Prickly Ash Seed Kernel: A New Bio-Fumigation Material Against Tobacco Black Shank


Abstract
:1. Introduction
2. Materials and Methods
2.1. Biological Materials
2.2. HS-SPME GC-MS Method
2.3. Chemical Composition of PSK Volatiles and Extracts
2.4. Mycelia Inhibition Effect of PSK Volatiles and Extracts
2.5. Pot Experiment
2.6. Effect of PSK on Soil Microbial Community
2.7. Data Analysis
3. Results and Discussion
3.1. Components of PSK Volatiles and Extracts
3.2. Antifungal Effects of PSK Against Phytophthora Nicotianae
3.3. Effects of PSK Fumigant on Plant Growth
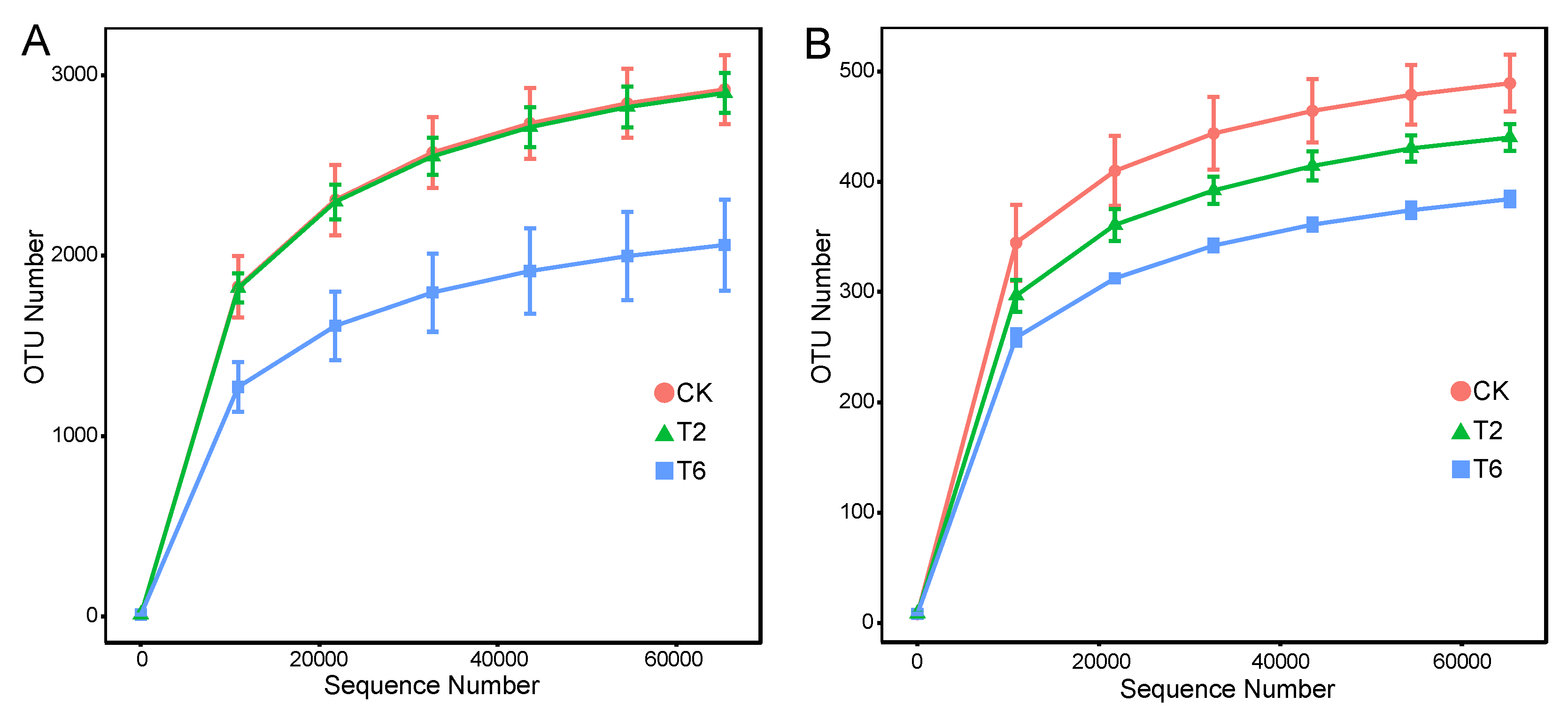
3.4. Effects of PSK Fumigant on Soil Microbial Community Richness and Diversity
3.5. Effect of PSK Fumigant on Microbial Structure
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
Appendix A



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compounds | Peak Area Percentage (%) | Identity (%) |
|---|---|---|
| Ethanone-1-(2-hydroxy-4,6-dimethoxyphenyl)- | 10.316749 | 94 |
| cis-9-Hexadecenoic acid | 6.208132 | 99 |
| n-Hexadecanoic acid | 15.668002 | 99 |
| Psilocin | 11.752521 | 90 |
| 9,12-Octadecadienoic acid (Z,Z)- | 15.248901 | 99 |
| cis-Vaccenic acid | 40.805695 | 99 |
| Addition of Prickly Ash Seed Kernel (g) | Inhibition Rate (%) |
|---|---|
| 1 | 35.19±0.99 d |
| 2 | 45.91±1.02 c |
| 4 | 52.05±0.87 b |
| 6 | 61.60±0.85 a |
References
- Panabières, F.; Ali, G.S.; Allagui, M.B.; Dalio, R.J.D.; Gudmestad, N.C.; Kuhn, M.; Roy, S.G.; Schena, L.; Zampounis, A. Phytophthora nicotianae diseases worldwide: New knowledge of a long-recognised pathogen. Phytopatho. Mediterr. 2016, 55, 20–40. [Google Scholar]
- Wang, H.C.; Chen, X.J.; Cai, L.T.; Cao, Y.; Lu, N.; Xia, H.Q.; Wang, M.S.; Shang, S.H. Race distribution and distribution of sensitivities to mefenoxam among isolates of Phytophthora parasitica var. nicotianae in Guizhou province of China. Crop Prot. 2013, 52, 136–140. [Google Scholar] [CrossRef]
- Reddy, P.P. Biofumigation. In Agro-ecological Approaches to Pest Management for Sustainable Agriculture; Springer: Singapore, 2017; pp. 43–59. [Google Scholar]
- Warmington, R.; Clarkson, J. Volatiles from biofumigant plants have a direct effect on carpogenic germination of sclerotia and mycelial growth of Sclerotinia sclerotiorum. Plant Soil 2016, 401, 213–229. [Google Scholar] [CrossRef] [Green Version]
- Goates, B.J.; Mercier, J. Control of common bunt of wheat under field conditions with the biofumigant fungus Muscodor albus. Eur. J. Plant Pathol. 2011, 131, 403–407. [Google Scholar] [CrossRef]
- Hansena, Z.R.; Keinath, A.P. Increased pepper yields following incorporation of biofumigation cover crops and the effects on soilborne pathogen populations and pepper diseases. Appl. Soil Ecol. 2013, 63, 67–77. [Google Scholar] [CrossRef]
- Madhavi, G.B.; Uma, D.G.; Kumar, V.K.; Babu, R.T.; Naidu, T.C.M. Evaluation of different brassica species and onion for their biofumigation effect against Rhizoctonia solani f. sp. sasakii in vitro. J. Res. Angrau 2015, 43, 22–28. [Google Scholar]
- Gong, B.; Bloszies, S.; Li, X.; Wei, M.; Yang, F.J.; Shi, Q.H.; Wang, X.F. Efficacy of garlic straw application against root-knot nematodes on tomato. Sci. Hortic. 2013, 161, 49–57. [Google Scholar] [CrossRef]
- Naz, I.; Palomares-Rius, J.E.; Khan, S.M.; Ali, S.; Ahmad, M.; Ali, A.; Khan, A. Control of Southern root knot nematode Meloidogyne incognita (Kofoid and White) Chitwood on tomato using green manure of Fumaria parviflora Lam (Fumariaceae). Crop Prot. 2015, 67, 121–129. [Google Scholar] [CrossRef]
- Rosa, M.; Luca, L.; D’Avinoc, L.; Gilda, P. Nitrogen and carbon mineralization in soils amended with biofumigant or non-biofumigant plant materials. Ind. Crop. Prod. 2015, 75, 65–72. [Google Scholar]
- He, C.; Gong, B.; Wang, C.; Wang, X.; Wei, M.; Yang, F.; Li, Y.; Shi, Q. Studies on root-knot nematode control effect and its mechanism by biofumigation with Zanthoxylum bungeanum seed meal in tomato seedlings. Chin. Veg. 2016, 6, 64–70. (in Chinese). [Google Scholar]
- Zhang, C.; Gao, J.; Han, T.; Tian, X.; Wang, F. Integrated control of tobacco black shank by combined use of riboflavin and Bacillus subtilis strain Tpb55. BioControl 2017, 62, 835–845. [Google Scholar] [CrossRef]
- Dong, X. Study on the Extraction, Antioxidant and Antibacterial Activities of the Active Substances from Zanthoxylum bungeanum Seeds; Sichuan Agricultural University: Chengdu, China, 2016. (in Chinese) [Google Scholar]
- Wang, C.; You, C.; Yang, K.; Guo, S.; Geng, Z.; Fan, L.; Du, S.; Deng, Z.; Wang, Y. Antifeedant activities of methanol extracts of four Zanthoxylum species and benzophenanthridines from stem bark of Zanthoxylum schinifolium against Tribolium castaneum. Ind. Crop. Prod. 2015, 74, 407–411. [Google Scholar] [CrossRef]
- Han, X.; Zhao, J.; Cao, J.; Zhang, C. Essential oil of Chrysanthemum indicum L.: Potential biocontrol agent against plant pathogen Phytophthora nicotianae. Environ. Sci. Pollut. Res. 2019, 26, 7013–7023. [Google Scholar] [CrossRef] [PubMed]
- Han, T.; You, C.; Zhang, L.; Feng, C.; Zhang, C.; Wang, J.; Kong, F. Biocontrol potential of antagonist Bacillus subtilis Tpb55 against tobacco black shank. BioControl 2016, 61, 195–205. [Google Scholar] [CrossRef]
- Bao, S.D. Soil Agricultural Chemistry Analysis; China Agricultural Publishing House: Beijing, China, 2000. (In Chinese) [Google Scholar]
- Fabienne, G. Comparative 16S rDNA and 16S rRNA sequence analysis indicates that Actinobacteria might be a dominant part of the metabolically active bacteria in heavy metal- contaminated bulk and rhizosphere soil. Environ. Microbiol. 2003, 10, 896–907. [Google Scholar]
- Bokulich, N.A.; Subramanian, S.; Faith, J.J.; Gevers, D.; Gordon, J.I.; Knight, R.; Mills, D.A.; Caporaso, J.G. Quality-filtering vastly improves diversity estimates from Illumina amplicon sequencing. Nat. Methods 2013, 10, 57–59. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Pena, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [Green Version]
- Edgar, R.C.; Haas, B.J.; Clemente, J.C.; Quince, C.; Knight, R. UCHIME improves sensitivity and speed of chimera detection. Bioinformatics 2011, 27, 2194–2200. [Google Scholar] [CrossRef] [Green Version]
- Hammer, K.A.; Carson, C.F.; Riley, T.V. Antifungal activity of the components of Melaleuca alternifolia (tea tree) oil. J. Appl. Microbiol. 2003, 95, 853–886. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Wang, Z.; Li, X.; Zhang, H.; Zhou, X.; Zhang, H. Analysis of volatile compounds in the pericarp of Zanthoxylum bungeanum Maxim. By ultrasonic nebulization extraction coupled with headspace single-drop microextraction and GC-MS. Chromatographia 2010, 71, 455–459. [Google Scholar] [CrossRef]
- Zhang, W.; Tan, S.; Yang, J.; Liao, Q.; Lan, J.; Lv, Y.; Tang, J. Comparison of volatile components in fresh and dried Zanthoxylum bungeanum Maxim. Food Sci. Biotechnol. 2019, 28, 1083–1092. [Google Scholar] [CrossRef] [PubMed]
- Fu, L.; Xie, H.; Shi, S. Multielement analysis of Zanthoxylum bungeanum Matim. Essential oil using ICP-MS/MS. Anal. Bioanal. Chem. 2018, 410, 3769–3778. [Google Scholar] [CrossRef] [PubMed]
- Sriwichai, T.; Sookwong, P.; Siddiqui, M.W.; Sommano, S.R. Aromatic profiling of Zanthoxyhum myriacanthum (makwhaen) essential oils from dried fruits using different initial drying techniques. Ind. Crop. Prod. 2019, 133, 284–291. [Google Scholar] [CrossRef]
- Boeck, P.; Leal, P.C.; Yunes, R.A.; Filho, V.C.; López, S.; Sortino, M.; Escalante, A.; Furlán, R.L.E.; Zacchino, S. Antifungal activity and studies on mode of action of novel xanthoxyline-derived chalcones. Arch. Pharm. Chem. Life Sci. 2005, 338, 87–95. [Google Scholar] [CrossRef] [PubMed]
- Wen, M.G.; Zhang, X.B.; Tian, J.N.; Ni, S.; Bian, H.; Lin, Y.; Liang, H. Binding interaction of xanthoxylin with bovine serum albumin. J. Solut. Chem. 2009, 38, 391–401. [Google Scholar] [CrossRef]
- Pinto, E.; Gonçalves, M.J.; Hrimpenga, K.; Pinto, J.; Vaz, S.; Vale-Silva, L.A.; Cavaleiro, C.; Salgueiro, L. Antifungal activity of the essential oil of Thymus villosus subsp. lusitanicus against Candida, Cryptococcus, Aspergillus and dermatophyte species. Ind. Crop. Prod. 2013, 51, 93–99. [Google Scholar] [CrossRef]
- Espinosa-García, F.J.; Langenheim, J.H. Effects of sabinene and γ-terpinene from coastal redwood leaves acting singly or in mixtures on the growth of some of their fungus endophytes. Biochem. Syst. Ecol. 1991, 19, 643–650. [Google Scholar] [CrossRef]
- Glamoclija, J.; Sokovic, M.; Grubisic, D.; Vukojevic, J.; Milinekovic, I.; Ristic, M. Antifungal activity of Critmum maritimum essential oil and its components against mushroom pathogen Mycogone perniciosa. Chem. Nat. Comp. 2009, 45, 96–97. [Google Scholar] [CrossRef]
- Tampieri, M.P.; Galuppi, R.; Macchioni, F.; Carelle, M.S.; Falcioni, L.; Cioni, P.L.; Morelli, I. The inhibition of Candida albicans by selected essential oils and their major components. Mycopathologia 2005, 159, 339–345. [Google Scholar] [CrossRef]
- D’Auria, F.D.; Tecca, M.; Strippoli, V.; Salvatore, G.; Battinelli, L.; Mazzanti, G. Antifungal activity of Lavandula angustifolia essential oil against Candida albicans yeast and mycelial form. Med. Mycol. 2005, 43, 391–396. [Google Scholar] [CrossRef] [Green Version]
- An, P.; Yang, X.; Yu, J.; Qi, J.; Ren, X.; Kong, Q. α-terpineol and terpene-4-ol, the critical components of tea tree oil, exert antifungal activities in vitro and in vivo against Aspergillus niger in grapes by inducing morphous damage and metabolic changes of fungus. Food Control 2019, 98, 42–53. [Google Scholar] [CrossRef]
- Rahmana, M.A.; Whitelaw-Weckerta, B.O. Consecutive applications of brassica green manures and seed meal enhances suppression of Meloidogyne javanica and increases yield of Vitis vinifera cv. Semillon. Appl. Soil Ecol. 2011, 47, 195–203. [Google Scholar] [CrossRef]
- Hollister, E.B.; Hu, P.; Wang, A.S.; Hons, F.M.; Gentry, T.J. Differential impacts of brassicaceous and nonbrassicaceous oilseed meals on soil bacterial and fungal communities. FEMS Microbiol. Immunol. 2012, 83, 632–641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weerakoon, D.M.N.; Reardon, C.L.; Paulitz, T.C.; Izzo, A.D. Long-term suppression of Pythium abappressorium induced by Brassica juncea seed meal amendment is biologically mediated. Soil Biol. Biochem. 2012, 51, 44–52. [Google Scholar] [CrossRef]
- Yim, B.; Hanschen, F.S.; Wrede, A.; Nitt, H.; Schreiner, M.; Smalla, K.; Winkelmann, T. Effects of biofumigation using Brassica juncea and Raphanus sativus in comparison to disinfection using Basamid on apple plant growth and soil microbial communities at three field sites with replant disease. Plant Soil 2016, 406, 389–408. [Google Scholar] [CrossRef]
- Ma, L.; Zheng, S.C.; Zhang, T.K.; Liu, Z.; Wang, X.; Zhou, X.; Yang, C.; Duo, J.; Mo, M. Effect of nicotine from tobacco root exudates on chemotaxis, growth, biocontrol efficiency, and colonization by Pseudomonas aeruginosa NXHG29. Antonie Leeuwenhoek 2018, 111, 1237–1257. [Google Scholar] [CrossRef]
- Hu, P.; Hollister, E.B.; Somenahally, A.C.; Hons, F.M.; Gentry, T.J. Soil bacterial and fungal communities respond differently to various isothiocyanates added for biofumigation. Front. Microbiol. 2014, 5, 729. [Google Scholar] [CrossRef]
- Kuthubutheen, A.J.; Pugh, G.J.F. The effects of fungicides on soil fungal populations. Soil Biol. Biochem. 1979, 11, 297–303. [Google Scholar] [CrossRef]
- Galletti, S.; Sala, E.; Leoni, O.; Burzi, P.L.; Cerato, C. Trichoderma spp. tolerance to Brassica carinata seed meal for a combined use in biofumigation. Biol. Control 2008, 45, 319–327. [Google Scholar] [CrossRef]
- Lidor, O.; Dror, O.; Hamershlak, D.; Shoshana, N.; Belausov, E.; Zahavi, T.; Mozes-Daube, N.; Naor, N.; Zchori-Fein, E.; Iasur-Kruhh, L.; et al. Introduction of a putative biocontrol agent into a range of phytoplasma- and liberibacter-susceptible crop plants. Pest Manag. Sci. 2018, 74, 811–819. [Google Scholar] [CrossRef]
- Malinich, E.A.; Bauer, C.E. The plant growth promoting bacterium Azospirillum brasilense is vertically transmitted in Phaseolus vulgaris (common bean). Symbiosis 2018, 76, 97–108. [Google Scholar] [CrossRef]





| Compounds | Retention Index | Identity (%) | Class | Peak Area Percentage (%) | |
|---|---|---|---|---|---|
| Literature | Experiment | ||||
| Hexanal | 800 | 799 | 98 | Ⅰ 2 | 1.16 |
| 1-Hexanol | 868 | 864 | 96 | Ⅱ 3 | 0.20 |
| Heptanal | 901 | 898 | 97 | Ⅰ | 0.62 |
| Un 1 | 922 | NA 6 | 4.39 | ||
| (R)-α-Pinene | 929 | 928 | 96 | Ⅲ 4 | 2.36 |
| Sabinene | 974 | 972 | 97 | Ⅲ | 13.45 |
| β-Pinene | 975 | 974 | 95 | Ⅲ | 1.15 |
| β-Myrcene | 991 | 989 | 98 | Ⅲ | 7.72 |
| Octanal | 1003 | 1003 | 98 | Ⅰ | 2.00 |
| α-Phellandrene | 1005 | 1003 | 95 | Ⅲ | 1.01 |
| α-Terpinene | 1017 | 1013 | 95 | Ⅲ | 2.19 |
| o-Cymene | 1022 | 1020 | 94 | Ⅲ | 1.90 |
| D-Limonene; β-Phellandrene | 1031 | 1026 | 95 | Ⅲ | 16.93 |
| Un | 1032 | NA | 5.76 | ||
| β-cis-Ocimene | 1038 | 1041 | 94 | Ⅲ | 3.01 |
| γ-Terpinene | 1054 | 1051 | 95 | Ⅲ | 3.58 |
| trans-Sabinene hydrate | 1061 | 1064 | 95 | Ⅲ | 2.27 |
| 1-Octanol | 1071 | 1067 | 97 | Ⅲ | 0.52 |
| Terpinolene | 1083 | 1080 | 97 | Ⅲ | 2.27 |
| Linalool | 1099 | 1101 | 93 | Ⅲ | 6.22 |
| Nonanal | 1104 | 1106 | 98 | Ⅰ | 2.18 |
| L-4-terpineneol | 1182 | 1181 | 95 | Ⅲ | 2.30 |
| Crypton | 1184 | 1185 | 96 | Ⅳ 5 | 1.19 |
| α-Terpineol | 1198 | 1195 | 97 | Ⅲ | 2.06 |
| Decanal | 1206 | 1206 | 99 | Ⅰ | 0.53 |
| Piperitone | 1260 | 1261 | 94 | Ⅲ | 8.36 |
| α-Terpinyl acetate | 1350 | 1351 | 95 | Ⅲ | 1.49 |
| Caryophyllene | 1419 | 1422 | 97 | Ⅲ | 0.68 |
| Xanthoxylin | 1675 | 1678 | 92 | Ⅳ | 2.49 |
| Compound | IC50 | R2 | Toxicity Regression Equations |
|---|---|---|---|
| Sabinene | 172.88 | 0.9916 | y = 0.5611x + 3.7444 |
| Piperitone | 51.99 | 0.9982 | y = 1.0613x + 3.1789 |
| D-Limonene | 448.15 | 0.9992 | y = 0.8291x + 2.8017 |
| Linalool | 18.03 | 0.9935 | y = 1.1721x + 3.5278 |
| β-Myrcene | NA 1 | - | - |
| PSK Addition (%) | Organic Matter (g/kg) | Hydrolyzable Nitrogen (mg/kg) | Available Phosphorus (mg/kg) | Available Potassium (mg/kg) | Total Phosphorus (mg/Kg) | pH |
|---|---|---|---|---|---|---|
| CK | 1.53 ± 0.16 c | 121.47 ± 3.26 a | 52.50 ± 0.49 a | 139.47 ± 2.20 c | 484.53 ± 9.20 b | 6.93 ± 0.03 a |
| 2 | 2.01 ± 0.22 b | 132.57 ± 1.97 b | 50.83 ± 1.25 a | 184.17 ± 6.40 b | 507.70 ± 9.80 ab | 6.74 ± 0.06 a |
| 6 | 2.66 ± 0.29 a | 141.83 ± 2.48 b | 50.97 ± 1.32 a | 245.40 ± 12.00 a | 542.90 ± 7.80 a | 6.35 ± 0.08 b |
| Treatment | Bacterial Community | Fungal Community | ||||||
|---|---|---|---|---|---|---|---|---|
| OTUs | Shannon | Chao1 | ACE | OTUs | Shannon | Chao1 | ACE | |
| CK | 2920 ±234 a | 9.6±0.4 a | 3177±224 a | 3139±204 a | 489±32 a | 5.7±0.2 a | 517±21a | 517±24 a |
| T2 | 2902±309 a | 9.6±0.2 a | 3112±120 a | 3123±127 a | 440±15 ab | 5.4±0.3 a | 467±15 ab | 470±15 ab |
| T6 | 2058±309 b | 8.5±0.3 b | 2243±331 b | 2248±333 b | 384±8 b | 5.3±0.2 a | 420±14 b | 418±13 b |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Liu, M.; Han, X.; Zheng, Y.; Chao, J.; Zhang, C.-S. Prickly Ash Seed Kernel: A New Bio-Fumigation Material Against Tobacco Black Shank. Agronomy 2020, 10, 770. https://doi.org/10.3390/agronomy10060770
Wang Y, Liu M, Han X, Zheng Y, Chao J, Zhang C-S. Prickly Ash Seed Kernel: A New Bio-Fumigation Material Against Tobacco Black Shank. Agronomy. 2020; 10(6):770. https://doi.org/10.3390/agronomy10060770
Chicago/Turabian StyleWang, Yaochen, Minghong Liu, Xiaobin Han, Yanfen Zheng, Jianmin Chao, and Cheng-Sheng Zhang. 2020. "Prickly Ash Seed Kernel: A New Bio-Fumigation Material Against Tobacco Black Shank" Agronomy 10, no. 6: 770. https://doi.org/10.3390/agronomy10060770
APA StyleWang, Y., Liu, M., Han, X., Zheng, Y., Chao, J., & Zhang, C. -S. (2020). Prickly Ash Seed Kernel: A New Bio-Fumigation Material Against Tobacco Black Shank. Agronomy, 10(6), 770. https://doi.org/10.3390/agronomy10060770