Management Intensity Controls Nitrogen-Use-Efficiency and Flows in Grasslands—A 15N Tracing Experiment

Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Sites and Experimental Design
2.2. Production and Use of 15N Labeled Cattle Slurry
2.3. Sampling and Sample Preparation
2.4. Calculation of 15N Recovery
2.5. Nitrogen Balance
2.6. Statistical Analysis
3. Results
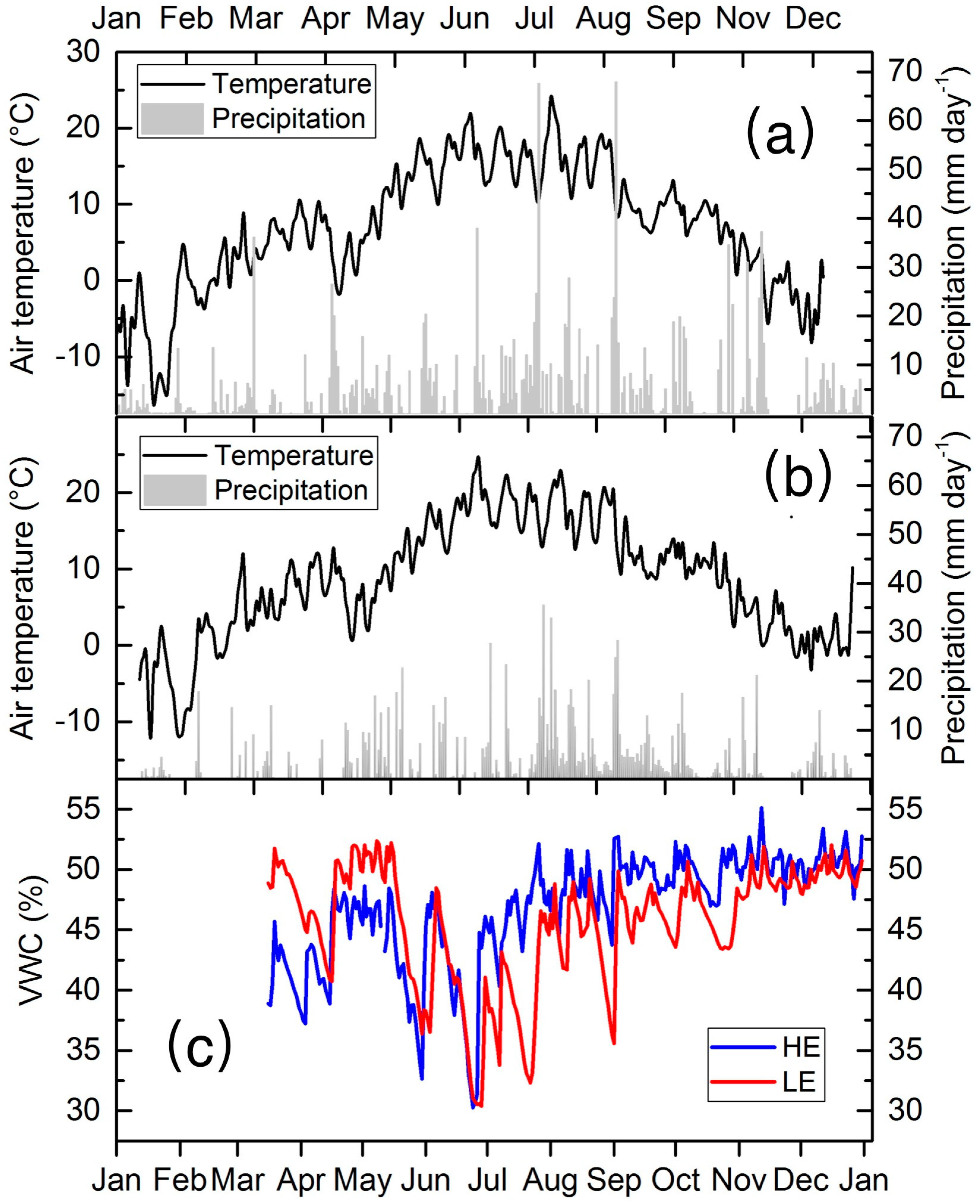
3.1. Meteorological Conditions
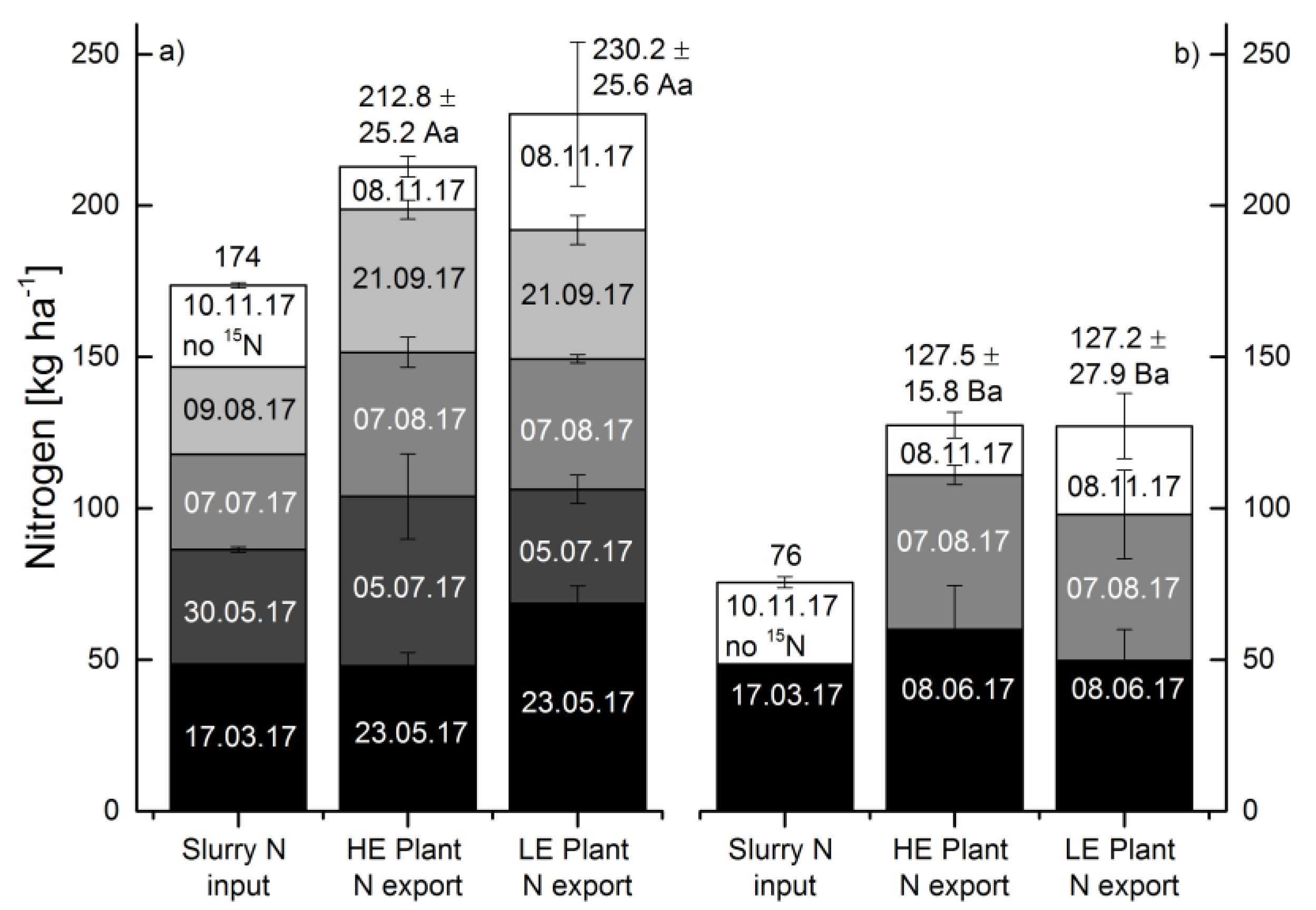
3.2. Plant Biomass Yields, N Content and Associated N Export
3.3. Slurry N Input
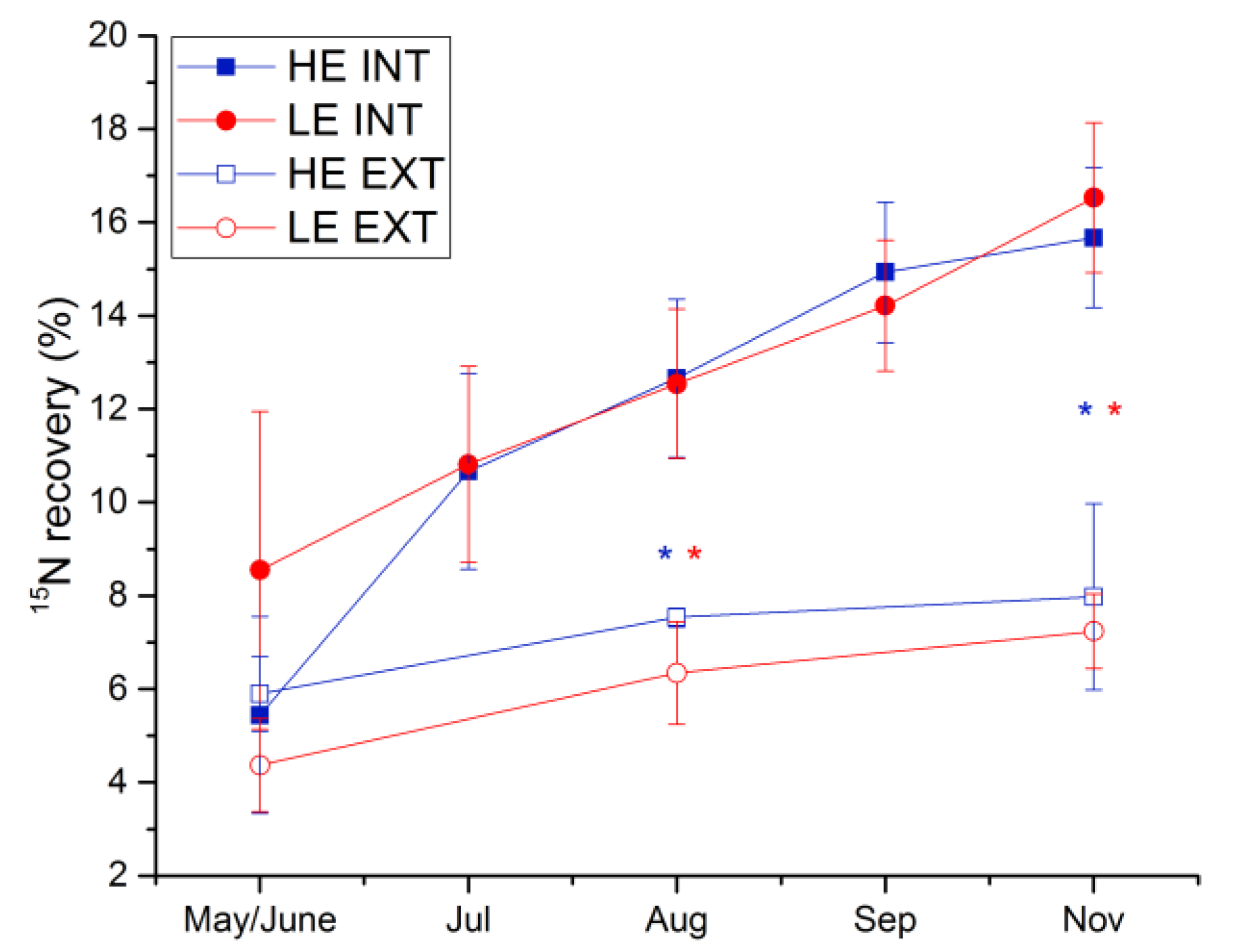
3.4. 15N Recovery from Labeled Fertilizer
3.4.1. Plant 15N Recovery
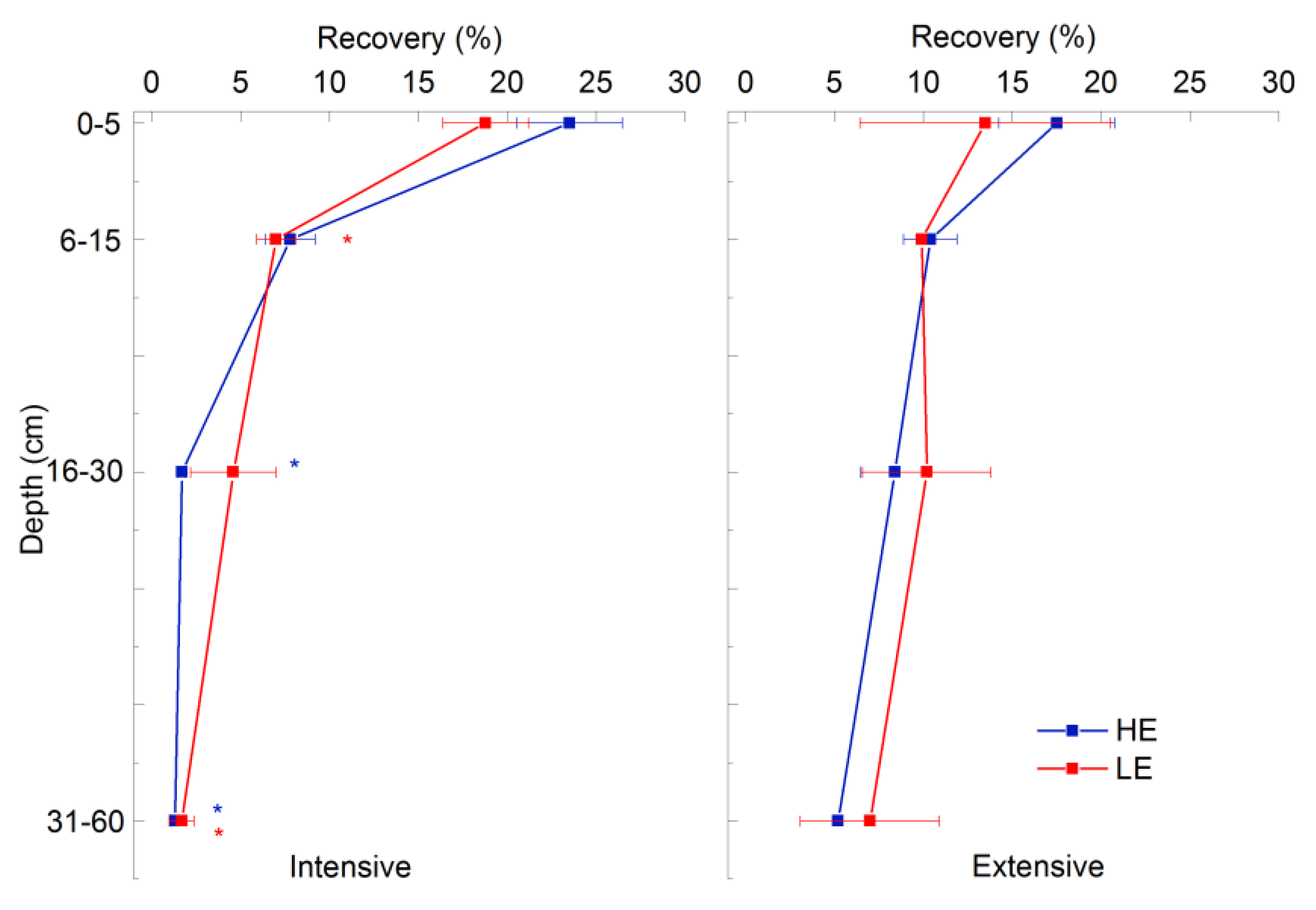
3.4.2. 15N Recovery in Total Soil Nitrogen
3.4.3. Total 15N Recovery
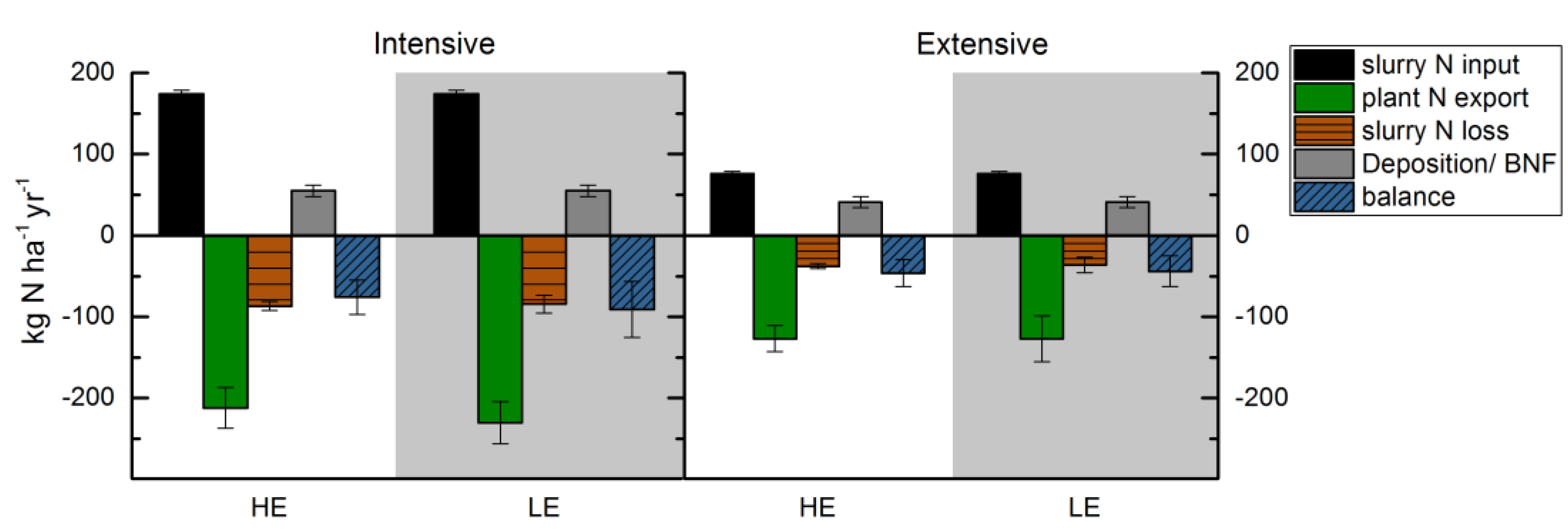
3.5. Nitrogen Balance
4. Discussion
4.1. N Cycling in Montane Grassland Soil
4.2. Effects of Climate Change Simulation
4.3. Effects of Intensification
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- BMELV. Statistisches Jahrbuch über Ernährung, Landwirtschaft und Forsten der Bundesrepublik Deutschland; Landwirtsschaftsverlag GmbH: Münster, Germany, 2012; Available online: https://www.bmel-statistik.de/fileadmin/SITE_MASTER/content/Jahrbuch/Agrarstatistisches-Jahrbuch-2012.pdf (accessed on 14 January 2020).
- Wiesmeier, M.; Hübner, R.; Barthold, F.; Spörlein, P.; Geuß, U.; Hangen, E.; Reischl, A.; Schilling, B.; von Lützow, M.; Kögel-Knabner, I. Amount, distribution and driving factors of soil organic carbon and nitrogen in cropland and grassland soils of southeast Germany (Bavaria). Agric. Ecosyst. Environ. 2013, 176, 39–52. [Google Scholar] [CrossRef]
- Von Glasenapp, M.; Thornton, T.F. Traditional ecological knowledge of Swiss alpine farmers and their resilience to socioecological change. Hum. Ecol. 2011, 39, 769–781. [Google Scholar] [CrossRef]
- Poschlod, P.; Wallis de Vries, M.F. The historical and socioeconomic perspective of calcareous grasslands—Lessons from the distant and recent past. Biol. Conserv. 2002, 104, 361–376. [Google Scholar] [CrossRef]
- Wilson, J.B.; Peet, R.K.; Dengler, J.; Pärtel, M. Plant species richness: The world records. J. Veg. Sci. 2012, 23, 796–802. [Google Scholar] [CrossRef]
- Rabot, E.; Wiesmeier, M.; Schlüter, S.; Vogel, H.-J. Soil structure as an indicator of soil functions: A review. Geoderma 2018, 314, 122–137. [Google Scholar] [CrossRef]
- Blum, W.E.H. Functions of soil for society and the environment. Rev. Environmental Sci. Bio/Technol. 2005, 4, 75–79. [Google Scholar] [CrossRef]
- Patterson, M. OECD Agriculture Statistics; OECD Publishing: Paris, France, 2012; Available online: http://www.fao.org/3/i9166e/i9166e_Chapter7_Dairy.pdf (accessed on 14 January 2020).
- Sebastià, M.-T.; Kirwan, L.; Connolly, J. Strong shifts in plant diversity and vegetation composition in grassland shortly after climatic change. J. Veg. Sci. 2008, 19, 299–306. [Google Scholar] [CrossRef]
- Berauer, B.J.; Wilfahrt, P.A.; Arfin-Khan, M.A.S.; Eibes, P.; von Heßberg, A.; Ingrisch, J.; Schloter, M.; Schuchardt, M.A.; Jentsch, A. Low resistance of montane and alpine grasslands to abrupt changes in temperature and precipitation regimes. Arct. Antarct. Alp. Res. 2019, 51, 215–231. [Google Scholar] [CrossRef]
- Wagner, S.; Berg, P.; Schädler, G.; Kunstmann, H. High resolution regional climate model simulations for Germany: Part II—Projected climate changes. Clim. Dyn. 2013, 40, 415–427. [Google Scholar] [CrossRef]
- Smiatek, G.; Kunstmann, H.; Knoche, R.; Marx, A. Precipitation and temperature statistics in high-resolution regional climate models: Evaluation for the European Alps. J. Geophys. Res. Atmos. 2009, 114. [Google Scholar] [CrossRef] [Green Version]
- Pepin, N.; Bradley, R.S.; Diaz, H.F.; Baraër, M.; Caceres, E.B.; Forsythe, N.; Fowler, H.; Greenwood, G.; Hashmi, M.Z.; Liu, X.D. Elevation-dependent warming in mountain regions of the world. Nat. Clim. Chang. 2015, 5, 424–430. [Google Scholar]
- Gobiet, A.; Kotlarski, S.; Beniston, M.; Heinrich, G.; Rajczak, J.; Stoffel, M. 21st century climate change in the European Alps—A review. Sci. Total Environ. 2014, 493, 1138–1151. [Google Scholar] [CrossRef] [PubMed]
- Warscher, M.; Wagner, S.; Marke, T.; Laux, P.; Smiatek, G.; Strasser, U.; Kunstmann, H. A 5 km resolution regional climate simulation for central europe: Performance in high mountain areas and seasonal, regional and elevation-dependent variations. Atmosphere 2019, 10, 682. [Google Scholar] [CrossRef] [Green Version]
- Rajczak, J.; Pall, P.; Schär, C. Projections of extreme precipitation events in regional climate simulations for Europe and the Alpine Region. J. Geophys. Res. Atmos. 2013, 118, 3610–3626. [Google Scholar] [CrossRef]
- Giorgi, F.; Torma, C.; Coppola, E.; Ban, N.; Schär, C.; Somot, S. Enhanced summer convective rainfall at Alpine high elevations in response to climate warming. Nat. Geosci. 2016, 9, 584–589. [Google Scholar] [CrossRef]
- Galler, P.V.J. Wirtschaftsdünger: Anfall, Lagerung, Verwertung, Umwelt; Landwirtschaftskammer: Salzburg, Austria, 2009; Available online: https://www.infothek-biomasse.ch/images//205_2009_LK_Hofduenger_naehrstoffe_AUT.pdf (accessed on 14 January 2020).
- Wang, C.; Chen, Z.; Unteregelsbacher, S.; Lu, H.; Gschwendtner, S.; Gasche, R.; Kolar, A.; Schloter, M.; Kiese, R.; Butterbach-Bahl, K. Climate change amplifies gross nitrogen turnover in montane grasslands of Central Europe in both summer and winter seasons. Glob. Chang. Biol. 2016, 22, 2963–2978. [Google Scholar] [CrossRef]
- Fu, J.; Gasche, R.; Wang, N.; Lu, H.; Butterbach-Bahl, K.; Kiese, R. Impacts of climate and management on water balance and nitrogen leaching from montane grassland soils of S-Germany. Environ. Pollut. 2017, 229, 119–131. [Google Scholar] [CrossRef]
- Zistl-Schlingmann, M.; Tobler, U.; Berauer, B.J.; Garcia-Franco, N.; Wilfahrt, P.A.; Wiesmeier, M.; Jentsch, A.; Wolf, B.; Kiese, R.; Dannenmann, M. Intensive slurry management and climate change promote nitrogen mining from organic matter-rich montane grassland soils. submitted.
- Walter, J.; Grant, K.; Beierkuhnlein, C.; Kreyling, J.; Weber, M.; Jentsch, A. Increased rainfall variability reduces biomass and forage quality of temperate grassland largely independent of mowing frequency. Agric. Ecosyst. Environ. 2012, 148, 1–10. [Google Scholar] [CrossRef]
- Cantarel, A.A.M.; Bloor, J.M.G.; Soussana, J.-F. Four years of simulated climate change reduces above-ground productivity and alters functional diversity in a grassland ecosystem. J. Veg. Sci. 2013, 24, 113–126. [Google Scholar] [CrossRef]
- Vries, F.T.; Bloem, J.; Quirk, H.; Stevens, C.J.; Bol, R.; Bardgett, R.D. Extensive management promotes plant and microbial nitrogen retention in temperate grassland. PLoS ONE 2012, 7, e51201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cardenas, L.; Bhogal, A.; Chadwick, D.; McGeough, K.; Misselbrook, T.; Rees, R.M.; Thorman, R.; Watson, C.; Williams, J.; Smith, K.; et al. Nitrogen use efficiency and nitrous oxide emissions from five UK fertilised grasslands. Sci. Total Environ. 2019, 661, 696–710. [Google Scholar] [CrossRef]
- Cantarel, A.A.M.; Bloor, J.M.G.; Deltroy, N.; Soussana, J.-F. Effects of climate change drivers on nitrous oxide fluxes in an upland temperate grassland. Ecosystems 2011, 14, 223–233. [Google Scholar] [CrossRef]
- Butterbach-Bahl, K.; Dannenmann, M. Soil carbon and nitrogen interactions and biosphere-atmosphere exchange of nitrous oxide and methane. In Recarbonization of the Biosphere; Springer: Berlin/Heidelberg, Germany, 2012; pp. 429–443. [Google Scholar]
- Wolf, B.; Zheng, X.; Brüggemann, N.; Chen, W.; Dannenmann, M.; Han, X.; Sutton, M.A.; Wu, H.; Yao, Z.; Butterbach-Bahl, K. Grazing-induced reduction of natural nitrous oxide release from continental steppe. Nature 2010, 464, 881–884. [Google Scholar] [CrossRef] [PubMed]
- Jensen, L.S.; Pedersen, I.S.; Hansen, T.B.; Nielsen, N.E. Turnover and fate of 15N-labelled cattle slurry ammonium-N applied in the autumn to winter wheat. Eur. J. Agron. 2000, 12, 23–35. [Google Scholar] [CrossRef]
- Paul, J.W.; Beauchamp, E.G. Availability of manure slurry ammonium for corn using 15N-labelled (NH4) 2SO4. Can. J. Soil Sci. 1995, 75, 35–42. [Google Scholar] [CrossRef] [Green Version]
- Rocha, K.F.; Mariano, E.; Grassmann, C.S.; Trivelin, P.C.O.; Rosolem, C.A. Fate of 15N fertilizer applied to maize in rotation with tropical forage grasses. Field Crop. Res. 2019, 238, 35–44. [Google Scholar] [CrossRef]
- Christian, D.G.; Poulton, P.R.; Riche, A.B.; Yates, N.E. The recovery of 15N-labelled fertilizer applied to Miscanthus× giganteus. Biomass Bioenergy 1997, 12, 21–24. [Google Scholar] [CrossRef]
- Scheiner, J.D.; Gutiérrez-Boem, F.H.; Lavado, R.S. Sunflower nitrogen requirement and 15N fertilizer recovery in Western Pampas, Argentina. Eur. J. Agron. 2002, 17, 73–79. [Google Scholar] [CrossRef]
- Rowlings, D.W.; Scheer, C.; Liu, S.; Grace, P.R. Annual nitrogen dynamics and urea fertilizer recoveries from a dairy pasture using 15N; effect of nitrification inhibitor DMPP and reduced application rates. Agric. Ecosyst. Environ. 2016, 216, 216–225. [Google Scholar] [CrossRef]
- Zhou, M.; Zhu, B.; Brüggemann, N.; Dannenmann, M.; Wang, Y.; Butterbach-Bahl, K. Sustaining crop productivity while reducing environmental nitrogen losses in the subtropical wheat-maize cropping systems: A comprehensive case study of nitrogen cycling and balance. Agric. Ecosyst. Environ. 2016, 231, 1–14. [Google Scholar] [CrossRef]
- Kiese, R.; Fersch, B.; Baessler, C.; Brosy, C.; Butterbach-Bahl, K.; Chwala, C.; Dannenmann, M.; Fu, J.; Gasche, R.; Grote, R. The TERENO Pre-Alpine Observatory: Integrating meteorological, hydrological, and biogeochemical measurements and modeling. Vadose Zone J. 2018, 17. [Google Scholar] [CrossRef]
- IUSS Working Group WRB. World Soil Resources Reports No. 106. In World Reference Base for Soil Resources 2014; update 2015; FAO: Rome, Italy, 2015. [Google Scholar]
- Pütz, T.; Kiese, R.; Wollschläger, U.; Groh, J.; Rupp, H.; Zacharias, S.; Priesack, E.; Gerke, H.H.; Gasche, R.; Bens, O. TERENO-SOILCan: A lysimeter-network in Germany observing soil processes and plant diversity influenced by climate change. Environ. Earth Sci. 2016, 75, 1242. [Google Scholar] [CrossRef]
- Guo, C.; Simon, J.; Gasche, R.; Naumann, P.S.; Bimüller, C.; Pena, R.; Polle, A.; Kögel-Knabner, I.; Zeller, B.; Rennenberg, H. Minor contribution of leaf litter to N nutrition of beech (Fagus sylvatica) seedlings in a mountainous beech forest of Southern Germany. Plant Soil 2013, 369, 657–668. [Google Scholar] [CrossRef]
- LfL. Wirtschaftsdünger und Gewässerschutz Lagerung und Ausbringung von Wirtschaftsdüngern in der Landwirtschaft. Available online: https://www.lfl.bayern.de/mam/cms07/publikationen/daten/informationen/p_34348.pdf (accessed on 14 January 2020).
- Jacot, K.A.; Lüscher, A.; Nösberger, J.; Hartwig, U.A. The relative contribution of symbiotic N2 fixation and other nitrogen sources to grassland ecosystems along an altitudinal gradient in the Alps. Plant Soil. 2000, 225, 201–211. [Google Scholar] [CrossRef]
- Ammann, C.; Flechard, C.R.; Leifeld, J.; Neftel, A.; Fuhrer, J. The carbon budget of newly established temperate grassland depends on management intensity. Agric. Ecosyst. Environ. 2007, 121, 5–20. [Google Scholar] [CrossRef]
- Hoekstra, N.J.; Lalor, S.T.J.; Richards, K.G.; O’Hea, N.; Lanigan, G.J.; Dyckmans, J.; Schulte, R.P.O.; Schmidt, O. Slurry 15 NH 4-N recovery in herbage and soil: Effects of application method and timing. Plant Soil 2010, 330, 357–368. [Google Scholar] [CrossRef] [Green Version]
- Nüsse, A.; Linsler, D.; Loges, R.; Reinsch, T.; Taube, F.; Ludwig, B. Effect of grassland harvesting frequency and N-fertilization on stocks and dynamics of soil organic matter in the temperate climate. Arch. Agron. Soil Sci. 2018, 64, 1925–1931. [Google Scholar] [CrossRef]
- Barnard, R.; Barthes, L.; Leadley, P.W. Short-term uptake of 15 N by a grass and soil micro-organisms after long-term exposure to elevated CO2. Plant Soil 2006, 280, 91–99. [Google Scholar] [CrossRef]
- Izaurralde, R.C.; Thomson, A.M.; Morgan, J.A.; Fay, P.A.; Polley, H.W.; Hatfield, J.L. Climate impacts on agriculture: Implications for forage and rangeland production. Agron. J. 2011, 103, 371–381. [Google Scholar] [CrossRef] [Green Version]
- Dukes, J.S.; Chiariello, N.R.; Cleland, E.E.; Moore, L.A.; Shaw, M.R.; Thayer, S.; Tobeck, T.; Mooney, H.A.; Field, C.B. Responses of grassland production to single and multiple global environmental changes. PLoS Biol. 2005, 3, e319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Egli, M.; Hitz, C.; Fitze, P.; Mirabella, A. Experimental determination of climate-change effects on above-ground and below-ground organic matter in alpine grasslands by translocation of soil cores. J. Plant Nutr. Soil Sci. 2004, 167, 457–470. [Google Scholar] [CrossRef]
- Soussana, J.-F.; Lemaire, G. Coupling carbon and nitrogen cycles for environmentally sustainable intensification of grasslands and crop-livestock systems. Agric. Ecosyst. Environ. 2014, 190, 9–17. [Google Scholar] [CrossRef]
- Harpole, W.S.; Tilman, D. Grassland species loss resulting from reduced niche dimension. Nature 2007, 446, 791–793. [Google Scholar] [CrossRef]
- Reich, P.B. The world-wide ‘fast–slow’plant economics spectrum: A traits manifesto. J. Ecol. 2014, 102, 275–301. [Google Scholar] [CrossRef]
- Kuzyakov, Y.; Xu, X. Competition between roots and microorganisms for nitrogen: Mechanisms and ecological relevance. New Phytol. 2013, 198, 656–669. [Google Scholar] [CrossRef]
- Inselsbacher, E.; Umana, N.H.-N.; Stange, F.C.; Gorfer, M.; Schüller, E.; Ripka, K.; Zechmeister-Boltenstern, S.; Hood-Novotny, R.; Strauss, J.; Wanek, W. Short-term competition between crop plants and soil microbes for inorganic N fertilizer. Soil Biol. Biochem. 2010, 42, 360–372. [Google Scholar] [CrossRef]
- Zistl-Schlingmann, M.; Feng, J.; Kiese, R.; Stephan, R.; Zuazo, P.; Willibald, G.; Wang, C.; Butterbach-Bahl, K.; Dannenmann, M. Dinitrogen emissions: An overlooked key component of the N balance of montane grasslands. Biogeochemistry 2019, 143, 15–30. [Google Scholar] [CrossRef] [Green Version]
- Hart, S.C.; Firestone, M.K.; Paul, E.A.; Smith, J.L. Flow and fate of soil nitrogen in an annual grassland and a young mixed-conifer forest. Soil Biol. Biochem. 1993, 25, 431–442. [Google Scholar] [CrossRef]
- Cheng, W.; Parton, W.J.; Gonzalez-Meler, M.A.; Phillips, R.; Asao, S.; McNickle, G.G.; Brzostek, E.; Jastrow, J.D. Synthesis and modeling perspectives of rhizosphere priming. New Phytol. 2014, 201, 31–44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dijkstra, F.A.; Carrillo, Y.; Pendall, E.; Morgan, J.A. Rhizosphere priming: A nutrient perspective. Front. Microbiol. 2013, 4, 216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernhardt-Römermann, M.; Römermann, C.; Sperlich, S.; Schmidt, W. Explaining grassland biomass—The contribution of climate, species and functional diversity depends on fertilization and mowing frequency. J. Appl. Ecol. 2011, 48, 1088–1097. [Google Scholar] [CrossRef] [Green Version]
- Bimüller, C.; Dannenmann, M.; Tejedor, J.; von Lützow, M.; Buegger, F.; Meier, R.; Haug, S.; Schroll, R.; Kögel-Knabner, I. Prolonged summer droughts retard soil N processing and stabilization in organo-mineral fractions. Soil Biol. Biochem. 2014, 68, 241–251. [Google Scholar] [CrossRef]
- Ammann, C.; Spirig, C.; Leifeld, J.; Neftel, A. Assessment of the nitrogen and carbon budget of two managed temperate grassland fields. Agric. Ecosyst. Environ. 2009, 133, 150–162. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Intensive | Extensive | |||
|---|---|---|---|---|
| Date | HE | LE | HE | LE |
| 23.05./08.06.17 | 2.2 ± 0.2 a | 3.8 ± 0.4 b | 3.7 ± 0.4 a | 3.7 ± 0.4 a |
| 05.07.17 | 3.1 ± 0.4 a | 2.4 ± 0.2 b | ||
| 07.08.17 | 1.8 ± 0.1 a | 1.9 ± 0 a | 2.8 ± 0.2 a | 3.0 ± 0.6 a |
| 21.09.17 | 2.1 ± 0.1 a | 1.9 ± 0.3 a | ||
| 08.11.17 | 0.6 ± 0.3 a | 1.5 ± 0.9 a | 0.9 ± 0.2 a | 1.6 ± 0.5 a |
| Sum | 9.8 ± 0.6 Aa | 11.6 ± 0.7 Ab | 7.5 ± 0.5 Ba | 8.3 ± 1.2 Ba |
| Intensive | Extensive | |||
|---|---|---|---|---|
| HE | LE | HE | LE | |
| N concentration | 2.3 ± 0.3 Aa | 2.1 ± 0.1 Aa | 1.7 ± 0.1 Ba | 1.6 ± 0.1 Ba |
| Intensive | Extensive | |||
|---|---|---|---|---|
| Year | HE | LE | HE | LE |
| 2012 | 291 ± 44 Aa | 316 ± 46 Aa | 165 ± 31 Ba | 167 ± 37 Ba |
| 2013 | 365 ± 13 Aa | 416 ± 26 Aa | 210 ± 40 Ba | 244 ± 28 Ba |
| 2014 | 310 ± 10 Aa | 283 ± 14 Aa | 157 ± 22 Ba | 156 ± 16 Ba |
| 2015 | 189 ± 7 Aa | 219 ± 4 Ab | 169 ± 7 B | 130 ± n.a. B |
| 2016 | n.a. | n.a. | n.a. | n.a |
| 2017 | 213 ± 25 Aa | 230 ± 26 Aa | 128 ± 16 Ba | 127 ± 28 Ba |
| Management | Site | N from Fertilizer (%) | N from Other Sources (%) |
|---|---|---|---|
| Intensive | HE | 10.9 ± 0.8 Aa | 89.1 |
| LE | 10.6 ± 0.6 Aa | 89.4 | |
| Extensive | HE | 3.1 ± 0.3 Ba | 96.9 |
| LE | 2.8 ± 0.3 Ba | 97.2 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zistl-Schlingmann, M.; Kwatcho Kengdo, S.; Kiese, R.; Dannenmann, M. Management Intensity Controls Nitrogen-Use-Efficiency and Flows in Grasslands—A 15N Tracing Experiment. Agronomy 2020, 10, 606. https://doi.org/10.3390/agronomy10040606
Zistl-Schlingmann M, Kwatcho Kengdo S, Kiese R, Dannenmann M. Management Intensity Controls Nitrogen-Use-Efficiency and Flows in Grasslands—A 15N Tracing Experiment. Agronomy. 2020; 10(4):606. https://doi.org/10.3390/agronomy10040606
Chicago/Turabian StyleZistl-Schlingmann, Marcus, Steve Kwatcho Kengdo, Ralf Kiese, and Michael Dannenmann. 2020. "Management Intensity Controls Nitrogen-Use-Efficiency and Flows in Grasslands—A 15N Tracing Experiment" Agronomy 10, no. 4: 606. https://doi.org/10.3390/agronomy10040606
APA StyleZistl-Schlingmann, M., Kwatcho Kengdo, S., Kiese, R., & Dannenmann, M. (2020). Management Intensity Controls Nitrogen-Use-Efficiency and Flows in Grasslands—A 15N Tracing Experiment. Agronomy, 10(4), 606. https://doi.org/10.3390/agronomy10040606