Design of a Real-Time Gas-Exchange Measurement System for Crop Stands in Environmental Scenarios


Abstract
:1. Introduction
2. Materials and Methods
2.1. CO2 and H2O Gas Exchange of Complete Crops
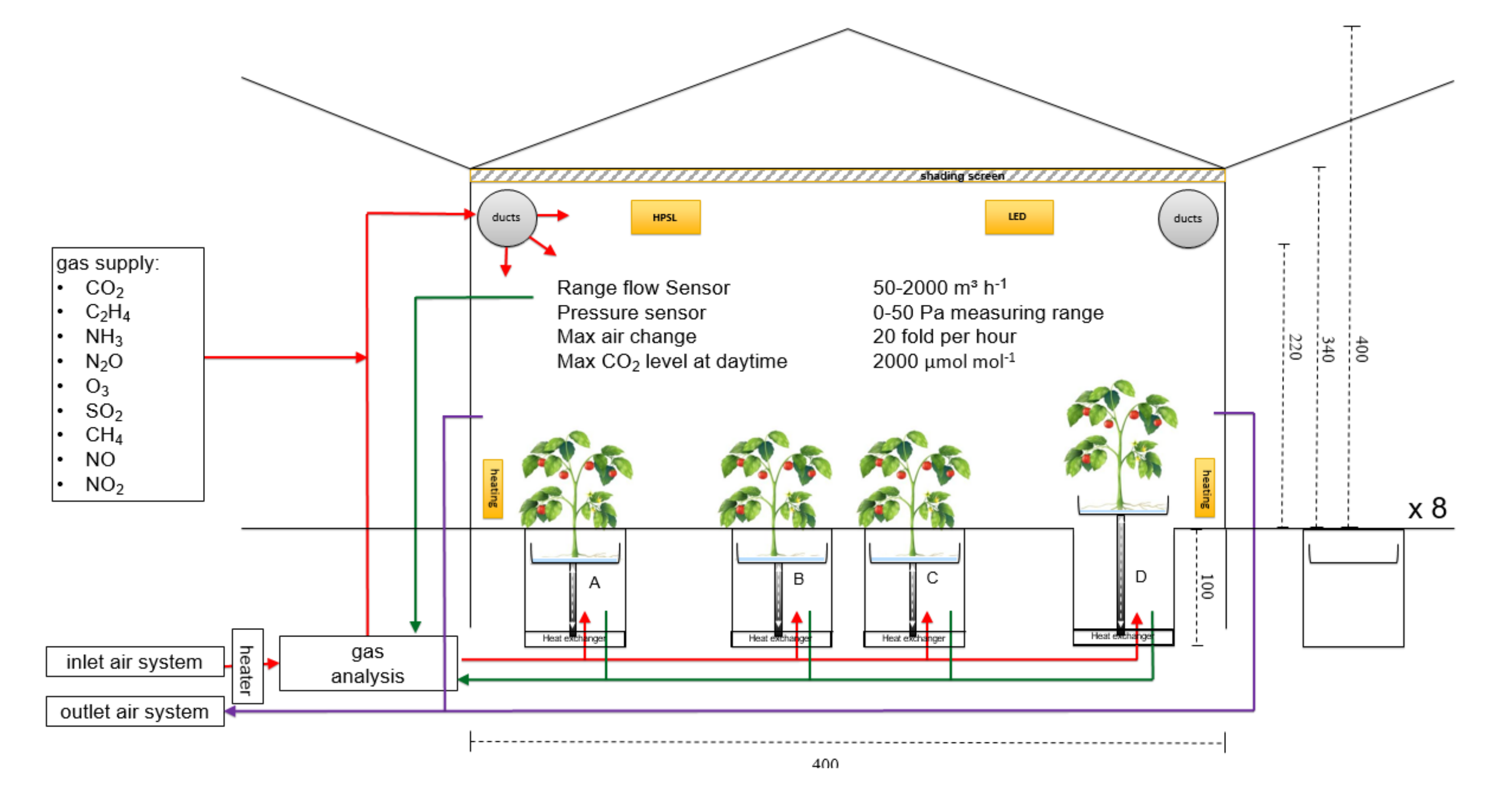
2.2. Configuration Measuring CO2 Gas Exchange
2.3. Calculation of the CO2 and H2O Gas Exchange
2.4. Removing Heat and Humidity from the Cabin Air
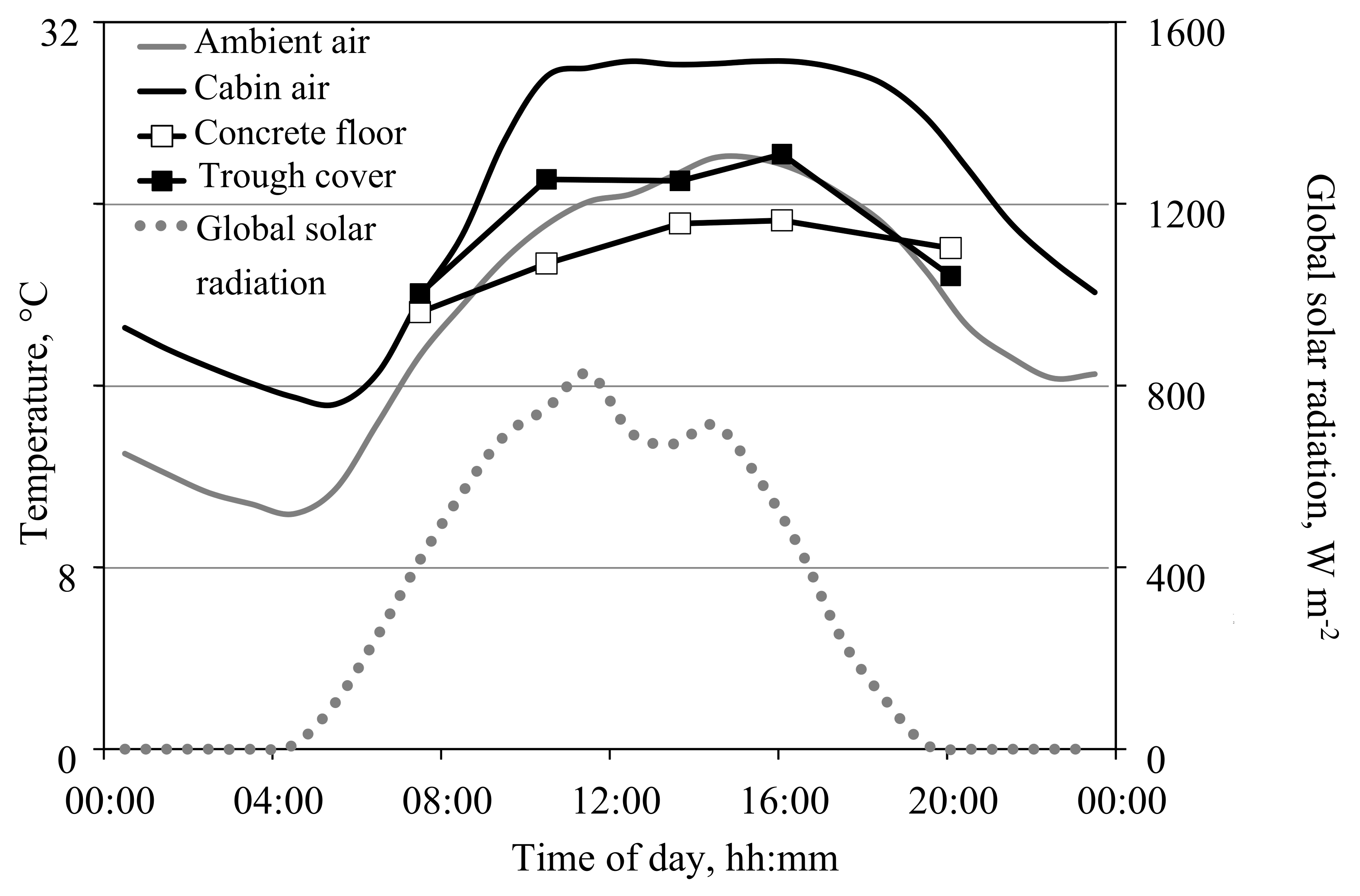
2.5. Environmental Constraints for Gas Exchange Measurements
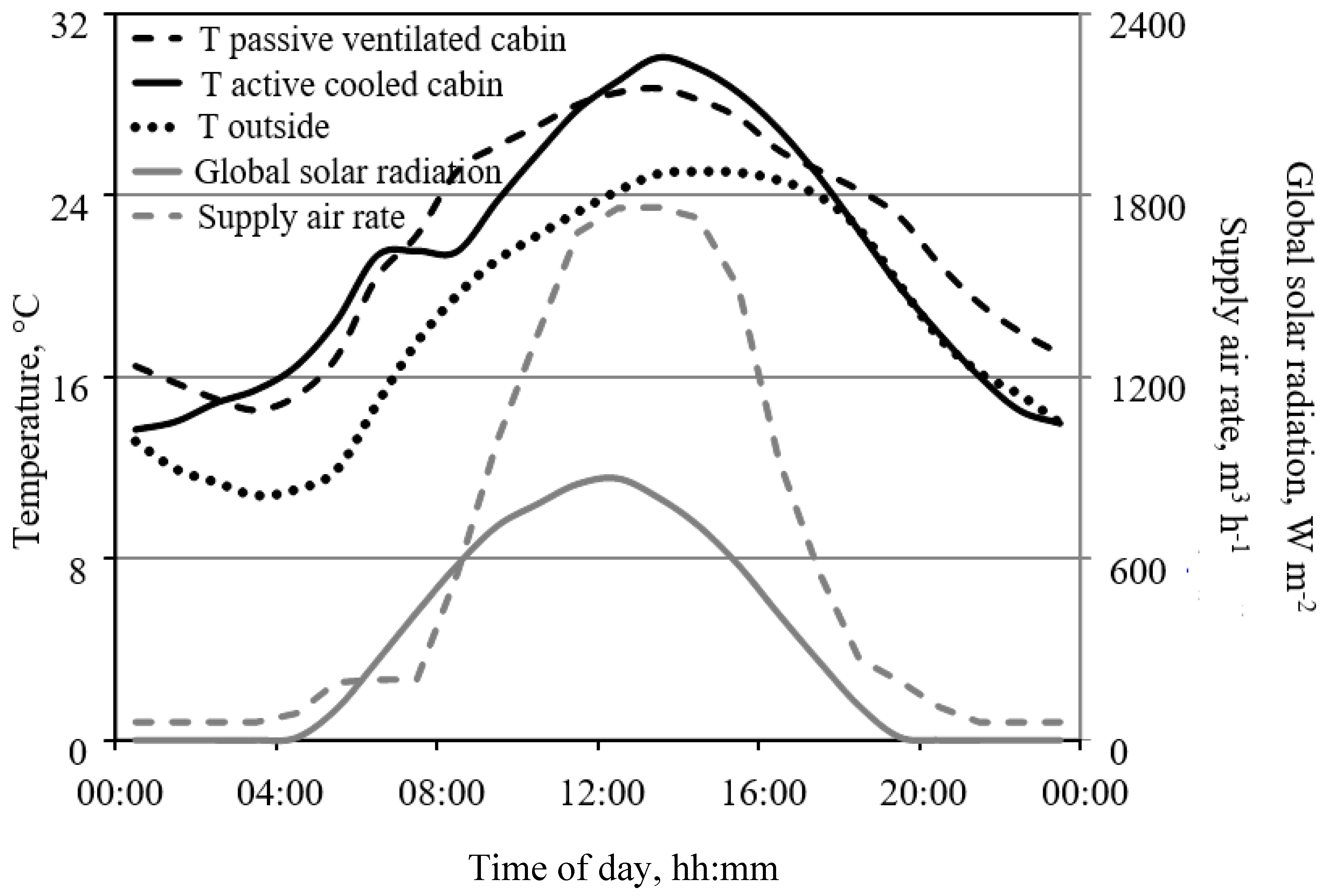
2.6. Air Exchange Rates of Closed Cabins and CO2 Sources
2.7. Controlling the Cabin Climate and Other Possible Treatments of the Plants
2.8. Separating the Gas Exchange of the Shoot and of the Root Zone
2.9. Crop Experiments
2.10. Comparing Measured and Simulated Crop Photosynthesis
3. Results and Discussion
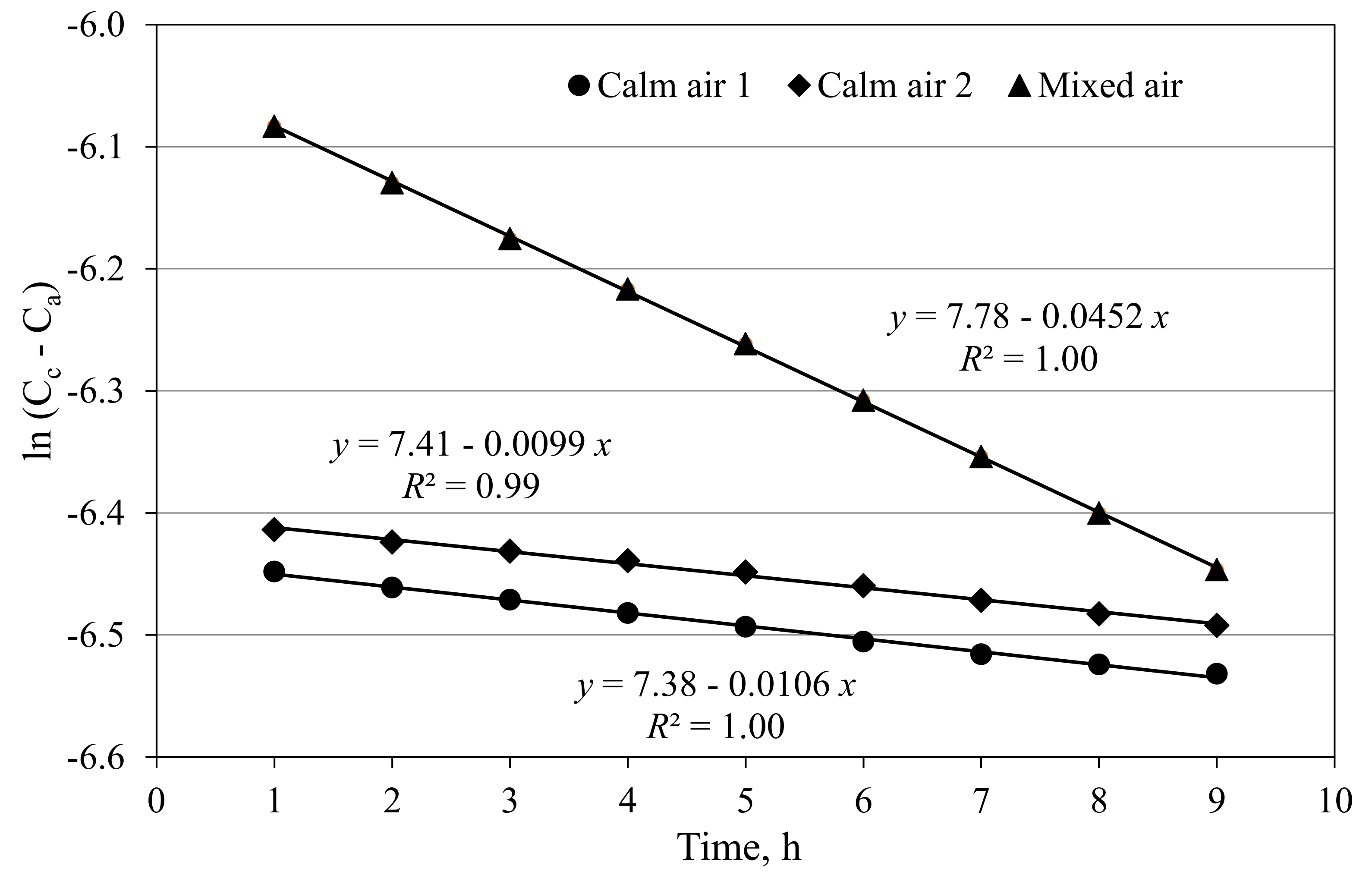
3.1. Air Exchange Rates of Closed Cabins
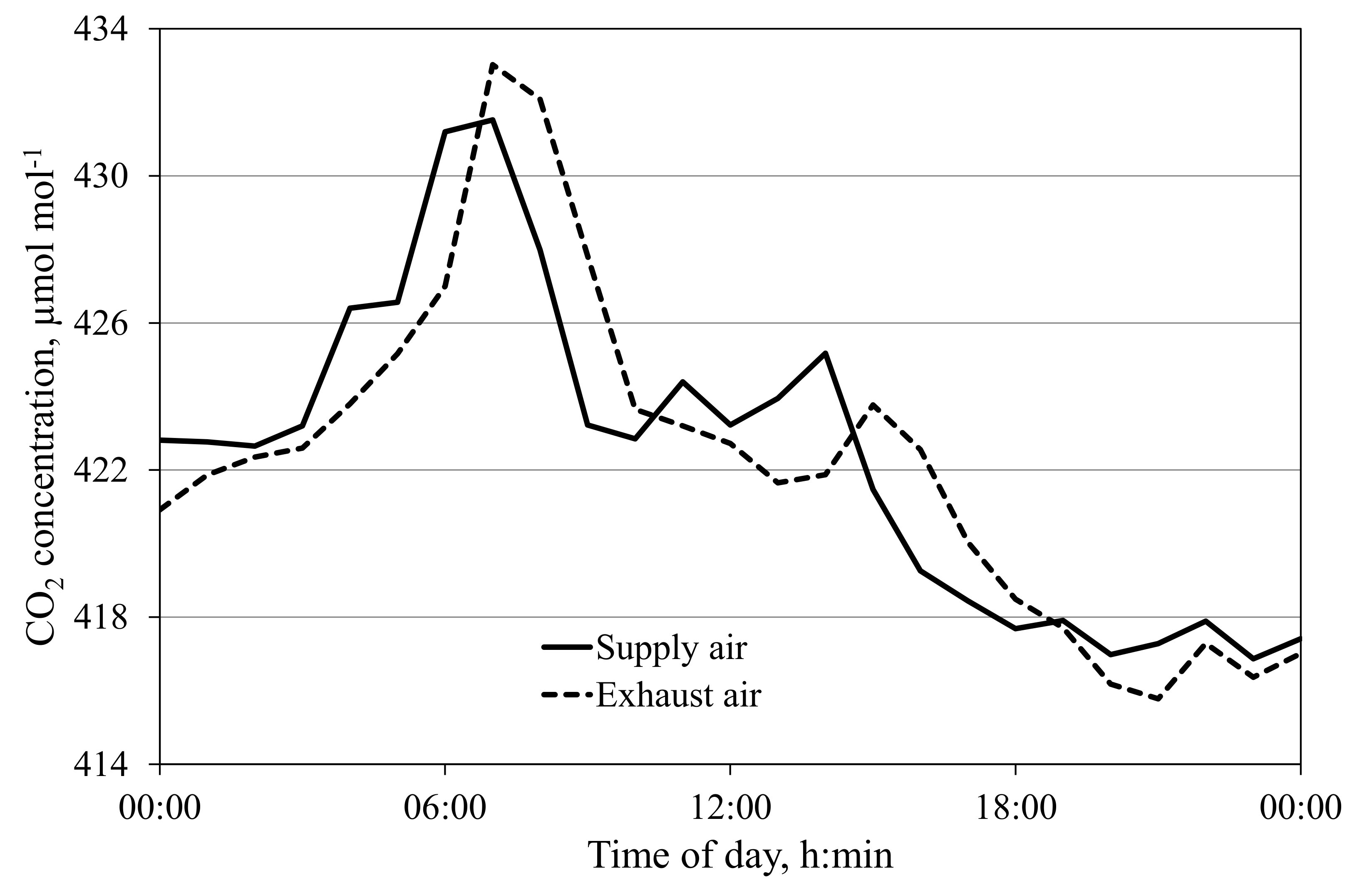
3.2. CO2 Sources and Sinks in the Cabins
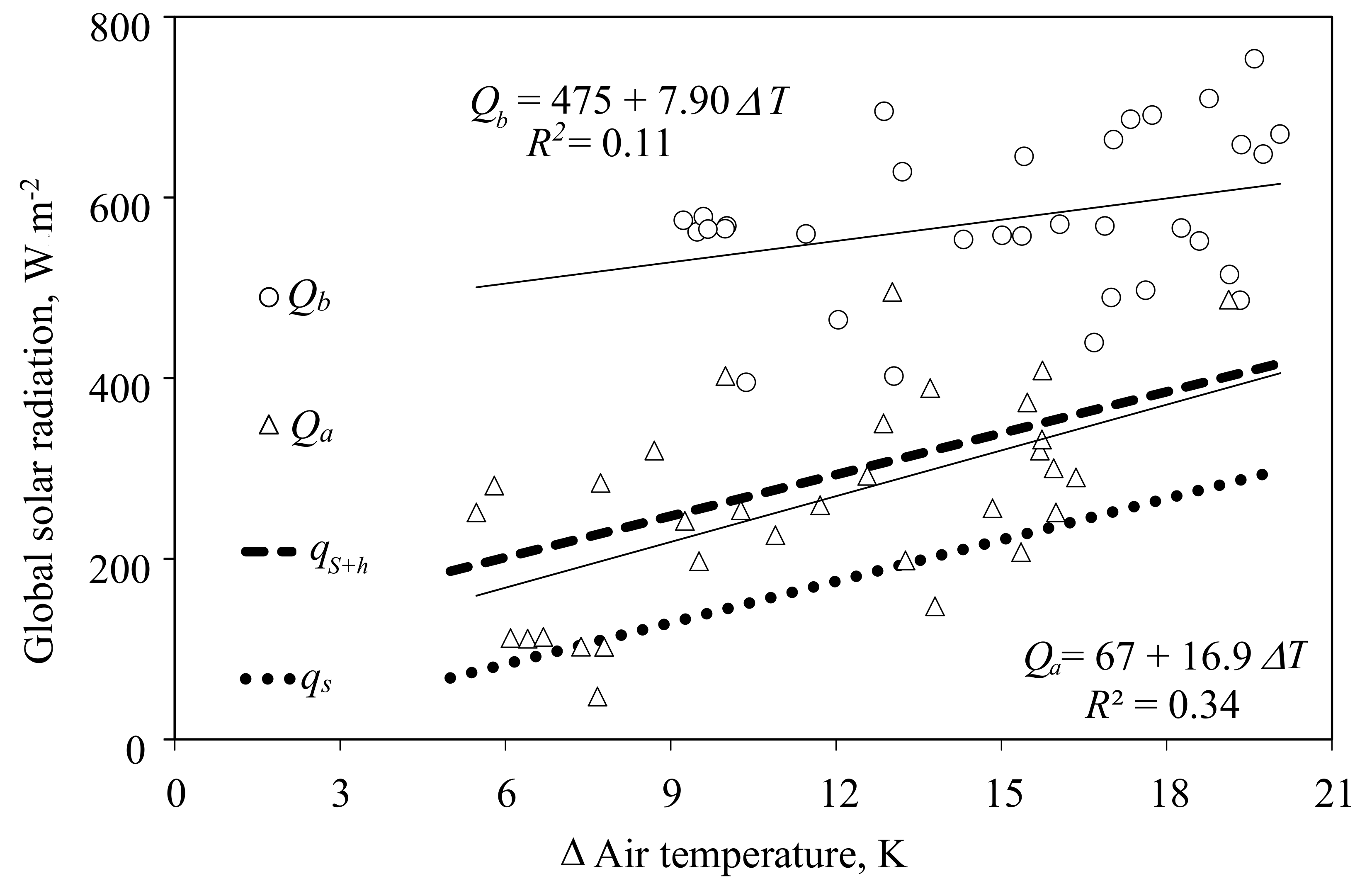
3.3. Possible Ranges of Environmental Conditions during the Measurements
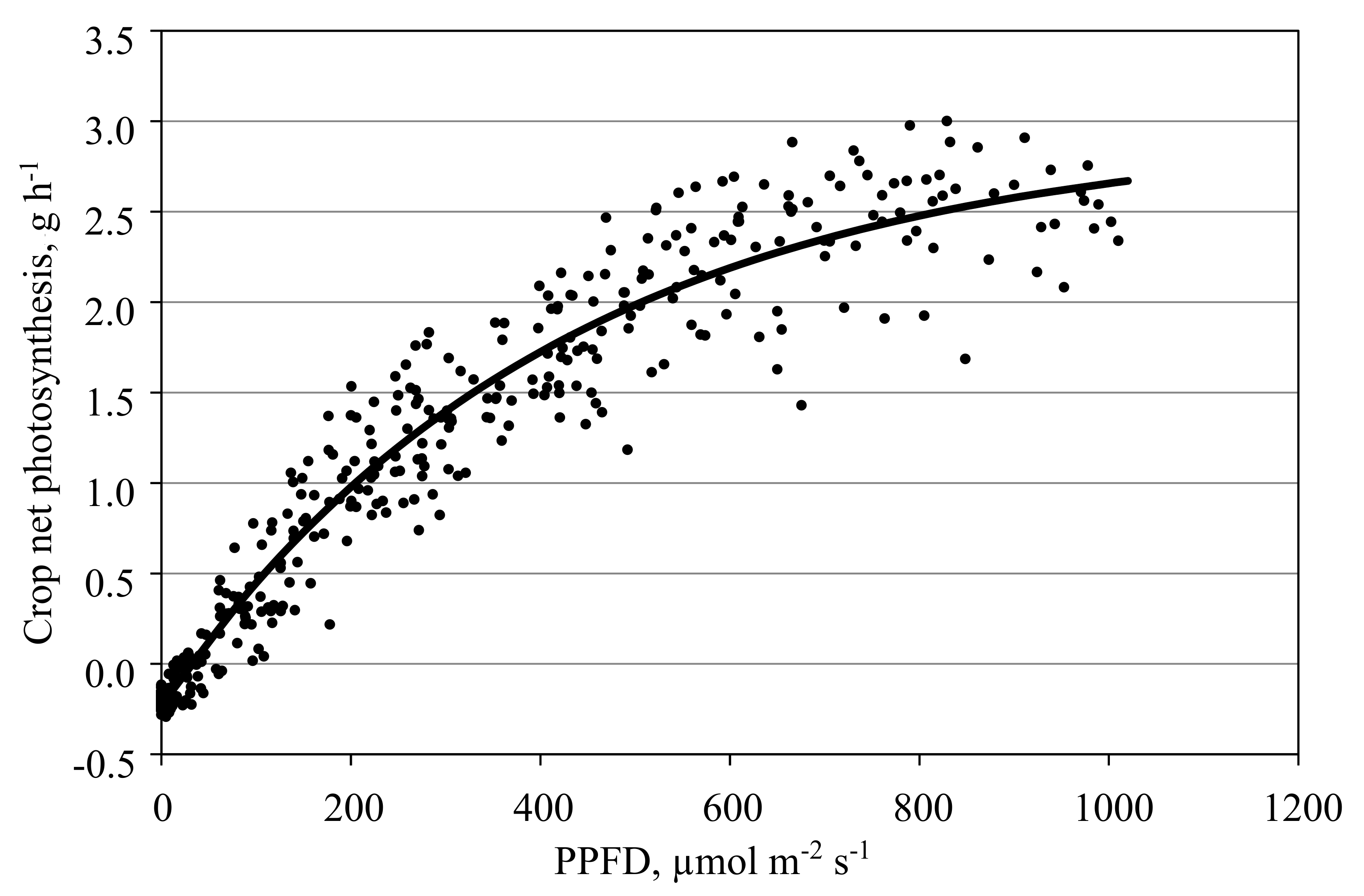
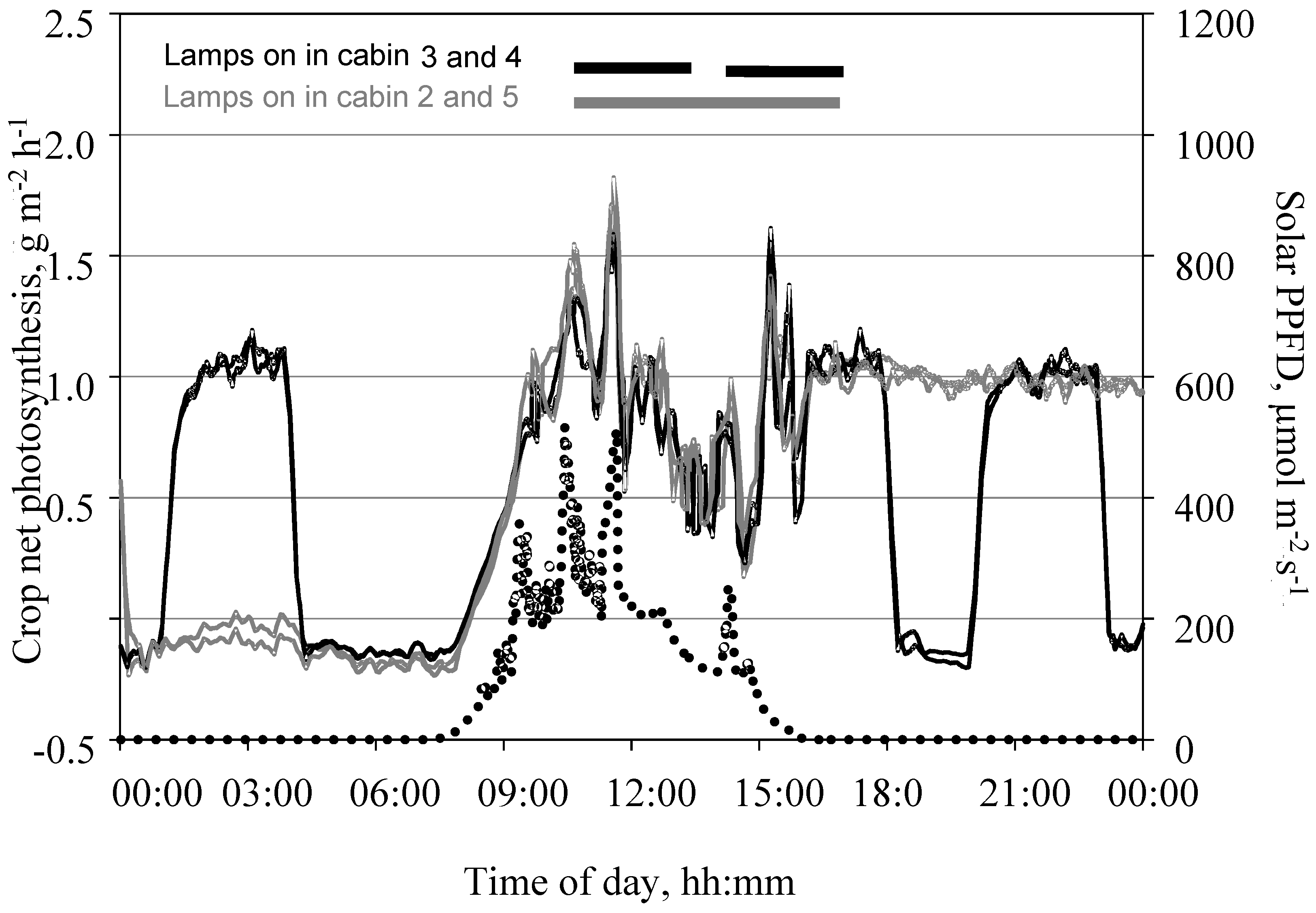
3.4. Measurement of Photosynthesis in the Closed Chamber Mode
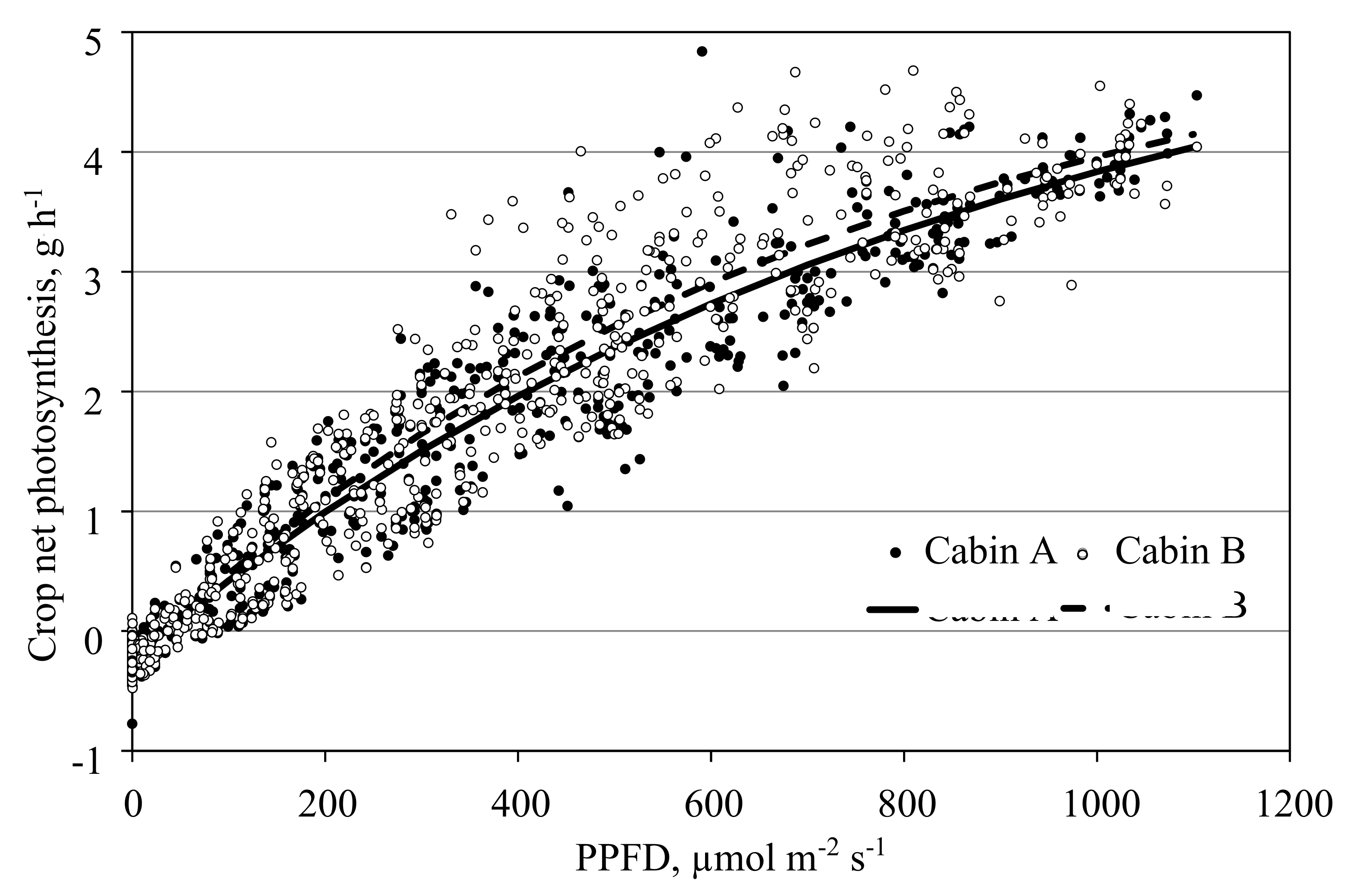
3.5. Measurement of Photosynthesis in the Open Chamber Mode
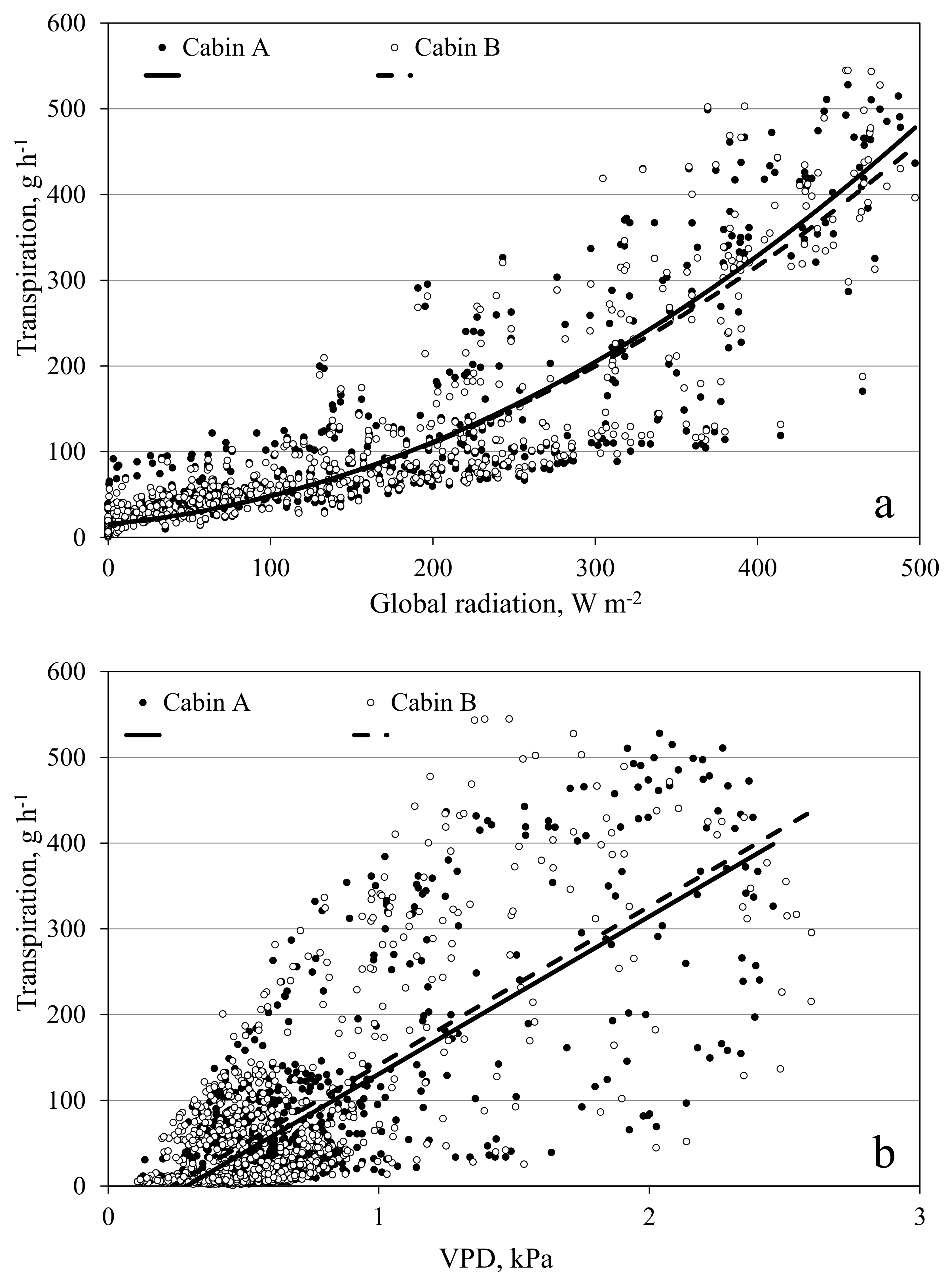
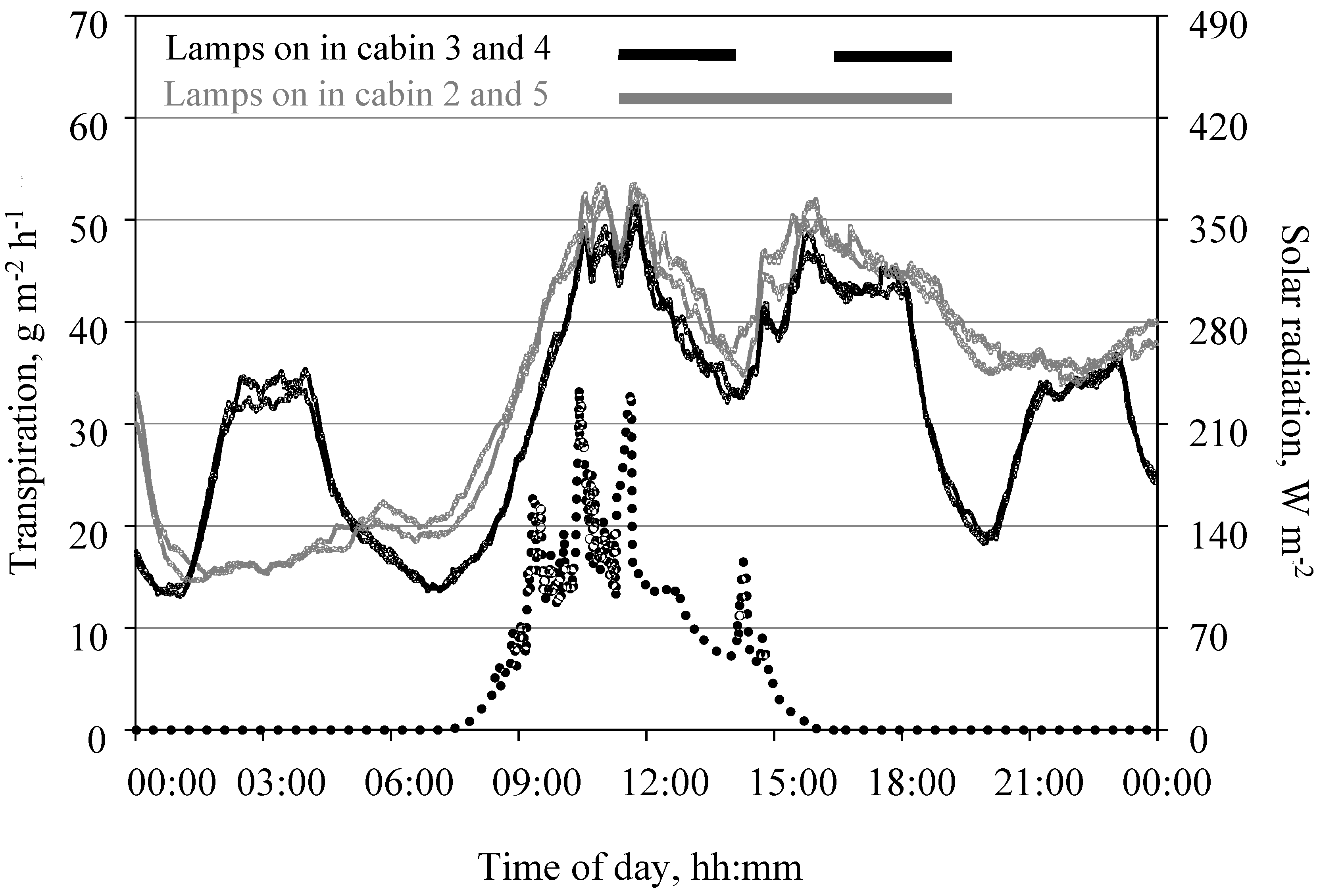
3.6. Measurement of Transpiration in the Open Chamber Mode
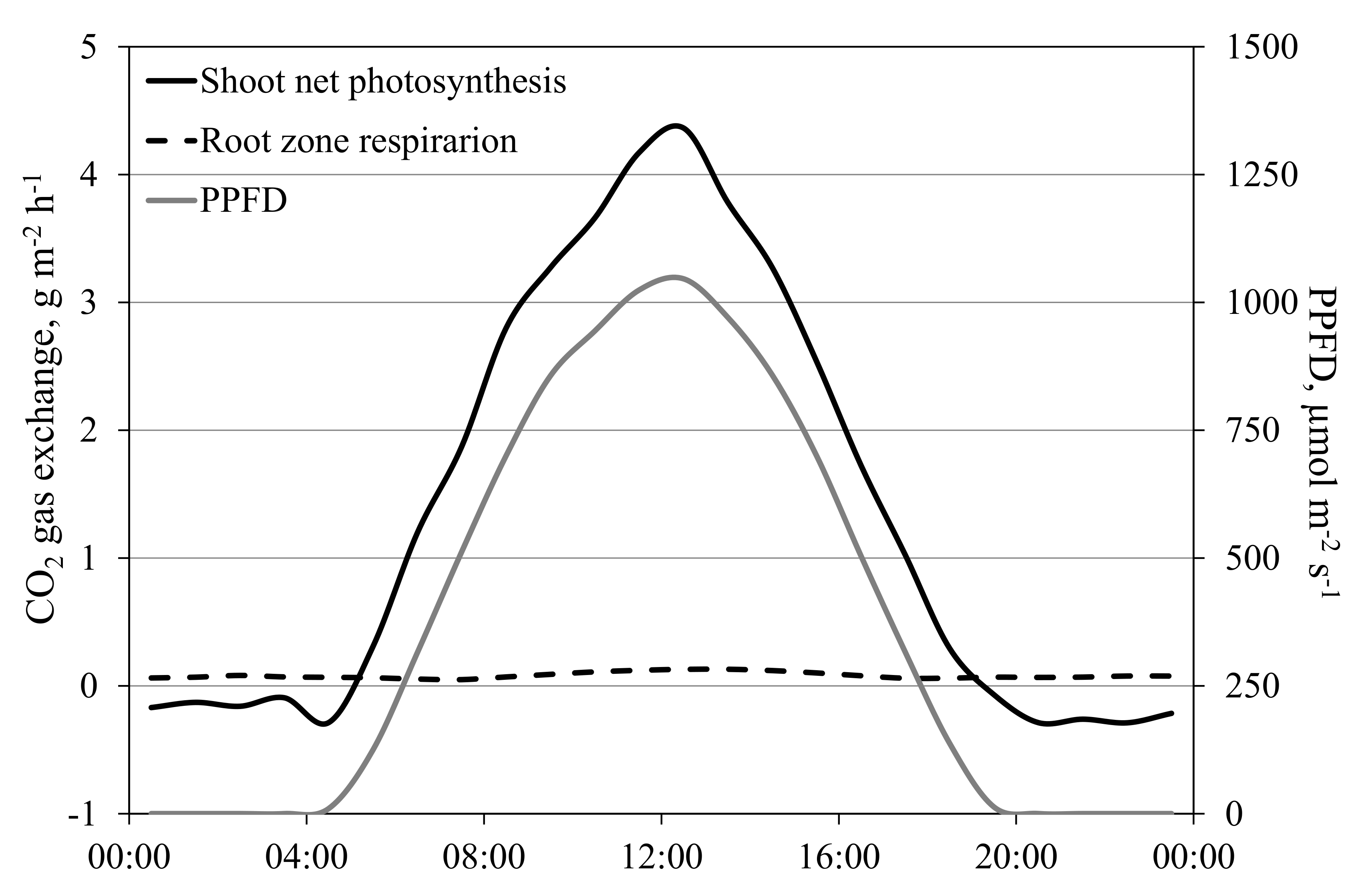
3.7. Separating the Gas Exchange of the Shoot and of the Root Zone
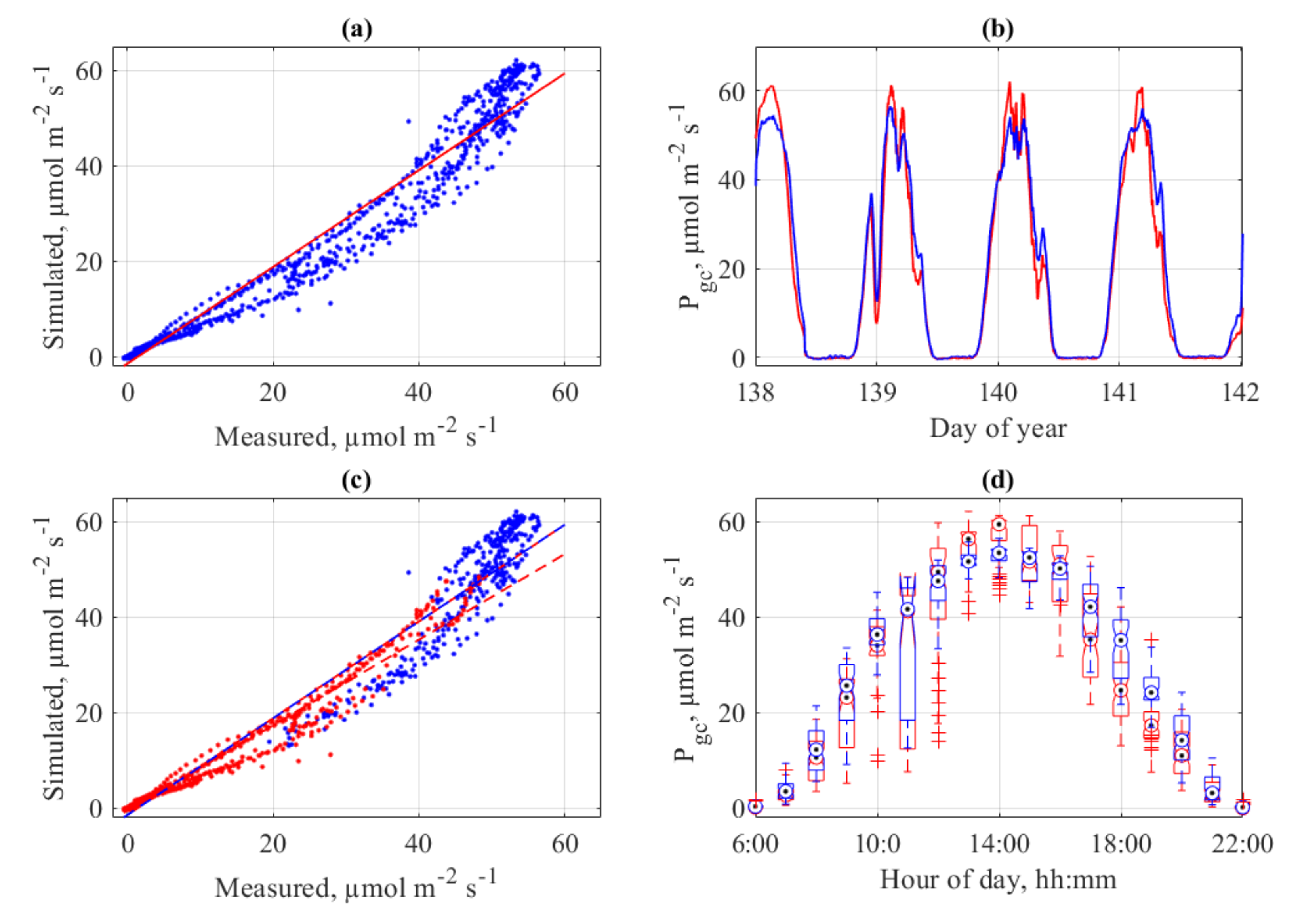
3.8. Models and Measurements
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Fanourakis, D.; Hyldgaard, B.; Giday, H.; Bouranis, D.; Körner, O.; Nielsen, K.L.; Ottosen, C.O. Differential effects of elevated air humidity on stomatal closing ability of Kalanchoë blossfeldiana among the C3 and CAM states. Environ. Exp. Bot. 2017, 143, 115–124. [Google Scholar] [CrossRef]
- Matteucci, G.; Manca, G. Measuring CO2 exchange at canopy scale: The eddy covariance technique. In Terrestrial Photosynthesis in a Changing Environment: A Molecular, Physiological, and Ecological Approach; Flexas, J., Loreto, F., Medrano, H., Eds.; Cambridge University Press: Cambridge, UK, 2012; pp. 206–218. [Google Scholar] [CrossRef]
- Baldocchi, D.D. Assessing the eddy covariance technique for evaluating carbon dioxide exchange rates of ecosystems: Past, present and future. Glob. Chang. Biol. 2003, 9, 479–492. [Google Scholar] [CrossRef] [Green Version]
- Körner, O.; Van Straten, G. Decision support for dynamic greenhouse climate control strategies. Comput. Electron. Agric. 2008, 60, 18–30. [Google Scholar] [CrossRef]
- Körner, O.; Challa, H. Process-based humidity control regime for greenhouse crops. Comput. Electron. Agric. 2003, 39, 173–192. [Google Scholar] [CrossRef]
- Heissner, A. Der CO2-Gaswechsel von Paprikapflanzen in Abhängigkeit von der Bestrahlungsstärke, der CO2-Konzentration, der Lufttemperatur und dem Dampfdrucksättigungsdefizit der Luft: Messungen und Modell. (CO2 gas-exchange of sweet-pepper plants depending on irradiation, CO2 concentration, air temperature and vapour pressure deficit: Measurements and model). Gartenbauwissenschaft 1997, 62, 78–90. [Google Scholar]
- Jensen, T.J.; Rosenqvist, E.; Aaslyng, J.M. A daylight climate chamber for testing greenhouse climate control strategies and calculating canopy carbon dioxide exchange. HortTechnology 2006, 16, 191–198. [Google Scholar] [CrossRef] [Green Version]
- Van Iersel, M.W.; Bugbee, B. A multiple chamber, semicontinuous, crop carbon dioxide exchange system: Design, calibration, and data interpretation. J. Am. Soc. Hortic. Sci. 2000, 125, 86–92. [Google Scholar] [CrossRef] [Green Version]
- Aaslyng, J.M.; Lund, J.B.; Ehler, N.; Rosenqvist, E. Intelligrow: A greenhouse component-based climate system. Environ. Model. Softw. 2003, 18, 657–666. [Google Scholar] [CrossRef]
- Kläring, H.-P.; Hauschild, I.; Heißner, A. Fruit removal increases root-zone respiration in cucumber. Ann. Bot. 2014, 114, 1735–1745. [Google Scholar] [CrossRef]
- Barthel, M.; Hammerle, A.; Sturm, P.; Baur, T.; Gentsch, L.; Knohl, A. The diel imprint of leaf metabolism on the d13C signal of soil respiration under control and drought conditions. New Phytol. 2011, 192, 925–938. [Google Scholar] [CrossRef]
- Nederhoff, E.M.; Vegter, J.G. Photosynthesis of stands of tomato, cucumber and sweet pepper measured in greenhouses under various CO2-concentrations. Ann. Bot. 1994, 73, 353–361. [Google Scholar] [CrossRef]
- Körner, O.; Van ’t Ooster, A.; Hulsbos, M. Design and performance of a measuring system for CO2 exchange of a greenhouse crop at different light levels. Biosyst. Eng. 2007, 97, 219–228. [Google Scholar] [CrossRef]
- Kläring, H.-P.; Klopotek, Y.; Krumbein, A.; Schwarz, D. The effect of reducing the heating set point on the photosynthesis, growth, yield and fruit quality in greenhouse tomato production. Agric. For. Meteorol. 2015, 214, 178–188. [Google Scholar] [CrossRef]
- Körner, O.; Heuvelink, E.; Niu, Q. Quantification of temperature, CO2, and light effects on crop photosynthesis as a basis for model-based greenhouse climate control. J. Hortic. Sci. Biotechnol. 2009, 84, 233–239. [Google Scholar] [CrossRef]
- Hand, D.W.; Clark, G.; Hannah, M.A.; Thornley, J.H.M.; Warren-Wilson, J.; Wilson, J.W. Measuring the canopy net photosynthesis of glasshouse crops. J. Exp. Bot. 1992, 43, 375–381. [Google Scholar] [CrossRef]
- Teitel, M.; Atias, M.; Schwartz, A.; Cohen, S. Use of a greenhouse as an open chamber for canopy gas exchange measurements: Methodology and validation. Agric. For. Meteorol. 2011, 151, 1346–1355. [Google Scholar] [CrossRef]
- Lake, J.V. Measurement and control of the rate of carbon dioxide assimilation by glasshouse crops. Nature 1966, 209, 97–98. [Google Scholar] [CrossRef]
- Nederhoff, E.; Vegter, J.G. Canopy photosynthesis of tomato, cucumber and sweet pepper in greenhouses: Measurements compared to models. Ann. Bot. 1994, 73, 421–427. [Google Scholar] [CrossRef]
- Nett, L.; Hauschild, I.; Kläring, H.P. Carbon sink reduction by fruit removal triggers respiration but not nitrous oxide emissions from the root zone of cucumber. Ann. Appl. Biol. 2019, 175, 111–118. [Google Scholar] [CrossRef]
- De Kreij, C.; Voogt, W.; Van den Bos, A.L.; Baas, R. Nutrient Solutions for the Growth of Tomato in Closed Systems (In Dutch); Proefstation voor Bloemistrij en Glasgroenten: Naaldwijk, The Netherlands, 1997. [Google Scholar]
- Schwarz, D.; Kläring, H.P. Allometry to estimate leaf area of tomato. J. Plant. Nutr. 2001, 24, 1291–1309. [Google Scholar] [CrossRef]
- Körner, O. Crop Based Climate Regimes for Energy Saving in Greenhouse Cultivation. Ph.D. Thesis, Wageningen University, Wageningen, The Netherlands, 2003. [Google Scholar]
- Thornley, J.H.M. Mathematical Models in Plant Physiology; Academic Press (Inc.): London, UK, 1976; p. 318. [Google Scholar]
- Farquhar, G.D.; Von Caemmerer, S.; Berry, J.A. A biochemical model of photosynthetic CO2 assimilation in leaves of C3 species. Planta 1980, 149, 78–90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gijzen, H. Ontwikkeling van een simulatiemodel voor transpiratie en wateropname en van een integral gewasmodel. In Development of a Simulation Model for Transpiration and Water Uptake and an Integral Crop Model; AB-DLO: Wageningen, The Netherlands, 1994; p. 90. [Google Scholar]
- Goudriaan, J.; Van Laar, H.H.; Van Keulen, H.; Louwerse, W. Photosynthesis, CO2 and Plant Production. In Wheat Growth and Modelling; Day, W., Atkins, R.K., Eds.; NATO ASI, Springer Scientific: New York, NY, USA, 1985; Volume 86, pp. 107–122. [Google Scholar]
- Goudriaan, J.; Van Laar, H.H. Modelling Potential Crop Growth Processes; Kluwer: Dordrecht, The Netherlands, 1994; p. 238. [Google Scholar]
- Gijzen, H.; Goudriaan, J. A flexible and explanatory model of light distribution and photosynthesis in row crops. Agric. For. Meteorol. 1989, 48, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Körner, O. Evaluation of crop photosynthesis models for dynamic climate control. Acta Hortic. 2004, 654, 295–302. [Google Scholar] [CrossRef]
- Schuch, I. Methods for Energetic Evaluation of Closed Greenhouses Considering Enthalpy Differences; Winter-Industries GmbH: Berlin, Germany, 2014. (In German) [Google Scholar]
- Nederhoff, E.M.; De Graaf, R. Effects of CO2 on leaf conductance and canopy transpiration of greenhouse grown cucumber and tomato. J. Hortic. Sci. Biotechnol. 1993, 68, 925–937. [Google Scholar] [CrossRef]
- Haberle, K.H.; Reiter, I.; Patzner, K.; Heyne, C.; Matyssek, R. Switching the light off: A break in photosynthesis and sap flow of forest trees under total solar eclipse. Meteorol. Z. 2001, 10, 201–206. [Google Scholar] [CrossRef]
- Blessing, C.H.; Barthel, M.; Gentsch, L.; Buchmann, N. Strong coupling of shoot assimilation and soil respiration during drought and recovery periods in beech as indicated by natural abundance δ13C measurements. Front. Plant. Sci. 2016, 7, 1710. [Google Scholar] [CrossRef] [Green Version]
- Ramírez, T.; Kläring, H.P.; Körner, O. Does interruption of supplementary lighting affect the long-term carbon dioxide exchange of greenhouse tomato crops? Biosyst. Eng. 2019, 187, 69–80. [Google Scholar] [CrossRef]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | Ref. | Method | Gas Supply | Type | Loc. | Size, m2 | Crops |
|---|---|---|---|---|---|---|---|
| 1992 | [16] | Null-balance at ambient CO2 with proportional pure CO2 injection | Technically pure CO2 with mass flow controller | open | UK | 1 × 162 | Cucumber |
| 1994 | [12] | Air exchange rates measured online using nitrous oxide (N2O) as tracer gas. | Technically pure CO2 with mass flow controller | semi-open | NL | 4 × 192 | Cucumber Tomato Sweet pepper |
| 2007 | [13] | Proportional pure CO2 injection with max. 10.2 g m−2 h−1 to air conditioning unit for elevated CO2 | Technically pure CO2 with mass flow controller | semi-closed | NL | 2 × 44 | Tomato Chrysanthemum |
| 2011 | [17] | Outside air drawn through a dry cooling pad with ambient CO2 | No CO2 supply | open | ISR | 2 × 360 | Sweet pepper |
| 2015 | [14] | Supplying outside air through ducts in a range of 3 to 12 m3 m−2 h−1 to ambient CO2 | No CO2 supply | semi-open | D | 6 × 64 | Tomato |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kläring, H.-P.; Körner, O. Design of a Real-Time Gas-Exchange Measurement System for Crop Stands in Environmental Scenarios. Agronomy 2020, 10, 737. https://doi.org/10.3390/agronomy10050737
Kläring H-P, Körner O. Design of a Real-Time Gas-Exchange Measurement System for Crop Stands in Environmental Scenarios. Agronomy. 2020; 10(5):737. https://doi.org/10.3390/agronomy10050737
Chicago/Turabian StyleKläring, Hans-Peter, and Oliver Körner. 2020. "Design of a Real-Time Gas-Exchange Measurement System for Crop Stands in Environmental Scenarios" Agronomy 10, no. 5: 737. https://doi.org/10.3390/agronomy10050737
APA StyleKläring, H. -P., & Körner, O. (2020). Design of a Real-Time Gas-Exchange Measurement System for Crop Stands in Environmental Scenarios. Agronomy, 10(5), 737. https://doi.org/10.3390/agronomy10050737