Biostimulant-Treated Seedlings under Sustainable Agriculture: A Global Perspective Facing Climate Change

 ,
, 
 ,
,  ,
,  ,
,  ,
,  ,
, 
 and
and
Abstract
:1. Introduction
2. Source of Biostimulants
3. Biostimulant Applications for Crop Agronomy
4. Implications of Biostimulant for Abiotic Stress Tolerance
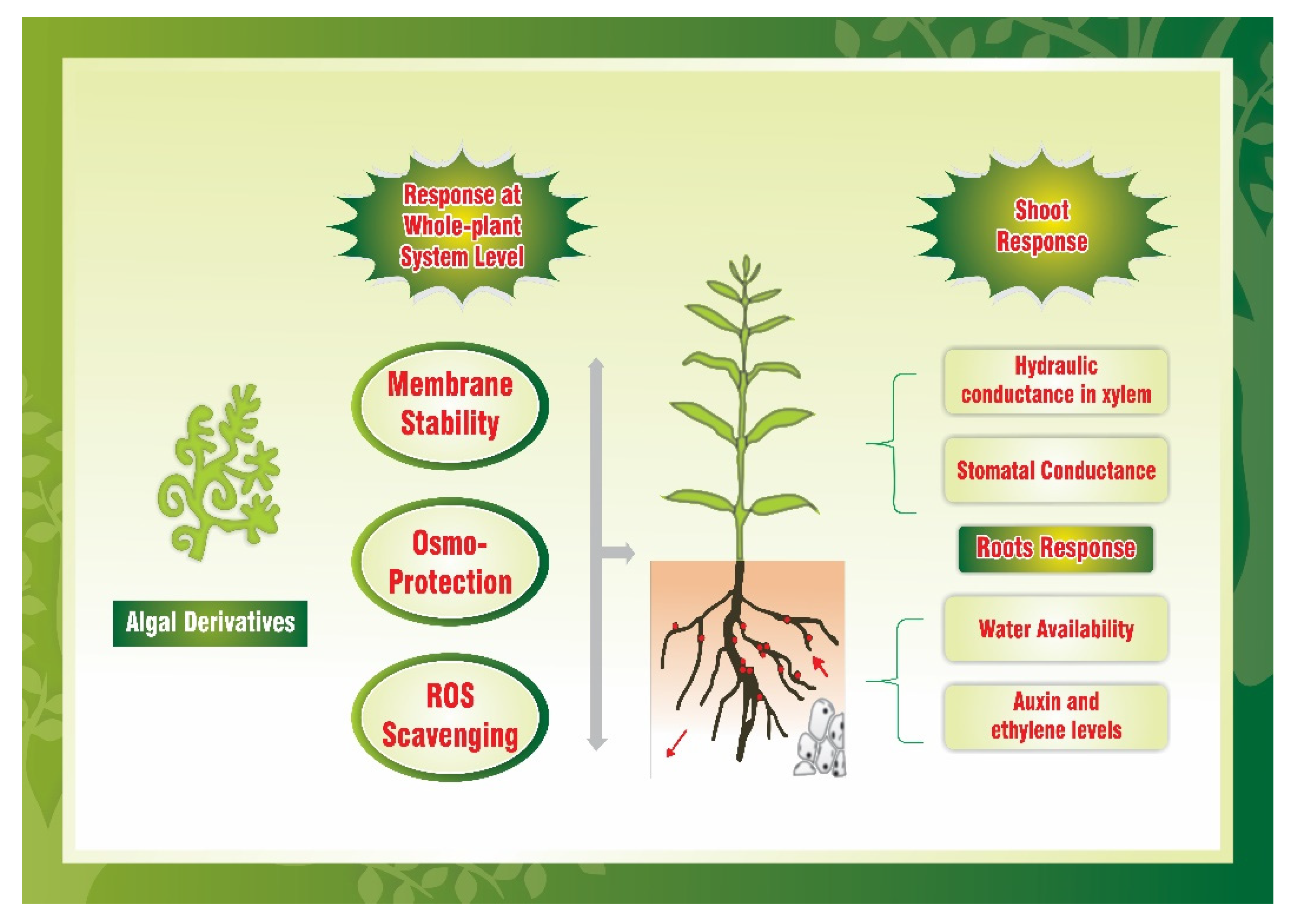
4.1. Drought
4.2. Salinity
4.3. Thermal Stress
5. Implication of Biostimulants on Antioxidant Potential
6. Role of Biostimulants on Nutrient Use Efficiency of Plants
7. Quality Commodity Ramification of Biostimulants for Agri-Production
8. Legal Framework and Limitation of using Biostimulants
9. Future Insights and Challenges
10. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Rouphael, Y.; Colla, G. Editorial: Biostimulants in agriculture. Front. Plant Sci. 2020, 11, 40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Punia, H.; Tokas, J.; Malik, A.; Satpal; Rani, A.; Gupta, P.; Kumari, A.; Mor, V.S.; Bhuker, A.; Kumar, S. Solar radiation and nitrogen use efficiency for sustainable agriculture. In Resources Use Efficiency in Agriculture; Kumar, S., Meena, R.S., Jhariya, M.K., Eds.; Springer Nature Singapore Pte Ltd.: Singapore, 2020; pp. 177–212. [Google Scholar]
- Ortiz-Ospina, E.; Roser, M. World Population Growth. OurWorldInData. 2016. Available online: https://ourworldindata.org/world-population-growth (accessed on 26 October 2020).
- Mu, S.; Zhou, S.; Chen, Y.; Li, J.; Ju, W.; Odeh, I.O.A. Assessing the impact of restoration-induced land conversion and management alternatives on net primary productivity in Inner Mongolian grassland, China. Glob. Planet. Chang. 2013, 108, 29–41. [Google Scholar] [CrossRef]
- Punia, H.; Madan, S.; Malik, A.; Sethi, S. Stability analysis for quality attributes in durum wheat (Triticum durum L.) genotypes. Bangladesh J. Bot. 2019, 48, 967–972. [Google Scholar] [CrossRef]
- Punia, H.; Tokas, J.; Malik, A.; Singh, S.; Phogat, D.; Bhuker, A.; Mor, V.S.; Rani, A.; Sheokand, R. Discerning morpho-physiological and quality traits contributing to salinity tolerance acquisition in sorghum [Sorghum bicolor (L.) Moench]. S. Afr. J. Bot. 2020. [Google Scholar] [CrossRef]
- Punia, H.; Tokas, J.; Malik, A.; Sangwan, S.; Baloda, S.; Singh, N.; Singh, S.; Bhuker, A.; Singh, P.; Yashveer, S.; et al. Identification and detection of bioactive peptides in milk and dairy products: Remarks about agro-foods. Molecules 2020, 25, 3328. [Google Scholar] [CrossRef]
- Calvo, P.; Nelson, L.; Kloepper, J.W. Agricultural uses of plant biostimulants. Plant Soil 2014, 383, 3–41. [Google Scholar] [CrossRef] [Green Version]
- Nardi, S.; Pizzeghello, D.; Schiavon, M.; Ertani, A. Plant biostimulants: Physiological responses induced by protein hydrolyzed-based products and humic substances in plant metabolism. Sci. Agricola 2016, 73, 18–23. [Google Scholar] [CrossRef] [Green Version]
- De Pascale, S.; Rouphael, Y.; Colla, G. Plant biostimulants: Innovative tool for enhancing plant nutrition in organic farming. Eur. J. Hortic. Sci. 2018, 82, 277–285. [Google Scholar] [CrossRef]
- Rouphael, Y.; De Micco, V.; Arena, C.; Raimondi, G.; Colla, G.; De Pascale, S. Effect of Ecklonia maxima seaweed extract on yield, mineral composition, gas exchange, and leaf anatomy of zucchini squash grown under saline conditions. J. Appl. Phycol. 2017, 29, 459–470. [Google Scholar] [CrossRef]
- Rouphael, Y.; Colla, G.; Graziani, G.; Ritieni, A.; Cardarelli, M.; De Pascale, S. Phenolic composition, antioxidant activity and mineral profile in two seed-propagated artichoke cultivars as affected by microbial inoculants and planting time. Food Chem. 2017, 234, 10–19. [Google Scholar] [CrossRef]
- Yakhin, O.I.; Lubyanov, A.A.; Yakhin, I.A.; Brown, P.H. Biostimulants in plant science: A global perspective. Front. Plant Sci. 2017, 7, 2049. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rouphael, Y.; Kyriacou, M.C.; Petropoulos, S.A.; De Pascale, S.; Colla, G. Improving vegetable quality in controlled environments. Sci. Hortic. 2018, 234, 275–289. [Google Scholar] [CrossRef]
- Zhang, X.; Schmidt, R.E. The impact of growth regulators on the α-tocopherol status in water-stressed Poa pratensis. Int. Turfgrass Soc. Res. J. 1997, 8, 1364–2137. [Google Scholar]
- Du Jardin, P. The Science of Plants Biostimulants: A Bibliographic Analysis; EU: Brisel, Belgium, 2012. [Google Scholar]
- Du Jardin, P. Plant biostimulants: Definition, concept, main categories and regulation. Sci. Hortic. 2015, 196, 3–14. [Google Scholar] [CrossRef] [Green Version]
- Regulation, E.U. 1009 of the European Parliament and of the Council of 5 June 2019 Laying Down Rules on the Making Available on the Market of EU Fertilising Products and Amending Regulations (EC) No 1069/2009 and (EC) No 1107/2009 and Repealing Regulation (EC) No 2003/200. Available online: https://eur-lex.europa.eu/legal-content/EN/ALL/?uri=CELEX%3A32019R1009 (accessed on 14 October 2020).
- Taylor, A.G.; Allen, P.S.; Bennett, M.A.; Bradford, K.J.; Burris, J.S.; Misra, M.K. Seed enhancements. Seed Sci. Res. 1998, 8, 245–256. [Google Scholar] [CrossRef]
- Snapp, S.; Price, R.; Morton, M. Seed priming of winter annual cover crops improves germination and emergence. Agron. J. 2008, 100, 1506–1510. [Google Scholar] [CrossRef]
- Harris, D.; Raghuwanshi, B.S.; Gangwar, J.S.; Singh, S.C.; Joshi, K.D.; Rashid, A.; Hollington, P.A. Participatory evaluation by farmers of on-farm seed priming in wheat in india, nepal and pakistan. Exp. Agric. 2001, 37, 403–415. [Google Scholar] [CrossRef]
- Vartha, E.W.; Clifford, P.T.P. Effects of seed coating on establishment and survival of grasses, surface-sown on tussock grasslands. N. Z. J. Exp. Agric. 1973, 1, 39–43. [Google Scholar] [CrossRef]
- Falloon, R.; Fletcher, R. Increased herbage production from perennial ryegrass following fungicide seed treatment. N. Z. J. Agric. Res. 1983, 26, 1–5. [Google Scholar] [CrossRef]
- Amirkhani, M.; Netravali, A.N.; Huang, W.; Taylor, A.G. Investigation of soy protein–based biostimulant seed coating for broccoli seedling and plant growth enhancement. HortScience 2016, 51, 1121–1126. [Google Scholar] [CrossRef] [Green Version]
- Parađiković, N.; Zeljković, S.; Tkalec, M.; Vinković, T.; Maksimović, I.; Haramija, J. Influence of biostimulant application on growth, nutrient status and proline concentration of begonia transplants. Biol. Agric. Hortic. 2017, 33, 89–96. [Google Scholar] [CrossRef]
- Fernández, J.A.; Ceglie, F.; Tuzel, Y.; Koller, M.; Koren, A.; Hitchings, R.; Tittarelli, F. Organic substrate for transplant production in organic nurseries. A review. Agron. Sustain. Dev. 2018, 38, 35. [Google Scholar]
- Traon, D.; Amat, L.; Zotz, F.; du Jardin, P. A Legal Framework for Plant Biostimulants and Agronomic Fertiliser Additives in the EU-Report to the European Commission; DG Enterprise & Industry: Copenhagen, Denmark, 2014. [Google Scholar]
- Rouphael, Y.; Spíchal, L.; Panzarová, K.; Casa, R.; Colla, G. High-throughput plant phenotyping for developing novel biostimulants: From lab to field or from field to lab? Front. Plant Sci. 2018, 9, 1197. [Google Scholar] [CrossRef] [PubMed]
- Kunicki, E.; Grabowska, A.; Sękara, A.; Wojciechowska, R. The effect of cultivar type, time of cultivation, and biostimulant treatment on the yield of spinach (Spinacia oleracea L.). Folia Hortic. 2010, 22, 9–13. [Google Scholar] [CrossRef] [Green Version]
- Canellas, L.P.; Olivares, F.L.; Aguiar, N.O.; Jones, D.L.; Nebbioso, A.; Mazzei, P.; Piccolo, A. Humic and fulvic acids as biostimulants in horticulture. Sci. Hortic. 2015, 196, 15–27. [Google Scholar] [CrossRef]
- Shah, Z.H.; Rehman, H.M.; Akhtar, T.; Alsamadany, H.; Hamooh, B.T.; Mujtaba, T.; Daur, I.; Al Zahrani, Y.; Alzahrani, H.A.S.; Ali, S.; et al. Humic substances: Determining potential molecular regulatory processes in plants. Front. Plant Sci. 2018, 9, 263. [Google Scholar] [CrossRef] [Green Version]
- Rang-Jin, X.; Kyriacou, M.; Colla, G. Vegetable grafting: A toolbox for securing yield stability under multiple stress conditions. Front. Plant Sci. 2018, 8, 2255. [Google Scholar]
- Assainar, S.K.; Abbott, L.K.; Mickan, B.S.; Whiteley, A.S.; Siddique, K.H.M.; Solaiman, Z.M. Response of wheat to a multiple species microbial inoculant compared to fertilizer application. Front. Plant Sci. 2018, 9, 1601. [Google Scholar] [CrossRef] [Green Version]
- Khan, W.; Rayirath, U.P.; Subramanian, S.; Jithesh, M.N.; Rayorath, P.; Hodges, D.M.; Critchley, A.T.; Craigie, J.S.; Norrie, J.; Prithiviraj, B. Seaweed extracts as biostimulants of plant growth and development. J. Plant Growth Regul. 2009, 28, 386–399. [Google Scholar] [CrossRef]
- Craigie, J.S. Seaweed extract stimuli in plant science and agriculture. J. Appl. Phycol. 2011, 23, 371–393. [Google Scholar] [CrossRef]
- Sharma, H.S.S.; Fleming, C.C.; Selby, C.; Rao, J.R.; Martin, T. Plant biostimulants: A review on the processing of macroalgae and use of extracts for crop management to reduce abiotic and biotic stresses. J. Appl. Phycol. 2013, 26, 465–490. [Google Scholar] [CrossRef]
- Battacharyya, D.; Babgohari, M.Z.; Rathor, P.; Prithiviraj, B. Seaweed extracts as biostimulants in horticulture. Sci. Hortic. 2015, 196, 39–48. [Google Scholar] [CrossRef]
- Nabti, E.; Jha, B.; Hartmann, A. Impact of seaweeds on agricultural crop production as biofertilizer. Int. J. Environ. Sci. Technol. 2016, 14, 1119–1134. [Google Scholar] [CrossRef]
- Carvalho, M.E.A.; Castro, P.R.C.; Novembre, A.D.C.; Chamma, H. Seaweed extract improves the vigor and provides the rapid emergence of dry bean seeds. Am. J. Agric. Environ. Sci. 2013, 13, 1104–1107. [Google Scholar]
- Carvalho, M.E.A.; Castro, P.R.; Gaziola, S.A.; Azevedo, R.A. Is seaweed extract an elicitor compound? Changing proline content in drought-stressed bean plants. Comun. Sci. 2018, 9, 292–297. [Google Scholar] [CrossRef]
- Colla, G.; Nardi, S.; Cardarelli, M.; Ertani, A.; Lucini, L.; Canaguier, R.; Rouphael, Y. Protein hydrolysates as biostimulants in horticulture. Sci. Hortic. 2015, 196, 28–38. [Google Scholar] [CrossRef]
- Parađiković, N.; Tkalec, M.; Zeljković, S.; Vinković, T. Biostimulant application in transplants production of Allium sativum L. and Rosa canina L. In Proceedings of the Fifth International Scientific Agricultural Symposium “Agrosym 2014”, Jahorina, Bosnia and Herzegovina, 23–26 October 2014; p. 694. [Google Scholar]
- Colla, G.; Hoagland, L.; Ruzzi, M.; Cardarelli, M.; Bonini, P.; Canaguier, R.; Rouphael, Y. Biostimulant action of protein hydrolysates: Unraveling their effects on plant physiology and microbiome. Front. Plant Sci. 2017, 8, 2202. [Google Scholar] [CrossRef] [Green Version]
- Rouphael, Y.; Cardarelli, M.; Bonini, P.; Colla, G. Synergistic action of a microbial-based biostimulant and a plant derived-protein hydrolysate enhances lettuce tolerance to alkalinity and salinity. Front. Plant Sci. 2017, 8, 131. [Google Scholar] [CrossRef] [Green Version]
- Wilson, H.T.; Amirkhani, M.; Taylor, A.G. Evaluation of gelatin as a biostimulant seed treatment to improve plant performance. Front. Plant Sci. 2018, 9, 1006. [Google Scholar] [CrossRef] [Green Version]
- Schmitt, A.; Koch, E.; Stephan, D.; Kromphardt, C.; Jahn, M.; Krauthausen, H.-J.; Forsberg, G.; Werner, S.; Amein, T.; Wright, S.A.I.; et al. Evaluation of non-chemical seed treatment methods for the control of Phoma valerianellae on lamb’s lettuce seeds. J. Plant Dis. Prot. 2009, 116, 200–207. [Google Scholar] [CrossRef]
- Singh, U.B.; Malviya, D.; Khan, W.; Singh, S.; Karthikeyan, N.; Imran, M.; Rai, J.P.; Sarma, B.K.; Manna, M.C.; Chaurasia, R.; et al. Earthworm grazed-Trichoderma harzianum biofortified spent mushroom substrates modulate accumulation of natural antioxidants and bio-fortification of mineral nutrients in tomato. Front. Plant Sci. 2018, 9, 1017. [Google Scholar] [CrossRef] [PubMed]
- Yaronskaya, E.; Vershilovskaya, I.; Poers, Y.; Alawady, A.E.; Averina, N.; Grimm, B. Cytokinin effects on tetrapyrrole biosynthesis and photosynthetic activity in barley seedlings. Planta 2006, 224, 700–709. [Google Scholar] [CrossRef] [PubMed]
- Couto, C.A.; Peixoto, C.P.; Vieira, E.L.; Carvalho, E.V.; Peixoto, V.A.B. Action of cinetina, butyric acid and gibberellic acid on the emergency of sunflower under aluminum stress/Acao da cinetina, acido indolbutirico e acido giberelico na emergencia do girassol sob estresse por aluminio. Comun. Sci. 2012, 3, 206–210. [Google Scholar]
- Qiu, Y.; Amirkhani, M.; Mayton, H.; Chen, Z.; Taylor, A. Biostimulant seed coating treatments to improve cover crop germination and seedling growth. Agronomy 2020, 10, 154. [Google Scholar] [CrossRef] [Green Version]
- Parađiković, N.; Vinković, T.; Teklić, T.; Vlado, G.; Milaković, Z. Biostimulant application in tomato transplants production. In Proceedings of the 43. Hrvatski I 3. Međunarodni Simpozij Agronoma, Opatija, Croatia, 18–21 February 2008. [Google Scholar]
- Vinković, T.; Parađiković, N.; Teklić, T.; Lisjak, M.; Štolfa, I.; Špoljarević, M.; Balićević, R. Mineral composition of tomato under influence of biostimulant composition. In Proceedings of the 45. Hrvatski I 5. Međunarodni Simpozij Agronoma, Osijek, Hrvatsk, 15–19 February 2010. [Google Scholar]
- Parađiković, N.; Vinković, T.; Vrček, I.V.; Tkalec, M.; Lončarić, Z.; Milaković, Z. Ca status in pepper fruit and leaves under influence of biostimulants treatment. In Proceedings of the 46th Croatian and 6th International Symposium on Agriculture, Opatija, Croatia, 14–18 February 2011. [Google Scholar]
- Parađiković, N.; Vinković, T.; Vrček, I.V.; Tkalec, M. Natural biostimulants reduce the incidence of BER in sweet yellow pepper plants (Capsicum annuum L.). Agric. Food Sci. 2013, 22, 307–317. [Google Scholar] [CrossRef] [Green Version]
- Tkalec, M.; Vinković, T.; Baličević, R.; Parađiković, N. Influence of biostimulants on growth and development of bell pepper (Capsicum annuum L.). Acta Agric. Serbica 2010, 15, 83–88. [Google Scholar]
- Lisjak, M.; Tomić, O.; Špoljarević, M.; Teklić, T.; Stanisavljević, A.; Balas, J. Garden cress germinability and seedling vigour after treatment with plant extracts. Poljoprivreda 2015, 21, 41–46. [Google Scholar] [CrossRef]
- Dudaš, S.; Šola, I.; Sladonja, B.; Erhatić, R.; Ban, D.; Poljuha, D. The effect of biostimulant and fertilizer on “low input” lettuce production. Acta Bot. Croat. 2016, 75, 253–259. [Google Scholar] [CrossRef] [Green Version]
- Špoljarević, M.; Štolfa, I.; Lisjak, M.; Stanisavljević, A.; Vinković, T.; Agić, D.; Parađiković, N.; Teklić, T.; Engler, M.; Klešić, K. Strawberry (Fragaria x ananassa Duch) leaf antioxidative response to biostimulators and reduced fertilization with N and K. Poljoprivreda 2010, 16, 50–56. [Google Scholar]
- Stanisavljević, A.; Lisjak, M.; Špoljarević, M.; Vinković, T.; Agić, D.; Štolfa, I.; Welzer, F.; Parađiković, N.; Teklić, T. Biostimulators impact on productivity in the cultivation of strawberries on foil. In Proceedings of the 45. Hrvatski i 5. Međunarodni simpozij agronoma, Opatija, Croatia, 15–19 February 2010; pp. 1138–1142. [Google Scholar]
- Bogunovic, I.; Duralija, B.; Gadze, J.; Kisić, I. Biostimulant usage for preserving strawberries to climate damages. Hortic. Sci. 2016, 42, 132–140. [Google Scholar] [CrossRef] [Green Version]
- Zeljković, S.; Parađiković, N.; Šušak, U.; Tkalec, M. Growth and development of basil transplants (Ocimum basilicum L.) under biostimulants application. AГPO3HAЊE 2015, 15, 415–424. [Google Scholar] [CrossRef] [Green Version]
- Tkalec, M.; Parađiković, N.; Zeljković, S.; Vinković, T. Efficiency of biostimulant on growth and development of wild rose. In Proceedings of the 47. Hrvatski i 7. Međunarodni Simpozij Agronoma; Zagreb, Hrvatska, 13–17 February 2012. [Google Scholar]
- Zeljković, S.B.; Parađiković, N.A.; Babić, T.S.; Đurić, G.D.; Oljača, R.M.; Vinković, T.M.; Tkalec, M.B. Influence of biostimulant and substrate volume on root growth and development of scarlet sage (Salvia splendens L.) transplants. J. Agric. Sci. Belgrade 2010, 55, 29–36. [Google Scholar] [CrossRef] [Green Version]
- Zeljković, S.; Parađiković, N.; Vinković, T.; Tkalec, M. Biostimulant application in the production of seedlings of seasonal flowers. Agroznanje-Agro-Knowl. J. 2011, 12, 175–181. [Google Scholar]
- Parađiković, N.; Vinković, T.; Radman, D. Influence of biostimulant on seed germination of some flower species. Sjemenarstvo 2008, 25, 25–33. [Google Scholar]
- Souri, M.K.; Bakhtiarizade, M. Biostimulation effects of rosemary essential oil on growth and nutrient uptake of tomato seedlings. Sci. Hortic. 2019, 243, 472–476. [Google Scholar] [CrossRef]
- Drobek, M.; Frąc, M.; Cybulska, J. Plant biostimulants: Importance of the quality and yield of horticultural crops and the improvement of plant tolerance to abiotic stress—A review. Agronomy 2019, 9, 335. [Google Scholar] [CrossRef] [Green Version]
- Rouphael, Y.; Franken, P.; Schneider, C.; Schwarz, D.; Giovannetti, M.; Agnolucci, M.; De Pascale, S.; Bonini, P.; Colla, G. Arbuscular mycorrhizal fungi act as biostimulants in horticultural crops. Sci. Hortic. 2015, 196, 91–108. [Google Scholar] [CrossRef]
- Ugolini, L.; Cinti, S.; Righetti, L.; Stefan, A.; Matteo, R.; D’Avino, L.; Lazzeri, L. Production of an enzymatic protein hydrolyzate from defatted sunflower seed meal for potential application as a plant biostimulant. Ind. Crop. Prod. 2015, 75, 15–23. [Google Scholar] [CrossRef]
- Scaglia, B.; Pognani, M.; Adani, F. The anaerobic digestion process capability to produce biostimulant: The case study of the dissolved organic matter (DOM) vs. auxin-like property. Sci. Total Environ. 2017, 589, 36–45. [Google Scholar] [CrossRef]
- Pacholczak, A.; Szydło, W.; Jacygrad, E.; Federowicz, M. Effect of auxins and the biostimulator AlgaminoPlant on rhizogenesis in stem cuttings of two dogwood cultivars (Cornus alba ‘Aurea’and ‘Elegantissima’). Acta Sci. Pol. Hortorum Cultus 2012, 11, 93–103. [Google Scholar]
- Xu, C.; Leskovar, D.I. Effects of A. nodosum seaweed extracts on spinach growth, physiology and nutrition value under drought stress. Sci. Hortic. 2015, 183, 39–47. [Google Scholar] [CrossRef]
- Spann, T.M.; Little, H.A. Applications of a commercial extract of the brown seaweed ascophyllum nodosum increases drought tolerance in container-grown ‘hamlin’ sweet orange nursery trees. HortScience 2011, 46, 577–582. [Google Scholar] [CrossRef] [Green Version]
- Hernández-Herrera, R.M.; Santacruz-Ruvalcaba, F.; Ruiz-López, M.A.; Norrie, J.; Hernández-Carmona, G. Effect of liquid seaweed extracts on growth of tomato seedlings (Solanum lycopersicum L.). J. Appl. Phycol. 2013, 26, 619–628. [Google Scholar] [CrossRef]
- Xu, Z.; Jiang, Y.; Zhou, G. Response and adaptation of photosynthesis, respiration, and antioxidant systems to elevated CO2 with environmental stress in plants. Front. Plant Sci. 2015, 6, 701. [Google Scholar] [CrossRef] [Green Version]
- Kumari, R.; Kaur, I.; Bhatnagar, A.K. Effect of aqueous extract of Sargassum johnstonii Setchell & Gardner on growth, yield and quality of Lycopersicon esculentum Mill. J. Appl. Phycol. 2011, 23, 623–633. [Google Scholar]
- Colla, G.; Rouphael, Y.; Di Mattia, E.; El-Nakhel, C.; Cardarelli, M. Co-inoculation of Glomus intraradices and Trichoderma atroviride acts as a biostimulant to promote growth, yield and nutrient uptake of vegetable crops. J. Sci. Food Agric. 2015, 95, 1706–1715. [Google Scholar] [CrossRef]
- Mukherjee, P.K.; Horwitz, B.A.; Herrera-Estrella, A.; Schmoll, M.; Kenerley, C.M. Trichoderma research in the genome era. Annu. Rev. Phytopathol. 2013, 51, 105–129. [Google Scholar] [CrossRef]
- Nicolás, C.; Hermosa, R.; Rubio, B.; Mukherjee, P.K.; Monte, E. Trichoderma genes in plants for stress tolerance-status and prospects. Encycl. Appl. Plant Sci. 2014, 228, 71–78. [Google Scholar] [CrossRef]
- Usuki, F.; Narisawa, K. A mutualistic symbiosis between a dark septate endophytic fungus, Heteroconium chaetospira, and a nonmycorrhizal plant, Chinese cabbage. Mycologia 2007, 99, 175–184. [Google Scholar] [CrossRef]
- Tavarini, S.; Passera, B.; Martini, A.; Avio, L.; Sbrana, C.; Giovannetti, M.; Angelini, L.G. Plant growth, steviol glycosides and nutrient uptake as affected by arbuscular mycorrhizal fungi and phosphorous fertilization in Stevia rebaudiana Bert. Ind. Crop. Prod. 2018, 111, 899–907. [Google Scholar] [CrossRef]
- Gaiero, J.R.; McCall, C.A.; Thompson, K.A.; Day, N.J.; Best, A.S.; Dunfield, K.E. Inside the root microbiome: Bacterial root endophytes and plant growth promotion. Am. J. Bot. 2013, 100, 1738–1750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, D.; Zhao, H.; Zhao, D.; Zhu, X.; Wang, Y.; Duan, Y.; Xuan, Y.H.; Chen, L. Isolation and identification of bacteria from rhizosphere soil and their effect on plant growth promotion and root-knot nematode disease. Biol. Control 2018, 119, 12–19. [Google Scholar] [CrossRef]
- Dias, M.P.; Bastos, M.S.; Xavier, V.B.; Cassel, E.; Astarita, L.V.; Santarém, E.R. Plant growth and resistance promoted by Streptomyces spp. in tomato. Plant Physiol. Biochem. 2017, 118, 479–493. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Xu, Y.; Lai, X. Antagonistic endophytic bacteria associated with nodules of soybean (Glycine max L.) and plant growth-promoting properties. Braz. J. Microbiol. 2018, 49, 269–278. [Google Scholar] [CrossRef] [PubMed]
- Goñi, O.; Quille, P.; O’Connell, S. Ascophyllum nodosum extract biostimulants and their role in enhancing tolerance to drought stress in tomato plants. Plant Physiol. Biochem. 2018, 126, 63–73. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.-S.; Zou, Y.-N.; Abd-Allah, E.F. Mycorrhizal association and ROS in plants. In Oxidative Damage to Plants; Elsevier: Amsterdam, The Netherlands, 2014; pp. 453–475. [Google Scholar]
- Morales-Payan, J. Influence of foliar sprays of an amino acid formulation on fruit yield of ’edward’ mango. Acta Hortic. 2015, 157–159. [Google Scholar] [CrossRef]
- Tejada, M.; Rodríguez-Morgado, B.; Paneque, P.; Parrado, J. Effects of foliar fertilization of a biostimulant obtained from chicken feathers on maize yield. Eur. J. Agron. 2018, 96, 54–59. [Google Scholar] [CrossRef]
- Ramírez-Pérez, L.J.; Morales-Díaz, A.B.; Adalberto, B.M.; De-Alba-Romenus, K.; González-Morales, S.; Juárez-Maldonado, A. Dynamic modeling of cucumber crop growth and uptake of N, P and K under greenhouse conditions. Sci. Hortic. 2018, 234, 250–260. [Google Scholar] [CrossRef]
- Panfili, I.; Bartucca, M.L.; Del Buono, D. The treatment of duckweed with a plant biostimulant or a safener improves the plant capacity to clean water polluted by terbuthylazine. Sci. Total Environ. 2019, 646, 832–840. [Google Scholar] [CrossRef]
- García-Martínez, A.M.; Díaz, A.; Tejada, M.; Bautista, J.; Rodríguez, B.; Santa-Maria, C.; Revilla, E.; Parrado, J. Enzymatic production of an organic soil biostimulant from wheat-condensed distiller solubles: Effects on soil biochemistry and biodiversity. Process. Biochem. 2010, 45, 1127–1133. [Google Scholar] [CrossRef]
- Milić, B.; Kalajdžić, J.; Keserović, Z.; Magazin, N.; Miodragović, M.; Popara, G. Bioregulators can improve fruit size, yield and plant growth of northern highbush blueberry (Vaccinium corymbosum L.). Sci. Hortic. 2018, 235, 214–220. [Google Scholar] [CrossRef]
- Luziatelli, F.; Ficca, A.G.; Colla, G.; Švecová, E.B.; Ruzzi, M. Foliar application of vegetal-derived bioactive compounds stimulates the growth of beneficial bacteria and enhances microbiome biodiversity in lettuce. Front. Plant Sci. 2019, 10, 60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahnert, A.; Haratani, M.; Schmuck, M.; Berg, G. Enriching beneficial microbial diversity of indoor plants and their surrounding built environment with biostimulants. Front. Microbiol. 2018, 9, 2985. [Google Scholar] [CrossRef] [PubMed]
- Rouphael, Y.; Colla, G. Synergistic biostimulatory action: Designing the next generation of plant biostimulants for sustainable agriculture. Front. Plant Sci. 2018, 9, 1655. [Google Scholar] [CrossRef] [Green Version]
- Lucini, L.; Rouphael, Y.; Cardarelli, M.; Bonini, P.; Baffi, C.; Colla, G.; Lucini, L.; Rouphael, Y.; Cardarelli, M.; Bonini, P.; et al. A vegetal biopolymer-based biostimulant promoted root growth in melon while triggering brassinosteroids and stress-related compounds. Front. Plant Sci. 2018, 9, 472. [Google Scholar] [CrossRef] [Green Version]
- Ertani, A.; Francioso, O.; Tinti, A.; Schiavon, M.; Pizzeghello, D.; Nardi, S. Evaluation of seaweed extracts from Laminaria and Ascophyllum nodosum spp. as biostimulants in Zea mays L. using a combination of chemical, biochemical and morphological approaches. Front. Plant Sci. 2018, 9, 428. [Google Scholar] [CrossRef]
- Palumbo, G.; Schiavon, M.; Nardi, S.; Ertani, A.; Celano, G.; Colombo, C.M. Biostimulant potential of humic acids extracted from an amendment obtained via combination of Olive Mill Wastewaters (OMW) and a pre-treated organic material derived from Municipal Solid Waste (MSW). Front. Plant Sci. 2018, 9, 1028. [Google Scholar] [CrossRef]
- Van Oosten, M.J.; Pepe, O.; De Pascale, S.; Silletti, S.; Maggio, A. The role of biostimulants and bioeffectors as alleviators of abiotic stress in crop plants. Chem. Biol. Technol. Agric. 2017, 4, 5. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.T.; Janisiewicz, W.J.; Liu, Z.; Callahan, A.M.; Evans, B.E.; Jurick, W.M.I.; Dardick, C. Exposure in vitro to an Environmentally Isolated Strain TC09 of Cladosporium sphaerospermum triggers plant growth promotion, early flowering, and fruit yield increase. Front. Plant Sci. 2019, 9, 1959. [Google Scholar] [CrossRef] [Green Version]
- Cristiano, G.; Pallozzi, E.; Conversa, G.; Tufarelli, V.; De Lucia, B. Effects of an animal-derived biostimulant on the growth and physiological parameters of potted snapdragon (Antirrhinum majus L.). Front. Plant Sci. 2018, 9, 861. [Google Scholar] [CrossRef]
- Sharma, A.; Kumar, V.; Yuan, H.; Kanwar, M.K.; Bhardwaj, R.; Thukral, A.K.; Zheng, B. Jasmonic acid seed treatment stimulates insecticide detoxification in Brassica juncea L. Front. Plant Sci. 2018, 9, 1609. [Google Scholar] [CrossRef] [PubMed]
- Aamir, M.; Kashyap, S.P.; Zehra, A.; Dubey, M.K.; Singh, V.K.; Ansari, W.A.; Upadhyay, R.S.; Singh, S. Trichoderma erinaceum bio-priming modulates the wrkys defense programming in tomato against the Fusarium oxysporum f. sp. lycopersici (Fol) challenged condition. Front. Plant Sci. 2019, 10, 911. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cramer, G.R.; Urano, K.; Velrot, S.; Pezzotti, M.; Shinozaki, K. Effects of abiotic stress on plants: A systems biology perspective. BMC Plant Biol. 2011, 11, 163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Vasconcelos, A.C.F.; Chaves, L.H.G. Biostimulants and their role in improving plant growth under abiotic stresses. In Biostimulants in Plant Science; Mirmajlessi, S.M., Radhakrishnan, R., Eds.; IntechOpen: London, UK, 2020. [Google Scholar]
- Cortivo, C.D.; Conselvan, G.B.; Carletti, P.; Barion, G.; Sella, L.; Vamerali, T. Biostimulant effects of seed-applied sedaxane fungicide: Morphological and physiological changes in maize seedlings. Front. Plant Sci. 2017, 8, 8. [Google Scholar] [CrossRef] [Green Version]
- Huang, J.; Levine, A.; Wang, Z. Plant abiotic stress. Sci. World J. 2013, 2013, 432836. [Google Scholar] [CrossRef] [Green Version]
- Teale, W.D.; Paponov, I.; Palme, K. Auxin in action: Signalling, transport and the control of plant growth and development. Nat. Rev. Mol. Cell Biol. 2006, 7, 847–859. [Google Scholar] [CrossRef]
- Tokas, J.; Punia, H. Milk Fat Globule Membrane (MFGM): An ingredient of dairy products as nutraceutical. Trends Pept. Protein Sci. 2019, 4, 1–7. [Google Scholar]
- Punia, H.; Bhadu, S.; Tokas, J. Nanotechnology and nanomedicine: Going small means aiming big. In Advances in Biochemistry & Applications in Medicine; Shrestha, R., Ed.; Open Access eBooks: Las Vegas, Nevada, USA, 2018. [Google Scholar]
- Punia, H.; Tokas, J.; Bhadu, S.; Mohanty, A.K.; Rawat, P.; Malik, A.; Singh, S. Proteome dynamics and transcriptome profiling in sorghum [Sorghum bicolor (L.) Moench] under salt stress. 3 Biotech 2020, 10, 1–10. [Google Scholar] [CrossRef]
- Taiz, L.; Zeiger, E. Stress physiology. Plant Physiol. 2006, 91, 750–751. [Google Scholar]
- Santos, C.M.G.; Vieira, E.L. Efeito de bioestimulante na germinação de sementes, vigor de plântulas e crescimento inicial do algodoeiro. Magistra Cruz Das Almas 2005, 17, 124–130. [Google Scholar]
- Salehi-Lisar, S.Y.; Bakhshayeshan-Agdam, H. Drought stress in plants: Causes, consequences, and tolerance. In Drought Stress Tolerance in Plants; Hossain, M.A., Wani, S.H., Bhattacharjee, S., Burritt, D.J., Tran, L.-S.P., Eds.; Springer: Cham, Switzerland, 2016; Volume 1, pp. 1–16. [Google Scholar]
- Lana, R.M.Q.; Lana, A.M.Q.; Gozuen, C.F.; Bonotto, I.; Trevisan, L.R. Aplicação de reguladores de crescimento na cultura do feijoeiro. Biosci. J. 2009, 25, 13–20. [Google Scholar]
- De Carvalho, T.C.; da Silva, S.S.; da Silva, R.C.; Panobianco, M.; Mógor, Á.F. Influência de bioestimulantes na germinação e desenvolvimento de plântulas de Phaseolus vulgaris sob restrição hídrica. Rev. Ciências Agrárias 2013, 36, 199–205. [Google Scholar]
- Fioreze, S.L.; Tochetto, C.; Coelho, A.E.; Melo, H.F. Effects of calcium supply on soybean plants. Comun. Sci. 2018, 9, 219–225. [Google Scholar] [CrossRef]
- Almeida, M.S.; Nunes, A.S.; Casagrande, R.R. Aplicação foliar de bioestimulante em híbrido de milho com e sem déficit hídrico. In Proceedings of the XIV Semin. Nac. Milho Safrinha, Cuiabá, Brazil, 21–23 November 2017; pp. 146–151. [Google Scholar]
- De Oliveira, F.A.; de Medeiros, J.F.; de Alves, R.C.; Lima, L.A.; dos Santos, S.T.; de Régis, L.R.L. Produção de feijão caupi em função da salinidade e regulador de crescimento. Rev. Bras. Eng. Agrícola Ambient. 2015, 19, 1049–1056. [Google Scholar] [CrossRef] [Green Version]
- Filho, H.C.L.W. Uso de Bioestimulantes e Enraizadores no Crescimento Inicial e Tolerância à Seca Em Cana-de-Açúcar [Dissertação]; Rio Largo Universidade Federal de Alagoas: Masejo, Alagoas, Brazil, 2011. [Google Scholar]
- Parihar, P.; Singh, S.; Singh, R.; Singh, V.P.; Prasad, S.M. Effect of salinity stress on plants and its tolerance strategies: A review. Environ. Sci. Pollut. Res. 2015, 22, 4056–4075. [Google Scholar] [CrossRef]
- de Lacerda, C.F.; Costa, R.N.T.; Bezerra, M.A.; Gheyi, H.R. Estratégias de Manejo Para Uso de Água Salina na Agricultura; Embrapa Agroindústria Tropical—Capítulo em livro científico (ALICE): Fortaleza, Brazil, 2010. [Google Scholar]
- Türkmen, Ö.; Dursun, A.; Turan, M.; Erdinç, Ç. Calcium and humic acid affect seed germination, growth, and nutrient content of tomato (Lycopersicon esculentum L.) seedlings under saline soil conditions. Acta Agric. Scand. Sect. B Plant Soil Sci. 2004, 54, 168–174. [Google Scholar] [CrossRef]
- Aktar, M.; Ali, A.Z.; Rahman, M. Application of nitrogen and boron on growth and nutrient contents of okra (Abelmoschus Esculentus L.) grown on soil. J. Asiat. Soc. Bangladesh Sci. 2015, 41, 173–181. [Google Scholar] [CrossRef]
- Aydin, A. Humic acid application alleviate salinity stress of bean (Phaseolus vulgaris L.) plants decreasing membrane leakage. Afr. J. Agric. Res. 2012, 7, 1073–1086. [Google Scholar] [CrossRef]
- Akaighe, N.; MacCuspie, R.I.; Navarro, D.A.; Aga, D.S.; Banerjee, S.; Sohn, M.; Sharma, V.K. Humic acid-induced silver nanoparticle formation under environmentally relevant conditions. Environ. Sci. Technol. 2011, 45, 3895–3901. [Google Scholar] [CrossRef]
- Azevedo, R.A.; Lea, P.J. Research on abiotic and biotic stress–What next? Ann. Appl. Biol. 2011, 159, 317–319. [Google Scholar] [CrossRef]
- García, A.C.; Santos, L.A.; Izquierdo, F.G.; Sperandio, M.V.L.; Castro, R.N.; Berbara, R.L.L. Vermicompost humic acids as an ecological pathway to protect rice plant against oxidative stress. Ecol. Eng. 2012, 47, 203–208. [Google Scholar] [CrossRef]
- de Neta, M.L.S.; de Oliveira, F.A.; Torres, S.B.; Souza, A.A.T.; da Silva, D.D.A.; dos Santos, S.T. Gherkin cultivation in saline medium using seeds treated with a biostimulant. Acta Sci. Agron. 2018, 40, e35216. [Google Scholar] [CrossRef] [Green Version]
- de Oliveira, F.A.; de Medeiros, J.F.; da Cunha, R.C.; de Souza, M.W.L.; Lima, L.A. Uso de bioestimulante como agente amenizador do estresse salino na cultura do milho pipoca. Rev. Ciência Agronômica 2016, 47, 307–315. [Google Scholar]
- Bulgari, R.; Trivellini, A.; Ferrante, A. Effects of two doses of organic extract-based biostimulant on greenhouse lettuce grown under increasing NaCl concentrations. Front. Plant Sci. 2019, 9, 9. [Google Scholar] [CrossRef] [PubMed]
- El-Baky, H.H.A.; Hussein, M.M.; El-Baroty, G.S. Algal extracts improve antioxidant defense abilities and salt tolerance of wheat plant irrigated with sea water. Afr. J. Biochem. Res. 2008, 2, 151–164. [Google Scholar]
- Nabati, D.A.; Schmidt, R.E.; Parrish, D.J. Alleviation of salinity stress in kentucky bluegrass by plant growth regulators and iron. Crop. Sci. 1994, 34, 198–202. [Google Scholar] [CrossRef]
- Latef, A.A.H.A.; Chaoxing, H. Effect of arbuscular mycorrhizal fungi on growth, mineral nutrition, antioxidant enzymes activity and fruit yield of tomato grown under salinity stress. Sci. Hortic. 2011, 127, 228–233. [Google Scholar] [CrossRef]
- Le Mire, G.; Nguyen, M.; Fassotte, B.; du Jardin, P.; Verheggen, F.; Delaplace, P.; Jijakli, H. Implementing biostimulants and biocontrol strategies in the agroecological management of cultivated ecosystems. Biotechnol. Agron. Société Environ. 2016, 20, 299–313. [Google Scholar]
- Upadhyay, S.K.; Singh, D.P. Effect of salt-tolerant plant growth-promoting rhizobacteria on wheat plants and soil health in a saline environment. Plant Biol. 2014, 17, 288–293. [Google Scholar] [CrossRef]
- Kai, H.; Iba, K. Temperature stress in plants. In eLS; John Wiley & Sons, Ltd: Hoboken, NJ, USA, 2014. [Google Scholar]
- Vendruscolo, E.P.; Martins, A.P.B.; Campos, L.F.C.; Seleguini, A.; dos Santos, M.M. Amenização de estresse térmico via aplicação de bioestimulante em sementes de meloeiro cantaloupe/thermal stress alleviation by biostimulant application on cantaloupe melon seeds. Rev. Bras. Eng. Biossistemas 2016, 10, 241–247. [Google Scholar] [CrossRef] [Green Version]
- Yildirim, E.; Dursun, A.; Güvenc, I.; Kumlay, A. The effects of different salt, biostimulant and temperature levels on seed germination of some vegetable species. Acta Hortic. 2002, 249–253. [Google Scholar] [CrossRef]
- Polo, J.; Barroso, R.; Ródenas, J.; Azcón-Bieto, J.; Cáceres, R.; Marfà, O. Porcine hemoglobin hydrolysate as a biostimulant for lettuce plants subjected to conditions of thermal stress. HortTechnology 2005, 16, 483–487. [Google Scholar] [CrossRef] [Green Version]
- Marfà, O.; Cáceres, R.; Polo, J.; Ródenas, J. Animal protein hydrolysate as a biostimulant for transplanted strawberry plants subjected to cold stress. Acta Hortic. 2009, 315–318. [Google Scholar] [CrossRef]
- Botta, A. Enhancing plant tolerance to temperature stress with amino acids: An approach to their mode of action. Acta Hortic. 2013, 29–35. [Google Scholar] [CrossRef]
- Xu, Y.; Huang, B. Responses of creeping bentgrass to trinexapac-ethyl and biostimulants under summer stress. HortScience 2010, 45, 125–131. [Google Scholar] [CrossRef] [Green Version]
- Turner, N.C.; Kramer, P.J. Adaptation of Plants to Water and High Temperature Stress; Library of Congress Cataloging in Publication Data; John Wiley & Sons, Inc: Hoboken, NJ, USA, 1980. [Google Scholar]
- Kauffman, G.L.; Kneivel, D.P.; Watschke, T.L. Effects of a biostimulant on the heat tolerance associated with photosynthetic capacity, membrane thermostability, and polyphenol production of perennial ryegrass. Crop. Sci. 2007, 47, 261–267. [Google Scholar] [CrossRef]
- Ali, S.Z.; Sandhya, V.; Grover, M.; Linga, V.R.; Bandi, V. Effect of inoculation with a thermotolerant plant growth promoting Pseudomonas putida strain AKMP7 on growth of wheat (Triticum spp.) under heat stress. J. Plant Interact. 2011, 6, 239–246. [Google Scholar] [CrossRef] [Green Version]
- Dobbelaere, S.; Croonenborghs, A.; Thys, A.; Broek, A.V.; Vanderleyden, J. Phytostimulatory effect of Azospirillum brasilense wild type and mutant strains altered in IAA production on wheat. Plant Soil 1999, 212, 153–162. [Google Scholar] [CrossRef]
- Huang, J.; Gu, M.; Lai, Z.; Fan, B.; Shi, K.; Zhou, Y.-H.; Yu, J.-Q.; Chen, Z. Functional Analysis of the Arabidopsis PAL Gene Family in Plant Growth, Development, and Response to Environmental Stress. Plant Physiol. 2010, 153, 1526–1538. [Google Scholar] [CrossRef] [Green Version]
- Shabala, L.; Mackay, A.; Tian, Y.; Jacobsen, S.-E.; Zhou, D.; Shabala, S. Oxidative stress protection and stomatal patterning as components of salinity tolerance mechanism in quinoa (Chenopodium quinoa). Physiol. Plant. 2012, 146, 26–38. [Google Scholar] [CrossRef]
- Han, M.; Li, G.; Liu, X.; Li, A.; Mao, P.; Liu, P.; Li, H.-H. Phenolic profile, antioxidant activity and anti-proliferative activity of crabapple fruits. Hortic. Plant J. 2019, 5, 155–163. [Google Scholar] [CrossRef]
- Abdalla, M.M. Boosting the growth of rocket plants in response to the application of Moringa oleifera extracts as a biostimulant. Life Sci. J. 2014, 11, 1113–1121. [Google Scholar]
- Rouphael, Y.; Colla, G.; Giordano, M.; El-Nakhel, C.; Kyriacou, M.C.; De Pascale, S. Foliar applications of a legume-derived protein hydrolysate elicit dose-dependent increases of growth, leaf mineral composition, yield and fruit quality in two greenhouse tomato cultivars. Sci. Hortic. 2017, 226, 353–360. [Google Scholar] [CrossRef]
- Tarantino, A.; Lops, F.; Disciglio, G.; Lopriore, G. Effects of plant biostimulants on fruit set, growth, yield and fruit quality attributes of ‘Orange rubis®’ apricot (Prunus armeniaca L.) cultivar in two consecutive years. Sci. Hortic. 2018, 239, 26–34. [Google Scholar] [CrossRef]
- Kumaraswamy, R.; Kumari, S.; Choudhary, R.C.; Sharma, S.; Pal, A.; Raliya, R.; Biswas, P.; Saharan, V. Salicylic acid functionalized chitosan nanoparticle: A sustainable biostimulant for plant. Int. J. Biol. Macromol. 2019, 123, 59–69. [Google Scholar] [CrossRef]
- Jindo, K.; Martim, S.A.; Navarro, E.C.; Pérez-Alfocea, F.; Hernandez, T.; Garcia, C.; Aguiar, N.O.; Canellas, L.P. Root growth promotion by humic acids from composted and non-composted urban organic wastes. Plant Soil 2012, 353, 209–220. [Google Scholar] [CrossRef]
- Ertani, A.; Schiavon, M.; Muscolo, A.; Nardi, S. Alfalfa plant-derived biostimulant stimulate short-term growth of salt stressed Zea mays L. plants. Plant Soil 2012, 364, 145–158. [Google Scholar] [CrossRef]
- Chen, T.H.H.; Murata, N. Glycinebetaine protects plants against abiotic stress: Mechanisms and biotechnological applications. Plant Cell Environ. 2010, 34, 1–20. [Google Scholar] [CrossRef]
- Hayat, S.; Ahmad, H.; Ali, M.; Ren, K.; Cheng, Z. Aqueous garlic extract stimulates growth and antioxidant enzymes activity of tomato (Solanum lycopersicum). Sci. Hortic. 2018, 240, 139–146. [Google Scholar] [CrossRef]
- Rehman, H.U.; Alharby, H.F.; Alzahrani, Y.; Rady, M.M. Magnesium and organic biostimulant integrative application induces physiological and biochemical changes in sunflower plants and its harvested progeny on sandy soil. Plant Physiol. Biochem. 2018, 126, 97–105. [Google Scholar] [CrossRef]
- Kulkarni, M.G.; Rengasamy, K.R.; Pendota, S.C.; Gruz, J.; Plačková, L.; Novák, O.; Doležal, K.; Amoo, S.O. Bioactive molecules derived from smoke and seaweed Ecklonia maxima showing phytohormone-like activity in Spinacia oleracea L. N. Biotechnol. 2019, 48, 83–89. [Google Scholar] [CrossRef] [PubMed]
- López-López, N.; López, A. Compost based ecological growing media according EU eco-label requirements. Sci. Hortic. 2016, 212, 1–10. [Google Scholar] [CrossRef]
- Giménez, A.; Fernández, J.A.; Pascual, J.A.; Ros, M.; López-Serrano, M.; Egea-Gilabert, C. An agroindustrial compost as alternative to peat for production of baby leaf red lettuce in a floating system. Sci. Hortic. 2019, 246, 907–915. [Google Scholar] [CrossRef]
- Sestili, F.; Rouphael, Y.; Cardarelli, M.; Pucci, A.; Bonini, P.; Canaguier, R.; Colla, G. Protein hydrolysate stimulates growth in tomato coupled with n-dependent gene expression involved in N assimilation. Front. Plant Sci. 2018, 9, 1233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fiorentino, N.; Ventorino, V.; Woo, S.L.; Pepe, O.; De Rosa, A.; Gioia, L.; Romano, I.; Lombardi, N.; Napolitano, M.; Colla, G.; et al. Trichoderma-based biostimulants modulate rhizosphere microbial populations and improve N uptake efficiency, yield, and nutritional quality of leafy vegetables. Front. Plant Sci. 2018, 9, 743. [Google Scholar] [CrossRef] [Green Version]
- Koskey, G.; Mburu, S.W.; Kimiti, J.M.; Ombori, O.; Maingi, J.M.; Njeru, E.M. Genetic characterization and diversity of Rhizobium isolated from root nodules of mid-altitude climbing bean (Phaseolus vulgaris L.) Varieties. Front. Microbiol. 2018, 9, 968. [Google Scholar] [CrossRef] [Green Version]
- Kamran, S.; Shahid, I.; Baig, D.N.; Rizwan, M.; Malik, K.A.; Mehnaz, S. Contribution of zinc solubilizing bacteria in growth promotion and zinc content of wheat. Front. Microbiol. 2017, 8, 2593. [Google Scholar] [CrossRef] [Green Version]
- Trejo-Téllez, L.I.; Estrada-Ortiz, E.; Gómez-Merino, F.C.; Becker, C.; Krumbein, A.; Schwarz, D. Flavonoid, nitrate and glucosinolate concentrations in Brassica species are differentially affected by photosynthetically active radiation, phosphate and phosphite. Front. Plant Sci. 2019, 10, 371. [Google Scholar] [CrossRef]
- Soppelsa, S.; Kelderer, M.; Casera, C.; Bassi, M.; Robatscher, P.; Andreotti, C. Use of biostimulants for organic apple production: Effects on tree growth, yield, and fruit quality at harvest and during storage. Front. Plant Sci. 2018, 9, 1342. [Google Scholar] [CrossRef] [PubMed]
- D’Amato, R.; De Feudis, M.; Hasuoka, P.E.; Regni, L.; Pacheco, P.H.; Onofri, A.; Businelli, D.; Proietti, P. The selenium supplementation influences olive tree production and oil stability against oxidation and can alleviate the water deficiency effects. Front. Plant Sci. 2018, 9, 1191. [Google Scholar] [CrossRef] [PubMed]
- Vergara, A.E.; Díaz, K.; Carvajal, R.; Espinoza, L.; Alcalde, J.A.; Pérez-Donoso, A.G. Exogenous applications of brassinosteroids improve color of red table grape (Vitis vinifera L. Cv. “Redglobe”) berries. Front. Plant Sci. 2018, 9, 363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koyama, R.; Roberto, S.R.; De Souza, R.T.; Borges, W.F.S.; Anderson, M.; Waterhouse, A.L.; Cantu, D.; Fidelibus, M.W.; Blanco-Ulate, B. Exogenous abscisic acid promotes anthocyanin biosynthesis and increased expression of flavonoid synthesis genes in Vitis vinifera × Vitis labrusca table grapes in a subtropical region. Front. Plant Sci. 2018, 9, 323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ugena, L.; Hýlová, A.; Podlešáková, K.; Humplík, J.F.; Doležal, K.; Diego, N.; De Spíchal, L. Characterization of biostimulant mode of action using novel multi-trait high-throughput screening of Arabidopsis germination and rosette growth. Front. Plant Sci. 2018, 9, 1327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paul, K.; Sorrentino, M.; Lucini, L.; Rouphael, Y.; Cardarelli, M.; Bonini, P.; Reynaud, H.; Canaguier, R.; Trtílek, M.; Panzarová, K.; et al. Understanding the biostimulant action of vegetal-derived protein hydrolysates by high-throughput plant phenotyping and metabolomics: A case study on tomato. Front. Plant Sci. 2019, 10, 47. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Biostimulants | Bioactive Components |
|---|---|
| Algreen ® | Seaweed extract (Ascophyllum nodosum, Laminaria sp., Sargassum spp.,), alginic acid; free amino acids, plant hormones, vitamins |
| Asahi SL (Atonik) ® | 0.1% sodium 5-nitroguaiacolate, 0.2% sodium ortho-nitrophenolate, and 0.3% sodium para-nitrophenolate |
| Bio-algeenS-92 ® | A. nodosum extract |
| Benefit ® | Free amino acids, vitamins, and nucleotides |
| Biozyme ® | A. nodosum extract, chelated micronutrients, zeatin, GA3, and IAA |
| Biplantol ®® | Universal macro-and microelements, uronic acids |
| Bio Rhizotonic ® | Algae extract, vitamins |
| Bio Root ® | Plant-based organic acids, soybean meal, alfalfa, rock phosphate, K-sulfate, and brewer’s yeast |
| Ergonfill ® | Keratin derivatives, protein hydrolysates, and cysteine |
| Equisetum extract (Acker-Schachtelhalm extrakt) | Flavonoids, plant acids, glycosides, Si |
| Fermented plant extract (Fermentierter Pflanzenextrakt ®) | Lactic acid bacteria, sugarcane molasses, yeasts, photosynthetic bacteria, pepper extracts, grasses, and garlic |
| Goëmar BM 86 ® | Algae A. nodosum extract |
| Grow-plex SP ® | Liquid humate |
| Kendal ® | Glutathione, protein hydrolysate, oligosaccharides, saponins, urea, |
| KE-Plantasalva ® | Bio-molasses, herbs extracts |
| Megafol ® | Auxin, amino acids, cytokines, gibberellins, betaines, vitamins |
| Radifarm ® | Amino acids, betaines, glycosides, microelements, organic acids, polysaccharides, saponins, vitamins |
| Roots 2 | Seaweed, vitamins, humic acid |
| Root Juice | Fulvic acid, humic acid, seaweed extract |
| Root & Shoot Builder | Amino acids, natural chelating agents, micronutrients, A. nodosum |
| Ruter AA ® | Micro- and Macronutrients, amino acids, |
| Slavol ® | Phosphate-mineralizing bacteria, auxins, nitrogen-fixing bacteria |
| Tablet ® | Trichoderma atroviride and Rhizophagus intraradices spores |
| Terra-Sorb ® | byproduct of enzymatic hydrolysis of amino acids |
| Tytanit ® | Titanium ascorbate |
| Stimulate ® | auxin, cytokinin, and gibberellic acid |
| Retrosal ® | zinc and calcium |
| Viva ® | Polysaccharides, proteins, polypeptides, amino acids, vitamin complexes, and humic acid |
| Plant Species | Biostimulants | Developmental Stage | Expected Outcomes | References |
|---|---|---|---|---|
| Tomato (Solanum lycopersicum L.) | Radifarm ® | Transplants | Enhanced roots growth | [52] |
| Radifarm ® + Megafol ® | Transplants | Improved nutrients uptake and distribution | [53] | |
| Bell peppers (Capsicum annuum L.) | Radifarm ® + Megafol ® | Fruit bearing | Increased macro- and micronutrient, especially Ca2+ ion concentration in leaves and fruits | [54] |
| Benefit ®; Megafol ®; Radifarm ®; Viva ® | From transplanting to harvest | Improved fruit yield | [55] | |
| Benefit ®; Megafol ®; Radifarm ®; Viva ® | 7th day seedlings | Increased fruit yield | [56] | |
| Garden cress (Lepidium sativum) L. | Acker-Schachtelhalm Extract ®; Biplantol Universal ®; Fermentierter ® Pflanzenex trakt ®; KE-Plantasalva ® | Germination | Improved water uptake and seedling growth, germination rate | [57] |
| Garlic (Allium sativum L.) | Radifarm ® | After transplanting | Increased seedling after transplanting | [44] |
| Lettuce (Lactuca sativa L.) | Bio-algeenS-90 ® | After transplanting | Improved growth and yield characteristics; Increased ascorbic acid content and dry matter and reduced pH | [58] |
| Strawberry (Fragaria x ananassa Duch) | Megafol ® + Viva ® | Fruit bearing | Activated antioxidative defense mechanism; decreased NK fertilization; enhanced fresh and dry weight | [59] |
| Kendal ® + Megafol ® + Viva ® | Flowering and fruit bearing | Increased fruit yield per plant | [60] | |
| Porcine blood-based biostimulant | Before flowering and onset of fruit ripening | Enhanced frost resistance, fruit weight; non-significant effects on fruit yield | [61] | |
| Basil (Ocimum basilicum L.) | Radifarm ® | Transplants | Increased above ground parts and root biomass | [62] |
| Dog rose (Rosa canina L.) | Radifarm ® | Robust growth in tissue culture | Enhanced root intensification | [63] |
| Radifarm ® | After transplanting | Increased seedling growth after transplanting | [44] | |
| Wax Begonia (Begonia semperflorens L.) | Radifarm ® | Transplants | Increased nutrient uptake with improved growth | [64] |
| Radifarm ® | After transplanting | Positive effects on morphological traits; enhanced nutrient uptake and proline level | [65] | |
| Marigold (Tagetes erecta L.) | Radifarm ® | Germination | Enhanced seedling fresh weight and germination energy | [66] |
| Primrose (Primula acaulis L.) | Radifarm ® | Transplants | Increased above ground parts and root intensification | [67] |
| Scarlet sage (Salvia splendens L.) | Radifarm ® | Transplants | Improved root mass and above ground parts | [64] |
| Source | Bioformulations | Plant Response |
|---|---|---|
| Consortium of beneficial fungi | Heteroconium chaetospira, Glomus viscosum, Glomus claroideum, Rhymbocarpus aggregatus, Glomus etunicatum, Trichoderma spp., Rhizophagus intraradices | The beneficial fungi promotes the growth and yield in tomato fruit [88] Stimulates protection against oxidative stress [89] |
| Marine algal biopreparations | Gelidium pectinutum, Sargassum wightii, Enteromorpha intestinalis, A. nodosum, Ecklonia maxima | Enhanced antioxidant capacity, chelation, extended shelf life of fruits, thermal and drought resistance [39,88] |
| Hydrolytic products | alfalfa hay, fruit and vegetable waste, pulses; natural and chemical (feathers, skin collagen animal tissue, casein, bone meal, fish waste | Improved yield [90], Enhanced NPK content and macro- and micronutrients in leaves [91,92] High protein content in cereals [91] Biotic and abiotic stresses tolerance [93] Improved soil fertility [94] |
| Anaerobic digested products | Lignin biomass, plants, and animal | Auxin-like properties [71,95] Improved nutrient availability [96] |
| Biostimulants Source | Cellular Mechanism | Physiological Mechanism | Agricultural Benefits | Environmental Benefits | References |
|---|---|---|---|---|---|
| Humic acids | Induce proton pumping, ATPases activity promote cell elongation and cell wall loosening | Enhanced accumulation of root biomass | Enhanced nutrient efficiency and root foraging ability | Improved yield and reduced utilization of fertilizers | [159] |
| Seaweed extracts | Upregulation of micronutrient transports encoding gens by application of A. nodosum (Brassica napus) | Increased root mass and mineral uptake | Enhanced accumulation of minerals in plant tissue | Biofortification of micronutrients (Mg, Fe, Cu, Zn) | [160] |
| Protein hydrolysate | Stimulation of biosynthesis of phenylalanine ammonia-lyase (PAL) via. enzymatic hydrolysis of alfalfa (Medicago sativa) and accumulation of flavonoids under salt stress | Protection against oxidative damage and UV rays | Improved crop resistance to abiotic stress (salt stress) | Improved crop yield under stress conditions (high salinity) | [153,161] |
| Glycine betaine | Protection against salt induced photodamage in quinoa | Maintenance of photosynthetic activity under salinity | Improved crop resistance to abiotic stress (salt stress) | Improved crop yield under stress conditions (high salinity) | [154,162] |
| Plant growth promoting Rhizobacteria | Induced auxin signaling pathways in roots via. application of Azospirillum brasilense wheat (Triticum aestivum) | Improved root mass and intensity | Enhanced nutrient efficiency and root foraging ability | Improved yield and reduced utilization of fertilizers | [152] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Malik, A.; Mor, V.S.; Tokas, J.; Punia, H.; Malik, S.; Malik, K.; Sangwan, S.; Tomar, S.; Singh, P.; Singh, N.; et al. Biostimulant-Treated Seedlings under Sustainable Agriculture: A Global Perspective Facing Climate Change. Agronomy 2021, 11, 14. https://doi.org/10.3390/agronomy11010014
Malik A, Mor VS, Tokas J, Punia H, Malik S, Malik K, Sangwan S, Tomar S, Singh P, Singh N, et al. Biostimulant-Treated Seedlings under Sustainable Agriculture: A Global Perspective Facing Climate Change. Agronomy. 2021; 11(1):14. https://doi.org/10.3390/agronomy11010014
Chicago/Turabian StyleMalik, Anurag, Virender S. Mor, Jayanti Tokas, Himani Punia, Shweta Malik, Kamla Malik, Sonali Sangwan, Saurabh Tomar, Pradeep Singh, Nirmal Singh, and et al. 2021. "Biostimulant-Treated Seedlings under Sustainable Agriculture: A Global Perspective Facing Climate Change" Agronomy 11, no. 1: 14. https://doi.org/10.3390/agronomy11010014
APA StyleMalik, A., Mor, V. S., Tokas, J., Punia, H., Malik, S., Malik, K., Sangwan, S., Tomar, S., Singh, P., Singh, N., Himangini, Vikram, Nidhi, Singh, G., Vikram, Kumar, V., Sandhya, & Karwasra, A. (2021). Biostimulant-Treated Seedlings under Sustainable Agriculture: A Global Perspective Facing Climate Change. Agronomy, 11(1), 14. https://doi.org/10.3390/agronomy11010014