1. Introduction
Winter wheat (
Triticum aestivum L. em Thell.) is the primary annual crop grown in the US Southern Great Plains (SGP). For example, winter wheat was planted on 1.82 million hectares of land in Oklahoma in 2017, producing 2.68 million tons of grain [
1]. Winter wheat is planted between September and October in the SGP, depending on the intended usage as pasture for grazing, dual-purpose (fall through winter grazing plus end of season grain harvest), or grain crop. In general, winter wheat cultivated as a forage crop is planted early (September), and later if planted for grain production.
Regardless of crop usage, a 3–4 month period of summer fallow exists in continuous monocultures of winter wheat, to conserve limited and sporadic amounts of precipitation as soil moisture to support winter wheat [
2]. However, legumes could be grown during the summer to serve as sources of green N for winter wheat [
3]. The use of annual legumes as cover crops during summer represents one of a series of systems of intensified production [
3,
4,
5] that may improve efficiencies in use of available soil water in the SGP [
6], increase soil carbon (C), suppress weed populations, and reduce nutrient runoff or leaching [
7].
In the SGP, winter wheat is primarily cultivated with conventional tillage, though different systems of reduced tillage are increasing in use to conserve soil moisture, increase soil aggregation, and decrease erosion [
8,
9]. As tillage influences soil physical, chemical, biological and environmental conditions, emissions of CO
2 and N
2O from soils can be influenced by tillage systems [
10,
11]. In general, systems of reduced tillage contribute to decreased soil emissions of CO
2 and increased soil organic carbon (SOC) over the long term [
12]. Tillage operations induce large peaks of soil respiration due to increased soil aeration and increased contact of crop residues with soil [
13]. In contrast, no-till systems avoid such tillage-induced emissions and delay decomposition of residues, but can have higher rates of soil respiration for longer periods due to increased availability of carbon substrates and soil moisture [
14].
Increased soil efflux of N
2O and CO
2 are expected after inclusion of legume-based cover crops in cropping systems applied to winter wheat, due to increased soil pools of C and N. However, the dynamics and magnitude of these fluxes largely depend on quality and quantity of biomass provided by the cover crop at termination [
15]. Additionally, environmental conditions in the soil, such as temperature and moisture, control C and N mineralization and thereby CO
2 and N
2O fluxes from decomposing green manures [
16]. Legume-based cover crops grown during the summer period of monocultures of winter wheat in the SGP are generally terminated 1–2 months prior to planting of winter wheat, to save soil moisture which is the critical factor for winter wheat production in the region [
17]. Additionally, the rates of N fixation by legumes decrease after flowering, so delayed termination may not add significant amounts of N inputs to the system. Alternatively, early-terminated legumes with less structural components can mineralize rapidly after termination, and the mineralized nutrients may be more efficiently transferred to winter wheat. However, if proper synchronization between N mineralization from green manures and uptake by the recipient crop is not achieved, yields of winter wheat may decrease and emissions of N
2O increase outside the growth period of winter wheat [
18].
Although interest for conservation agricultural practices such as no-till and legume green manures is increasing in the US SGP, there is limited information on influence of these managements on fluxes of CO2 and N2O from winter wheat cropping system in this region. Therefore, in this study, we tested the influences of tillage system, rates of applied N fertilizers and cowpea as a legume green manure on canopy and soil fluxes of CO2 and soil fluxes of N2O during a growing period of winter wheat cultivated in the US SGP. Fluxes of CO2 and N2O were monitored in a side-by-side small-plot experiment using closed chamber methods. The working hypotheses were: (i) fluxes of CO2 and N2O from winter wheat cultivation would be lower from no-till than from conventional tillage; (ii) Fluxes of CO2 and N2O would increase with increasing rates of inorganic N fertilizer; and (iii) Fluxes of CO2 and N2O would be greater from plots cultivated with cowpea as a cover crop than unfertilized control plots.
2. Materials and Methods
2.1. Study Site and Soil Properties
The study was conducted at USDA-ARS Grazinglands Research Laboratory at two sites near El Reno, OK, USA. The sites were defined as components of upland and bottomland areas of the North Canadian River drainage basin [
19]. The predominant soil type in upland site (35°32′45″ N, 98°00′44″ W; 421 m elevation) was a Norge silt loam (fine-silty, mixed, thermic, Udic Paleustolls) which is moderately well drained with 1%–3% slope. The topsoil (0–15 cm) had average total organic carbon (TOC) of 1.39%, total N content of 0.11%, and resulting C/N of 12.6. The predominant soil type in bottomland site (35°34′21″ N, 98°02′12″ W; 411 m elevation) was Dale silt loam (fine-silty, mixed, thermic, Pachic Haplustolls) with 0%–1% slope (USDA-NRCS, 1999). At the bottomland site, the average TOC of the topsoil 1.31%, while total N content of 0.10% and the C/N ratio was 13.1.
2.2. Experimental Design
The experiment was conducted as a split-plot in a completely randomized design, on a site basis. Nitrogen (inorganic and organic) treatments were the main plots which included two rates of inorganic fertilizers (45 and 90 kg N ha−1) applied at planting of winter wheat as dry urea, an annual summer legume green manure (cowpea; Vigna unguiculata L.) as a N source, and an unfertilized control. These main plots were split into subplots of two tillage systems; no-till (NT) and conventional tillage (CT). The CT system applied in this study was undertaken to mimic best management practices applicable to conventional tillage, including: fewest operations possible to prepare soil for planting and weed control; leave residues and roughness on soil surface to minimize erosion. Thus, there were a total of 8 treatment combinations (tillage-N source) with 3 replicated plots (4 m × 10 m) per combination on both sites.
2.3. Agronomic Management
The field experiment was initiated in 2011 as a long-term (>10 years) study, to define the effects of different green N crops on winter wheat-based systems (plant biomass, soil nutrients, water balance) of production. The current experiment on fluxes of CO2 and N2O was conducted during the 2016–2017 growing season (October through May) of winter wheat, which was oriented from the start of the summer period of (June) 2016, to the wheat harvest in the following May 2017. In 2016, wheat was harvested on 10 June. The CT plots were tilled (disked once, rototilled once) on 13 June. Cowpea seeds (cv. Red Ripper) inoculated with a Bradyrhizobium strain were planted on 15 June at 2.5 cm depth with sowing rate of 35 kg ha−1. Cowpea was fertilized with 100 kg ha−1 di-ammonium phosphate (DAP; 18% N, 46% P2O5, 0% K2O) on 17 June. DAP fertilizer was used as the P source, as P-only fertilizer was not available to producers in the regions. Therefore, 18 kg inorganic N ha–1 were applied to the cowpea plots.
Weeds in NT plots were controlled with spraying Roundup® herbicide as a 1% glyphosate (N-(phosphonomethyl) glycine) solution at planting (16 June 2016) and termination of cowpea (11 August 2016). Cowpea cultivated under the NT system was terminated on 17 August by shredding biomass and spraying the stubble. Cowpea cultivated under CT system was terminated two days later by disking (once) and roto-tilling (once). Prior to termination, total aboveground biomass yield of cowpea was determined by drying biomass harvested from 0.4 m × 0.4 m quadrats from each plot. The biomass was dried at 60 °C to constant weight in a forced-draft oven. Biomass N concentration was determined by a flash combustion method (Model VarioMacro; Elementar Americas, Inc., Mt. Laurel, NJ, USA). Amount of N per hectare in aboveground cowpea biomass was calculated as a product of dry biomass yield and N concentrations. The 45-N and 90-N plots were fertilized on 21 September and winter wheat (cv. Jagger) was sown in all plots on 22 September 2016. Winter wheat was harvested on 31 May 2017.
Crop management applied to the experimental plots in the previous years (2011–2015) followed similar timing and application rates as in the current study year. All weed control application to the NT plots was undertaken with herbicide. Tillage operations on CT plots were conducted only for planting of wheat and cowpea, and other applications required for weed control were largely herbicide applications. On CT plots, weeds during the fallow periods were primarily controlled with herbicide, though tillage was used in cases where weed populations were large, or had achieved stages of maturity where control with herbicides was less effective. Although the CT system used mechanical disturbance of soil to prepare seedbeds or control weeds, all tillage operations were restricted to the upper 0.20 m of the soil profile. Crop residues and surface roughness were present on the soil surface throughout the fallow periods during summer, which are common requirements for best management practices for CT systems.
2.4. Measurement and Calculation of N2O Fluxes
Fluxes of N2O were measured using a closed chamber method. A PVC collar (0.65 × 0.65 m) was inserted to 0.10 m depth in all plots (n = 24 for both sites) on 27 September (five days after planting of wheat). The collars remained intact throughout the measurement periods. Fluxes were generally measured at two-week intervals until the harvest of winter wheat.
The collars had a 0.04 m wide outer rim that was aboveground and parallel to the soil surface to support the top chamber used for flux measurement. Fluxes were measured by placing an opaque (white color) chamber (0.70 × 0.70 × 0.41 m3) on the collars. Chamber headspace air was continuously mixed by a small battery-driven fan. Concentrations of N2O and CO2 was determined by a portable Fourier transform infrared (FTIR) based analyzer (DX4040; Gasmet Technology Oy, Helsinki, Finland) that was connected to the chamber through 3 mm inlet and outlet tubing. The chamber was enclosed for 8 min during each measurement and concentrations were recorded at 40 s intervals.
Fluxes were calculated by linear regression using the MATLAB
® (MathWorks, Inc., Natick, MA, USA) routine of Kutzbach et al. [
20]. The first few records after chamber enclosure were discarded as dead-band based on visual inspection of the CO
2 concentrations recorded simultaneously with N
2O concentrations. Fluxes of N
2O were assumed to be zero when probability of linear regression was insignificant (
p > 0.05).
2.5. Measurements of Canopy and Soil CO2 Fluxes
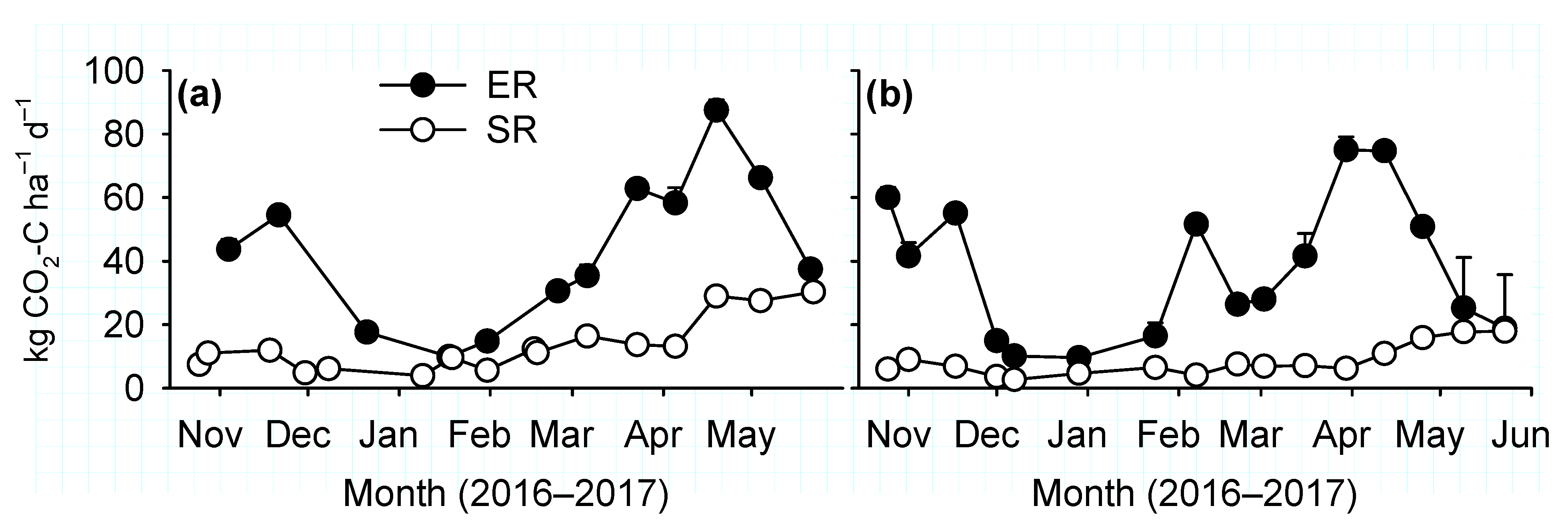
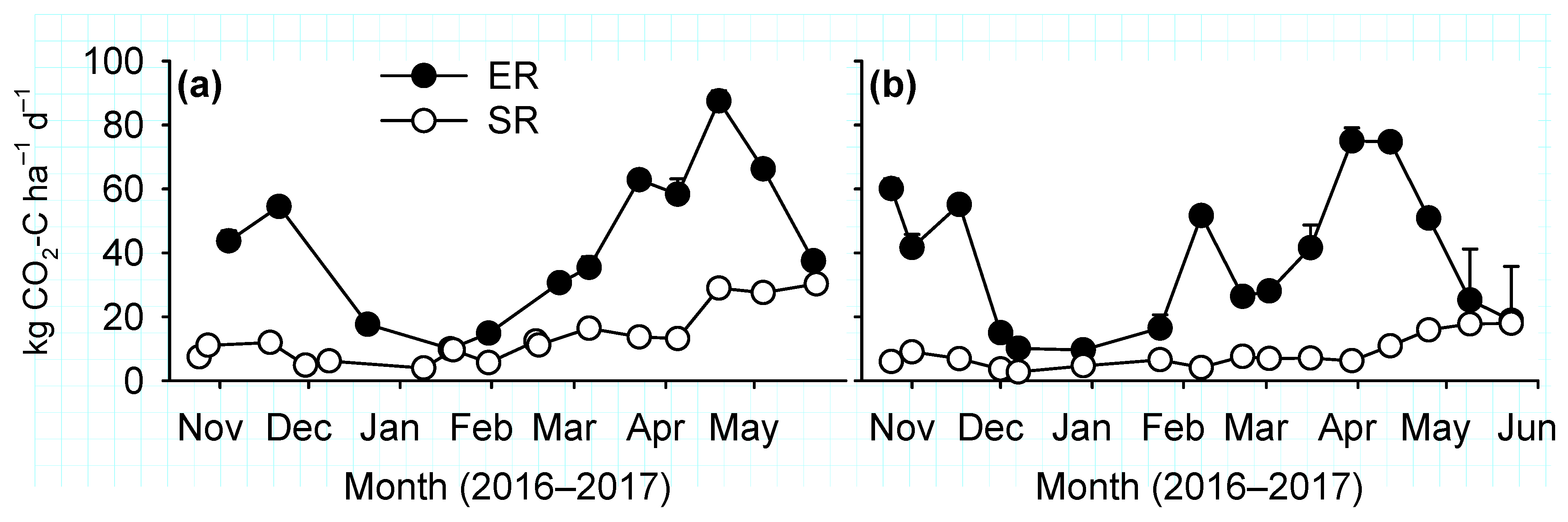
Canopy fluxes of CO2 were measured using a closed transparent Plexiglas chamber with similar dimension as the opaque chamber used to measure N2O flux. The chamber included sensors for temperature and photosynthetically active radiation (PAR), and a fan to mix headspace air. Concentrations of CO2 and H2O was determined by a portable infrared gas analyser (EGM-5; PP Systems, Amesbury, MA, USA) that was connected to the chamber through 3 mm inlet and outlet tubing. The gas analyser recorded all the measurements at 1 s intervals. On each measurement date, fluxes were measured consecutively at all collars under both ambient light and darkened conditions. At first, net ecosystem exchange (NEE) of CO2 was measured at ambient light for 1 min with the fully transparent chamber. Then the chamber was covered with a white cloth to block 100% PAR for ecosystem respiration (ER) flux measurements for 1 min after plant adaptation to dark conditions for 1 min.
Soil respiration (SR) fluxes were measured using a plant and root exclusion method [
21] on cylindrical PVC cores (diameter, 0.10 m; height, 0.30 m) inserted to 0.28 m soil depth. A core was inserted in all plots at about 1 m distance from the collar installed for measurements of canopy CO
2 and N
2O fluxes. The cores were inserted between two rows of winter wheat after germination. Growth of weeds was rarely seen inside the cores, and those encountered were removed by hand before SR measurements. Fluxes were measured using an EGM-5 gas analyzer connected to a cylindrical (height, 0.15 m; diameter, 0.10 m) chamber (model SRC-2; PP Systems). During the flux measurements, the chamber was enclosed for 60 seconds and CO
2 concentrations were recorded at 1 s intervals with the first 10 s of measurements discarded as a dead-band.
Fluxes of CO
2 were measured at midday between 9:30 and 13:00. Measurements were generally taken at regular intervals of two weeks. For measurements of canopy CO
2 fluxes, a Plexiglas chamber extension with similar dimensions as the top chamber was used when the height of crops exceeded chamber height. Fluxes of CO
2 were calculated by linear regression using the MATLAB
® routine by Kutzbach et al. [
20]. The first 10 s of recordings after chamber deployment were discarded as a dead-band.
2.6. Measurements of Environmental Variables
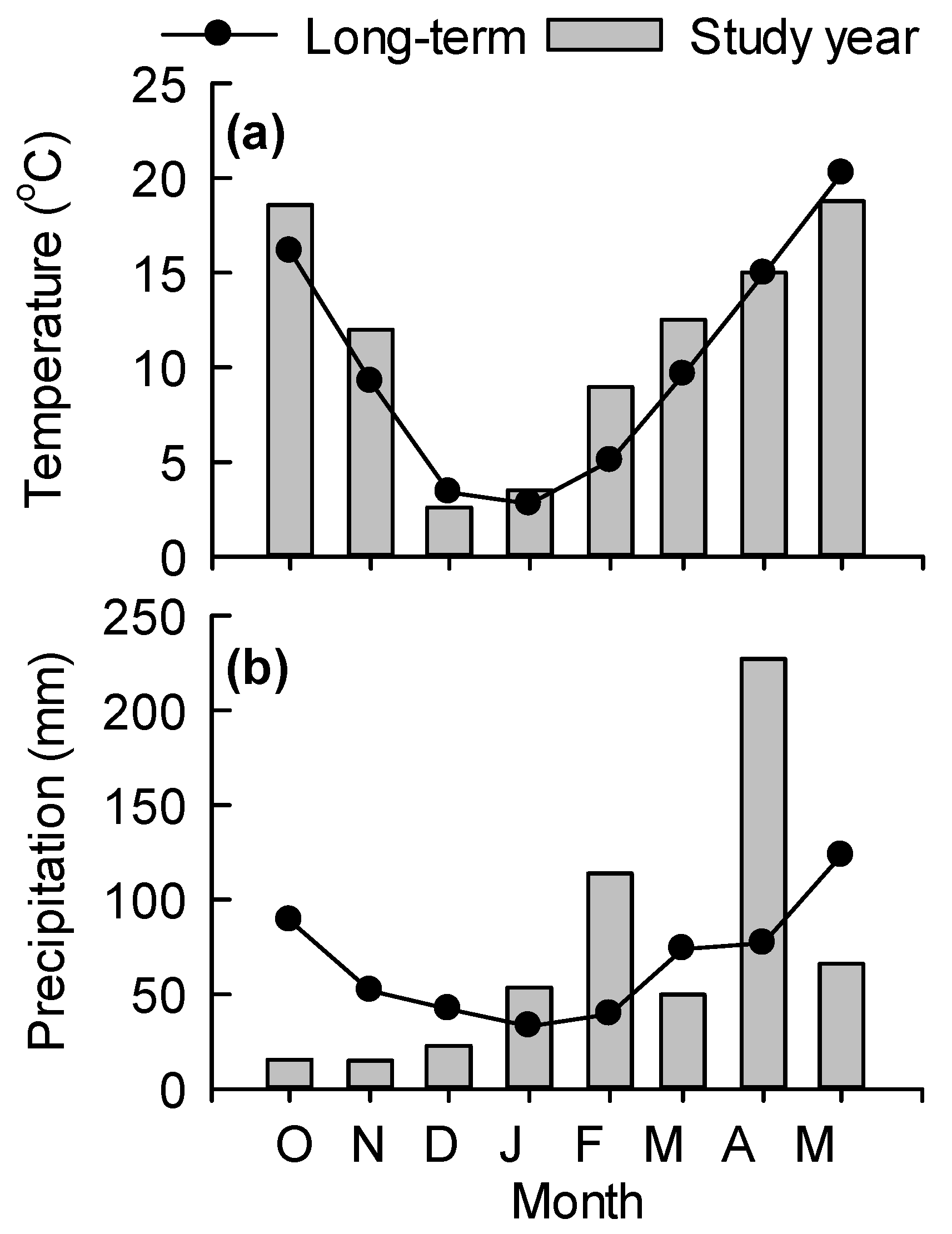
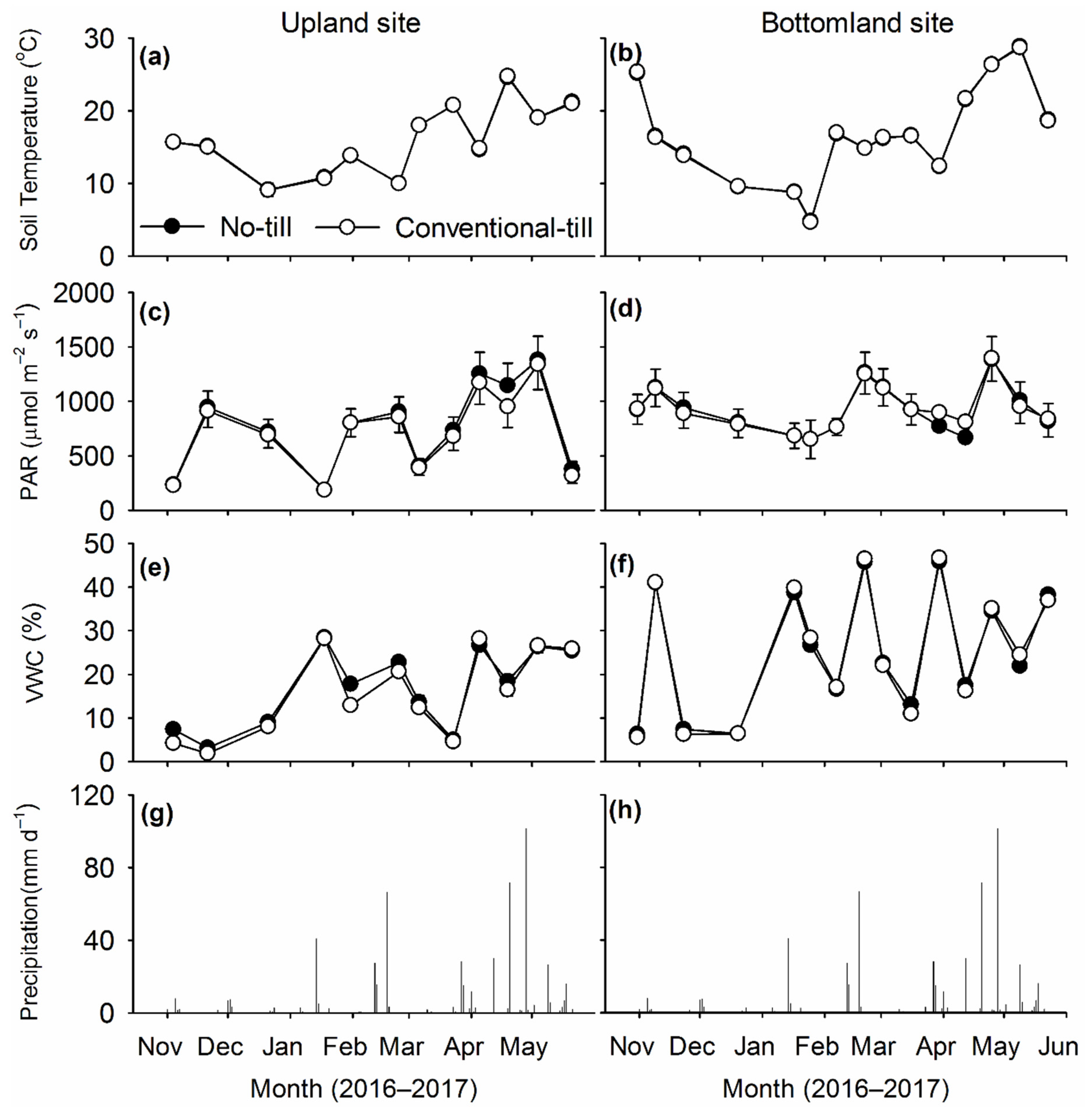
Volumetric water content (VWC) and temperature at the soil surface (0–5 cm) were measured outside each collar at flux measurement using Stevens® Hydra Probe® soil moisture sensors (Stevens Water Monitoring Systems, Inc., Portland, OR, USA). Air temperature during chamber enclosure and precipitation measurements during the study period were obtained from a weather station (Oklahoma Mesonet, Oklahoma Climatological Survey) roughly 1 km from the study site.
2.7. Measurements of Soil Mineral N Concentration
Soil samples (0–0.15 m) were taken from all plots on 2 dates (1 October 2016 and 1 May 2017) to determine concentrations of mineral N [nitrate (NO3−) and ammonium (NH4+)] during early and late growth of winter wheat. Two soil cores (diameter, 0.02 m) were taken from each plot at both soil sampling dates and subsequently pooled to form a composite sample. A subsample (10 g) was extracted in 1.0 M KCl and concentrations of NO3– and NH4+ were analyzed by flow injection method (Timberline Instruments, Boulder, CO, USA).
2.8. Statistical Analysis
Measurements from the subplots receiving combinations of tillage system and N treatments in each site are presented as average and standard errors unless otherwise stated. The effects of tillage system and N treatment were analysed by analysis of variance (ANOVA) using a PROC GLIMMIX in SAS software (SAS Inc., Cary, NC, USA). Since the fluxes were not measured on the same days on the two sites, statistical analyses were individually applied to data from each site. For pairwise comparisons, Fisher’s LSD method was used at 5% level.
4. Discussion
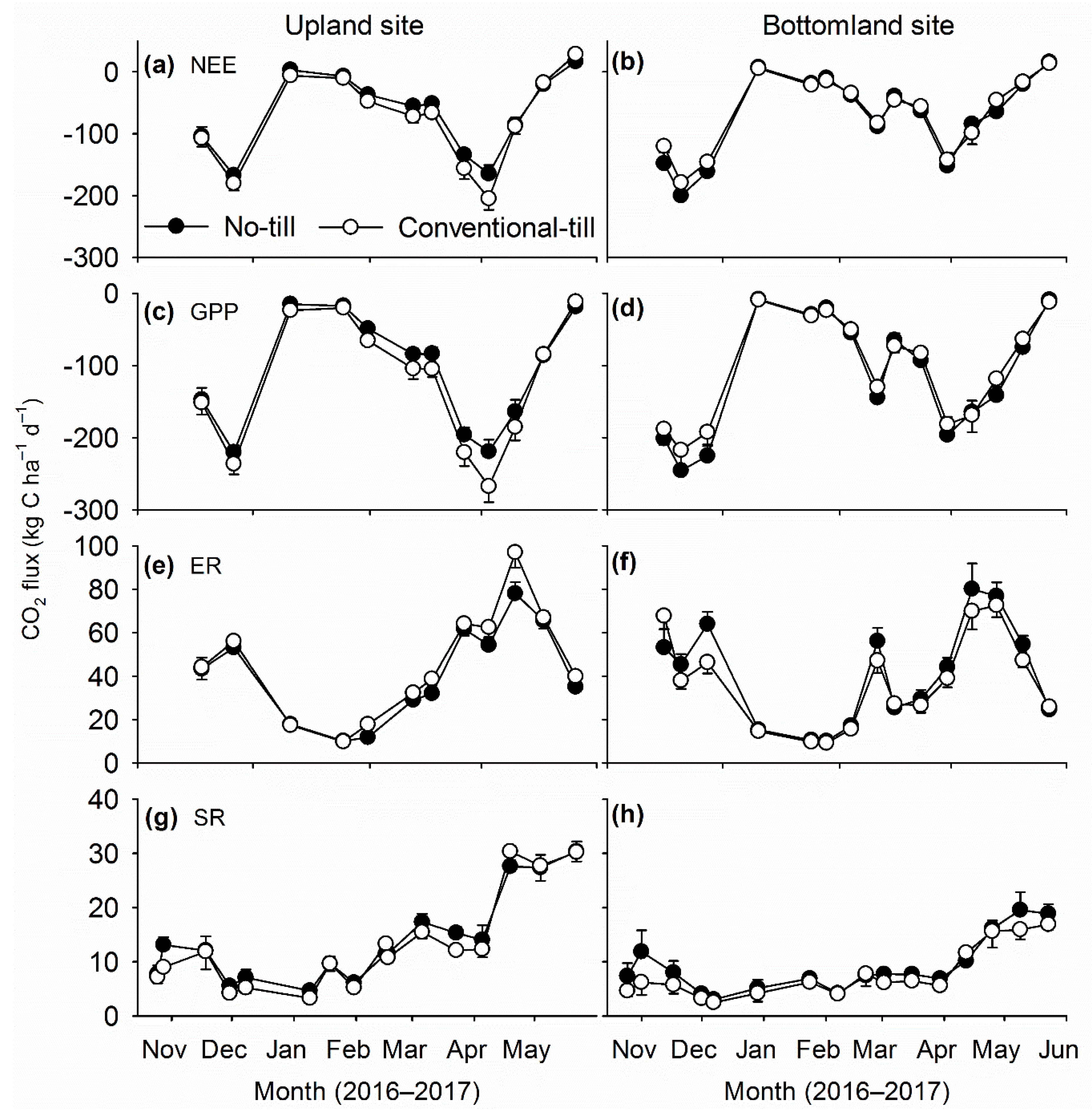
We observed minimal and inconsistent influences of tillage system on canopy fluxes of CO
2 from winter wheat cultivated in central Oklahoma during this study. Albeit influence of tillage system on GPP was statistically significant, the difference was only ~10%, and opposite responses were recorded on the two sites. As with GPP, ER differed by ~10% among tillage systems, and opposite responses were noted on the two sites. Consistent differences in GPP and ER rates on the two sites indicated the difference in ER rates between tillage systems was mostly due to differences in plant growth. A possible reason for similar canopy CO
2 fluxes from the contrasting tillage systems might be related to the lower intensity tillage applied under CT, which was restricted to the upper 20 cm of soil, and residues were left on the soil surface. Previous studies in the SGP have documented similar yield responses of winter wheat to NT and CT systems [
3,
22,
23], and were corroborated by similar levels of GPP for tillage systems in this study. The short distance between sites, and difference in soil properties and fertility, allowed a test of effects of nitrogen and tillage treatments across a broad set of landscapes. Generally, soil in the bottomland areas are considered highly fertile in the region, and hence highly productive. In the upland site, which is less fertile and has a shallower profile than the bottomland site, a CT system would have contributed for better cycling of nutrients through decomposition of biomass residues. This might have contributed for better growth of winter wheat under CT system which contributed for greater canopy CO
2 fluxes.
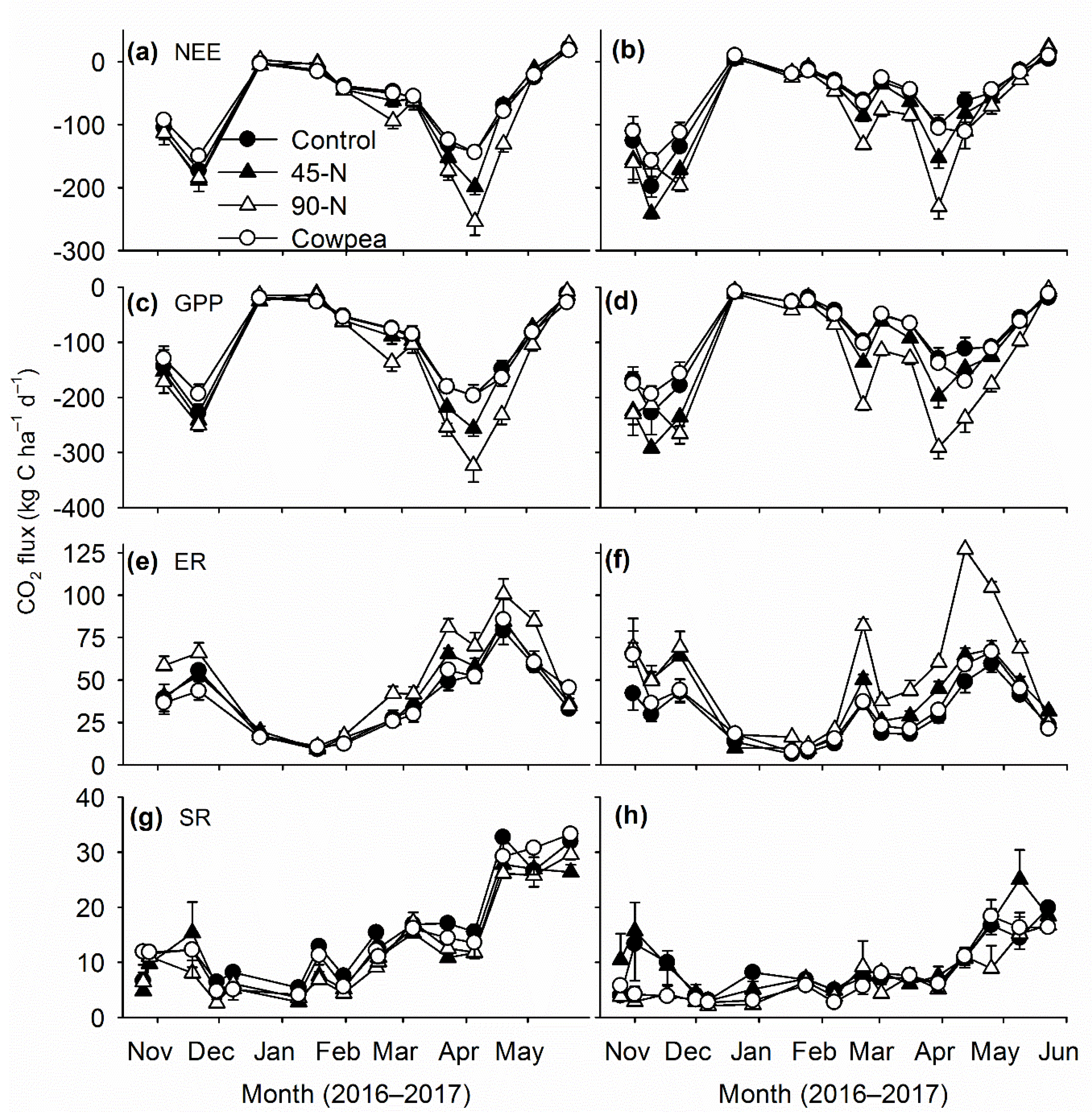
Contrary to system of tillage, large and consistent responses of canopy CO
2 fluxes to N rates were seen on both sites. Canopy fluxes of CO
2 increased with increasing rates of N fertilizer, which indicated strong growth responses of winter wheat to amounts of applied N fertilizer. Increased GPP fluxes with increasing level of N fertilizer corroborate results from previous studies in the SGP, where most studies reported strong positive responses of grain or forage yield of winter wheat to amount of applied N fertilizer [
4,
24,
25]. The results of canopy CO
2 fluxes also corroborate results on crop growth and yield of winter wheat, which was reported in a parallel study [
26]. Higher rates of ER fluxes in fertilized treatments was mostly contributed by increased plant respiration due to better crop growth, as influence of rates of N fertilizer on soil respiration was not significant.
The similar magnitudes and dynamics of canopy CO
2 fluxes from cowpea and 0-N treatments indicated that cowpea was not an effective source of green N for winter wheat. Additionally, the flux rates from cowpea treatments were significantly lower than from the 45-N and 90-N treatments. These results corroborate the previous findings in the region, as poor growth and yield responses of winter wheat to legume-based green N are reported frequently [
3,
4,
25,
27,
28]. Lack of interaction effects between the tillage systems and N treatments indicated that cowpea-based green manure could not replace inorganic N fertilizers in either tillage system. In 2016, cowpea produced 1.1 (±0.2) and 1.9 (±0.5) Mg ha
−1 aboveground dry biomass (mean ± standard deviation) on CT and NT plots at bottomland site, 1.7 (±0.7) and 2.0 (±0.6) Mg ha
−1 aboveground dry biomass on CT and NT plots at upland site. The biomass contained about 32–80 kg N ha
−1 at termination. The proportion of biologically fixed N by cowpea was not determined in this study but a previous study indicated that cowpea terminated 50 days after planting derived 55%–60% of total N in biomass from the atmosphere [
29]. Thus, although cowpea might have increased the pool of N in soils through biological fixation, the biomass N was not transferred effectively to the following crop of winter wheat. This resulted to poor growth of winter wheat, and canopy CO
2 fluxes remained similar to the unfertilized treatment. Future research should focus on improving biomass and N productivity of summer legumes and management approaches, such as timing of termination to synchronize N mineralization from decomposing green manures and N demand of winter wheat.
Increased soil respiration is expected after inclusion of cover crops in a monoculture due to increased inputs of C [
30,
31]. However, this was not observed during the current study, as the cowpea treatment did not have higher soil respiration rates than treatments without cover crops. A possible reason for the similar level of soil respiration from cowpea and fertilized treated plots might be due to similar, or lower, annual yields of biomass produced by the cowpea treatment. Winter wheat yields were lower from the cowpea treatment than the 45-N and 90-N treatments in the previous years. Although grain of winter wheat was harvested, residue was retained in the field, and thus, the fertilized treatments (45-N and 90-N) provided greater amounts of winter wheat residues than cowpea treatments. Another possible reason for similar SR fluxes was decomposition of a large proportion of cowpea biomass during the period between termination of cowpea and start of the flux measurements. Cowpea was terminated on the 17th of August in 2016, to conserve moisture to support germination and early growth of winter wheat, but SR fluxes were measured from the 25
th of October. A total of 140 mm rainfall was received during this period which might have created soil conditions conducive to decomposition of cowpea biomass since soil moisture is one of the key environmental factor to control decomposition of cover crop residues in these study sites [
16,
21]. A recent study documented that legume green manures with low C/N ratio (<15) may lose a large proportion of biomass C within a month after termination, when soil moisture and temperature are favorable for biomass decomposition [
16]. The cowpea biomass in this study had average C/N ratio of 15 at termination. Thus, although cowpea could have increased SR fluxes immediately after termination in August, long-term increases in soil respiration, an indicator of improved soil health, was not observed.
In this study, we did not observe strong responses of N
2O emissions in response to N fertilizers and cowpea green manures, since emissions from all treatments remained low. The first three months during flux measurements were dry, with only 53 mm precipitation received during October through December 2016. Therefore, soil conditions were not conducive for denitrification, a major microbial pathway for N
2O production in the soil [
32]. Although frequent large precipitation events occurred in February and April, emissions did not increase during that time period. The low emissions during these periods was possibly due to efficient uptake of available inorganic N in soil by winter wheat, which was growing rapidly as evidenced by high rates of GPP flux. Legume green manures generally increase emissions of N
2O after termination [
15,
33], but no significant N
2O emissions were detected from the cowpea treatment in this study. Since there was a gap between termination of cowpea and inception of measurement, and significant amounts of rainfall were received during the gap, rainfall induced pulses of N
2O emissions from the cowpea treatment were likely missed. Nevertheless, our results indicated that cowpea neither transferred biomass N effectively to a following crop of winter wheat, nor the N in biomass contributed to increased N
2O emissions during the active growing period of winter wheat. Further studies are required to understand the environmental fate of biologically fixed N of summer legumes in the SGP, and to develop systems of crop management that allow effective transfer of N to crops of winter wheat.











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}