Morphological Diversity and Bioactive Compounds in Wall Rocket (Diplotaxis erucoides (L.) DC.)





Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Cultivation Conditions
2.2. Morphoagronomic Characterization
2.3. Chemical Analysis
2.4. Statistical Analysis
3. Results
3.1. Plant Habit-Dependent Traits and Whole Plant Characterization
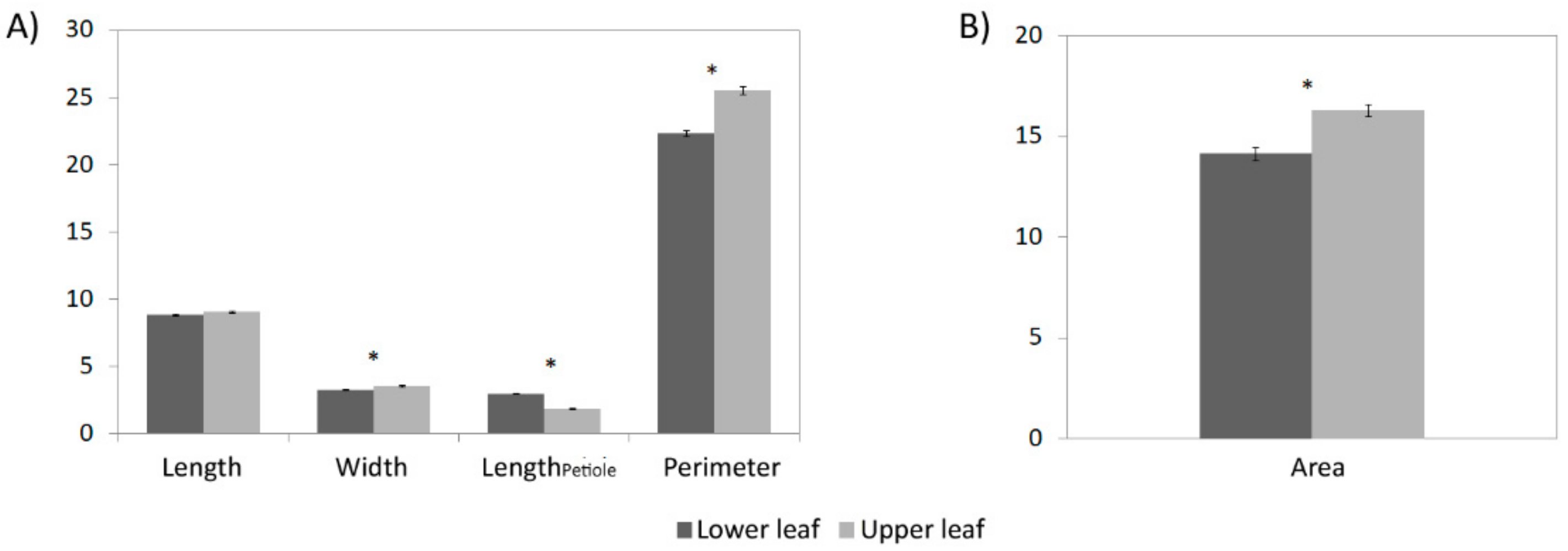
3.2. Leaf Related Traits
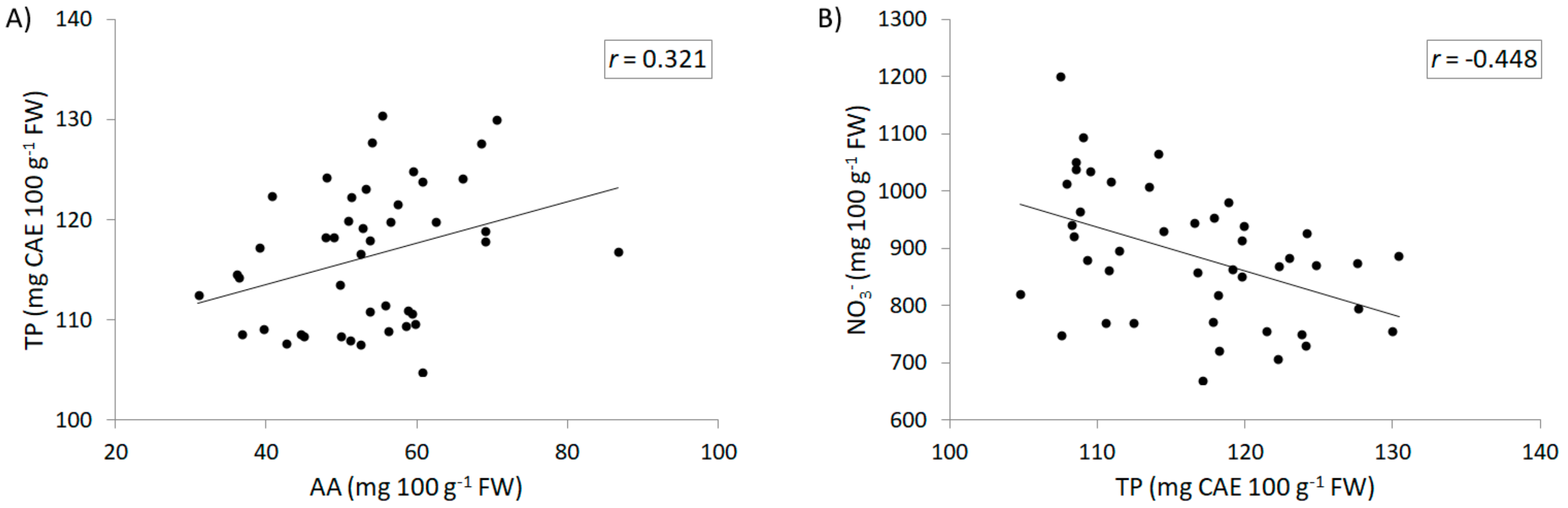
3.3. Chemical Composition
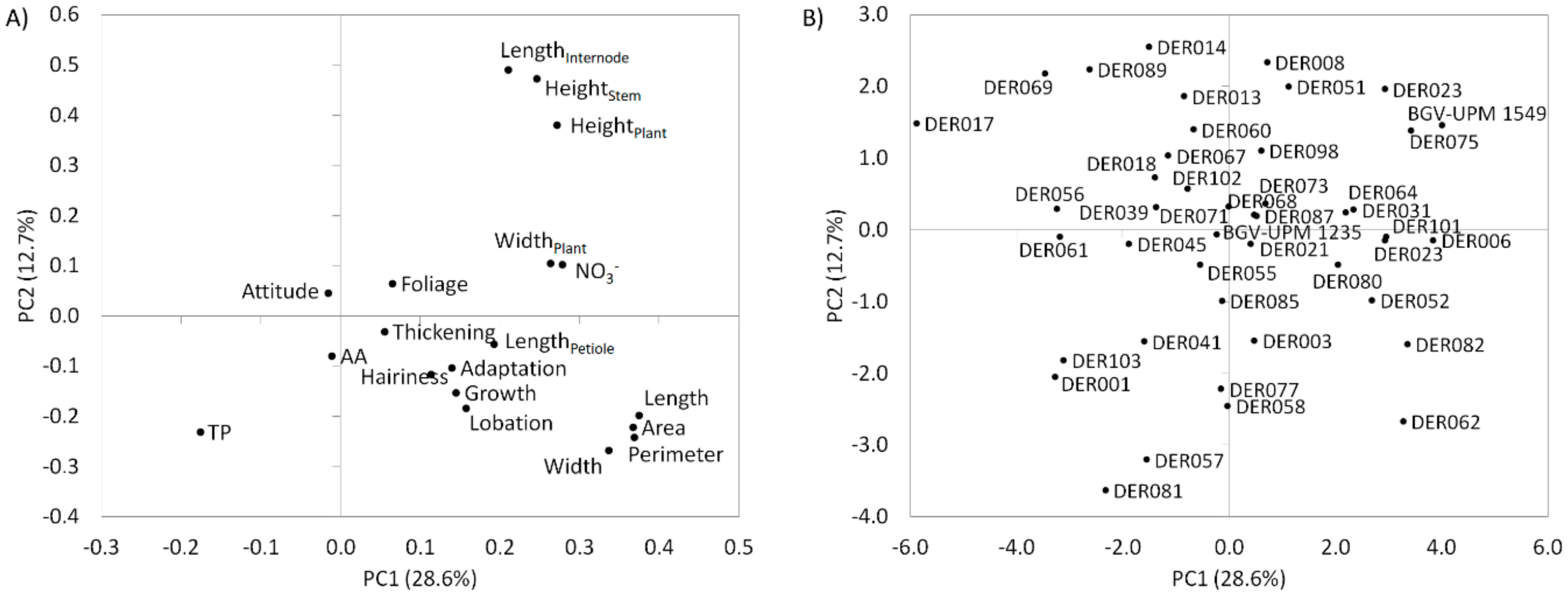
3.4. Principal Component Analysis (PCA)
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Shikov, A.N.; Tsitsilin, A.N.; Pozharitskaya, O.N.; Makarov, V.G.; Heinrich, M. Traditional and current food use of wild plants listed in the Russian Pharmacopoeia. Front. Pharmacol. 2017, 8, 841. [Google Scholar] [CrossRef] [PubMed]
- Shin, T.; Fujikawa, K.; Moe, A.Z.; Uchiyama, H. Traditional knowledge of wild edible plants with special emphasis on medicinal uses in Southern Shan State, Myanmar. J. Ethnobiol. Ethnomed. 2018, 14, 48. [Google Scholar] [CrossRef] [PubMed]
- Łuczaj, Ł.; Pieroni, A.; Tardío, J.; Pardo-de-Santayana, M.; Sõukand, R.; Svanberg, I.; Kalle, R. Wild food plant use in 21st century Europe: The disappearance of old traditions and the search for new cuisines involving wild edibles. Acta Soc. Bot. Pol. 2012, 81, 359–370. [Google Scholar] [CrossRef]
- Pinela, J.; Carvalho, A.M.; Ferreira, I.C.F.R. Wild edible plants: Nutritional and toxicological characteristics, retrieval strategies and importance for today’s society. Food Chem. Toxicol. 2017, 110, 165–188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parada, M.; Carrió, E.; Vallès, J. Ethnobotany of food plants in the Alt Empordà region (Catalonia, Iberian Peninsula). J. Appl. Bot. Food Qual. 2011, 84, 11–25. [Google Scholar]
- Couplan, F. Le régal Végétal. Reconnaître et Cuisiner les Plantes Comestibles; Sang de la Terre: Paris, France, 2015. [Google Scholar]
- Licata, M.; Tuttolomondo, T.; Leto, C.; Virga, G.; Bonsangue, G.; Cammalleri, I.; Gennaro, M.C.; La Bella, S. A survey of wild plant species for food use in Sicily (Italy)—Results of a 3-year study in four Regional Parks. J. Ethnobiol. Ethnomed. 2016, 12, 12. [Google Scholar] [CrossRef] [Green Version]
- Guarrera, P.M.; Savo, V. Wild food plants used in traditional vegetable mixtures in Italy. J. Ethnopharmacol. 2016, 185, 202–234. [Google Scholar] [CrossRef]
- Spadafora, N.D.; Amaro, A.L.; Pereira, M.J.; Müller, C.T.; Pintado, M.; Rogers, H.J. Multi-trait analysis of post-harvest storage in rocket salad (Diplotaxis tenuifolia) links sensorial, volatile and nutritional data. Food Chem. 2016, 211, 114–123. [Google Scholar] [CrossRef] [Green Version]
- Egea-Gilabert, C.; Fernández, J.A.; Migliaro, D.; Martínez-Sánchez, J.J.; Vicente, M.J. Genetic variability in wild vs. cultivated Eruca vesicaria populations as assessed by morphological, agronomical and molecular analyses. Sci. Hortic. 2009, 121, 260–266. [Google Scholar] [CrossRef]
- Disciglio, G.; Tarantino, A.; Frabboni, L.; Gagliardi, A.; Giuliani, M.M.; Tarantino, E.; Gatta, G. Qualitative characterisation of cultivated and wild edible plants: Mineral elements, phenols content and antioxidant capacity. Ital. J. Agron. 2017, 12, 1036. [Google Scholar] [CrossRef] [Green Version]
- Schiattone, M.I.; Viggiani, R.; Di Venere, D.; Sergio, L.; Cantore, V.; Todorovic, M.; Perniola, M.; Candido, V. Impact of irrigation regime and nitrogen rate on yield, quality and water use efficiency of wild rocket under greenhouse conditions. Sci. Hortic. 2018, 229, 182–192. [Google Scholar] [CrossRef]
- Bondonno, C.P.; Blekkenhorst, L.C.; Liu, A.H.; Bondonno, N.P.; Ward, N.C.; Croft, K.D.; Hodgson, J.M. Vegetable-derived bioactive nitrate and cardiovascular health. Mol. Asp. Med. 2018, 61, 83–91. [Google Scholar] [CrossRef] [PubMed]
- Lundberg, J.O.; Carlström, M.; Weitzberg, E. Metabolic effects of dietary nitrate in health and disease. Cell Metab. 2018, 28, 9–22. [Google Scholar] [CrossRef] [PubMed]
- Herraiz, F.J.; Vilanova, S.; Andújar, I.; Torrent, D.; Plazas, M.; Gramazio, P.; Prohens, J. Morphological and molecular characterization of local varieties, modern cultivars and wild relatives of an emerging vegetable crop, the pepino (Solanum muricatum), provides insight into its diversity, relationships and breeding history. Euphytica 2015, 206, 301–318. [Google Scholar] [CrossRef]
- BGV-UPM. Colecciones. Available online: http://www.bancodegermoplasma.upm.es/colecciones.html. (accessed on 24 April 2019).
- Taranto, F.; Francese, G.; Di Dato, F.; D’Alessandro, A.; Greco, B.; Sanajà, V.O.; Pentangelo, A.; Mennella, G.; Tripodi, P. Leaf metabolic, genetic, and morphophysiological profiles of cultivated and wild rocket salad (Eruca and Diplotaxis spp.). J. Agric. Food Chem. 2016, 64, 5824–5836. [Google Scholar] [CrossRef]
- Bell, L.; Methven, L.; Signore, A.; Oruna-Concha, M.J.; Wagstaff, C. Analysis of seven salad rocket (Eruca sativa) accessions: The relationships between sensory attributes and volatile and non-volatile compounds. Food Chem. 2017, 218, 181–191. [Google Scholar] [CrossRef] [Green Version]
- Herraiz, F.J.; Vilanova, S.; Plazas, M.; Gramazio, P.; Andújar, I.; Rodríguez-Burruezo, A.; Fita, A.; Anderson, G.J.; Prohens, J. Phenological growth stages of pepino (Solanum muricatum) according to the BBCH scale. Sci. Hortic. 2015, 183, 1–7. [Google Scholar] [CrossRef]
- IPGRI. Descriptors for Rocket (Eruca spp.); International Plant Genetic Resources Institute: Rome, Italy, 1999. [Google Scholar]
- Guijarro-Real, C.; Adalid-Martínez, A.M.; Gregori-Montaner, A.; Prohens, J.; Rodríguez-Burruezo, A.; Fita, A. Factors affecting germination of Diplotaxis erucoides and their effect on selected quality properties of the germinated products. Sci. Hortic. 2020, 261, 109013. [Google Scholar] [CrossRef]
- Rodríguez, G.R.; Mouseenko, J.B.; Robbins, M.D.; Morejón, N.H.; Francis, D.M.; van der Knaap, E. Tomato Analyzer: A useful software application to collect accurate and detailed morphological and colorimetric data from two-dimensional objects. J. Vis. Exp. 2010, 37, e1856. [Google Scholar] [CrossRef] [Green Version]
- Singleton, V.; Rossi, J. Colorimetry of total phenolics with phosphomolybdic phosphotungstic acid reagents. Am. J. Enol. Vitic. 1965, 16, 144–158. [Google Scholar]
- Guijarro-Real, C.; Prohens, J.; Rodriguez-Burruezo, A.; Adalid-Martínez, A.M.; López-Gresa, M.P.; Fita, A. Wild edible fool’s watercress, a potential crop with high nutraceutical properties. PeerJ 2019, 7, e6296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Egea-Gilabert, C.; Ruiz-Hernández, M.V.; Parra, M.Á.; Fernández, J.A. Characterization of purslane (Portulaca oleracea L.) accessions: Suitability as ready-to-eat product. Sci. Hortic. 2014, 172, 73–81. [Google Scholar] [CrossRef]
- Rodríguez-Burruezo, A.; Prohens, J.; Nuez, F. Genetic analysis of quantitative traits in pepino (Solanum muricatum) in two growing seasons. J. Am. Soc. Hortic. Sci. 2002, 127, 271–278. [Google Scholar] [CrossRef] [Green Version]
- Clewer, A.G.; Scarisbrick, D.H. Practical Statistics and Experimental Design for Plant and Crop Science; John Wiley & Sons Ltd.: West Sussex, UK, 2001. [Google Scholar]
- Wrickle, G.; Weber, W.E. Quantitative Genetics and Selection in Plant Breeding; De Gruyter Online: Berlin, Germany, 1986. [Google Scholar]
- Metsalu, T.; Vilo, J. ClustVis: A web tool for visualizing clustering of multivariate data using Principal Component Analysis and heatmap. Nucleic Acids Res. 2015, 43, W566–W570. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Laborde, J.B. Diplotaxis DC. In Flora Ibérica: Plantas Vasculares de la Península Ibérica e Islas Baleares; Nieto, G., Ed.; Real Jardín Botánico, CSIC: Madrid, Spain, 1990; pp. 346–362. [Google Scholar]
- Prohens, J.; Gramazio, P.; Plazas, M.; Dempewolf, H.; Kilian, B.; Díez, M.J.; Fita, A.; Herraiz, F.J.; Rodríguez-Burruezo, A.; Soler, S.; et al. Introgressiomics: A new approach for using crop wild relatives in breeding for adaptation to climate change. Euphytica 2017, 213, 158. [Google Scholar] [CrossRef]
- Mousavizadeh, S.J.; Hassandokht, M.R.; Kashi, A. Multivariate analysis of edible Asparagus species in Iran by morphological characters. Euphytica 2015, 206, 445–457. [Google Scholar] [CrossRef]
- D’Antuono, L.F.; Elementi, S.; Neri, R. Glucosinolates in Diplotaxis and Eruca leaves: Diversity, taxonomic relations and applied aspects. Phytochemistry 2008, 69, 187–199. [Google Scholar] [CrossRef]
- Di Gioia, F.; Avato, P.; Serio, F.; Argentieri, M.P. Glucosinolate profile of Eruca sativa, Diplotaxis tenuifolia and Diplotaxis erucoides grown in soil and soilless systems. J. Food Compos. Anal. 2018, 69, 197–204. [Google Scholar] [CrossRef]
- Colonna, E.; Rouphael, Y.; Barbieri, G.; De Pascale, S. Nutritional quality of ten leafy vegetables harvested at two light intensities. Food Chem. 2016, 199, 702–710. [Google Scholar] [CrossRef]
- Salvatore, S.; Pellegrini, N.; Brenna, O.V.; Del Rio, D.; Frasca, G.; Brighenti, F.; Tumino, R. Antioxidant characterization of some sicilian edible wild greens. J. Agric. Food Chem. 2005, 53, 9465–9471. [Google Scholar] [CrossRef]
- Bennett, R.N.; Rosa, E.A.S.; Mellon, F.A.; Kroon, P.A. Ontogenic profiling of glucosinolates, flavonoids, and other secondary metabolites in Eruca sativa (salad rocket), Diplotaxis erucoides (wall rocket), Diplotaxis tenuifolia (wild rocket), and Bunias orientalis (Turkish rocket). J. Agric. Food Chem. 2006, 54, 4005–4015. [Google Scholar] [CrossRef] [PubMed]
- Francisco, M.; Velasco, P.; Moreno, D.A.; García-Viguera, C.; Cartea, M.E. Cooking methods of Brassica rapa affect the preservation of glucosinolates, phenolics and vitamin C. Food Res. Int. 2010, 43, 1455–1463. [Google Scholar] [CrossRef] [Green Version]
- Bell, L.; Oloyede, O.O.; Lignou, S.; Wagstaff, C.; Methven, L. Taste and flavor perceptions of glucosinolates, isothiocyanates, and related compounds. Mol. Nutr. Food Res. 2018, 62, 1700990. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bianco, V.V.; Santamaria, P.; Elia, A. Nutritional value and nitrate content in edible wild species used in Southern Italy. Acta Hortic. 1998, 476, 71–90. [Google Scholar] [CrossRef]
- Tang, L.; Luo, W.; Tian, S.; He, Z.; Stoffella, P.J.; Yang, X. Genotypic differences in cadmium and nitrate co-accumulation among the Chinese cabbage genotypes under field conditions. Sci. Hortic. 2016, 201, 92–100. [Google Scholar] [CrossRef]
- Bahadoran, Z.; Mirmiran, P.; Jeddi, S.; Azizi, F.; Ghasemi, A.; Hadaegh, F. Nitrate and nitrite content of vegetables, fruits, grains, legumes, dairy products, meats and processed meats. J. Food Compos. Anal. 2016, 51, 93–105. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Descriptora | Code | Typee | Scale/Units |
|---|---|---|---|
| Plant habit-dependent traits | |||
| Plant growth rate | Growth | Qual ord | 3 = slow; 7 = fast |
| Adaptation levelb | Adaptation | Qual ord | 3 = low; 7 = high |
| Leaf growth attitude | Attitude | Qual categ | 1 = semi-prostrate; 2 = horizontal; 3 = semi-erect |
| Whole plant traits | |||
| Size-related traits | |||
| Plant height | HeightPlant | Quantit | cm |
| Plant width | WidthPlant | Quantit | cm |
| Stem height | HeightStem | Quantit | cm |
| Internode length | LengthInternode | Quantit | cm |
| Descriptive traits | |||
| Stem thickening | Thickening | Qual ord | 3 = thin; 7 = thick |
| Stem color | ColorStem | Qual categ | 1 = light green; 2 = green, 3 = dark green; 4 = red/purple green; 5 = red/purple |
| Stem hairiness | Hairiness | Qual ord | 3 = sparse; 7 = dense |
| Foliagec | Foliage | Quantit | |
| Leaf traits | |||
| Size-related traits | |||
| Leaf lengthd | Length | Quantit | cm |
| Leaf width | Width | Quantit | cm |
| Petiole length | LengthPetiole | Quantit | cm |
| Leaf perimeterd | Perimeter | Quantit | cm |
| Leaf aread | Area | Quantit | cm2 |
| Descriptive traits | |||
| Leaf margin shape | Margin | Qual categ | 1 = entire; 2 = crenate; 3 = dentate |
| Leaf blade shape | Shape | Qual categ | 1 = orbicular; 2 = elliptic; 3 = obovate; 4 = spatulate |
| Leaf apex shape | Apex | Qual categ | 1 = acute; 2 = rounded; 3 = broadly rounded |
| Leaf lobation intensity | Lobation | Qual ord | 0 = absent; 5 = deep lobation |
| Petiole and midvein color | ColorPetiole | Qual categ | 1 = white; 2 = light green; 3 = green; 4 = purple; 5 = red |
| Sum of squares (%) | |||||
| Descriptora | Mean | Range | Population | Residual | H2 |
| HeightPlant (cm) | 8.92 | (6.48–10.74) | 41.6*** | 58.4 | 0.28 |
| WidthPlant (cm) | 13.33 | (10.56–16.60) | 26.6* | 73.4 | 0.09 |
| HeightStem (cm) | 3.99 | (2.38–6.30) | 47.5*** | 52.5 | 0.35 |
| LengthInternode (cm) | 1.04 | (0.56–1.56) | 42.3*** | 57.7 | 0.28 |
| Foliage (number of leaves) | 8.33 | (7.40–9.40) | 30.1** | 69.9 | 0.13 |
| HeightPlant | HeightStem | LengthInternode | |
|---|---|---|---|
| WidthPlanta | 0.431** | 0.399** | 0.303* |
| HeightPlant | 0.707*** | 0.672*** | |
| HeightStem | 0.926*** |
| Descriptora | Lower Leaf | Upper Leaf |
|---|---|---|
| Shape | 1: 15.0%b | 1: 2.3%a |
| - | 2: 9.5% | |
| 3: 83.2%ns | 3: 76.8%ns | |
| 4: 1.8%a | 4: 11.4%b | |
| Apex | 1: 2.3%a | 1: 29.6%b |
| 2: 87.7%b | 2: 62.7%a | |
| 3: 10.0%ns | 3: 7.7%ns | |
| Margin | 1: 5.5% | - |
| 1.5: 47.7%b | 1.5: 0.9%a | |
| 2: 45.4%b | 2: 30.9%a | |
| 2.5: 1.4%a | 2.5: 50.9%b | |
| - | 3: 17.3% | |
| Lobation | 0: 99.6%b | 0: 6.8%a |
| 1: 0.4%a | 1: 25.9%b | |
| - | 2: 46.8% | |
| - | 3: 20.5% |
| Sum of squares (%) | |||||
| Descriptora | Mean | Range | Population | Residual | H2 |
| Length (cm) | 9.05 | (7.27–10.47) | 32.4*** | 67.56 | 0.16 |
| Width (cm) | 3.56 | (2.71–4.37) | 43.0*** | 57.0 | 0.30 |
| Perimeter (cm) | 25.42 | (20.52–32.02) | 43.4*** | 56.6 | 0.30 |
| Area (cm2) | 16.37 | (10.68–24.17) | 43.2*** | 56.8 | 0.30 |
| LengthPetiole (cm) | 1.84 | (0.75–2.95) | 29.7* | 70.4 | 0.13 |
| LengthPetiole | Width | Perimeter | Area | |
|---|---|---|---|---|
| Lengtha | 0.700*** | 0.696*** | 0.941*** | 0.800*** |
| LengthPetiole | 0.309*** | 0.554*** | 0.371*** | |
| Width | 0.759*** | 0.922*** | ||
| Perimeter | 0.784*** |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guijarro-Real, C.; Prohens, J.; Rodríguez-Burruezo, A.; Fita, A. Morphological Diversity and Bioactive Compounds in Wall Rocket (Diplotaxis erucoides (L.) DC.). Agronomy 2020, 10, 306. https://doi.org/10.3390/agronomy10020306
Guijarro-Real C, Prohens J, Rodríguez-Burruezo A, Fita A. Morphological Diversity and Bioactive Compounds in Wall Rocket (Diplotaxis erucoides (L.) DC.). Agronomy. 2020; 10(2):306. https://doi.org/10.3390/agronomy10020306
Chicago/Turabian StyleGuijarro-Real, Carla, Jaime Prohens, Adrián Rodríguez-Burruezo, and Ana Fita. 2020. "Morphological Diversity and Bioactive Compounds in Wall Rocket (Diplotaxis erucoides (L.) DC.)" Agronomy 10, no. 2: 306. https://doi.org/10.3390/agronomy10020306
APA StyleGuijarro-Real, C., Prohens, J., Rodríguez-Burruezo, A., & Fita, A. (2020). Morphological Diversity and Bioactive Compounds in Wall Rocket (Diplotaxis erucoides (L.) DC.). Agronomy, 10(2), 306. https://doi.org/10.3390/agronomy10020306