Altering Tetrapyrrole Biosynthesis by Overexpressing Ferrochelatases (Fc1 and Fc2) Improves Photosynthetic Efficiency in Transgenic Barley

 ,
,  and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Identification of Two Barley FC Genes
2.2. Phylogenetic Analysis
2.3. cDNA Cloning and Binary Plasmid Construction
2.4. Barley Transformation and Analysis of Transgenic Plants
2.5. Protein Isolation and Immunoblot Assay
2.6. Transient Expression of HvFC1-Green Fluorescent Protein (GFP) Fusion
2.7. Plant Material and Growth Conditions
2.8. Photosynthetic Measurements
2.9. Leaf n and Fe analysis
2.10. Chlorophyll Content
2.11. Statistical Analysis
3. Results
3.1. Identification and Sequence Analysis of Two Types of Ferrochelatases in Barley
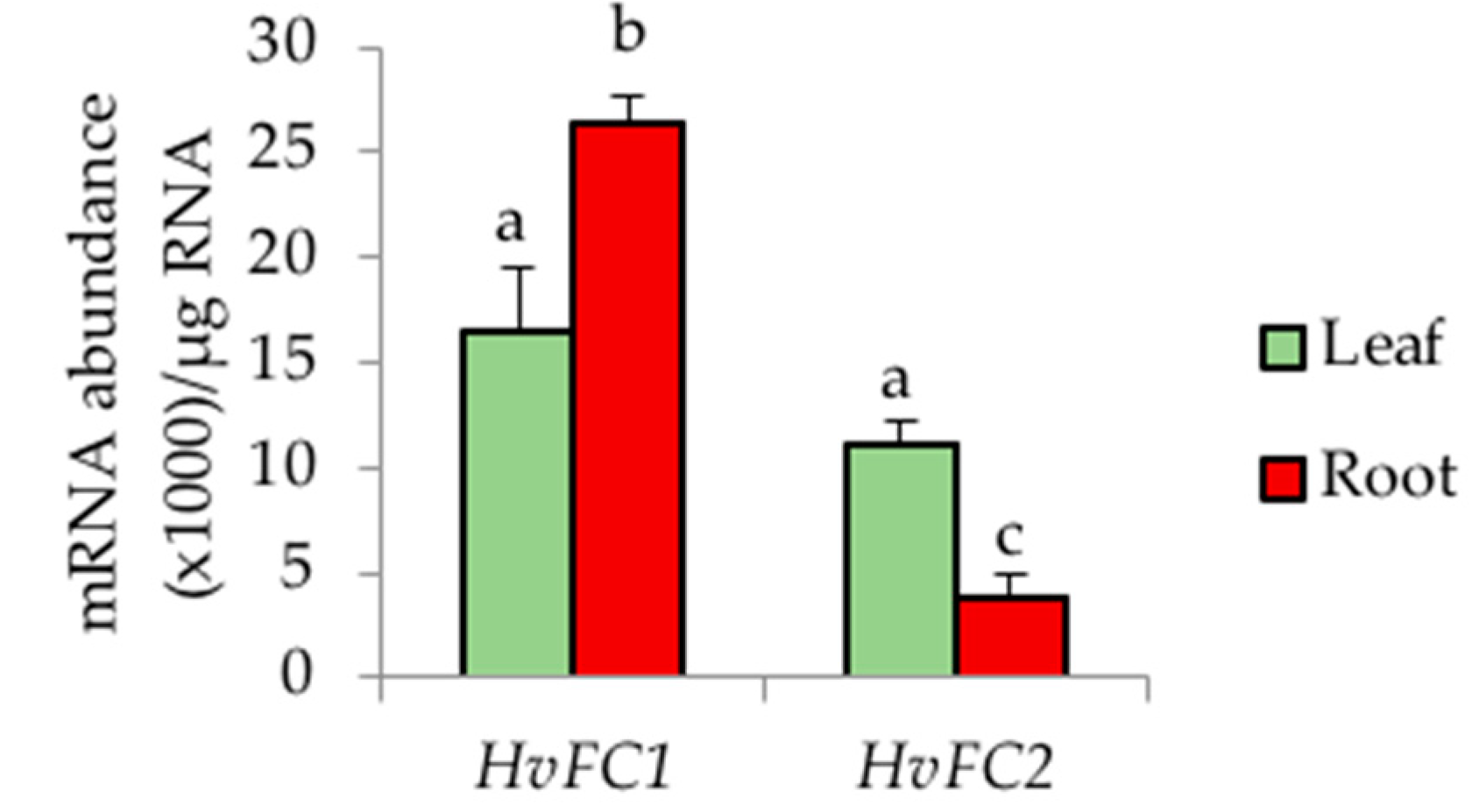
3.2. Two Types of Barley Ferrochelatases Have Differential Tissue-Specific Expression Patterns
3.3. Barley FC1 Is Targeted to Plastids
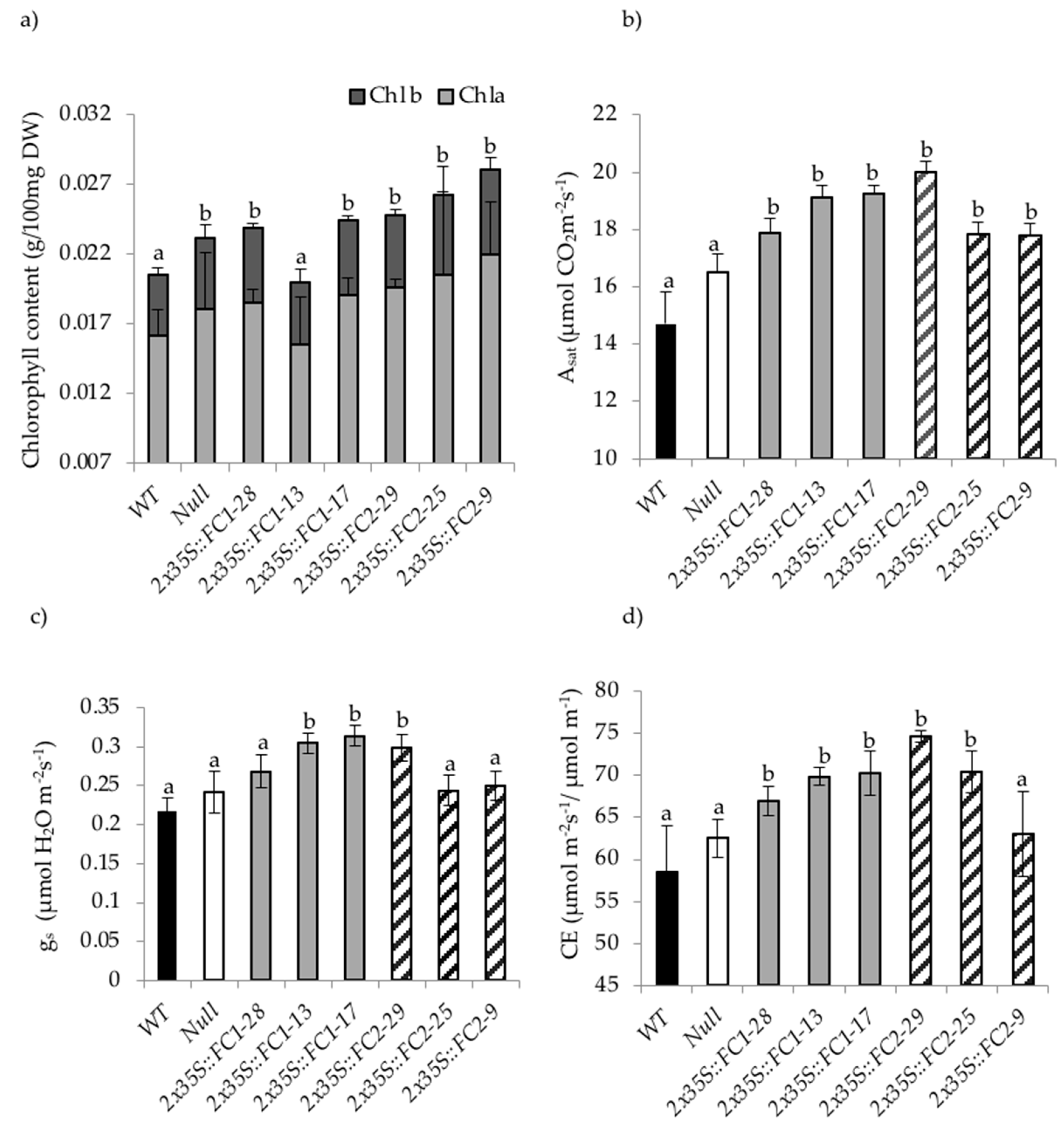
3.4. Increasing HvFC Expression Affects Photosynthetic Performance
4. Discussion
4.1. Two Barley FCs Differ in Structure and Expression
4.2. Both HvFC1 and HvFC2 Are Localised in Chloroplast
4.3. Both Barley FC Isoforms Contribute to Photosynthetic Performance
5. Patent
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Reynolds, M.; Manes, Y.; Izanloo, A.; Langridge, P. Phenotyping approaches for physiological breeding and gene discovery in wheat. Ann. Appl. Biol. 2009, 155, 309–320. [Google Scholar] [CrossRef]
- Reynolds, M.P.; Van-Ginkel, M.; Ribaut, J.M. Avenues for genetic modification of radiation use efficiency in wheat. J. Exp. Bot. 2000, 51, 459–473. [Google Scholar] [CrossRef] [PubMed]
- Sharma-Natu, P.; Ghildiyal, M. Potential targets for improving photosynthesis and crop yield. Curr. Sci. 2005, 88, 1918–1928. [Google Scholar]
- Tanaka, R.; Tanaka, A. Tetrapyrrole biosynthesis in higher plants. Annu. Rev. Plant Biol. 2007, 58, 321–346. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Schliep, M.; Willows, R.D.; Cai, Z.-L.; Neilan, B.A.; Scheer, H. A Red-Shifted Chlorophyll. Science 2010, 329, 1318–1319. [Google Scholar] [CrossRef]
- Cramer, W.; Soriano, G.; Ponomarev, M.; Huang, D.; Zhang, H.; Martinez, S.; Smith, J. Some new structural aspects and old controversies concerning the cytochrome b 6 f complex of oxygenic photosynthesis. Annu. Rev. Plant Biol. 1996, 47, 477–508. [Google Scholar] [CrossRef]
- Kurisu, G.; Zhang, H.; Smith, J.L.; Cramer, W.A. Structure of the cytochrome b6f complex of oxygenic photosynthesis: Tuning the cavity. Science 2003, 302, 1009–1014. [Google Scholar] [CrossRef] [Green Version]
- Smith, A.G.; Cornah, J.E.; Roper, J.M. Compartmentation of tetrapyrrole metabolism in higher plants. In A Plant Carbohydrate Metabolism; Bryant, J., Ed.; BIOS Scientific Publishers: Oxford, UK, 1999; pp. 281–294. [Google Scholar]
- Terry, M.J.; Smith, A.G. A model for tetrapyrrole synthesis as the primary mechanism for plastid-to-nucleus signaling during chloroplast biogenesis. Front. Plant Sci. 2013, 4, 14. [Google Scholar] [CrossRef] [Green Version]
- Woodson, J.D.; Perez-Ruiz, J.M.; Chory, J. Heme synthesis by plastid ferrochelatase i regulates nuclear gene expression in plants. Curr. Biol. 2011, 21, 897–903. [Google Scholar] [CrossRef] [Green Version]
- Woodson, J.D.; Perez-Ruiz, J.M.; Schmitz, R.J.; Ecker, J.R.; Chory, J. Sigma factor-mediated plastid retrograde signals control nuclear gene expression. Plant J. 2013, 73, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Nagahatenna, D.S.K.; Langridge, P.; Whitford, R. Tetrapyrrole-based drought stress signaling. Plant Biotechnol. 2015, 13, 447–459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagahatenna, D.S.K.; Whitford, R. Ferrochelatase compositions and methods to increase agronomic performance of plants. U.S. Patent WO2016054462A1, 7 April 2016. [Google Scholar]
- Al-Karadaghi, S.; Hansson, M.; Nikonov, S.; Jönsson, B.; Hederstedt, L. Crystal structure of ferrochelatase: The terminal enzyme in heme biosynthesis. Structure 1997, 5, 1501–1510. [Google Scholar] [CrossRef]
- Cornah, J.E.; Terry, M.J.; Smith, A.G. Green or red: What stops the traffic in the tetrapyrrole pathway? Trends Plant Sci. 2003, 8, 224–230. [Google Scholar] [CrossRef]
- Mochizuki, N.; Tanaka, R.; Grimm, B.; Masuda, T.; Moulin, M.; Smith, A.G.; Tanaka, A.; Terry, M.J. The cell biology of tetrapyrroles: A life and death struggle. Trends Plant Sci. 2010, 15, 488–498. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, R.; Kobayashi, K.; Masuda, T. Tetrapyrrole metabolism in Arabidopsis thaliana. Arab. Book 2011, 9, e0145. [Google Scholar] [CrossRef] [Green Version]
- Moulin, M.; Smith, A. Regulation of tetrapyrrole biosynthesis in higher plants. Biochem. Soc. Trans. 2005, 33, 737–742. [Google Scholar] [CrossRef]
- Smith, A.G.; Santana, M.A.; Wallace-Cook, A.D.; Roper, J.M.; Labbe-Bois, R. Isolation of a cDNA encoding chloroplast ferrochelatase from Arabidopsis thaliana by functional complementation of a yeast mutant. J. Biol. Chem. 1994, 269, 13405–13413. [Google Scholar]
- Little, H.N.; Jones, O.T. The subcellular loclization and properties of the ferrochelatase of etiolated barley. Biochem. J. 1976, 156, 309–314. [Google Scholar] [CrossRef]
- Chow, K.S.; Singh, D.P.; Walker, A.R.; Smith, A.G. Two different genes encode ferrochelatase in Arabidopsis: Mapping, expression and subcellular targeting of the precursor proteins. Plant J. 1998, 15, 531–541. [Google Scholar] [CrossRef]
- Nagai, S.; Koide, M.; Takahashi, S.; Kikuta, A.; Aono, M.; Sasaki-Sekimoto, Y.; Ohto, H.; Takamiya, K.; Masuda, T. Induction of isoforms of tetrapyrrole biosynthetic enzymes, AtHEMA2 and AtFC1, under stress conditions and their physiological functions in Arabidopsis. Plant Physiol. 2007, 144, 1039–1051. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scharfenberg, M.; Mittermayr, L.; Roepenack-Lahaye, E.V.; Schlicke, H.; Grimm, B.; Leister, D.; Kleine, T. Functional characterization of the two ferrochelatases in Arabidopsis thaliana. Plant Cell Environ. 2014, 38, 280–298. [Google Scholar] [CrossRef]
- Singh, D.; Cornah, J.; Hadingham, S.; Smith, A. Expression analysis of the two ferrochelatase genes in Arabidopsis in different tissues and under stress conditions reveals their different roles in haem biosynthesis. Plant Mol. Biol. 2002, 50, 773–788. [Google Scholar] [CrossRef] [PubMed]
- Papenbrock, J.; Mishra, S.; Mock, H.-P.; Kruse, E.; Schmidt, E.-K.; Petersmann, A.; Braun, H.-P.; Grimm, B. Impaired expression of the plastidic ferrochelatase by antisense RNA synthesis leads to a necrotic phenotype of transformed tobacco plants. Plant J. 2001, 28, 41–50. [Google Scholar] [CrossRef]
- Roper, J.M.; Smith, A.G. Molecular localisation of ferrochelatase in higher plant chloroplasts. Eur. J. Biochem. 1997, 246, 32–37. [Google Scholar] [CrossRef] [PubMed]
- Chow, K.-S.; Singh, D.P.; Roper, J.M.; Smith, A.G. A Single Precursor Protein for Ferrochelatase-I from Arabidopsis Is Imported in Vitro into Both Chloroplasts and Mitochondria. J. Biol. Chem. 1997, 272, 27565–27571. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, T.; Masuda, T.; Singh, D.P.; Tan, F.-C.; Tsuchiya, T.; Shimada, H.; Ohta, H.; Amith, A.G.; Takamiya, K. Two Types of Ferrochelatase in Photosynthetic and Nonphotosynthetic Tissues of Cucumber: Their difference in phylogeny, gene expression, and localization. J. Biol. Chem. 2002, 277, 4731–4737. [Google Scholar] [CrossRef] [Green Version]
- Hay, D.; Ortega-Rodes, P.; Fan, T.; Schnurrer, F.; Brings, L.; Hedtke, B.; Grimm, B. Transgenic Tobacco Lines Expressing Sense or Antisense FERROCHELATASE 1 RNA Show Modified Ferrochelatase Activity in Roots and Provide Experimental Evidence for Dual Localization of Ferrochelatase 1. Plant Cell Physiol. 2016, 57, 2576–2585. [Google Scholar] [CrossRef] [Green Version]
- Lister, R.; Chew, O.; Rudhe, C.; Lee, M.-N.; Whelan, J. Arabidopsis thaliana ferrochelatase-I and-II are not imported into Arabidopsis mitochondria. FEBS Lett. 2001, 506, 291–295. [Google Scholar] [CrossRef] [Green Version]
- Masuda, T.; Suzuki, T.; Shimada, H.; Ohta, H.; Takamiya, K. Subcellular localization of two types of ferrochelatase in cucumber. Planta 2003, 217, 602–609. [Google Scholar] [CrossRef]
- Espinas, N.A.; Kobayashi, K.; Sato, Y.; Mochizuki, N.; Takahashi, K.; Tanaka, R.; Masuda, T. Allocation of Heme Is Differentially Regulated by Ferrochelatase Isoforms in Arabidopsis Cells. Front. Plant Sci. 2016, 7, 1326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, T.; Roling, L.; Meiers, A.; Brings, L.; Ortega-Rodes, P.; Hedtke, B.; Grimm, B. Complementation studies of the Arabidopsis fc1 mutant substantiate essential functions of ferrochelatase 1 during embryogenesis and salt stress. Plant Cell Environ. 2019, 42, 618–632. [Google Scholar] [CrossRef]
- Miyamoto, K.; Tanaka, R.; Teramoto, H.; Masuda, T.; Tsuji, H.; Inokuchi, H. Nucleotide Sequences of cDNA Clones Encoding Ferrochelatase from Barley and Cucumber. Plant Physiol 1994, 105, 769–770. [Google Scholar] [CrossRef] [PubMed]
- Hall, B.G. Building phylogenetic trees from molecular data with MEGA. Mol. Biol. Evol. 2013, 30, 1229–1235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Curtis, M.; Grossniklaus, U. A Gateway Cloning Vector Set for High-Throughput Functional Analysis of Genes in Planta. Plant Physiol. 2003, 133, 462–469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tingay, S.; McElroy, D.; Kalla, R.; Fieg, S.; Wang, M.; Thornton, S.; Brettell, R. Agrobacterium tumefaciens-mediated barley transformation. Plant J. 1997, 11, 1369–1376. [Google Scholar] [CrossRef]
- Matthews, P.R.; Wang, M.-B.; Waterhouse, P.M.; Thornton, S.; Fieg, S.J.; Gubler, F.; Jacobsen, J.V. Marker gene elimination from transgenic barley, using co-transformation with adjacenttwin T-DNAs’ on a standard Agrobacterium transformation vector. Mol. Breed. 2001, 7, 195–202. [Google Scholar] [CrossRef]
- Sambrook, J.; Russell, D.W. Molecular Cloning: A Laboratory Manual, 3rd ed.; Cold Spring Harbor: New York, NY, USA, 2001; Volume 5, pp. 35–39. [Google Scholar]
- Burton, R.A.; Shirley, N.J.; King, B.J.; Harvey, A.J.; Fincher, G.B. The CesA Gene Family of Barley. Quantitative Analysis of Transcripts Reveals Two Groups of Co-Expressed Genes. Plant Physiol. 2004, 134, 224–236. [Google Scholar] [CrossRef] [Green Version]
- Garnett, T.; Conn, V.; Plett, D.; Conn, S.; Zanghellini, J.; Mackenzie, N.; Enju, A.; Francis, K.; Holtham, L.; Roessner, U.; et al. The response of the maize nitrate transport system to nitrogen demand and supply across the lifecycle. New Phytol. 2013, 198, 82–94. [Google Scholar] [CrossRef] [Green Version]
- Wheal, M.S.; Fowles, T.O.; Palmer, L.T. A cost-effective acid digestion method using closed polypropylene tubes for inductively coupled plasma optical emission spectrometry (ICP-OES) analysis of plant essential elements. Anal. Methods 2011, 3, 2854–2863. [Google Scholar] [CrossRef]
- Hiscox, J.D.; Israelstam, G.F. A method for the extraction of chlorophyll from leaf tissue without maceration. Can. J. Bot. 1979, 57, 1332–1334. [Google Scholar] [CrossRef]
- Dittami, S.M.; Michel, G.; Collen, J.; Boyen, C.; Tonon, T. Chlorophyll-binding proteins revisited—A multigenic family of light-harvesting and stress proteins from a brown algal perspective. BMC Evol. Biol. 2010, 10, 365–379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Natesan, S.K.A.; Sullivan, J.A.; Gray, J.C. Stromules: A characteristic cell-specific feature of plastid morphology. J. Exp. Bot. 2005, 56, 787–797. [Google Scholar] [CrossRef] [PubMed]
- Arimura, S.; Tsutsumi, N. A dynamin-like protein (ADL2b), rather than FtsZ, is involved in Arabidopsis mitochondrial division. Proc. Natl. Acad. Sci. USA 2002, 99, 5727–5731. [Google Scholar] [CrossRef] [Green Version]
- Arimura, S.; Yamamoto, J.; Aida, G.P.; Nakazono, M.; Tsutsumi, N. Frequent fusion and fission of plant mitochondria with unequal nucleoid distribution. Proc. Natl. Acad. Sci. USA 2004, 101, 7805–7808. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, T.; Masuda, T.; Inokuchi, H.; Shimada, H.; Ohta, H.; Takamiya, K. Overexpression, enzymatic properties and tissue localization of a Ferrochelatase of cucumber. Plant Cell Physiol. 2000, 41, 192–199. [Google Scholar] [CrossRef] [Green Version]
- Nijs, I.; Behaeghe, T.; Impens, I. Leaf Nitrogen Content as a Predictor of Photosynthetic Capacity in Ambient and Global Change Conditions. J. Biogeogr. 1995, 22, 177–183. [Google Scholar] [CrossRef]
- Sobotka, R.; Tichy, M.; Wilde, A.; Hunter, C.N. Functional assignments for the carboxyl-terminal domains of the ferrochelatase from Synechocystis PCC 6803: The CAB domain plays a regulatory role, and region II is essential for catalysis. Plant Physiol. 2011, 155, 1735–1747. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, K.; Takabayashi, A.; Tanaka, A.; Tanaka, R. Functional analysis of light-harvesting-like protein 3 (LIL3) and its light-harvesting chlorophyll-binding motif in Arabidopsis. J. Biol. Chem. 2014, 289, 987–999. [Google Scholar] [CrossRef] [Green Version]
- Storm, P.; Tibiletti, T.; Hall, M.; Funk, C. Refolding and enzyme kinetic studies on the ferrochelatase of the cyanobacterium Synechocystis sp. PCC 6803. PLoS ONE 2013, 8, e55569. [Google Scholar] [CrossRef] [Green Version]
- Kang, K.; Lee, K.; Park, S.; Lee, S.; Kim, Y.; Back, K. Overexpression of Rice Ferrochelatase I and II Leads to Increased Susceptibility to Oxyfluorfen Herbicide in Transgenic Rice. J. Plant Biol. 2010, 53, 291–296. [Google Scholar] [CrossRef]
- Vothknecht, U.C.; Kannangara, C.G.; von Wettstein, D. Barley glutamyl tRNA (Glu) reductase: Mutations affecting haem inhibition and enzyme activity. Phytochemistry 1998, 47, 513–519. [Google Scholar] [CrossRef]







© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nagahatenna, D.S.K.; Tiong, J.; Edwards, E.J.; Langridge, P.; Whitford, R. Altering Tetrapyrrole Biosynthesis by Overexpressing Ferrochelatases (Fc1 and Fc2) Improves Photosynthetic Efficiency in Transgenic Barley. Agronomy 2020, 10, 1370. https://doi.org/10.3390/agronomy10091370
Nagahatenna DSK, Tiong J, Edwards EJ, Langridge P, Whitford R. Altering Tetrapyrrole Biosynthesis by Overexpressing Ferrochelatases (Fc1 and Fc2) Improves Photosynthetic Efficiency in Transgenic Barley. Agronomy. 2020; 10(9):1370. https://doi.org/10.3390/agronomy10091370
Chicago/Turabian StyleNagahatenna, Dilrukshi S. K., Jingwen Tiong, Everard J. Edwards, Peter Langridge, and Ryan Whitford. 2020. "Altering Tetrapyrrole Biosynthesis by Overexpressing Ferrochelatases (Fc1 and Fc2) Improves Photosynthetic Efficiency in Transgenic Barley" Agronomy 10, no. 9: 1370. https://doi.org/10.3390/agronomy10091370
APA StyleNagahatenna, D. S. K., Tiong, J., Edwards, E. J., Langridge, P., & Whitford, R. (2020). Altering Tetrapyrrole Biosynthesis by Overexpressing Ferrochelatases (Fc1 and Fc2) Improves Photosynthetic Efficiency in Transgenic Barley. Agronomy, 10(9), 1370. https://doi.org/10.3390/agronomy10091370