Phenology-Adjusted Stress Severity Index to Assess Genotypic Responses to Terminal Drought in Field Grown Potato


Abstract
:1. Introduction
2. Materials and Methods
2.1. Site Description
2.2. Genotypes, Experimental Setup, and Crop Management
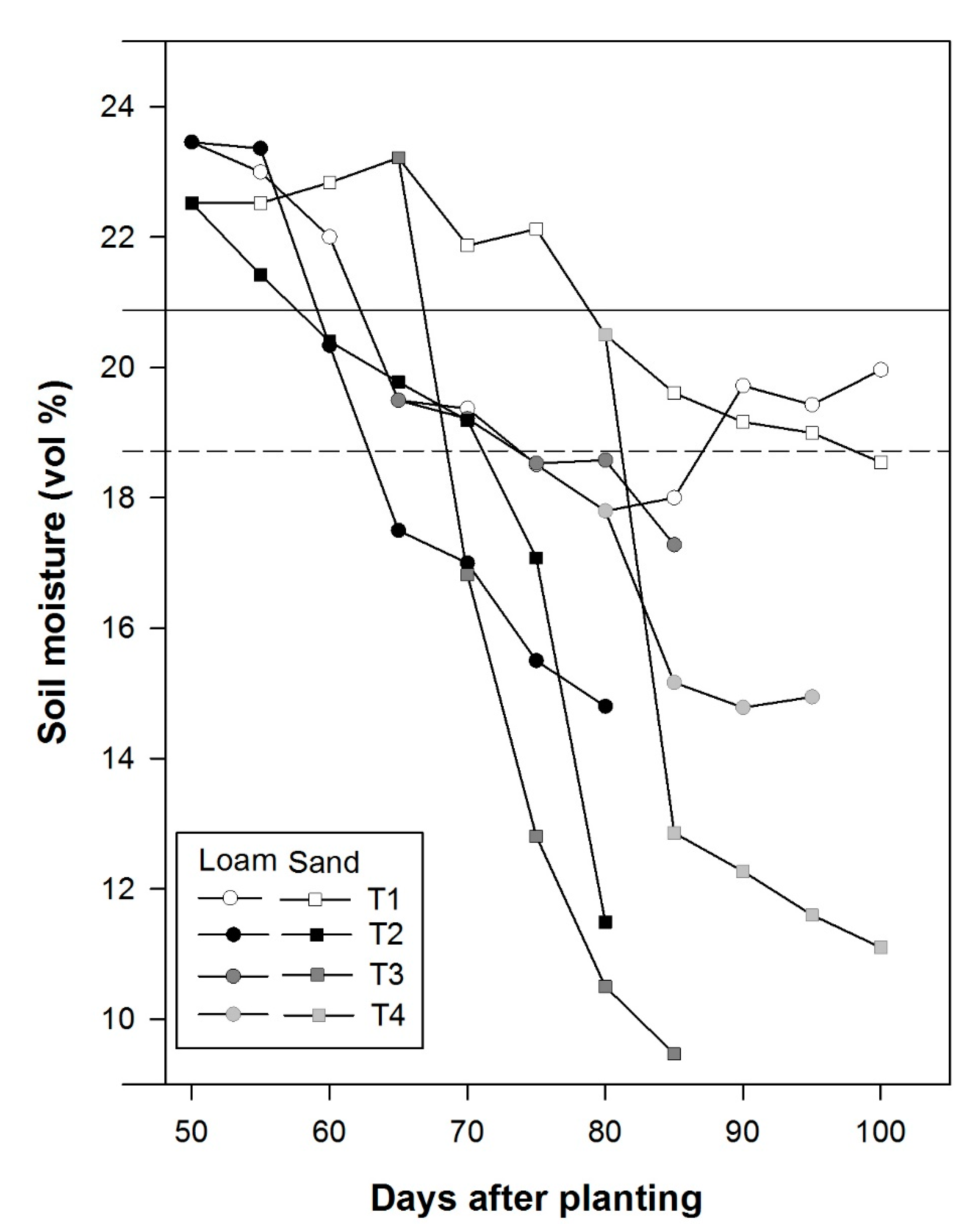
2.3. Irrigation Treatments and Soil Moisture Measurements
2.4. Samplings, Yield, and Yield Components
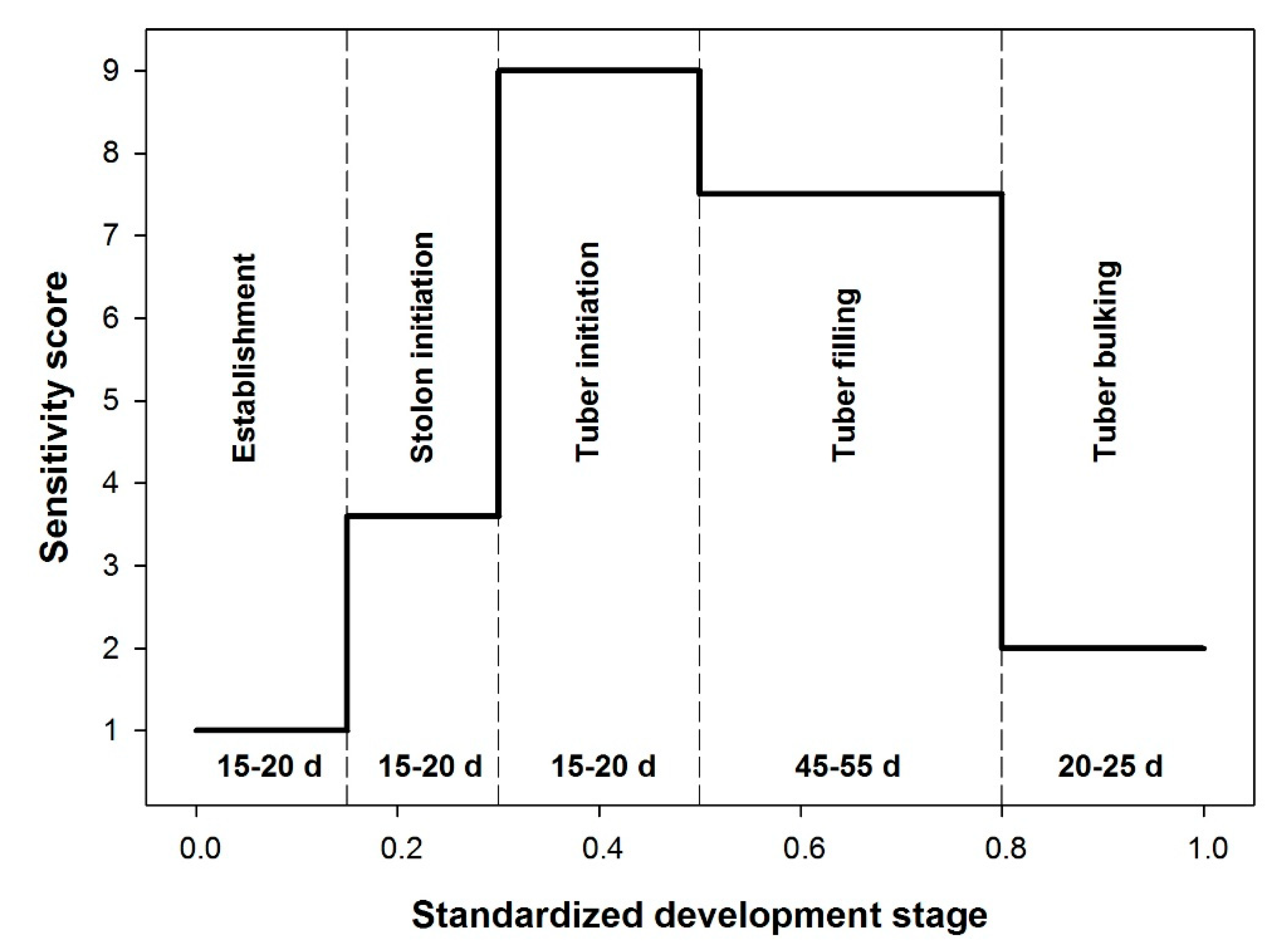
2.5. Phenology and Development Stage Specific Sensitivity to Moisture Deficit Stress
2.6. Soil Water and Stress Severity Index
2.7. Data Analyses
3. Results
3.1. Genotypic Performance under Water Deficit
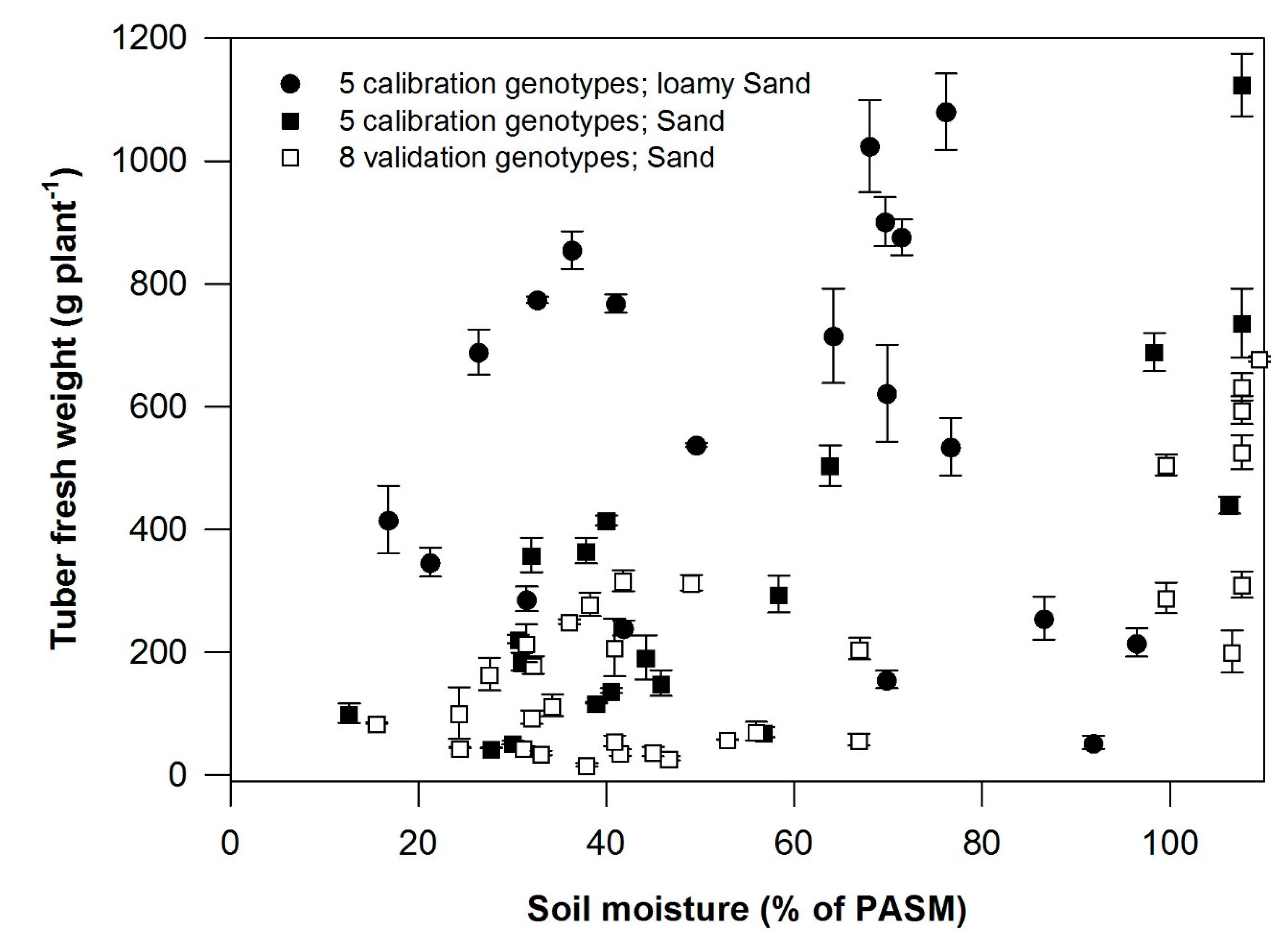
3.2. Plant Available Soil Moisture and Tuber Yield
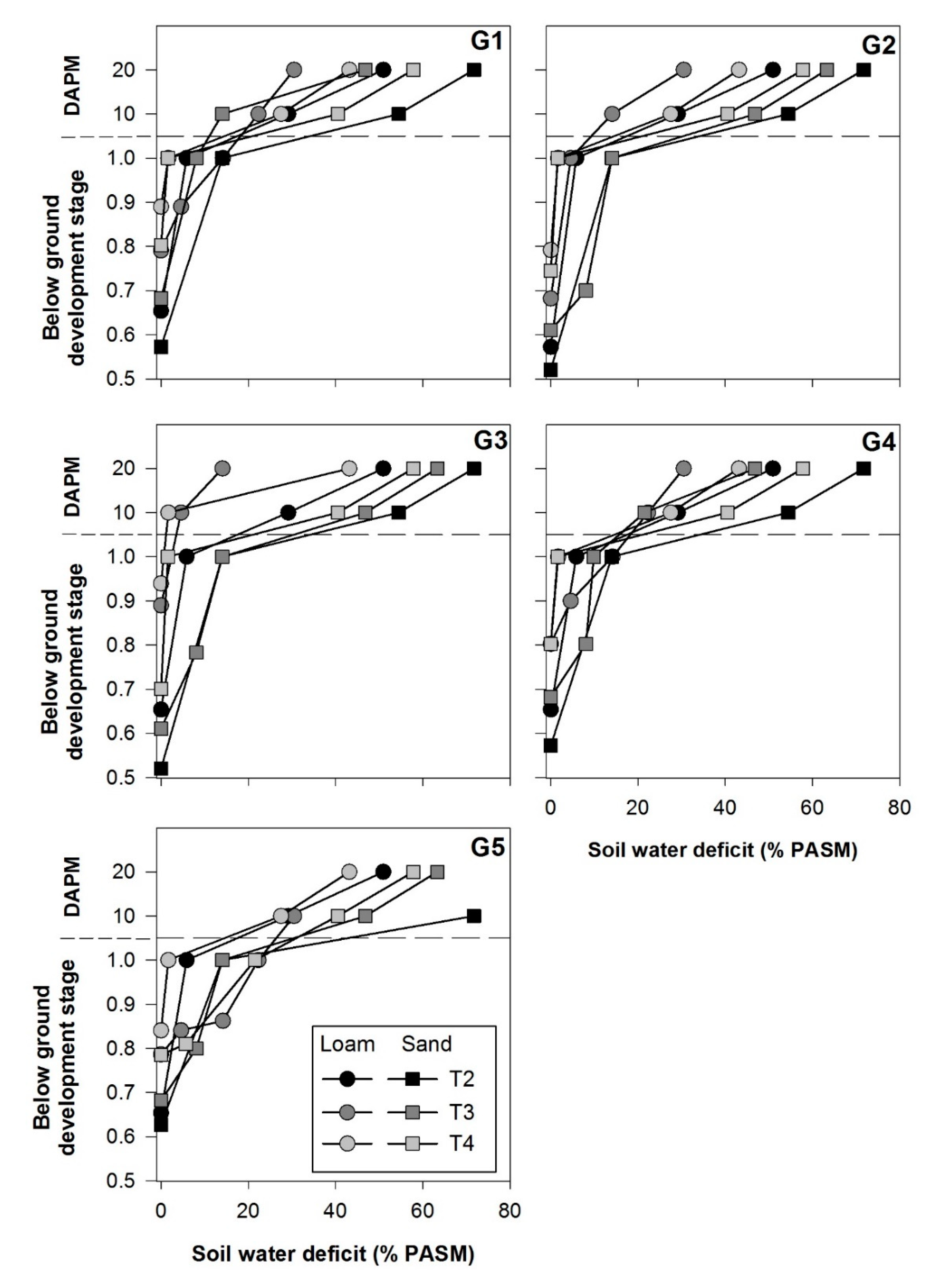
3.3. Soil Moisture Deficit and Below-Ground Development
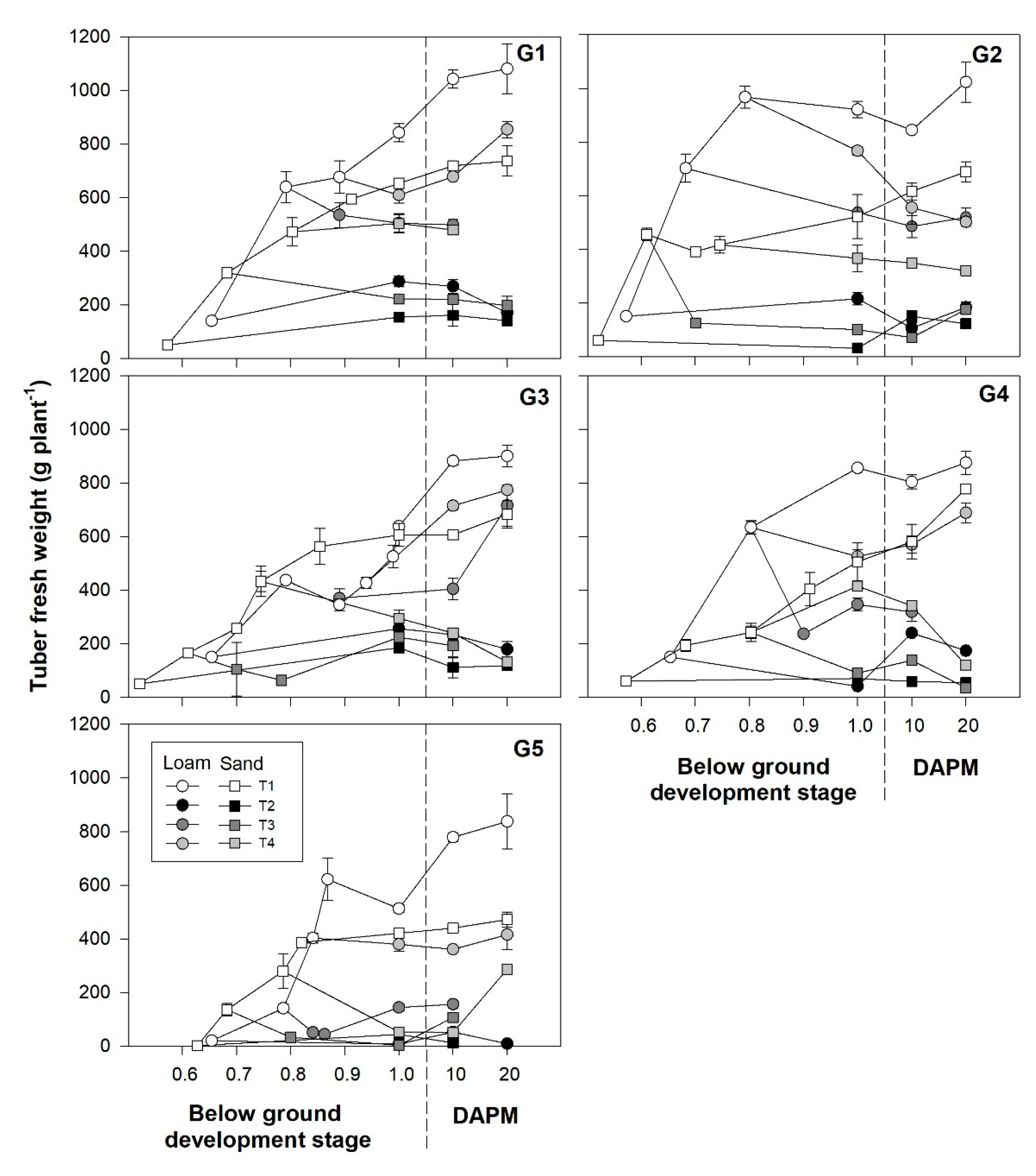
3.4. Below Ground Development and Tuber Fresh Weight
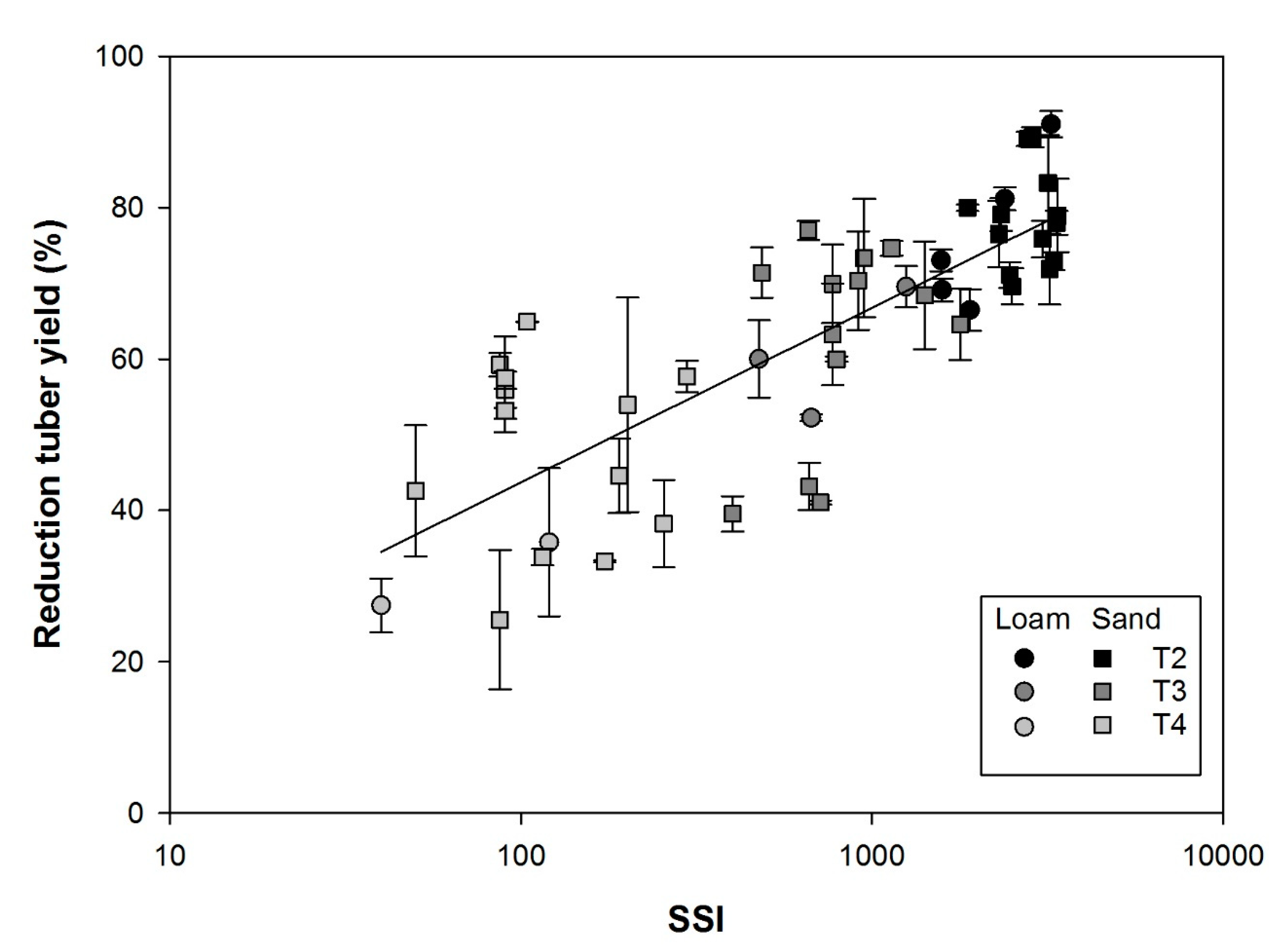
3.5. Stress Severity Index and Tuber Yield Reduction
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Texture | Depth (cm) | E.C. (1:1, dS/m) | CaCO3 (%) | P (ppm) | K (ppm) | |
|---|---|---|---|---|---|---|
| Loam | 0–15 | 0.84 | 0.14 | 16.88 | 499.2 | |
| 15–30 | 0.62 | 0.12 | 3.52 | 321.4 | ||
| Sand | 0–15 | 2.81 | 0 | 19.82 | 447.4 | |
| 15–30 | 1.01 | 0 | 11.66 | 325.8 | ||
| Exchangeable Cations (meg/100 g) | ||||||
| CEC | Ca2+ | Mg2+ | K+ | Na+ | ||
| Loam | 0–15 | 6.79 | 3.36 | 1.67 | 1.31 | 0.44 |
| 15–30 | 6.08 | 3.29 | 1.42 | 0.91 | 0.46 | |
| Sand | 0–15 | 4.86 | 2.58 | 1.33 | 0.72 | 0.23 |
| 15–30 | 3.84 | 1.81 | 1.13 | 0.67 | 0.24 | |
Appendix B
| No. | CIP Number | Group | Duration-Majes | Variety Name |
|---|---|---|---|---|
| G1 | CIP 392797.22 | Lowland tropic virus-resistant | Early | Unica |
| G2 | CIP 397078.12 | Lowland tropic virus-resistant | Intermediate | |
| G3 | CIP 392025.7 | Lowland tropic virus-resistant | Intermediate | |
| G4 | CIP 397073.16 | Early | ||
| G5 | CIP 301040.63 | Lowland tropic virus and late blight resistant | Late | |
| G6 | CIP 720088 | Late | Achirana | |
| G7 | CIP 397077.16 | Lowland tropic virus-resistant | Early | |
| G8 | CIP 394034.7 | Lowland tropic virus-resistant | Early | |
| G9 | CIP 301056.54 | Early | ||
| G10 | CIP 393227.66 | Very early | ||
| G11 | CIP 301024.95 | Lowland tropic virus and late blight resistant | Early | |
| G12 | CIP 397055.2 | Late | ||
| G13 | CIP 397069.5 | Virus resistance | Early |
References
- Levy, D.; Veilleux, R.E. Adaptation of potato to high temperatures and salinity—A review. Am. J. Potato Res. 2007, 84, 487–506. [Google Scholar] [CrossRef]
- Food and Agriculture Organization of the United Nations (FAO). Crop Water Information: Potato. Available online: http://www.fao.org/nr/water/cropinfo_potato.html (accessed on 13 July 2020).
- Stalham, M.A.; Allen, E.J. Effect of variety, irrigation regime and planting date on depth, rate, duration and density of root growth in the potato (Solanum tuberosum) crop. J. Agric. Sci. 2001, 137, 251–270. [Google Scholar] [CrossRef]
- Costa, L.D.; Vedove, G.D.; Gianquinto, G.; Giovanardi, R.; Peressotti, A. Yield, water use efficiency and nitrogen uptake in potato: Influence of drought stress. Potato Res. 1997, 40, 19–34. [Google Scholar] [CrossRef]
- Obidiegwu, J.E.; Bryan, G.J.; Jones, H.G.; Prashar, A. Coping with drought: Stress and adaptive responses in potato and perspectives for improvement. Front. Plant Sci. 2015, 6, 1–23. [Google Scholar] [CrossRef] [Green Version]
- Onder, S.; Caliskan, M.E.; Onder, D.; Caliskan, S.; Çalışkan, M.E. Different irrigation methods and water stress effects on potato yield and yield components. Agric. Water Manag. 2005, 73, 73–86. [Google Scholar] [CrossRef]
- Ramirez, D.A.; Yactayo, W.; Rens, L.R.; Rolando, J.; Palacios, S.; De Mendiburu, F.; Mares, V.; Barreda, C.; Loayza, H.; Monneveux, P.; et al. Defining biological thresholds associated to plant water status for monitoring water restriction effects: Stomatal conductance and photosynthesis recovery as key indicators in potato. Agric. Water Manag. 2016, 177, 369–378. [Google Scholar] [CrossRef]
- Saravia, D.; Farfán-Vignolo, E.R.; Gutierrez, R.; De Mendiburu, F.; Schafleitner, R.; Bonierbale, M.; Khan, M.A. Yield and Physiological Response of Potatoes Indicate Different Strategies to Cope with Drought Stress and Nitrogen Fertilization. Am. J. Potato Res. 2016, 93, 288–295. [Google Scholar] [CrossRef] [Green Version]
- Schafleitner, R.; Gutierrez, R.; Espino, R.; Gaudin, A.; Pérez, J.; Martinez, M.; Dominguez, A.; Tincopa, L.; Alvarado, C.; Numberto, G.; et al. Field Screening for Variation of Drought Tolerance in Solanum tuberosum L. by Agronomical, Physiological and Genetic Analysis. Potato Res. 2007, 50, 71–85. [Google Scholar] [CrossRef]
- Carli, C.; Yuldashev, F.; Khalikov, D.; Condori, B.; Mareš, V.; Monneveux, P. Effect of different irrigation regimes on yield, water use efficiency and quality of potato (Solanum tuberosum L.) in the lowlands of Tashkent, Uzbekistan: A field and modeling perspective. Field Crop. Res. 2014, 163, 90–99. [Google Scholar] [CrossRef]
- Reddy, J.M.; Jumaboev, K.; Bobojonov, I.; Carli, C.; Eshmuratov, D. Yield and water use efficiency of potato varieties under different soil-moisture stress conditions in the Fergana Valley of Central Asia. Agroecol. Sustain. Food Syst. 2016, 40, 407–431. [Google Scholar] [CrossRef]
- Rauf, S.; Al-Khayri, J.M.; Zaharieva, M.; Monneveux, P.; Khalil, F. Breeding Strategies to Enhance Drought Tolerance in Crops. Adv. Plant Breed. Strateg. Agron. Abiotic Biot. Stress Trait. 2016, 2, 397–445. [Google Scholar] [CrossRef]
- Puértolas, J.; Ballester, C.; Elphinstone, E.D.; Dodd, I.C.; Ballester, C. Two potato (Solanum tuberosum) varieties differ in drought tolerance due to differences in root growth at depth. Funct. Plant Biol. 2014, 41, 1107–1118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lahlou, O.; Ledent, J.-F. Root mass and depth, stolons and roots formed on stolons in four cultivars of potato under water stress. Eur. J. Agron. 2005, 22, 159–173. [Google Scholar] [CrossRef]
- MacKerron, D.K.L.; Jefferies, R.A. The influence of early soil moisture stress on tuber numbers in potato. Potato Res. 1986, 29, 299–312. [Google Scholar] [CrossRef]
- Vos, J.; Haverkort, A.J. Water availability and potato crop performance. In Potato Biology and Biotechnology: Advances and Perspectives; Elsevier: Amsterdam, The Netherlands, 2007; pp. 333–351. [Google Scholar]
- Tourneux, C.; Devaux, A.; Camacho, R.M.; Mamani, P.; Ledent, J.-F. Original article Effect of water shortage on six potato genotypes in the highlands of Bolivia (II): Water relations, physiological parameters. Agronomie 2003, 23, 181–190. [Google Scholar] [CrossRef] [Green Version]
- Schittenhelm, S.; Sourell, H.; Löpmeier, F.-J. Drought resistance of potato cultivars with contrasting canopy architecture. Eur. J. Agron. 2006, 24, 193–202. [Google Scholar] [CrossRef]
- Cabello, R.; Monneveux, P.; Mendiburu, F.; Bonierbale, M. Comparison of yield based drought tolerance indices in improved varieties, genetic stocks and landraces of potato (Solanum tuberosum L.). Euphytica 2013, 193, 147–156. [Google Scholar] [CrossRef]
- Wang, X.; Cai, J.; Jiang, N.; Liu, F.; Dai, T.; Cao, W. Pre-anthesis high-temperature acclimation alleviates damage to the flag leaf caused by post-anthesis heat stress in wheat. J. Plant Physiol. 2011, 168, 585–593. [Google Scholar] [CrossRef]
- Rodríguez, P.L.; Sanjuanelo, C.D.; Ñústez, L.C.E.; Moreno-Fonseca, L.P. Growth and phenology of three Andean potato varieties (Solanum tuberosum L.) under water stress. Agron. Colomb. 2016, 34, 141. [Google Scholar] [CrossRef]
- Jensen, C.; Mogensen, V.; Mortensen, G.; Fieldsend, J.; Milford, G.; Andersen, M.N.; Thage, J. Seed glucosinolate, oil and protein contents of field-grown rape (Brassica napus L.) affected by soil drying and evaporative demand. Field Crop. Res. 1996, 47, 93–105. [Google Scholar] [CrossRef]
- Ali, M.; Jensen, C.R.; Mogensen, V.O.; Bahrun, A. Drought adaptation of fleld grown wheat in relation to soil physical conditions. Plant Soil 1999, 208, 149–159. [Google Scholar] [CrossRef]
- Rivera-Hernández, B.; Carrillo-Avila, E.; Obrador-Olán, J.J.; Juárez-López, J.F.; Aceves-Navarro, L.A.; García-López, E. Soil moisture tension and phosphate fertilization on yield components of A-7573 sweet corn (Zea mays L.) hybrid, in Campeche, Mexico. Agric. Water Manag. 2009, 96, 1285–1292. [Google Scholar] [CrossRef]
- Mould, R.D.; Rutherfoord, R.J. The effect of moisture stress during consecutive growth stages on tuber yield and quality of BP1 potatoes (Solanum tuberosum L.). Crop Prod. 1980, 9, 89–92. [Google Scholar] [CrossRef]
- Asch, F.; Bahrun, A.; Jensen, C.R. Root-shoot communication of field-grown maize drought-stressed at different rates as modified by atmospheric conditions. J. Plant Nutr. Soil Sci. 2009, 172, 678–687. [Google Scholar] [CrossRef]
- Stark, J.C.; Love, S.L.; King, B.A.; Marshall, J.M.; Bohl, W.H.; Salaiz, T. Potato Cultivar Response to Seasonal Drought Patterns. Am. J. Potato Res. 2013, 90, 207–216. [Google Scholar] [CrossRef]
- Haverkort, A.J.; Van De Waart, M.; Bodlaender, K.B.A. The effect of early drought stress on numbers of tubers and stolons of potato in controlled and field conditions. Potato Res. 1990, 33, 89–96. [Google Scholar] [CrossRef]
- Doorenbos, J.; Kassam, A.H. Yield response to water. FAO Irrig. Drain. Pap. 1979, 33. [Google Scholar]
- Van Loon, C.D. The effect of water stress on potato growth, development, and yield. Am. J. Potato Res. 1981, 58, 51–69. [Google Scholar] [CrossRef]
- Spitters, C.J.T.; Schapendonk, A.H.C.M. Evaluation of breeding strategies for drought tolerance in potato by means of crop growth simulation. Plant Soil 1990, 123, 193–203. [Google Scholar] [CrossRef]
- Muthoni, J.; Kabira, J. Potato Production under Drought Conditions: Identification of Adaptive Traits. Int. J. Hortic. 2016, 6. [Google Scholar] [CrossRef]
- Boguszewska-Mańkowska, D.; Zarzyńska, K.; Nosalewicz, A. Drought Differentially Affects Root System Size and Architecture of Potato Cultivars with Differing Drought Tolerance. Am. J. Potato Res. 2019, 97, 54–62. [Google Scholar] [CrossRef]
- Simunek, J.; Sejna, M.; van Genuchten, M.T. The Hydrus-1D Software Package for Simulating the Movement of Water, Heat, and Multiple Solutes in Variably Saturated Media, Version 4.17, HYDRUS Software Series 3; Department of Environmental Sciences, University of California Riverside: Riverside, CA, USA, 2018. [Google Scholar]
- Maestrini, B.; Matthijs, B.; Thomas, B.; Lambertus, A.P.L. Can We Use the Relationship Between Within-Field Elevation and NDVI as an Indicator of Drought-Stress? In Environmental Software Systems. Data Science in Action—13th IFIP WG 5.11 International Symposium. In Proceedings of the International Symposium on Environmental Software Systems, Wageningen, The Netherlands, 5–7 February 2020; pp. 122–131, ISBN 9783030398156F. [Google Scholar]
- Li, X.; Ramirez, D.A.; Qin, J.; Dormatey, R.; Bi, Z.; Sun, C.; Wang, H.; Bai, J. Water restriction scenarios and their effects on traits in potato with different degrees of drought tolerance. Sci. Hortic. 2019, 256, 108525. [Google Scholar] [CrossRef]
- Qin, J.; Bian, C.; Liu, J.; Zhang, J.; Jin, L. An efficient greenhouse method to screen potato genotypes for drought tolerance. Sci. Hortic. 2019, 253, 61–69. [Google Scholar] [CrossRef]
- Nouri, A.; Nezami, A.; Kafi, M.; Hassanpanah, D. Growth and yield response of potato genotypes to deficit irrigation. Int. J. Plant Prod. 2016, 10, 139–158. [Google Scholar] [CrossRef]
- Sprenger, H.; Rudack, K.; Schudoma, C.; Neumann, A.; Seddig, S.; Peters, R.; Zuther, E.; Kopka, J.; Hincha, D.K.; Walther, D.; et al. Assessment of drought tolerance and its potential yield penalty in potato. Funct. Plant Biol. 2015, 42, 655–667. [Google Scholar] [CrossRef]
- Jensen, C.R.; Mogensen, V.O.; Poulsen, H.-H.; Henson, I.E.; Aagot, S.; Hansen, E.; Ali, M.; Wollenweber, B. Soil water matric potential rather than water content determines drought responses in field-grown lupin (Lupinus angustifolius). Aust. J. Agric. Res. 1998, 25, 353–363. [Google Scholar] [CrossRef]
- Jensen, C.; Battilani, A.; Plauborg, F.; Psarras, G.; Chartzoulakis, K.; Janowiak, F.; Stikic, R.; Jovanovic, Z.; Li, G.; Qi, X.; et al. Deficit irrigation based on drought tolerance and root signalling in potatoes and tomatoes. Agric. Water Manag. 2010, 98, 403–413. [Google Scholar] [CrossRef]
- Ramirez, D.A.; Yactayo, W.; Gutierrez, R.; Mareš, V.; De Mendiburu, F.; Posadas, A.; Quiroz, R.A. Chlorophyll concentration in leaves is an indicator of potato tuber yield in water-shortage conditions. Sci. Hortic. 2014, 168, 202–209. [Google Scholar] [CrossRef]
- Ray, S.S.; Daş, G.; Singh, J.P.; Panigrahy, S. Evaluation of hyperspectral indices for LAI estimation and discrimination of potato crop under different irrigation treatments. Int. J. Remote Sens. 2006, 27, 5373–5387. [Google Scholar] [CrossRef]






| Month/Year | Mean Air Temperature | Mean Relative Air Humidity | Mean VPD | Mean Daily PPFD | Mean Wind Speed |
|---|---|---|---|---|---|
| (°C) ± SD | (%) ± SD | (kPa) ± SD | (mol m−2 s−1) ± SD | (m s−1) ± SD | |
| Jul ‘13 | 17.0 ± 1.8 | 37.6 ± 9.9 | 1.36 ± 0.33 | 833 ± 109 | 0.88 ± 0.25 |
| Aug ‘13 | 15.7 ± 1.9 | 39.2 ± 14.8 | 1.25 ± 0.37 | 1005 ± 128 | 0.75 ± 0.14 |
| Sep ‘13 | 17.3 ± 1.2 | 45.8 ± 9.4 | 1.24 ± 0.27 | 1189 ± 40 | 0.81 ± 0.21 |
| Oct ‘13 | 17.9 ± 1.3 | 40.2 ± 7.8 | 1.37 ± 0.18 | 1203 ± 115 | 0.74 ± 0.13 |
| Nov ‘13 | 17.4 ± 1.7 | 48.1 ± 14.0 | 1.22 ± 0.37 | 1233 ± 59 | 1.03 ± 0.26 |
| Aug ‘14 | 18.4 ± 2.0 | 31.1 ± 11.3 | 1.63 ± 0.39 | 1058 ± 73 | 1.17 ± 0.24 |
| Sep ‘14 | 18.0 ± 1.9 | 43.3 ± 13.1 | 1.33 ± 0.39 | 1168 ± 83 | 1.23 ± 0.22 |
| Oct ‘14 | 19.0 ± 1.6 | 39.8 ± 11.4 | 1.49 ± 0.36 | 1275 ± 237 | 1.55 ± 0.32 |
| Nov ‘14 | 17.9 ± 2.0 | 54.4 ± 14.3 | 1.11 ± 0.38 | 1309 ± 96 | 1.34 ± 0.34 |
| Dec ‘14 | 18.9 ± 1.0 | 53.3 ± 13.0 | 1.17 ± 0.32 | 1333 ± 60 | 1.23 ± 0.11 |
| Soil Type | Depth (cm) | pH | Soil Texture | BD g cm−3 | Soil Water | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| Sand (%) | Clay (%) | Silt (%) | OM (%) | FC (vol %) | PW (vol %) | PASM (mm) | ||||
| Sand | 0–15 | 7.4 ± 0.24 | 94.0 ± 3.46 | 4.4 ± 0.89 | 0.4 ± 0.89 | 0.9 ± 0.18 | 1.15 | 20.8 | 5.0 | 63.2 |
| 15–30 | 7.5 ± 0.41 | 96.0 ± 1.41 | 4.0 ± 1.41 | 0.0 ± 0.00 | 0.7 ± 0.13 | |||||
| Loam | 0–15 | 8.1 ± 0.18 | 80.8 ± 1.79 | 11.2 ± 1.79 | 8.0 ± 1.41 | 1.5 ± 0.49 | 1.18 | 23.2 | 8.5 | 58.8 |
| 15–30 | 7.9 ± 0.19 | 81.6 ± 1.67 | 10.0 ± 1.41 | 8.4 ± 1.67 | 1.0 ± 0.25 | |||||
| Loamy Sand | Sand | |
|---|---|---|
| (L plant−1) | ||
| T1 | 152 | 83 |
| T2 | 48 | 37 |
| T3 | 71 | 42 |
| T4 | 89 | 47 |
| G | T | Max. Tuber Yield | Tuber Number | Above Ground Biomass | Mean Root Length | Max. Leaf Area Index | |||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Loam | Sand | Loam | Sand | Loam | Sand | Loam | Sand | Loam | Sand | ||||||||||||||||||||
| g pl−1 | SE | g pl−1 | SE | SE | SE | g pl−1 | SE | g pl−1 | SE | cm | SE | cm | SE | m2 m−2 | SE | m2 m−2 | SE | ||||||||||||
| 1 | 1 | 1080 ± 93 | 736 ± 83 | 14 ± 0.5 | 15 ± 1.9 | 35 ± 0.7 | 29 ± 0.8 | 20 ± 2.0 | 13 ± 1.4 | 3.0 ± 0.1 | 2.1 ± 0.2 | ||||||||||||||||||
| 2 | 1 | 1024 ± 113 | 689 ± 55 | 27 ± 0.7 | 22 ± 2.1 | 77 ± 1.0 | 64 ± 2.0 | 19 ± 0.6 | 16 ± 0.2 | 4.0 ± 0.2 | 3.3 ± 0.2 | ||||||||||||||||||
| 3 | 1 | 901 ± 60 | 1282 ± 65 | 13 ± 1.1 | 18 ± 1.5 | 74 ± 5.0 | 25 ± 0.0 | 21 ± 0.2 | 11 ± 0.2 | 3.5 ± 0.1 | 2.2 ± 0.0 | ||||||||||||||||||
| 4 | 1 | 876 ± 43 | 1124 ± 77 | 23 ± 0.5 | 20 ± 0.7 | 47 ± 2.0 | 53 ± 2.0 | 17 ± 0.5 | 14 ± 0.9 | 3.7 ± 0.3 | 2.5 ± 0.2 | ||||||||||||||||||
| 5 | 1 | 838 ± 1029 | 472 ± 42 | 23 ± 0.5 | 22 ± 1.4 | 83 ± 4.0 | 101 ± 4.0 | 21 ± 0.2 | 22 ± 3.0 | 3.5 ± 0.3 | 3.0 ± 0.2 | ||||||||||||||||||
| 1 | 2 | 287 ± 20 | 141 ± 12 | 15 ± 1.4 | 10 ± 2.3 | 10 ± 0.3 | 24 ± 1.8 | 10 ± 0.7 | 6 ± 0.0 | 2.3 ± 0.1 | 2.1 ± 0.1 | ||||||||||||||||||
| 2 | 2 | 216 ± 23 | 150 ± 21 | 8 ± 0.9 | 21 ± 2.6 | 19 ± 3.0 | 19 ± 1.0 | 11 ± 0.7 | 15 ± 0.7 | 2.4 ± 0.3 | 1.5 ± 0.1 | ||||||||||||||||||
| 3 | 2 | 256 ± 35 | 185 ± 14 | 10 ± 0.0 | 12 ± 2.5 | 26 ± 2.0 | 12 ± 0.0 | 14 ± 0.7 | 6 ± 0.2 | 2.2 ± 0.1 | 1.4 ± 0.2 | ||||||||||||||||||
| 4 | 2 | 240 ± 12 | 70 ± 8 | 10 ± 0.9 | 5 ± 1.2 | 13 ± 2.0 | 16 ± 1.0 | 13 ± 0.7 | 9 ± 1.0 | 3.2 ± 0.3 | 2.1 ± 0.1 | ||||||||||||||||||
| 5 | 2 | 53 ± 11 | 44 ± 1 | 7 ± 0.5 | 2 ± 0.5 | 33 ± 1.0 | 19 ± 2.0 | 16 ± 0.8 | 13 ± 1.4 | 2.2 ± 0.2 | 1.7 ± 0.2 | ||||||||||||||||||
| 1 | 3 | 535 ± 47 | 222 ± 7 | 7 ± 1.2 | 4 ± 0.2 | 41 ± 0.2 | 15 ± 1.7 | 13 ± 1.2 | 2 ± 0.2 | 1.3 ± 0.1 | 0.6 ± 0.1 | ||||||||||||||||||
| 2 | 3 | 538 ± 3 | 101 ± 16 | 17 ± 0.7 | 15 ± 1.7 | 24 ± 4.0 | 20 ± 1.0 | 13 ± 0.5 | 8 ± 1.7 | 2.4 ± 0.3 | 1.4 ± 0.3 | ||||||||||||||||||
| 3 | 3 | 716 ± 77 | 192 ± 45 | 12 ± 1.9 | 15 ± 0.7 | 44 ± 2.0 | 18 ± 4.0 | 16 ± 2.2 | 2 ± 0.2 | 2.1 ± 0.1 | 1.6 ± 0.1 | ||||||||||||||||||
| 4 | 3 | 347 ± 24 | 138 ± 4 | 16 ± 1.2 | 7 ± 2.0 | 21 ± 2.0 | 11 ± 2.0 | 17 ± 0.3 | 2 ± 0.7 | 2.1 ± 0.3 | 0.7 ± 0.1 | ||||||||||||||||||
| 5 | 3 | 156 ± 14 | 33 ± 5 | 23 ± 1.6 | 6 ± 0.5 | 12 ± 0.0 | 17 ± 4.0 | 11 ± 1.2 | 11 ± 2.3 | 2.6 ± 0.0 | 1.4 ± 0.2 | ||||||||||||||||||
| 1 | 4 | 855 ± 31 | 504 ± 33 | 5 ± 0.8 | 8 ± 1.4 | 27 ± 1.9 | 18 ± 0.7 | 7 ± 0.5 | 5 ± 1.2 | 1.4 ± 0.1 | 0.8 ± 0.2 | ||||||||||||||||||
| 2 | 4 | 768 ± 15 | 366 ± 21 | 21 ± 0.7 | 24 ± 1.2 | 46 ± 1.0 | 32 ± 7.0 | 16 ± 1.2 | 13 ± 0.7 | 2.4 ± 0.4 | 1.6 ± 0.1 | ||||||||||||||||||
| 3 | 4 | 774 ± 5 | 295 ± 30 | 8 ± 2.1 | 8 ± 1.9 | 24 ± 1.0 | 13 ± 2.0 | 12 ± 0.7 | 4 ± 1.2 | 1.6 ± 0.1 | 0.5 ± 0.2 | ||||||||||||||||||
| 4 | 4 | 689 ± 37 | 415 ± 8 | 13 ± 1.2 | 12 ± 0.5 | 23 ± 2.0 | 14 ± 2.0 | 15 ± 1.4 | 12 ± 1.6 | 1.9 ± 0.1 | 1.2 ± 0.2 | ||||||||||||||||||
| 5 | 4 | 416 ± 55 | 53 ± 3 | 22 ± 1.7 | 4 ± 0.7 | 56 ± 1.0 | 11 ± 2.0 | 13 ± 1.2 | 14 ± 1.2 | 2.6 ± 0.2 | 1.1 ± 0.3 | ||||||||||||||||||
| Means | |||||||||||||||||||||||||||||
| 1 | 901 ± 57 | a | 861 ± 42 | a | 20 ± 0.7 | a | 19 ± 1.5 | a | 63 ± 2.5 | a | 54 ± 1.8 | a | 20 ± 0.7 | a | 15 ± 1.1 | a | 3.5 ± 0.2 | a | 2.6 ± 0.2 | a | |||||||||
| 2 | 210 ± 20 | d | 118 ± 11 | c | 10 ± 0.7 | c | 10 ± 1.8 | b | 20 ± 1.7 | c | 18 ± 1.2 | b | 13 ± 0.7 | bc | 10 ± 0.7 | b | 2.5 ± 0.2 | b | 1.8 ± 0.2 | b | |||||||||
| 3 | 458 ± 33 | c | 137 ± 15 | bc | 15 ± 1.3 | b | 9 ± 1.0 | b | 28 ± 1.6 | b | 16 ± 2.5 | b | 14 ± 1.1 | c | 5 ± 1.0 | c | 2.1 ± 0.2 | b | 1.1 ± 0.2 | c | |||||||||
| 4 | 700 ± 29 | b | 327 ± 19 | b | 14 ± 1.3 | bc | 11 ± 1.1 | b | 35 ± 1.4 | b | 18 ± 2.7 | b | 13 ± 1.0 | b | 10 ± 1.2 | a | 2.0 ± 0.2 | b | 1.0 ± 0.2 | c | |||||||||
| LSD | 176 | 161 | 3.5 | 4.2 | 2.5 | 2.1 | 0.8 | 0.8 | 0.5 | 0.47 | |||||||||||||||||||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hoelle, J.; Asch, F.; Khan, A.; Bonierbale, M. Phenology-Adjusted Stress Severity Index to Assess Genotypic Responses to Terminal Drought in Field Grown Potato. Agronomy 2020, 10, 1298. https://doi.org/10.3390/agronomy10091298
Hoelle J, Asch F, Khan A, Bonierbale M. Phenology-Adjusted Stress Severity Index to Assess Genotypic Responses to Terminal Drought in Field Grown Potato. Agronomy. 2020; 10(9):1298. https://doi.org/10.3390/agronomy10091298
Chicago/Turabian StyleHoelle, Julia, Folkard Asch, Awais Khan, and Merideth Bonierbale. 2020. "Phenology-Adjusted Stress Severity Index to Assess Genotypic Responses to Terminal Drought in Field Grown Potato" Agronomy 10, no. 9: 1298. https://doi.org/10.3390/agronomy10091298
APA StyleHoelle, J., Asch, F., Khan, A., & Bonierbale, M. (2020). Phenology-Adjusted Stress Severity Index to Assess Genotypic Responses to Terminal Drought in Field Grown Potato. Agronomy, 10(9), 1298. https://doi.org/10.3390/agronomy10091298