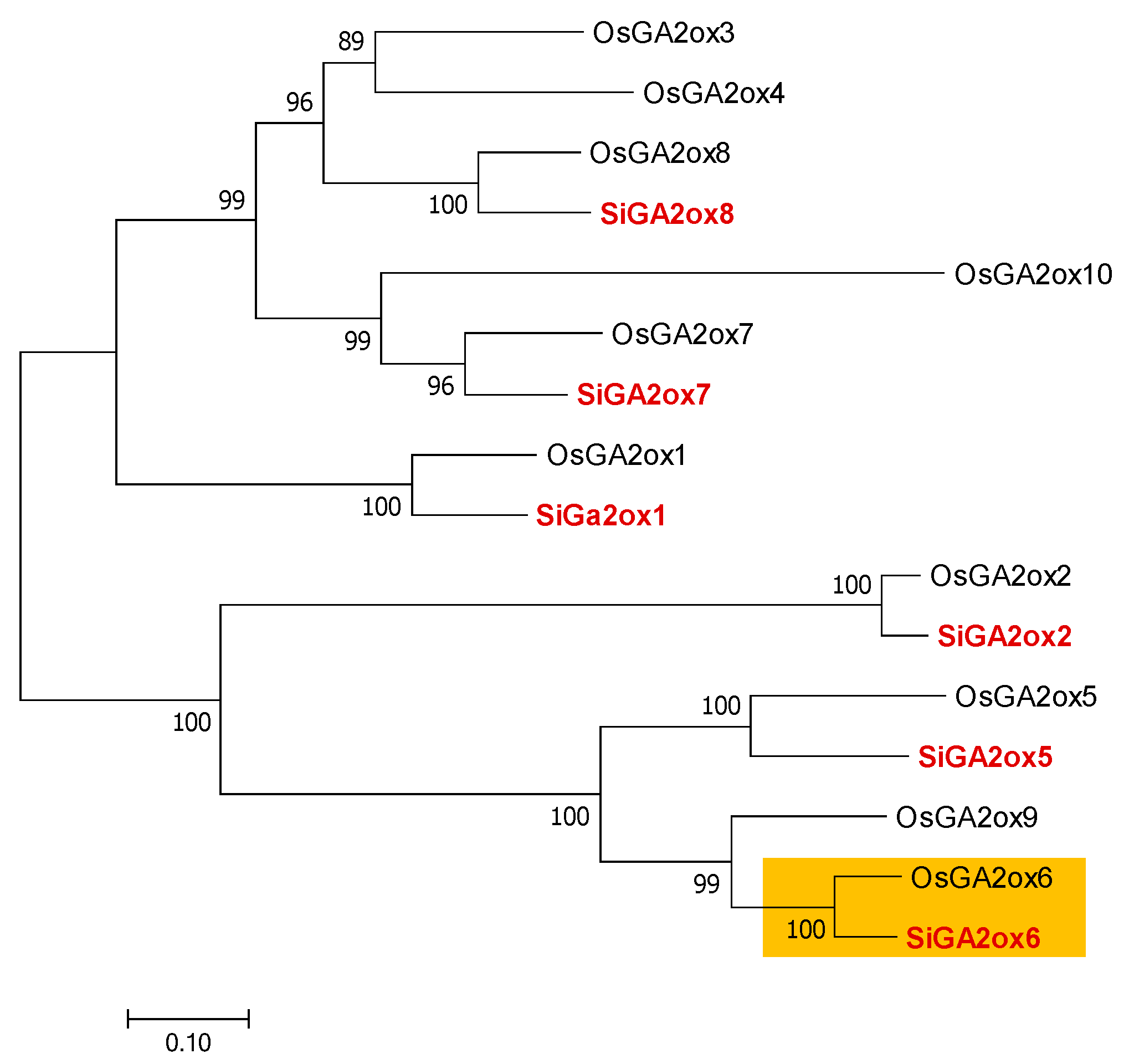
3.4. Molecular Characterization of Mutants through Transcriptomics
| CO 3 | : | Control |
| CO 3-100-7-12 | : | Mutant line with better photosynthetic efficiency |
| CO 3-200-13-4 | : | Mutant line with low photosynthetic efficiency |
| CO 3-100-18-22 | : | Lodged mutant type |
| CO 3-300-7-4 | : | Non-lodged mutant type |
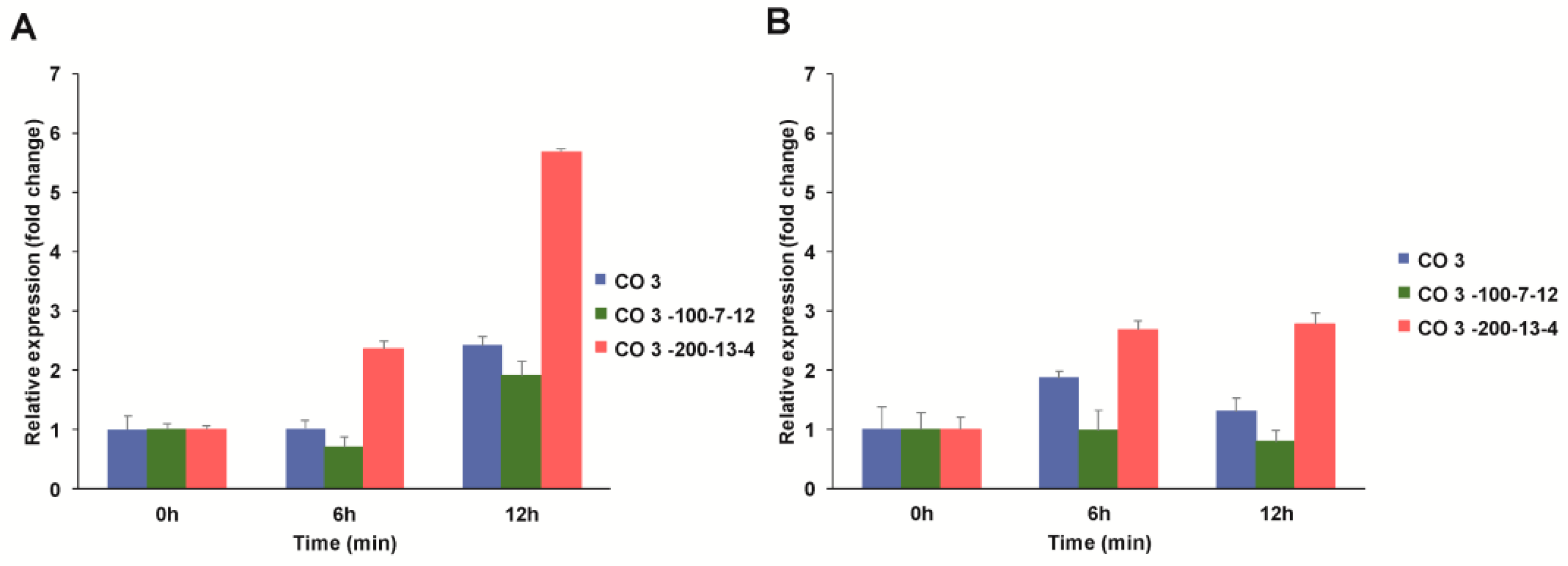
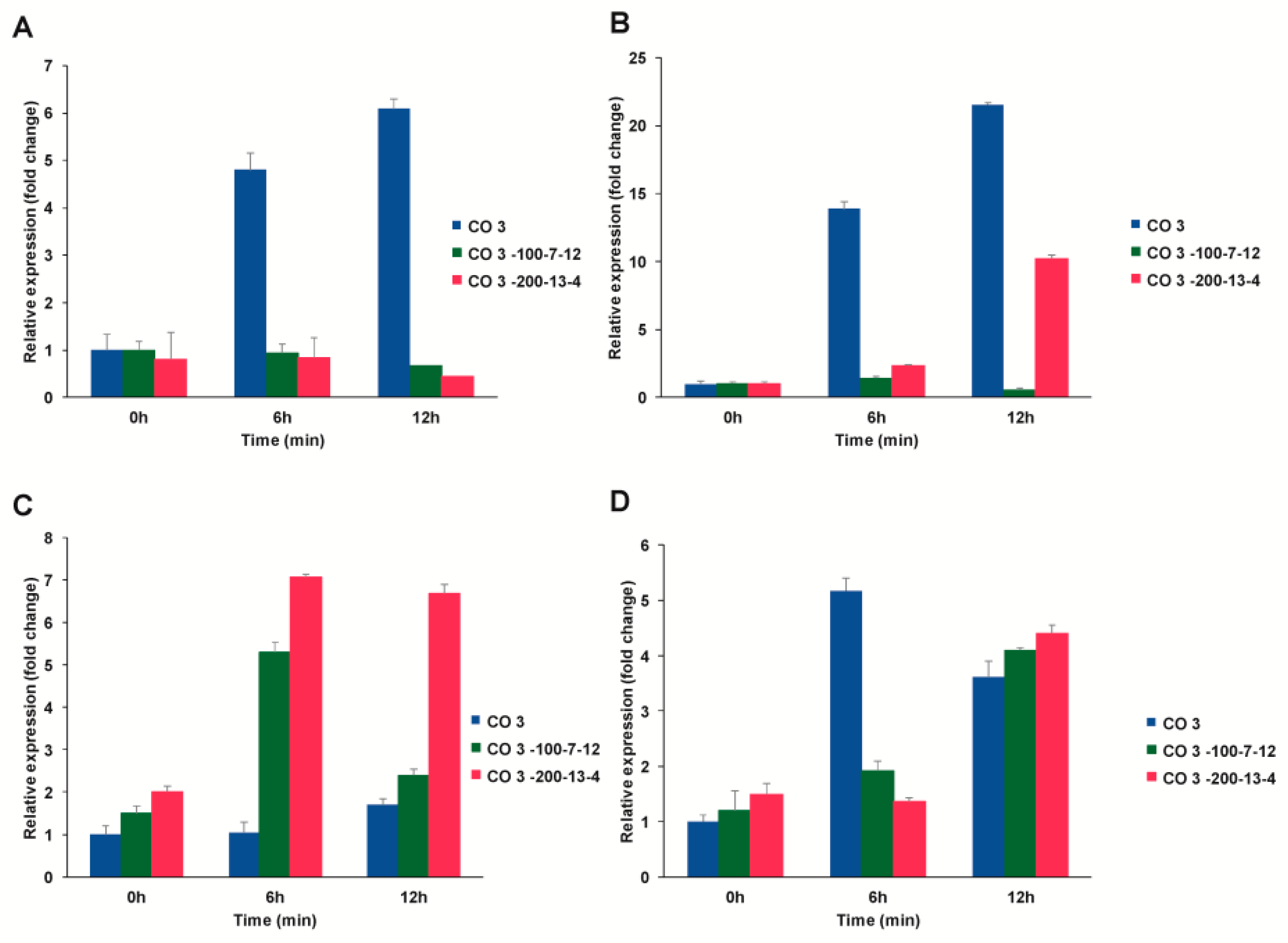
3.4.1. Expression Profiling of C4 Photosynthetic Genes
3.4.1.1. Pyruvate, Orthophosphate Dikinase (PPDK)
3.4.1.2. Nicotinamide Adenine Dinucleotide Phosphate Hydrogen. (NADPH)
3.4.1.3. Nicotinamide Adenine Dinucleotide Phosphate –Malate dehydrogenase (NADP-MDH)
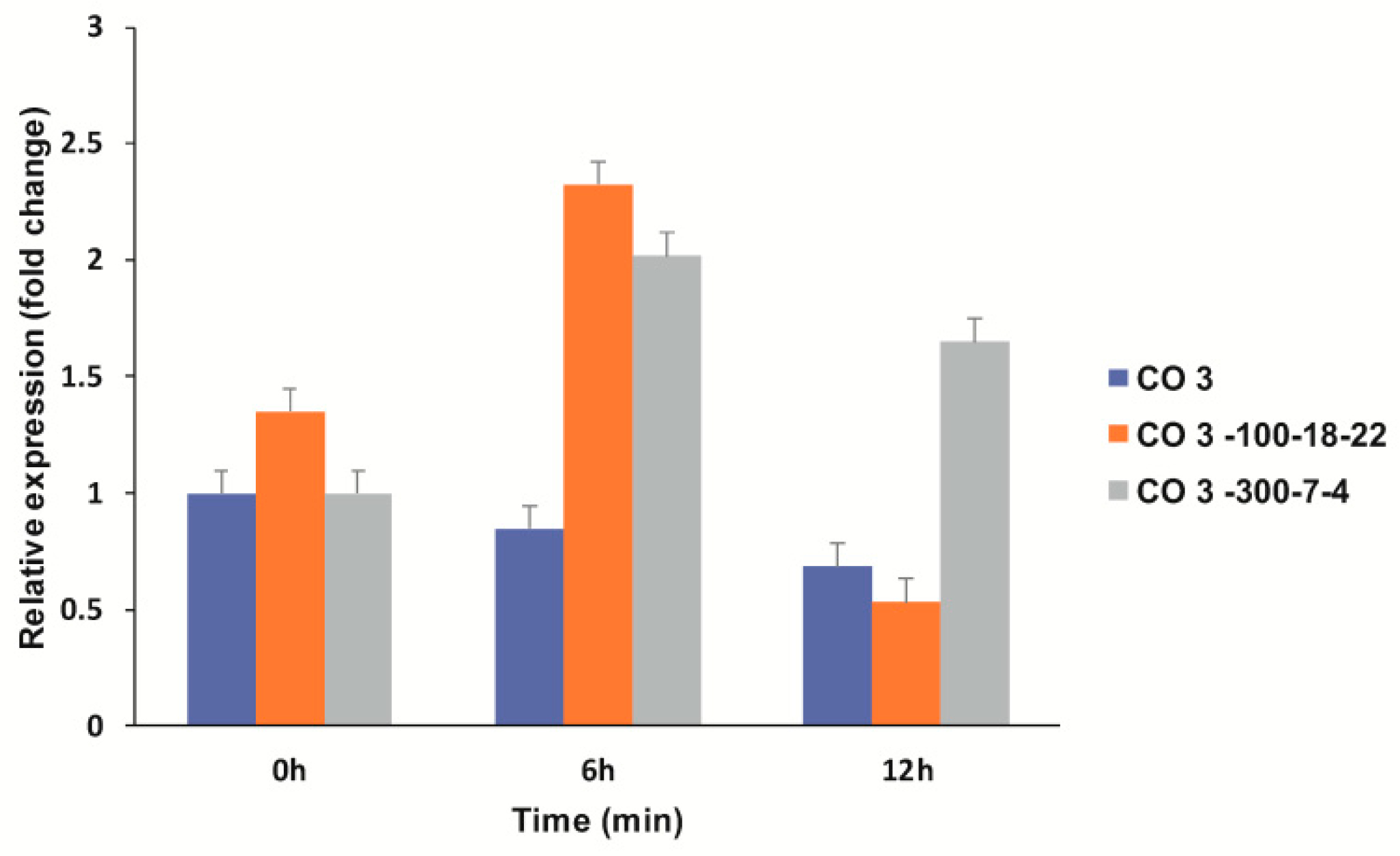
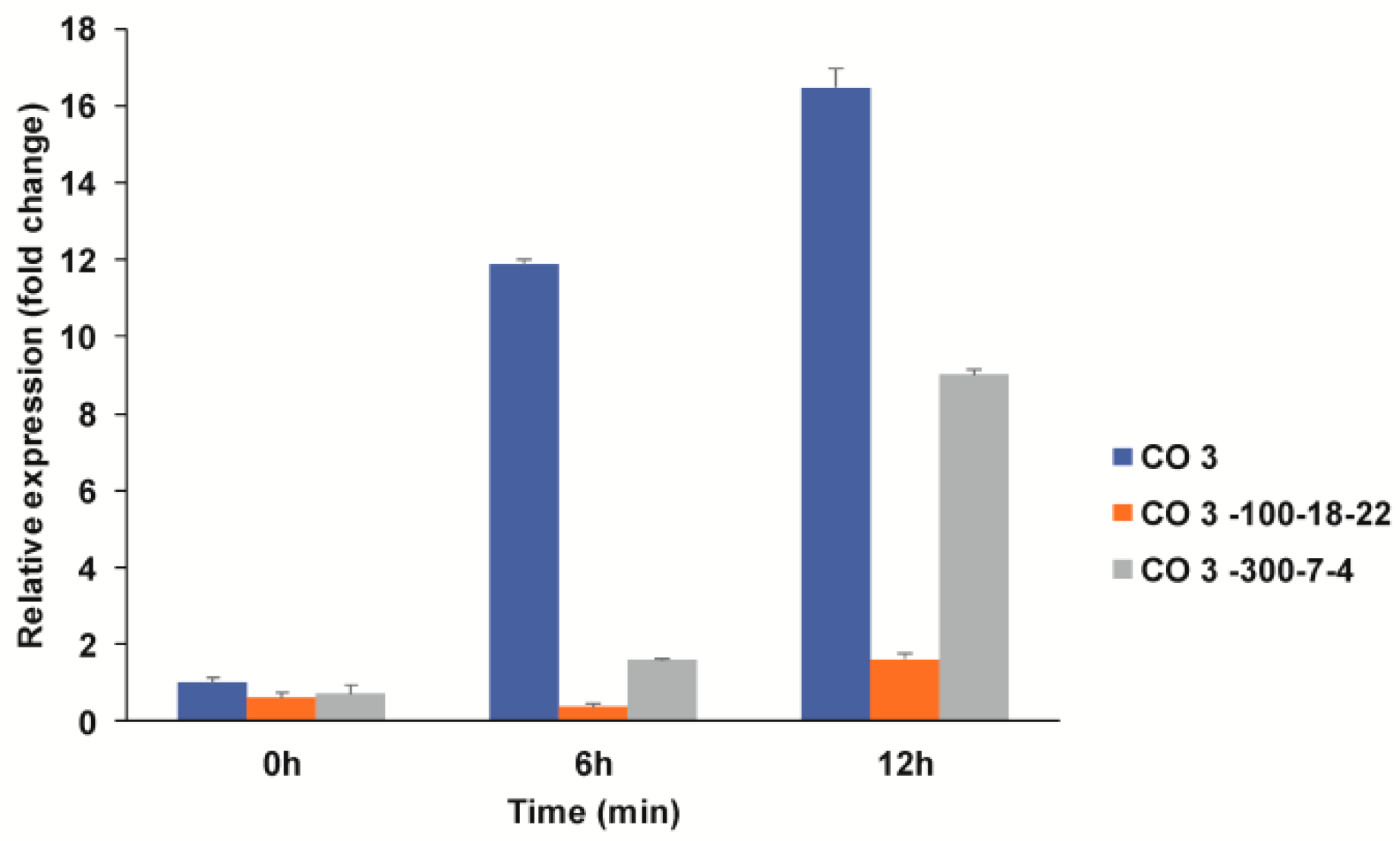
3.4.2. Expression Profiling of Lodging-Related Genes
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Padulosi, S.; Mal, B.; Ravi, S.B.; Gowda, J.; Gowda, K.T.K.; Shanthakumar, G.; Yenagi, N.; Dutta, N. Food security and climate change: Role of plant genetic resources of minor millets. Ind. J. Plant Genet. Res. 2009, 22, 1–16. [Google Scholar]
- De Wet, J.M.J.; Prasada Rao, K.E.; Mengesha, M.H.; Brink, D.E. Diversity in kodo millet, Paspalum scrobiculatum. Econ. Bot. 1983, 37, 159–163. [Google Scholar] [CrossRef] [Green Version]
- Weng, F.; Zhang, W.; Wu, X.; Xu, X.; Ding, Y.; Li, G.; Liu, Z.; Wang, S. Impact of low-temperature, over cast and rainy weather during the reproductive growth stage on lodging resistance of rice. Sci. Rep. 2017, 7, 46596. [Google Scholar] [CrossRef]
- Hegde, P.S.; Chandra, T.S. ESR spectroscopic study reveals higher free radical quenching potential in kodo millet (Paspalum scrobiculatum) compared to other millets. Food Chem. 2005, 92, 177–182. [Google Scholar] [CrossRef]
- Yadava, H.S.; Jain, A.K. Advances in kodo millet research. Directorate of Information and Publications of Agriculture. Indian Counc. Agric. Res. 2006.
- ISTA. International rules for seed testing. Seed Sci. Technol. 1985, 13, 299–355. [Google Scholar]
- Finney, D.J. Probit Analysis, 10th ed.; Cambridge University Press: Cambridge, UK, 1971. [Google Scholar]
- Finney, D.J. Statistical Method in Biological Assay; Charles Griffin & Co.: London, UK, 1978. [Google Scholar]
- PoornimaJency, J.; Ravikesavan, R.; Sumathi, P.; Raveendran, M. Effect of chemical mutagen on germination percentage and seedling parameters in Kodomillet variety CO 3. Int. J. Chem. Std. 2017, 5, 166–169. [Google Scholar]
- Sampson, J. A method of replicating dry or moist surfaces for examination by light microscopy. Nature 1961, 191, 932. [Google Scholar] [CrossRef]
- Kashiwagi, T.; Ishimaru, K. Identification and functional analysis of a locus for improvement of lodging resistance in rice. Plant Physiol. 2004, 134, 676–683. [Google Scholar] [CrossRef] [Green Version]
- Longeman, J.; Schell, J.; Willmitzer, L. Improved method for the isolation of RNA from plant tissues. Anal. Biochem. 1987, 163, 16–20. [Google Scholar] [CrossRef]
- Kumar, K.; Muthamilarasan, M.; Manoj, P. Reference genes for quantitative real-time PCR analysis in the model plant foxtail millet (Setariaitalica L.) subjected to abiotic stress conditions. Plant Cell Tiss. Organ. Cult. 2013, 115, 13–22. [Google Scholar] [CrossRef]
- Smith, H.H. Comparative genetic effects of different physical mutagens in higher plants. In Induced Mutations and Plant Breeding Improvement; IAEA: Vienna, Austria, 1972; pp. 75–93. [Google Scholar]
- PoornimaJency, J.; Ravikesavan, R.; Sumathi, P.; Raveendran, M. Determination of lethal dose and effect of physical mutagen on germination percentage and seedling parameters in kodomillet variety CO 3. Electron. J. Plant Breed. 2016, 7, 1122–1126. [Google Scholar]
- Subramanian, A.; Nirmalakumari, A.; Veerabadhiran, P. Mutagenic Efficiency and Effectiveness in Kodomillet (Paspalum scrobiculatum L.). Madras Agric. J. 2011, 98, 1–3. [Google Scholar]
- Schluter, U.; Muschak, M.; Berger, D.; Altmann, T. Photosynthetic performance of an Arabidopsis mutant with elevated stomatal density (sdd1-1) under different light regimes. J. Exp. Bot. 2003, 54, 867–874. [Google Scholar] [CrossRef]
- Tanaka, Y.; Sugano1, S.S.; Shimada, T.; Nishimura, I.H. Enhancement of leaf photosynthetic capacity through increased stomatal density in Arabidopsis. New Phytol. 2013, 198, 757–764. [Google Scholar] [CrossRef]
- Sreeja, R.; Subramanian, A.; Nirmalakumari, A.; Kannanbapu, J.R. Selection criteria for culm strength in Kodo millet (Paspalum scrobiculatum L.) to suit mechanical harvesting. Electron. J. Plant Breed. 2014, 5, 459–466. [Google Scholar]
- Terashima, K.; Akita, S.; Sakai, N. Eco-physiological characteristics related with lodging tolerance of rice in direct sowing cultivation. Jpn. J. Crop Sci. 1992, 61, 380–387. [Google Scholar] [CrossRef] [Green Version]
- Won, J.G.; Hirahara, Y.; Yoshida, T.; Imabayashi, S. Selection of rice lines using SPGP seedling method for direct seeding. Plant Prod. Sci. 1998, 1, 280–288. [Google Scholar] [CrossRef]
- Zuber, U.; Winzeler, H.; Messmer, M.M.; Keller, B.; Schmid, J.E.; Stamp, P. Morphological traits associated with lodging resistance of spring wheat (Triticumaestivum L.). J. Agron. Crop Sci. 1999, 182, 17–24. [Google Scholar] [CrossRef]
- Cenci, C.A.; Grando, S.; Ceccarelli, S. Culm anatomy in barley (Hordeum vulgare). Can. J. Bot. 1984, 62, 2023–2027. [Google Scholar] [CrossRef]
- Roshan, K.S.; Jaishankar, J.; Muthamilarasan, M.; Shweta, S.; Dangi, A.; Manoj, P. Proteins in C4 model, foxtail millet identifies potential candidates for crop improvement under abiotic stress. Sci. Rep. 2016, 6, 3264. [Google Scholar]
- Pinto, H.; Sharwood, R.E.; Tissue, D.T.; Ghannoum, O. Photosynthesis of C3, C3-C4, and C4 grasses at glacial CO2. J. Exp. Bot. 2014, 65, 3669–3681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Furbank, R.T.; Chitty, J.A.; Jenkins, C.L.D.; Taylor, W.C.; Trevanion, J.S.; Caemmerer, S.V.; Ashton, A.R. Genetic manipulation of key photosynthetic enzymes in the C4 plant Flaveria bidentis. Aust. J. Plant. Physiol. 1997, 24, 477–485. [Google Scholar] [CrossRef]
- Wang, D.A.R.P.; Stephen, J.; Moose, P.; Long, P.S. Cool C4 Photosynthesis: Pyruvate Pi Dikinase Expression and Activity Corresponds to the Exceptional Cold Tolerance of Carbon Assimilation in Miscanthus × giganteus. Plant Physiol. 2008, 148, 557–567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.M.; Xu, W.G.; Hu, L.; Zhang, L.; Li, Y.; Du, X.H. Expression of Maize Gene Encoding C4-Pyruvate Orthophosphate Dikinase(PPDK) and C4-Phosphoenolpyruvate Carboxylase (PEPC) in Transgenic Arabidopsis. Plant Mol. Biol. Rep. 2012, 30, 1367. [Google Scholar] [CrossRef]
- Muthamilarasan, M.; Roshan, K.S.; Singh, N.K.; Prasad, M. C4 photosynthetic enzymes of foxtail millet. Sci. Reps. 2018.
- Takeuchi, Y.; Akagi, H.; Kamasawa, N.; Osumi, M.; Honda, H. Aberrant chloroplasts in transgenic rice plants expressing a high level of maize NADP-dependent malic enzyme. Planta 2000, 211, 265–274. [Google Scholar] [CrossRef]
- Christin, P.A.; Salamin, N.; Kellogg, E.A.; Vicentini, A.; Besnard, G. Integrating phylogeny into studies of C4 variation in the grasses. Plant Physiol. 2009, 149, 82–87. [Google Scholar] [CrossRef] [Green Version]
- Ding, Y.; Ma, Q.H. Characterization of a cytosolic malate dehydrogenase cDNA which encodes an isozyme toward oxaloacetate reduction in wheat. Biochimie 2004, 86, 509–518. [Google Scholar] [CrossRef]
- Beeler, S.; Liu, H.C.; Stadler, M.; Schreier, T.; Eicke, S.; Lue, W.L.; Truernit, E.S.; Zeeman, S.C.; Chen, J.; Kotting, O. Plastidial NAD-Dependent Malate Dehydrogenase is Critical for Embryo Development and Heterotrophic Metabolism in Arabidopsis. Plant Physiol. 2014, 164, 1175–1190. [Google Scholar] [CrossRef] [Green Version]
- Menckhoff, L.; Ehret, N.M.; Buck, F.; Vuleti, M.; Luthje, S. Plasma membrane-associated malate dehydrogenase of maize (Zea mays L.) roots: Native versus recombinant protein. J. Prot. 2013, 80, 66–77. [Google Scholar] [CrossRef] [PubMed]
- Yao, J.; Ma, H.; Zhang, P.; Ren, L.; Yang, X.; Yao, G.; Zhang, P.; Zhou, M. Inheritance of stem strength and its correlations with culm morphological traits in wheat (Triticum aestivum L.). Can. J. Plant Sci. 2011, 91, 1065–1070. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.A.; Li, Q.; Ge, X.Y.; Yang, C.L.; Luo, X.L.; Zhang, A.H.; Xiao, J.L.; Tian, Y.C.; Xia, G.X.; Chen, X.Y.; et al. The mitochondrial malate dehydrogenase 1 gene GhmMDH1 is involved in plant and root growth under phosphorus deficiency deficiency conditions in cotton. Sci. Rep. 2015, 5, 10343. [Google Scholar] [CrossRef] [PubMed]
- Wei, C.; Song, Z.J.; Fang, Z.; Hu, W.U.N. Photosynthetic Features of Transgenic Rice Expressing Sorghum C4 Type NADP-ME. Acta Bot. Sinica 2004, 46, 873–882. [Google Scholar]
- Lo, S.F.; Yang, S.Y.; Chen, K.T.; Hsing, Y.I.; Zeevaart, J.A.; Chen, L.J.; Yu, S.M. A Novel Class of Gibberellin 2-Oxidases Control Semi dwarfism, Tillering, and Root Development in Rice. Plant Cell 2008, 20, 2603–2618. [Google Scholar] [CrossRef] [Green Version]
- Lee, D.J.; Zeevaart, A.D.J. Molecular Cloning of GA 2-Oxidase3 from Spinach and Its Ectopic Expression in Nicotiana sylvestris. Plant Physiol. 2005, 138, 243–254. [Google Scholar] [CrossRef] [Green Version]
- Pearce, S.; Saville, R.; Vaughan, S.P.; Chandler, P.M.; Wilhelm, E.P.; Sparks, C.A.; Kaff, N.A.; Korolev, A.; Boulton, M.I.; Phillips, A.L.; et al. Molecular Characterization of Rht-1 Dwarfing Genes in Hexaploid Wheat. Plant Physiol. 2011, 157, 1820–1831. [Google Scholar] [CrossRef] [Green Version]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Primer Sequence | Number of Bases |
|---|---|---|
| SiPPDK2 | F: GGTCGCAAAGCATGGCCTAA | 20 |
| R: GAAGGCTCCCCACCATGTT | 19 | |
| SiNADP-ME2 | F: TGAGCGCTGTGGTGCAAA | 18 |
| R:GGCAAAGTCCTCAAACTGAATGA | 23 | |
| SiNADP-ME9 | F: AGATTGGGCCCTTCTTATTGGT | 22 |
| R: GTAACGCAGCTCGCTCCATT | 20 | |
| SiNADP-MDH1 | F: GGCGTGACCACCCTAGATGTT | 21 |
| R: TACATTGGCCTTCCCAGCAT | 20 | |
| SiNADP-MDH4 | F: GCAGCAGTACGAGCGATTCA | 20 |
| R: GCCCCGCGTGTTGTTCT | 17 | |
| SiNADP-MDH10 | F: GTGGGAGAGGTTCTTGGACTTG | 22 |
| R: AGCATGCCCACCAATGACA | 19 | |
| SiNADP-MDH11 | F: GGAATGGAGCGAGCTGACTT | 20 |
| R: CCCCTGTTCCGCAAAAATC | 19 | |
| Rht-B | F: ATGAAGCGSGAGTACCAGGA | 20 |
| R:TCTGCGCCACGTCCGCCATGTC | 22 | |
| SiGA2ox6 | F: CGCCCTCATCGTCAA | 15 |
| R: ACGCTCTTGTATCTGTTGTTG | 21 |
| S.No. | Mutants | Stomatal Number | Stomatal Length (µm) | Chlorophyll Index | Flag Leaf Length (cm) | Flag Leaf Breadth (cm) |
|---|---|---|---|---|---|---|
| 1 | CO 3-100-1-5 | 45.00 ± 1.46 | 9.03 ± 0.17 | 38.97 ± 0.91 | 22.77 ± 0.50 | 1.30 ± 0.02 |
| 2 | CO 3-100-7-12 (high efficient) | 79.00 ± 1.46 | 10.27 ± 0.17 | 47.97 ± 0.91 | 28.47 ± 0.50 | 1.33 ± 0.02 |
| 3 | CO 3-200-1-3 | 58.00 ± 1.46 | 7.99 ± 0.12 | 41.37 ± 0.70 | 18.10 ± 0.52 | 1.20 ± 0.01 |
| 4 | CO 3-200-4-1 | 71.00 ± 1.46 | 9.16 ± 0.12 | 35.20 ± 0.70 | 19.57 ± 0.52 | 1.17 ± 0.01 |
| 5 | CO 3-200-14-1 | 56.00 ± 1.46 | 8.85 ± 0.12 | 37.00 ± 0.70 | 20.27 ± 0.52 | 1.30 ± 0.01 |
| 6 | CO3-40.25-12-4 | 58.00 ± 1.33 | 9.90 ± 0.13 | 37.57 ± 0.60 | 32.33 ± 0.62 | 1.30 ± 0.03 |
| 7 | CO 3-40.25-30-2 | 61.00 ± 1.33 | 9.46 ± 0.13 | 41.97 ± 0.60 | 29.10 ± 0.62 | 1.57 ± 0.03 |
| CO 3 200-13-4 (low efficient) | 27.00 ± 1.46 | 9.18 ± 0.12 | 34.10 ± 0.70 | 28.17 ± 0.52 | 1.00 ± 0.01 | |
| Wild type | 55.00 ± 2.03 | 9.34 ± 0.49 | 35.62 ±1.38 | 32.38 ± 0.40 | 1.17 ± 0.01 |
| S.No. | Mutants | Culm Thickness (mm) | Culm Strength (newtons) |
|---|---|---|---|
| 1 | CO 3-100-7-3 | 3.30 ± 0.07 | 34.73 ± 1.62 |
| 2 | CO 3-100-10-5 | 3.47 ± 0.07 | 22.08 ± 1.62 |
| 3 | CO 3-200-16-3 | 3.53 ± 0.05 | 29.73 ± 1.80 |
| 4 | CO 3-200-17-2 | 3.53 ± 0.05 | 24.18 ± 1.80 |
| 5 | CO 3-200-19-4 | 3.90 ± 0.05 | 39.63 ± 1.80 |
| 6 | CO 3-300-2-5 | 2.90 ± 0.24 | 43.65 ± 1.75 |
| 7 | CO 3-300-7-4 (highly stable) | 4.00 ± 0.24 | 39.73 ± 1.75 |
| 8 | CO 3-40.25-13-2 | 3.13 ± 0.05 | 30.27 ± 1.63 |
| 9 | CO 3-40.25-18-5 | 3.10 ± 0.05 | 34.49 ± 1.63 |
| CO3-100-18-22 (unstable) | 0.77 ± 0.07 | 6.88 ± 1.62 | |
| Wild type | 2.53 ± 0.12 | 24.53 ± 1.50 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jency, J.P.; Rajasekaran, R.; Singh, R.K.; Muthurajan, R.; Prabhakaran, J.; Mehanathan, M.; Prasad, M.; Ganesan, J. Induced Mutagenesis Enhances Lodging Resistance and Photosynthetic Efficiency of Kodomillet (Paspalum Scrobiculatum). Agronomy 2020, 10, 227. https://doi.org/10.3390/agronomy10020227
Jency JP, Rajasekaran R, Singh RK, Muthurajan R, Prabhakaran J, Mehanathan M, Prasad M, Ganesan J. Induced Mutagenesis Enhances Lodging Resistance and Photosynthetic Efficiency of Kodomillet (Paspalum Scrobiculatum). Agronomy. 2020; 10(2):227. https://doi.org/10.3390/agronomy10020227
Chicago/Turabian StyleJency, James Poornima, Ravikesavan Rajasekaran, Roshan Kumar Singh, Raveendran Muthurajan, Jeyakumar Prabhakaran, Muthamilarasan Mehanathan, Manoj Prasad, and Jeeva Ganesan. 2020. "Induced Mutagenesis Enhances Lodging Resistance and Photosynthetic Efficiency of Kodomillet (Paspalum Scrobiculatum)" Agronomy 10, no. 2: 227. https://doi.org/10.3390/agronomy10020227
APA StyleJency, J. P., Rajasekaran, R., Singh, R. K., Muthurajan, R., Prabhakaran, J., Mehanathan, M., Prasad, M., & Ganesan, J. (2020). Induced Mutagenesis Enhances Lodging Resistance and Photosynthetic Efficiency of Kodomillet (Paspalum Scrobiculatum). Agronomy, 10(2), 227. https://doi.org/10.3390/agronomy10020227