Genetic Diversity of Local Peach (Prunus persica) Accessions from La Palma Island (Canary Islands, Spain)

 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. DNA Extraction and PCR Amplification
2.3. Fingerprinting and Genetic Diversity Analyses
3. Results
3.1. SSR Polymorphism and Molecular Characterization of La Palma Local Peach Accessions
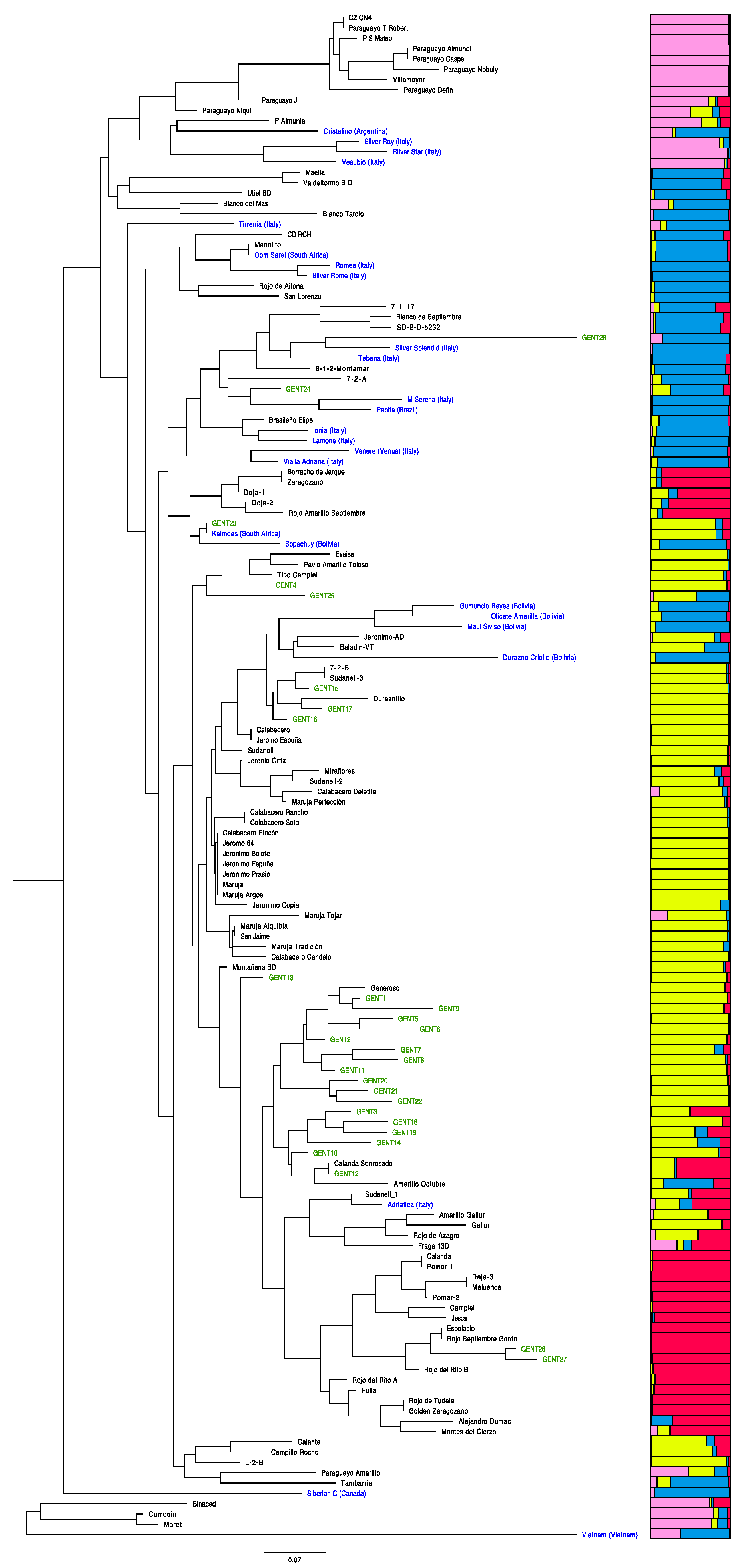
3.2. Comparison of the SSR Polymorphism of La Palma Local Peach Varieties with Other Accessions from Spain and Additional Countries
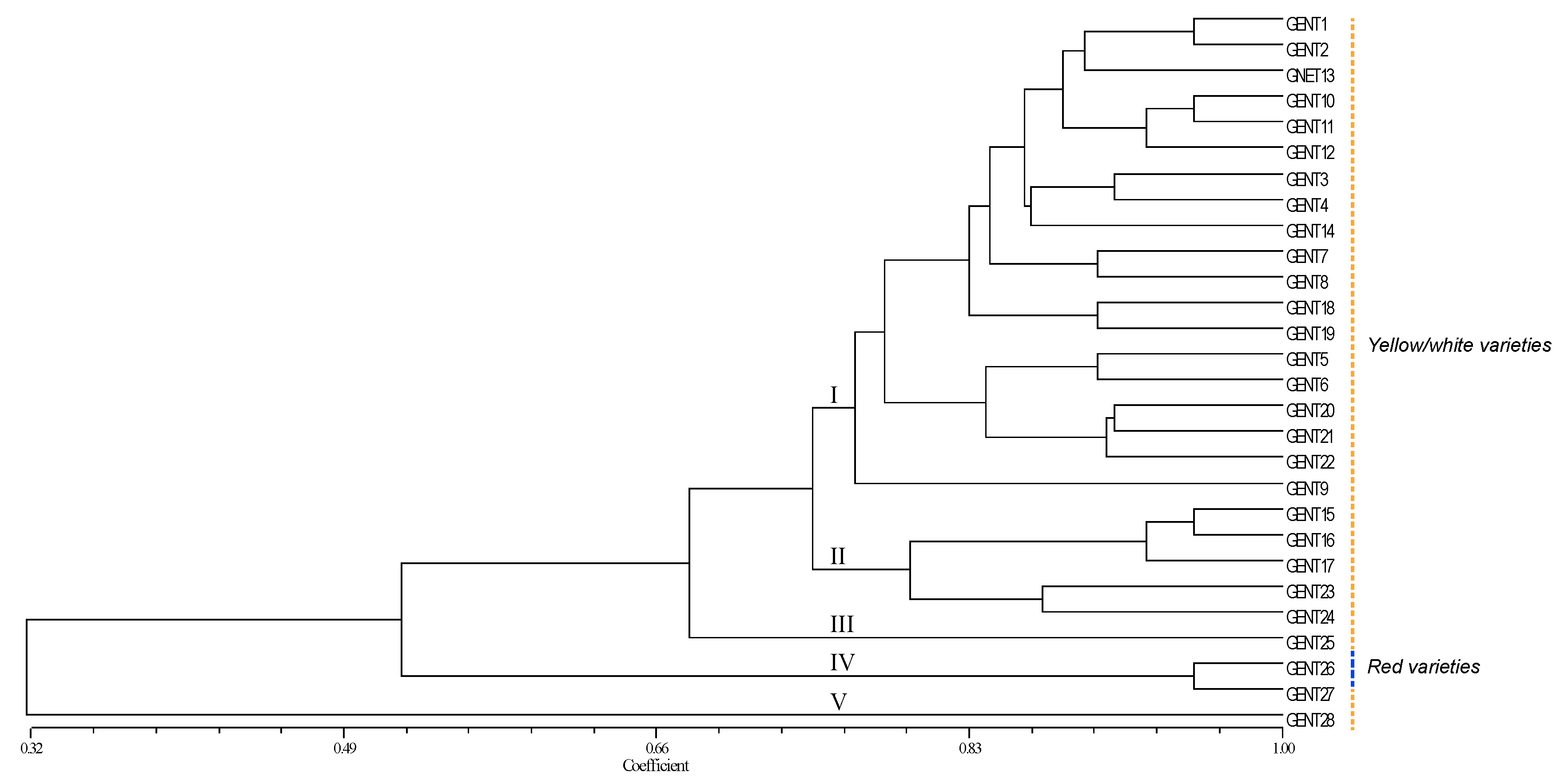
3.3. Genetic Relationships
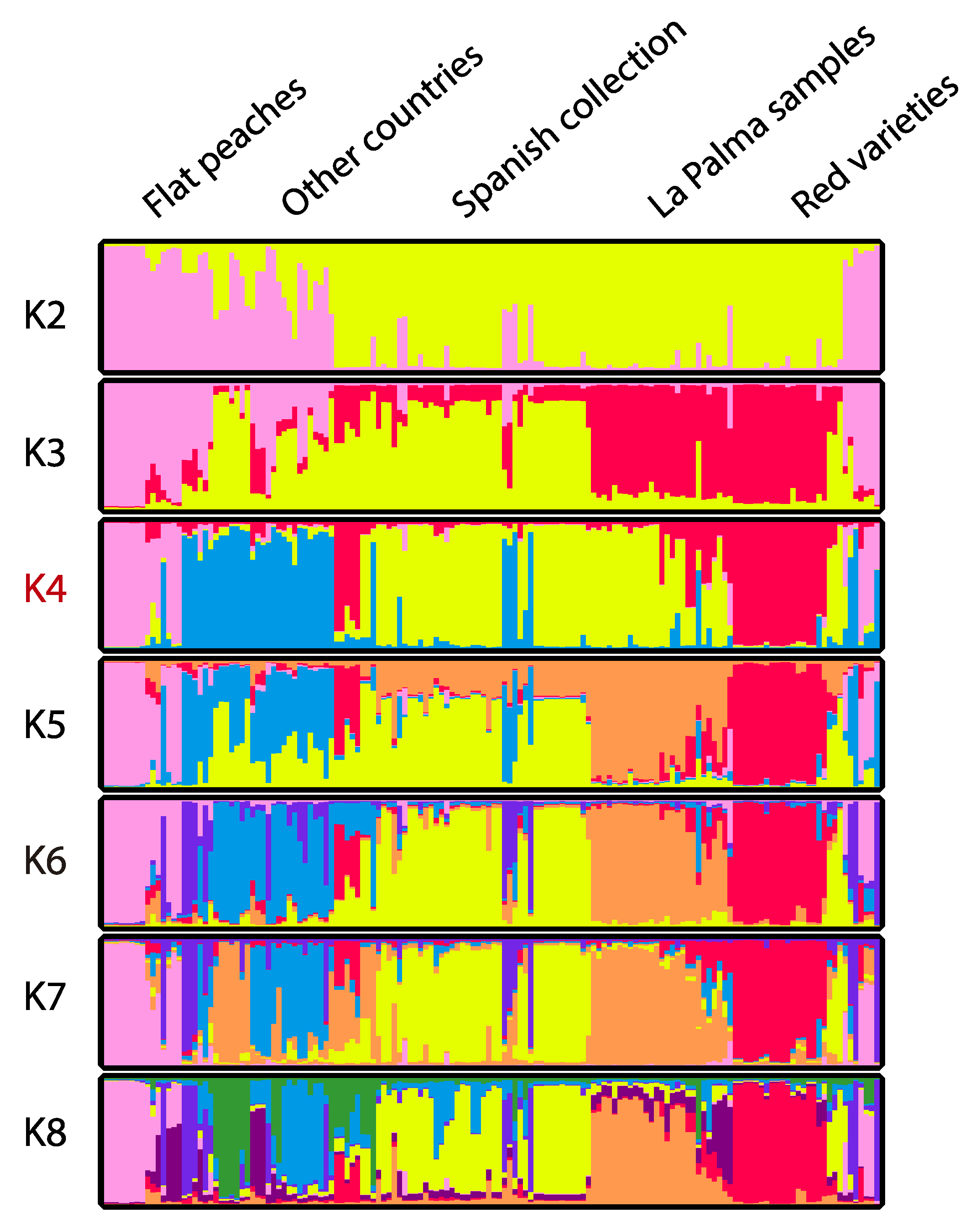
3.4. Genetic Structure
4. Discussion
4.1. La Palma Peach Diversity
4.2. Comparison of the Diversity of La Palma Peach Accessions with Those from Other Origins
4.3. Genetic Structure
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Faust, M.; Timon, B. Origin and dissemination of peach. Hortic. Rev. 1995, 17, 331–379. [Google Scholar]
- Huang, H.W.; Cheng, Z.P.; Zhang, Z.H.; Wang, Y. History of cultivation and trends in China. In The Peach: Botany, Production and Uses; Layne, D.R., Ed.; CABI: Wallingford, UK, 2008; pp. 37–60. [Google Scholar]
- Zheng, Y.; Crawford, G.; Chen, X. Archaeological evidence for peach (Prunus persica) cultivation and domestication in China. PLoS ONE 2014, 9, e106595. [Google Scholar] [CrossRef] [Green Version]
- Yu, Y.; Fu, J.; Xu, Y.; Zhang, J.; Ren, F.; Zhao, H.; Tian, S.; Guo, W.; Tu, X.; Zhao, J.; et al. Genome re-sequencing reveals the evolutionary history of peach fruit edibility. Nature communications 2018, 9, 5404. [Google Scholar] [CrossRef] [Green Version]
- FAOSTAT. Available online: http://www.fao.org/faostat/en/#data (accessed on 13 February 2020).
- Hesse, C.O. Peaches. In Advances in Fruit Breeding; Janick, J., Moore, J.N., Eds.; Purdue University Press: West Lafayette, IN, USA, 1975; pp. 285–335. [Google Scholar]
- Baird, W.V.; Ballard, R.E.; Rajapakse, S.; Abbott, A.G. Progress in Prunus mapping and application of molecular markers to germplasm improvement. HortSci 1996, 31, 1099–1106. [Google Scholar] [CrossRef]
- Li, T.H.; Li, Y.X.; Li, Z.C.; Zhang, H.L.; Qi, Y.W.; Wang, T. Simple sequence repeat analysis of genetic diversity in primary core collection of peach (Prunus persica). J. Integr. Plant Biol. 2008, 50, 102–110. [Google Scholar] [CrossRef] [PubMed]
- Xie, R.; Li, X.; Chai, M.; Song, L.; Jia, H.; Wu, D.; Chen, M.; Chen, K.; Aranzana, M.J.; Gao, Z. Evaluation of the genetic diversity of Asian peach accessions using a selected set of SSR markers. Sci. Hortic 2010, 125, 622–629. [Google Scholar] [CrossRef]
- Li, X.; Meng, X.; Jia, H.; Yu, M.; Ma, R.; Wang, L.; Cao, K.; Shen, Z.; Niu, L.; Tian, J.; et al. Peach genetic resources: Diversity, population structure and linkage disequilibrium. BMC Genet. 2013, 14, 84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aranzana, J.M.; Carbó, J.; Arús, P. Microsatellite variability in peach [Prunus persica (L.) Batsch]: Cultivar identification, marker mutation, pedigree inferences and population structure. Theor. Appl. Genet 2003, 106, 1341–1352. [Google Scholar] [CrossRef] [PubMed]
- The International Peach Genome Initiative. The high-quality draft genome of peach (Prunus persica) identifies unique patterns of genetic diversity, domestication and genome evolution. Nat. Genet. 2013, 45, 487–494. [Google Scholar] [CrossRef] [Green Version]
- Martín, C.; Herrero, M.; Hormaza, J.I. Molecular characterization of apricot germplasm from an old stone collection. PLoS ONE 2011, 6, e23979. [Google Scholar] [CrossRef] [Green Version]
- Wünsch, A.; Carrera, M.; Hormaza, J.I. Molecular characterization of local Spanish peach [Prunus persica (L.) Batsch] germplasm. Genet. Resour. Crop Evol. 2006, 53, 925–932. [Google Scholar] [CrossRef]
- Bouhadida, M.; Moreno, M.A.; Gonzalo, M.J.; Alonso, J.M.; Gogorcena, Y. Genetic variability of introduced and local Spanish peach cultivars determined by SSR markers. Tree Genet. Genomes 2011, 7, 257–270. [Google Scholar] [CrossRef] [Green Version]
- Larranaga, N.; Albertazzi, F.J.; Fontecha, G.; Palmieri, M.; Rainer, H.; van Zonneveld, M.; Hormaza, J.I. A Mesoamerican origin of cherimoya (Annona cherimola Mill.). Implications for the conservation of plant genetic resources. Mol. Ecol. 2017, 26, 4116–4130. [Google Scholar] [CrossRef] [PubMed]
- Hammer, K.; Arrowsmith, N.; Gladis, T. Agrobiodiversity with emphasis on plant genetic resources. Naturwissenschaften 2003, 90, 241–250. [Google Scholar] [CrossRef]
- Santana-Pérez, G. Comercio palmero en el tránsito del siglo XVI al XVII: Tras el signo del ataque de Drake. Tebeto: Anuario del Archivo Histórico Insular de Fuerteventura 2014, 7, 73–89. (In Spanish) [Google Scholar]
- Mendizabal, I.; Sandoval, K.; Berniell-Lee, G.; Calafell, F.; Salas, A.; Martínez-Fuentes, A.; Comas, D. Genetic origin, admixture, and asymmetry in maternal and paternal human lineages in Cuba. BMC Evol. Biol. 2008, 8, 213. [Google Scholar] [CrossRef] [Green Version]
- Gómez-Carballa, A.; Ignacio-Veiga, A.; Álvarez-Iglesias, V.; Pastoriza-Mourelle, A.; Ruíz, Y.; Pineda, L.; Carracedo, A.; Salas, A. A melting pot of multicontinental mtDNA lineages in admixed Venesuelans. Am. J. Phys. Anthropol. 2012, 147, 78–87. [Google Scholar] [CrossRef]
- Suárez-Bosa, M.; Suárez-Viera, F. Emigration and Canary Islands entrepreneurship in Cuba, 1850–1950. Secuencia 2013, 87, 97–126. (In Spanish) [Google Scholar]
- Brito, W.R. La Agricultura en la isla de La Palma; Instituto de Estudios Canarios: La Laguna: Tenerife, Spain, 1982. (In Spanish) [Google Scholar]
- Viera y Clavijo, J.D. Diccionario de Historia Natural de las Islas Canarias; Real Sociedad de Amigos del País de Las Palmas de Gran Canaria: Gran Canaria, Spain, 1866. (In Spanish) [Google Scholar]
- Herrero, J. Cartografía de Frutales de Hueso y Pepita; Estación Experimental de Aula Dei (EEAD-CSIC): Zaragoza, Spain, 1964. (In Spanish) [Google Scholar]
- ISTAC. Available online: http://www.gobiernodecanarias.org/istac/jaxi-istac/tabla.do (accessed on 9 December 2019).
- Varshney, R.K.; Graner, A.; Sorrells, M.E. Genomics-assisted breeding for crop improvement. Trends Plant Sci. 2005, 10, 621–630. [Google Scholar] [CrossRef] [Green Version]
- Cheng, Z.; Huang, H. SSR fingerprinting Chinese peach cultivars and landraces (Prunus persica) and analysis of their genetic relationships. Sci. Hortic. 2009, 120, 188–193. [Google Scholar] [CrossRef]
- Bouhadida, M.; Casas, A.M.; Moreno, M.A.; Gogorcena, Y. Molecular characterization of Miraflores peach variety and relatives using SSRs. Sci. Hortic. 2007, 111, 140–145. [Google Scholar] [CrossRef] [Green Version]
- Testolin, R.; Marrazzo, T.; Cipriani, G.; Quarta, R.; Verde, I.; Dettori, M.T.; Pancaldi, M.; Sansavini, S. Microsatellite DNA in peach (Prunus persica L. Batsch) and its use in fingerprinting and testing the genetic origin of cultivars. Genome 2000, 43, 512–520. [Google Scholar] [CrossRef] [PubMed]
- Aranzana, M.J.; Abbassi, E.K.; Howad, W.; Arús, P. Genetic variation, population structure and linkage disequilibrium in peach commercial varieties. BMC Genet. 2010, 11, 69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoon, J.; Liu, D.; Song, W.; Liu, W.; Zhang, A.; Li, S. Genetic diversity and ecogeographical phylogenetic relationships among peach and nectarine cultivars based on simple sequence repeat (SSR) markers. J. Am. Soc. Hortic. Sci. 2006, 131, 513–521. [Google Scholar] [CrossRef] [Green Version]
- Hormaza, J.I. Molecular characterization and similarity relationships among apricot (Prunus armeniaca L.) genotypes using simple sequence repeats. Theor. Appl. Genet. 2002, 104, 321–328. [Google Scholar] [CrossRef]
- QGIS Development Team. Version 2.8.1-Wien. QGIS Geographic Information System. Open Source Geospatial Foundation Project. Available online: http://www.qgis.org/ (accessed on 23 February 2020).
- Rohlf, F.J. NTSYS-PC: Numerical Taxonomy and Multivariable Analysis System—Version 2.0; Exeter Software: New York, NY, USA, 1993. [Google Scholar]
- Nei, M.; Li, W.H. Mathematical model for studying genetic variation in terms of restriction endonucleases. Proc. Natl. Acad. Sci. USA 1979, 76, 5269–5273. [Google Scholar] [CrossRef] [Green Version]
- Wright, S. The genetical structure of populations. Ann. Eugen. 1949, 15, 323–354. [Google Scholar] [CrossRef]
- Jombart, T. adegenet: A R package for the multivariate analysis of genetic markers. Bioinformatics 2008, 24, 1403–1405. [Google Scholar] [CrossRef] [Green Version]
- Jombart, T.; Ahmed, I. adegenet 1.3-1: New tools for the analysis of genome-wide SNP data. Bioinformatics 2011, 27, 3070–3071. [Google Scholar] [CrossRef] [Green Version]
- Adamack, A.T.; Gruber, B. PopGenReport: Simplifying basic population genetic analyses in R. Methods Ecol. Evol. 2014, 5, 384–387. [Google Scholar] [CrossRef]
- Gruber, B.; Adamack, A.T. Landgenreport: A new R function to simplify landscape genetic analysis using resistance surface layers. Mol. Ecol. Resour. 2015, 15, 1172–1178. [Google Scholar] [CrossRef] [PubMed]
- Fig Tree. Available online: http://tree.bio.ed.ac.uk/software/figtree/ (accessed on 23 March 2020).
- Hubisz, M.J.; Falush, D.; Stephens, M.; Pritchard, J.K. Inferring weak population structure with the assistance of sample group information. Mol. Ecol. Resour. 2009, 9, 1322–1332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the number of clusters of individuals using the software STRUCTURE: A simulation study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Earl, D.A.; von Holdt, B.M. Structure Harvester: A website and program for visualizing STRUCTURE output and implementing the Evanno method. Conserv. Genet. Resour. 2012, 4, 359–361. [Google Scholar] [CrossRef]
- Jakobsson, M.; Rosenberg, N.A. CLUMPP: A cluster matching and permutation program for dealing with label switching and multimodality in analysis of population structure. Bioinformatics 2007, 23, 1801–1806. [Google Scholar] [CrossRef] [Green Version]
- Rosenberg, N.A. DISTRUCT: A program for the graphical display of population structure. Mol. Ecol. Notes 2004, 4, 137–138. [Google Scholar] [CrossRef]
- Sosinski, B.; Gannavarapu, M.; Hager, L.D.; Beck, L.E.; King, G.J.; Ryder, C.D.; Rajapakse, S.; Baird, W.V.; Ballard, R.E.; Abbott, A.G. Characterization of microsatellite markers in peach [Prunus persica (L.) Batsch]. Theor. Appl. Genet. 2000, 101, 421–428. [Google Scholar] [CrossRef]
- Cipriani, G.; Lot, G.; Huang, W.G.; Marrazzo, M.T.; Peterlunger, E.; Testolin, R. AC/GT and AG/CT microsatellite repeats in peach [Prunus persica (L) Batsch]: Isolation, characterisation and cross-species amplification in Prunus. Theor. Appl. Genet. 1999, 99, 65–72. [Google Scholar] [CrossRef]
- Aranzana, M.J.; Garcia-Mas, J.; Carbo, J.; Arús, P. Development and variability analysis of microsatellite markers in peach. Plant Breed. 2002, 121, 87–92. [Google Scholar] [CrossRef]
- Padilla, G.; Socias i Company, R.; Ordás, A. Molecular characterization of almond accessions from the island of La Palma (Canary Islands, Spain) using SSR markers. Plant Genet. Resour. 2014, 12, 323–329. [Google Scholar] [CrossRef]
- Pereira-Lorenzo, S.; Ramos-Cabrer, A.M.; Gonzalez-Diaz, A.J.; Diaz-Hernandez, M.B. Genetic assessment of local apple cultivars from La Palma, Spain, using simple sequence repeats (SSRs). Sci. Hortic. 2008, 117, 160–166. [Google Scholar] [CrossRef]
- Pereira-Lorenzo, S.; Costa, R.M.L.; Ramos-Cabrer, A.M.; Ciordia-Ara, M.; Ribeiro, C.A.M.; Borges, O.; Barreneche, T. Chestnut cultivar diversification process in the Iberian Peninsula, Canary Islands, and Azores. Genome 2011, 54, 301–315. [Google Scholar] [CrossRef] [PubMed]
- Pereira-Lorenzo, S.; Ramos-Cabrer, A.M.; Ferreira, V.; Díaz-Hernández, M.B.; Carnide, V.; Pinto-Carnide, O.; Rodrigues, R.; Velázquez-Barrera, M.E.; Rios-Mesa, D.; Ascasíbar-Errasti, J.; et al. Genetic diversity and core collection of Malus × domestica in northwestern Spain, Portugal and the Canary Islands by SSRs. Sci. Hortic. 2018, 240, 49–56. [Google Scholar] [CrossRef]
- Marsal, G.; Méndez, J.J.; Mateo, J.M.; Ferrer, S.; Canals, J.M.; Zamora, F.; Fort, F. Molecular characterization of Vitis vinifera L. local cultivars from volcanic areas (Canary Islands and Madeira) using SSR markers. OENO One 2019, 53. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| SSR Locus | Number of Alleles | Ho | He | Ne | F |
|---|---|---|---|---|---|
| Pchgms-3 a | 9 | 0.372 | 0.589 | 2.437 | 0.370 |
| UDP96-001 b | 8 | 0.150 | 0.627 | 2.683 | 0.761 |
| UDP96-003 b | 6 | 0.149 | 0.241 | 1.318 | 0.383 |
| UDP96-008 b | 4 | 0.149 | 0.189 | 1.234 | 0.216 |
| UDP96-018 b | 4 | 0.345 | 0.508 | 2.034 | 0.322 |
| UDP98-406 b | 6 | 0.108 | 0.160 | 1.190 | 0.323 |
| UDP96-015 b | 8 | 0.338 | 0.649 | 2.848 | 0.479 |
| UDP98-407 b | 5 | 0.128 | 0.400 | 1.666 | 0.679 |
| UDP98-416 c | 5 | 0.101 | 0.348 | 1.533 | 0.709 |
| CPPCT6 d | 4 | 0.142 | 0.389 | 1.637 | 0.635 |
| Samples | Sample Size | Mean n° of Alleles/Locus | N° of Polymorphic Loci | Ho | He | Fis | N° of Private Alleles |
|---|---|---|---|---|---|---|---|
| La Palma | 28 | 3.2 | 9 | 0.075 | 0.280 | 0.732 | 5 |
| Spanish collection | 95 | 4.1 | 10 | 0.232 | 0.398 | 0.417 | 6 |
| Other countries | 26 | 4.7 | 10 | 0.208 | 0.476 | * 0.564 | 11 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pérez, V.; Larrañaga, N.; Abdallah, D.; Wünsch, A.; Hormaza, J.I. Genetic Diversity of Local Peach (Prunus persica) Accessions from La Palma Island (Canary Islands, Spain). Agronomy 2020, 10, 457. https://doi.org/10.3390/agronomy10040457
Pérez V, Larrañaga N, Abdallah D, Wünsch A, Hormaza JI. Genetic Diversity of Local Peach (Prunus persica) Accessions from La Palma Island (Canary Islands, Spain). Agronomy. 2020; 10(4):457. https://doi.org/10.3390/agronomy10040457
Chicago/Turabian StylePérez, Verónica, Nerea Larrañaga, Donia Abdallah, Ana Wünsch, and José I. Hormaza. 2020. "Genetic Diversity of Local Peach (Prunus persica) Accessions from La Palma Island (Canary Islands, Spain)" Agronomy 10, no. 4: 457. https://doi.org/10.3390/agronomy10040457
APA StylePérez, V., Larrañaga, N., Abdallah, D., Wünsch, A., & Hormaza, J. I. (2020). Genetic Diversity of Local Peach (Prunus persica) Accessions from La Palma Island (Canary Islands, Spain). Agronomy, 10(4), 457. https://doi.org/10.3390/agronomy10040457