Contribution of Agro-Environmental Factors to Yield and Plant Diversity of Olive Grove Ecosystems (Olea europaea L.) in the Mediterranean Landscape



Abstract
:1. Introduction
2. Materials and Methods
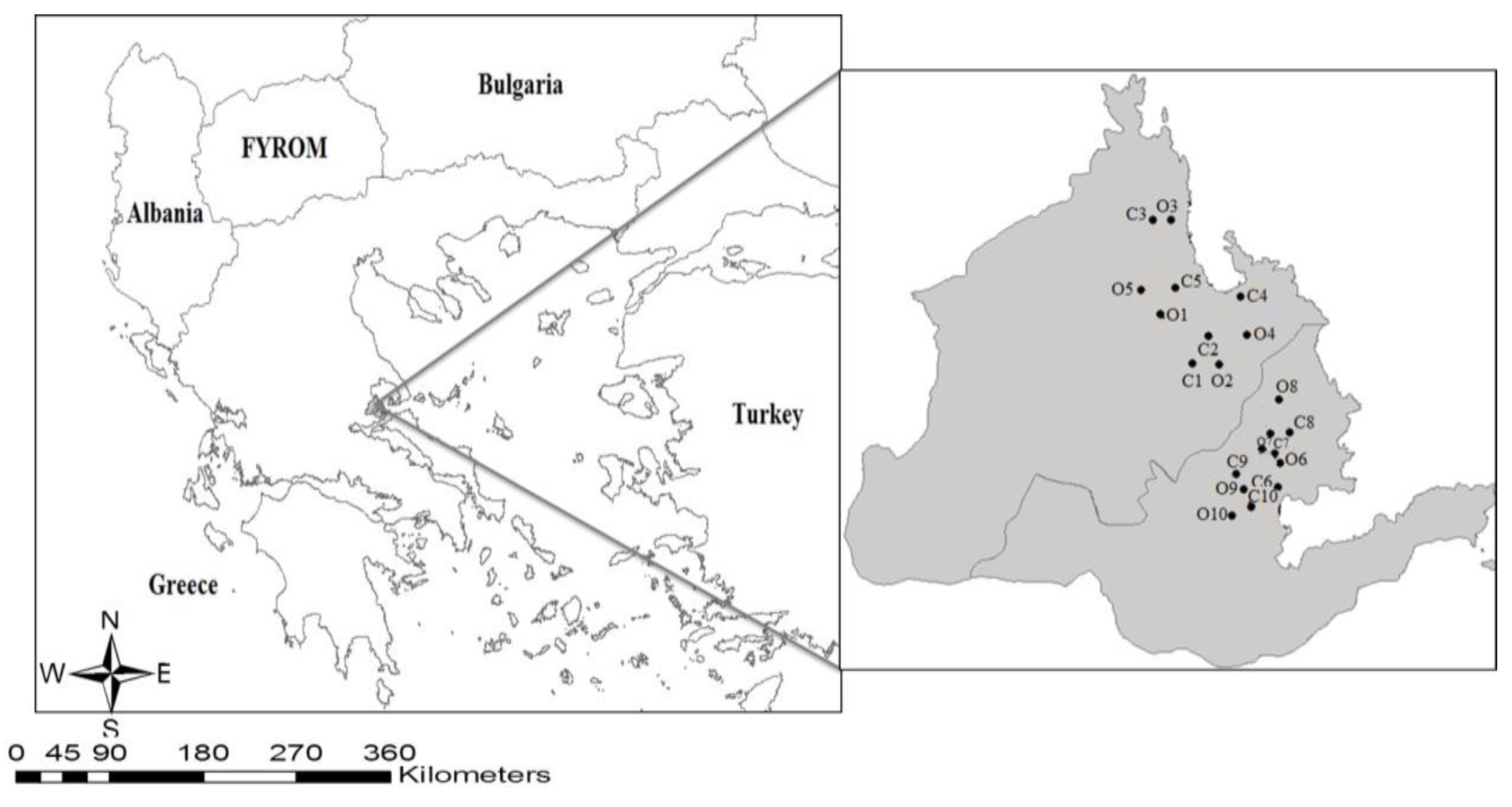
2.1. Study Area
2.2. Sampling
2.3. Statistical Analyses
- Y = the dependent variable (also called the predicted variable).
- β1,β2…βp = a weight (also called a coefficient). Determines how much weight one variable contributes to the model. If everything in the equation holds constant, b0 gives the predicted change in Y for a unit change in X.
- X1,2…p = a variable.
- β0 = the intercept—always a constant.
3. Results
3.1. Factors Influencing Yield of Olive Grove Management Systems
3.2. Factors Influencing Herbaceous Plant Species Richness of Olive Grove Management Systems
3.3. Identifying Indicator Plant Species in Olive Grove Management Systems
4. Discussion
4.1. The Contribution of Agricultural and Environmental Factors in the Yields of Organic and Conventional Olive Groves
4.2. The Role of Agricultural and Environmental Factors in the Yields of Organic and Conventional Olive Groves
4.2.1. Organic Olive Groves
4.2.2. Conventional Olive Groves
4.3. Typical Herbaceous Plant Species
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
| Variables | Sampling Methods | Organic Olive Groves | Conventional Olive Groves |
|---|---|---|---|
| Herbaceous plant cover (Hpc) (%) | Line point | 84.40 ± 5.96 | 73.80 ± 5.36 |
| Shannon’s diversity of herbaceous plants (Sd) | Line point | 2.63 ± 0.06 | 2,37 ± 0,04 |
| Herbaceous plant biomass (Hpb) (gr/0.25 m2) | 0.25 m2 plot | 69.40 ± 5.21 | 58.80 ± 4.39 |
| Woody plant density (Wpd) (individuals/100 m) | 10*10 m plot | 7.14 ± 0.42 | 4.25 ± 0.64 |
| Earthworm density (Ed) (individuals/0.25 m2) | 0.25 m2 plot | 25.24 ± 11.69 | 4.20 ± 1.54 |
| Isopod density (Id) (individuals/100 trap days) | Pitfall traps | 9.3 ± 2.1 | 6,86 ± 2.3 |
| Farm size (Fs) (ha) | GPS (Garmin eTrex Venture HC) | 138.30 ± 193.79 | 155.00 ± 109.29 |
| Altitude (Al)(m) | GPS (Garmin eTrex Venture HC) | 80.34 ± 51.71 | 62.85 ± 47.19 |
| Slope (Sl) (%) | Clinometer (Suunto Tandem) | 29.65 ± 21.05 | 23.96 ± 17.97 |
| Air temperature (At) (°C) | Digital Thermo-Hygrometer, TFA | 17.49 ± 1.77 | 18.66 ± 1.47 |
| Relative humidity (Rh) (%) | Digital Thermo-Hygrometer, TFA | 69.01 ± 8.65 | 63.85 ± 6.25 |
| Organic fertilizer K (OfK) (kg/m3) | Questionary | 81.00 ± 14.49 | |
| Inorganic fertilizer N (IfN)(kg/m3) | Questionary | 9.51 ± 0.74 | |
| Inorganic fertilizer Κ(IfK) (kg/m3) | Questionary | 20.64 ± 2.58 | |
| Manure (Man) (kg) | Questionary | 9.80 ± 0.42 | |
| * Herbicide (Her) | Questionary | 0 | 1 |
| Sand (San) (%) | Cylindrical sampler | 51.00 ± 12.57 | 55.80 ± 8.09 |
| Clay (Cl|) (%) | Cylindrical sampler | 23.60 ± 8.94 | 17.60 ± 5.96 |
| Silt (Sil) (%) | Cylindrical sampler | 25.40 ± 6.22 | 26.60 ± 6.46 |
| pH (pH) (%) | Cylindrical sampler | 7.01 ± 1.14 | 6.77 ± 0.75 |
| CEC (CEC) (meq/100 gr) | Cylindrical sampler | 18.50 ± 6.88 | 12.55 ± 4.23 |
| CaCO3 (CaCO3)(%) | Cylindrical sampler | 5.37 ± 9.43 | 2.02 ± 4.80 |
| P (P) (mg/kg) | Cylindrical sampler | 4.36 ± 1.80 | 2.75 ± 0.48 |
| K (K) (mg/kg) | Cylindrical sampler | 192.95 ± 186.45 | 108.15 ± 52.69 |
| Organic matter (Om)(%) | Cylindrical sampler | 3.46 ± 0.93 | 0.82 ± 0.16 |
| C/N (C/N) | Cylindrical sampler | 9.08 ± 3.36 | 4.83 ± 2.45 |
| Bulk density (Bd) (gr/cm3) | Cylindrical sampler | 0.99 ± 0.04 | 1.30 ± 0.09 |
| Species | Family | Organic Olive Groves | Conventional Olive Groves |
|---|---|---|---|
| Aegilops geniculata | Poaceae | + | + |
| Aira elegantissima | + | + | |
| Alopecurus myosuroides | + | + | |
| Briza maxima | + | + | |
| Bromus tectorum | + | ||
| Cynosurus echinatus | + | ||
| Dactylis glomerata | + | + | |
| Gaudinia fragilis | + | + | |
| Hordeum bulbosum | + | + | |
| Hordeum murinum | + | + | |
| Lagurus ovatus | + | ||
| Lolium perenne | + | ||
| Piptatherum miliaceum | + | + | |
| Psilurus incurvus | + | + | |
| Setaria verticillata | + | + | |
| Sorghum halepense | + | + | |
| Avena barbata | + | + | |
| Anthemis arvensis | Asteraceae | + | + |
| Anthemis chia | + | + | |
| Calendula arvensis | + | ||
| Carduus pycnocephalus | + | + | |
| Glebionis segetum | + | + | |
| Cichorium intybus | + | ||
| Crepis rubra | + | + | |
| Crupina crupinastrum | + | + | |
| Onopordum acanthium | + | ||
| Onopordum illyricum | + | ||
| Onopordum tauricum | + | ||
| Matricaria recutita | + | + | |
| Leontodon tuberosus | + | + | |
| Sonchus oleraceus | + | + | |
| Xanthium spinosum | + | ||
| Lupinus angustifolius | Fabaceae | + | + |
| Medicago lupulina | + | + | |
| Trifolium angustifolium | + | + | |
| Trifolium arvense | + | + | |
| Trifolium campestre | + | + | |
| Vicia cracca | + | ||
| Capsella bursa-pastoris | Brassicaceae | + | |
| Raphanus raphanistrum | + | + | |
| Rapistrum rugosum | + | ||
| Parietaria officinalis | + | ||
| Sinapis arvensis | + | + | |
| Alcea biennis | Malvaceae | + | + |
| Malva sylvestris | + | + | |
| Arisarum vulgare | Araceae | + | + |
| Arum maculatum | + | ||
| Dracunculus vulgaris | + | ||
| Anemone coronaria | Ranunculaceae | + | |
| Anemone pavonina | |||
| Asphodeline lutea | Asphodelaceae | + | |
| Asphodelus aestivus | + | + | |
| Asphodelus ramosus | + | ||
| Daucus carota | Apiaceae | + | + |
| Eryngium campestre | + | + | |
| Orlaya daucoides | + | + | |
| Orlaya grandiflora | + | ||
| Oenanthe pimpinelloides | + | ||
| Pallenis spinosa | + | + | |
| Smyrnium rotundifolium | + | ||
| Smyrnium perfoliatum | + | + | |
| Ferulago nodosa | + | ||
| Convolvulus althaeoides | Convolvulaceae | + | + |
| Convolvulus elegantissimus | + | + | |
| Fumaria officinalis | Papaveraceae | + | + |
| Papaver nigrotinctum | + | + | |
| Papaver rhoeas | + | + | |
| Agrostemma githago | Caryophylaceae | + | + |
| Stellaria media | + | + | |
| Silene cretica | + | + | |
| Amaranthus deflexus | Amaranthaceae | + | + |
| Anacamptis pyramidalis | Orchidaceae | + | + |
| Neottia nidus-avis | + | ||
| Anagallis arvensis | Primulaceae | + | + |
| Asterolinon linum-stellatum | + | + | |
| Cyclamen graecum | + | + | |
| Bellardia trixago | Orobanchaceae | + | + |
| Verbascum undulatum | + | ||
| Bituminaria bituminosa | Fabaceae | + | + |
| Onobrychis caput-galli | + | + | |
| Scorpiurus muricatus | + | + | |
| Campanula spatulata | Campanulaceae | + | + |
| Erodium cicutarium | Geraniaceae | + | + |
| Geranium robertianum | + | + | |
| Geranium tuberosum | + | ||
| Lilium candidum | Liliaceae | ||
| Muscari comosum | Hyacinthaceae | + | + |
| Narcissus tazetta | Amaryllidaceae | ||
| Tuberaria guttata | Cistaceae | + | + |
| Lamium amplexicaule | Lamiaceae | + | + |
| Phlomis fruticosa | + | ||
| Salvia verbenaca | + | + | |
| Salvia viridis | + | + | |
| Micromeria nervosa | + | ||
| Carex flacca | Cyperaceae | + | |
| Echium plantagineum | Boraginaceae | + | + |
| Knautia integrifolia | Dipsacaceae | + | + |
| Scabiosa stellata | + | ||
| Euphorbia helioscopia | Euphorbiaceae | + | + |
| Mercurialis annua | + | + | |
| Chenopodium album | Chenopodiaceae | + | + |
| Galium aparine | Rubiaceae | + | + |
| Gladiolus italicus | Iridaceae | + | + |
| Plantago major | Plantaginaceae | + | |
| Geum coccineum | Rosaceae | + | |
| Tribulus terrestris | Zygophyllaceae | + | |
| Urtica dioica | Urticaceae | + | + |
| Species | Family | Organic Olive Groves | Conventional Olive Groves |
|---|---|---|---|
| Arbutus andrachne | Ericaceae | ||
| Arbutus unedo | Ericaceae | ||
| Crataegus monogyna | Rosaceae | + | + |
| Calicotome villosa | Fabaceae | ||
| Cercis siliquastrum | Caesalpiniaceae | + | + |
| Cistus creticus | Cistaceae | ||
| Erica manipuliflora | Ericaceae | + | + |
| Ficus carica | Moraceae | ||
| Fumana thymifolia | Cistaceae | ||
| Juniperus oxycedrus | Cupressaceae | + | |
| Juniperus phoenicea | Cupressaceae | + | + |
| Myrtus communis | Myrtaceae | ||
| Olea europaea | Oleaceae | + | + |
| Olea. europaea var. sylvestris | Oleaceae | + | + |
| Paliurus spina-christi | Rhamnaceae | + | + |
| Phlomis fruticosa | Lamiaceae | + | + |
| Pistacia lentiscus | Anacardiaceae | + | + |
| Pistacia terebinthus | Anacardiaceae | ||
| Pyrus spinosa | Rosaceae | + | + |
| Quercus coccifera | Fagaceae | + | + |
| Quercus pubescens | Fagaceae | ||
| Rhamnusalaternus | Rhamnaceae | ||
| Rubus fruticosus | Rosaceae | + | + |
| Satureja thymbra | Lamiaceae | + | |
| Smilax aspera | Smilacaceae | + | |
| Spartium junceum | Fabaceae | + | |
| Ulmus glabra | Ulmaceae | + | |
| Vitex agnus-castus | Verbenaceae | + | + |
| Species | Family | Organic Olive Groves | Conventional Olive Groves |
|---|---|---|---|
| Armadillidium tuberculatum | Armadillidiidae | + | + |
| Armadillidium vulgare | Armadillidiidae | + | + |
| Armadillo officinalis | Armadillidae | + | + |
| Leptotrichus naupliensis | Porcellionidae | + | |
| Porcellio laevis | Porcellionidae | + | + |
| Porcellio obsoletus | Porcellionidae | + | |
| Porcellionides pruinosus | Porcellionidae | + | + |
| Species | Family | Organic Olive Groves | Conventional Olive Groves |
|---|---|---|---|
| Aporrectodea caliginosa | Lumbricidae | + | + |
| Aporrectodea trapezoides | Lumbricidae | + | |
| Dendrobaena byblica | Lumbricidae | + | + |
| Dendrobaena cognettii | Lumbricidae | + | |
| Dendrobaena veneta | Lumbricidae | + | |
| Microscolex dubius | Megascolecidae | + | + |
| Microscolex phosphoreus | Megascolecidae | + | |
| Octodrilus complanatus | Lumbricidae | + | + |
| Octodrilus croaticus | Lumbricidae | + | + |
References
- Pienkowski, M.; Beaufoy, G. The Environmental Impact of Olive Oil Production in the European Union: Practical Options for Improving the Environmental Impact; European Forum on Nature Conservation and Pastoralism: Peterborough, UK, 2002. [Google Scholar]
- Sokos, C.K.; Mamolos, A.P.; Kalburtji, K.L.; Birtsas, P.K. Review: Farming and wildlife in Mediterranean agroecosystems. J. Nat. Conserv. 2012, 21, 81–92. [Google Scholar] [CrossRef]
- Underwood, E.C.; Viers, J.H.; Klausmeyer, K.R.; Cox, R.L.; Shaw, M.R. Threats and biodiversity in the Mediterranean biome. Divers. Distrib. 2009, 15, 188–197. [Google Scholar] [CrossRef]
- Zhang, Z.; Zhou, J. From ecosystems to human welfare: The role and conservation of biodiversity Dos ecossistemas ao bem estar humano: O papel e a conservação da biodiversidade. Ciênc. Rural Santa Maria 2019, 49, e20170875. [Google Scholar] [CrossRef]
- Kaniewski, D.; Van Campo, E.; Boiy, T. Primary domestication and early uses of the emblematic olive tree: Palaeobotanical, historical and molecular evidence from the Middle East. Biol. Rev. 2012, 87, 885–899. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mercuri, A.M.; Mazzanti, M.B.; Florenzano, A. Olea, Juglans and Castanea: The OJC group as pollen evidence of the development of human-induced environments in the Italian peninsula. Quat. Int. 2013, 303, 24–42. [Google Scholar] [CrossRef]
- Valamoti, S.M.; Gkatzogia, E.; Ntinou, M. Did Greek colonisation bring olive growing to the north? An integrated archaeobotanical investigation of the spread of Olea europaea in Greece from the 7th to the 1st millennium BC. Veg. Hist. Archaeobot. 2018, 27, 177–195. [Google Scholar] [CrossRef]
- Zohary, D.; Hopf, M.; Weiss, E. Domestication of Plants in the Old World: The Origin and Spread of Domesticated Plants in Southwest Asia, Europe, and the Mediterranean Basin, 4th ed.; Oxford University Press: Oxford, UK, 2012; p. 296. [Google Scholar]
- Wingård, S. Olivolja Och Bordsoliver—Rapport; Jordbruksverket: Jönköping, Sweden, 2010; p. 78. [Google Scholar]
- International Olive Oil Council, IOOC. World Olive Oil Figures. Available online: http://www.internationaloliveoil.org/estaticos/view/131-world-olive-oil-figures (accessed on 22 September 2020).
- Melero, S.; Porras, J.C.R.; Herencia, J.F.; Madejon, E. Chemical and biochemical properties in a silty loam soil under conventional and organic management. Soil Till. Res. 2005, 6, 162–170. [Google Scholar] [CrossRef]
- Ministry of Rural Development and Food, 2013. Available online: http://www.minagric.gr (accessed on 14 August 2020).
- Soberȯn, J.; Rodriguez, P.; Vazquez-Dominguez, P. Implications of the hierarchical structure of biodiversity for the development of ecological indicators of sustainable use. Ambio 2000, 29, 136–142. [Google Scholar] [CrossRef]
- Petanidou, T.; Kizos, T. Socioeconomic Dimensions of Changes in the Agricultural Landscape of the Mediterranean Basin: A Case Study of the Abandonment of Cultivation Terraces on Nisyros Island, Greece. Environ. Manage. 2008, 41, 250–266. [Google Scholar] [CrossRef]
- Gaston, K. Global patterns in biodiversity. Nature 2000, 405, 220–227. [Google Scholar] [CrossRef]
- Dubois, A. The relationships between taxonomy and conservation biologyin the century of extinctions. C.R. Biol. 2003, 326, S9–S21. [Google Scholar] [CrossRef]
- Gao, J. N-terminal acetylation promotes synaptonemal complex assembly in C. elegans. Genes Dev. 2016, 30, 2404–2416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, S.Z. Growth regulator-induced betacyanin accumulation and dopa-4,5-dioxygenase (DODA) gene expression in euhalophyte Suaeda salsa calli. In Vitro Cell. Dev. Biol. Plant 2011, 47, 391–398. [Google Scholar] [CrossRef]
- Feng, Z.T. Effects of NaCl stress on the growth and photosynthetic characteristics of Ulmus pumila L. seedlings in sand culture. Photosynthetica 2014, 52, 313–320. [Google Scholar] [CrossRef]
- Zhou, J.J. Overexpression of OsPIL15, a phytochromeinteracting factor- like protein gene, represses etiolated seedling growth in rice. J. Integr. Plant Biol. 2014, 56, 373–387. [Google Scholar] [CrossRef]
- Yu, J. Concurrent highly pathogenic porcine reproductive and respiratory syndrome virus infection accelerates Haemophilus parasuis infection in conventional pigs. Vet. Microbiol. 2012, 158, 316–321. [Google Scholar] [CrossRef]
- Chen, T.S. Nitric oxide participates in waterlogging tolerance through enhanced adventitious root formation in the euhalophyte Suaeda salsa. Funct. Plant Biol. 2016, 43, 244–253. [Google Scholar] [CrossRef]
- Liu, W. Protein Kinase LTRPK1 Influences Cold Adaptation and Microtubule Stability in Rice. J. Plant Growth Regul. 2013, 32, 483–490. [Google Scholar] [CrossRef]
- Song, J. The role of the seed coat in adaptation of dimorphic seeds of the euhalophyte Suaeda salsa to salinity. Plant Species Biol. 2017, 32, 107–114. [Google Scholar] [CrossRef]
- Deng, Y.Q. Identification and functional analysis of the autofluorescent substance in Limonium bicolor salt glands. Plant Physiol. Biochem. 2015, 97, 20–27. [Google Scholar] [CrossRef]
- Fu, C. AtFes1A is Essential for Highly Efficient Molecular Chaperone Function in Arabidopsis. J. Plant Biol. 2015, 58, 366–373. [Google Scholar] [CrossRef]
- Zhang, Y.H. Reduced function of the RNA-binding protein FPA rescues a T-DNA insertion mutant in the Arabidopsis ZHOUPI gene by promoting transcriptional read-through. Plant Mol. Biol. 2016, 91, 549–561. [Google Scholar] [CrossRef] [PubMed]
- Yuan, C. Austdiol, fulvic acid and citromycetin derivatives from an endolichenic fungus, Myxotrichum sp. Phytochem. Lett. 2013, 6, 662–666. [Google Scholar] [CrossRef]
- Gao, J. NatB domain-containing CRA-1 antagonizes hydrolase ACER-1 linking acetyl-CoA metabolism to the initiation of recombination during C. elegans meiosis. PLoS Genet. 2015, 11, e1005029. [Google Scholar] [CrossRef] [Green Version]
- Xie, S.B.; Zhou, J. Harnessing Plant Biodiversity for the Discovery of Novel Anticancer Drugs Targeting Microtubules. Front. Plant Sci. 2017, 8, 720. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.Y. A Maternal Low-Fiber Diet Predisposes Offspring to Improved Metabolic Phenotypes in Adulthood in an Herbivorous Rodent. Physiol. Biochem. Zool. 2017, 90, 75–84. [Google Scholar] [CrossRef]
- Integrated Developmental Programme of Rural Areas (IDPRA) of Region of Thessaly, Greece. Case of Othrys, Magnesia; University of Thessaly: Volos, Greece, 2002.
- Pieper, R.P. Measurement Techniques for Herbaceous and Shrubby Vegetation; New Mexico State University: Las Cruses, NM, USA, 1978; pp. 1–148. [Google Scholar]
- Tutin, T.G.; Burges, N.A.; Chater, A.O.; Edmondson, J.R.; Heywood, V.H.; Moore, D.M.; Valentine, D.H.; Walters, S.M.; Webb, D.A. Flora Europaea; Cambridge University Press: Cambridge, UK, 1968. [Google Scholar]
- Tutin, T.G.; Heywood, V.H.; Burges, N.A.; Moore, D.M.; Valentine, D.H.; Walters, S.M.; Webb, D.A. Flora Europaea, 2nd ed.; Cambridge University Press: Cambridge, UK, 1993; Volume 1. [Google Scholar]
- Strid, A.; Tan, K. Flora Hellenica; Koeltz Scientific Books: Koenigstein, Germany, 2002. [Google Scholar]
- Dimopoulos, P.; Raus, T.; Bergmeier, E.; Constantinidis, T.; Iatrou, G.; Kokkini, S.; Strid, A.; Tzanoudakis, D. Vascular Plants of Greece: An Annotated Checklist; Botanic Garden and Botanical Museum Berlin-Dahlem: Berlin, Germany, 2013; p. 372. [Google Scholar]
- Dimopoulos, P.; Raus, T.; Bergmeier, E.; Constantinidis, T.; Iatrou, G.; Kokkini, S.; Strid, A.; Tzanoudakis, D. Vascular plants of Greece: An annotated checklist. Sup. Willdenowia 2016, 46, 301–347. [Google Scholar] [CrossRef] [Green Version]
- Zar, J. Biostatistical Analysis; Prentice-Hall, Inc.: Englewood Cliffs, NJ, USA, 1999; p. 960. [Google Scholar]
- McCullagh, P.; Nelder, J.A. Generalized linear models. In Standard Book on Generalized Linear Models; Chapman and Hall: London, UK, 1989; p. 512. [Google Scholar]
- Solomou, A.; Sfougaris, A.; Sfenthourakis, S. Terrestrial isopods as bioindicators for environmental monitoring in olive groves and natural ecosystems. J. Nat. Hist. 2019, 53, 1721–1735. [Google Scholar] [CrossRef]
- Pearce, J.; Ferrier, S. An evaluation of alternative algorithms for fitting species distribution models using logistic regression. Ecol. Modell. 2000, 128, 127–147. [Google Scholar] [CrossRef]
- Mittelbach, G.G.; Steiner, C.F.; Scheiner, S.M.; Gross, K.L.; Reynolds, H.L.; Waide, R.B.; Willig, M.R.; Dodson, S.I.; Gough, L. What is the observed relationship between species richness and productivity? Ecology 2001, 82, 2381–2396. [Google Scholar] [CrossRef]
- Crawley, M.J. GLIM for Ecologists; Blackwell Scientific: Oxford, UK, 1993; p. 379. [Google Scholar]
- Schwarz, M.; Zimmermann, N.E. A new GLM-based method for mapping tree cover continuous fields using regional MODIS reflectance data. Remote Sens. Environ. 2005, 95, 428–443. [Google Scholar] [CrossRef]
- Akaike, H. On entropy maximization principle. In Applications of Statistics; Krishnaiah, P.R., Ed.; North-Holland Publishing Company: Amsterdam, The Netherlands, 1977; pp. 27–41. [Google Scholar]
- Holm, S. A simple sequentially rejective multiple test procedure. Scand. J. Stat. 1979, 6, 65–70. [Google Scholar]
- Wohlgemuth, T.; Nobis, M.; Kienast, F.; Plattner, M. Modelling vascular plant diversity at the landscape scale using systematic samples. J. Biogeogr. 2008, 35, 1226–1240. [Google Scholar] [CrossRef]
- Team, R.C. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020; Available online: https://www.R-project.org (accessed on 23 November 2020).
- Dufrene, M.; Legendre, P. Species assemblages and indicator species: The need for a flexible asymmetrical approach. Ecol Monogr. 1997, 67, 345–366. [Google Scholar] [CrossRef]
- Dufrȇne, M. IndVal or How to Identify Indicator Species of a Sample Typology. Available online: http://mrw.wallonie.be/dgrne/sibw/outils/indval/home.htmlS (accessed on 10 January 2019).
- Seaby, R.M.; Henderson, P.A. Species Diversity and Richness, Version 4; Pisces Conservation Ltd.: Lymington, UK, 2006. [Google Scholar]
- Steve, D. Sustainable farming Compost Tea. 2009. Available online: http://www.soilsoup.com (accessed on 2 September 2020).
- Georgi, I.E. Effect of soil aluminum concentrations on bioaccumulation in earthworms of the genus Octodrilus. Postgraduate Thesis, Agricultural University of Athens, Department of Plant Production Science, Science and Modern Systems of Plant Production, Plant Protection and Architecture, Athens, Greece, 2010. [Google Scholar]
- Solomou, A.D.; Sfougaris, A.I.; Vavoulidou, E.M.; Csuzdi, C. The effects of farming practices on earthworm dynamics in olive groves of central Greece: (Oligochaeta). Zool. Middle East. 2012, 58, 119–126. [Google Scholar] [CrossRef]
- Solomou, A.D.; Sfougaris, A.I.; Vavoulidou, E.M.; Csuzdi, C. Species richness and density of earthworms in relation to soil factors in olive orchard production systems in Central Greece. Commun. Soil Sci. Plant Anal. 2013, 44, 301–311. [Google Scholar] [CrossRef]
- Briccoli Bati, C.; Santilli, E.; Guagliardi, I.; Toscano, P. Cultivation Techniques. 2012. Available online: http://creativecommons.org/licenses/by/3.0 (accessed on 4 July 2012).
- Gómez-Casero, M.T.; López-Granados, F.; Pena-Barragán, J.M.; Jurado-Expósito, M.; García-Torres, L. Assessing Nitrogen and Potassium Deficiencies in Olive Orchards through Discriminant Analysis of Hyperspectral Data. J. Amer. Soc. Hort. Sci. 2007, 132, 611–618. [Google Scholar] [CrossRef] [Green Version]
- Stirling, G.; Wilsey, B. Empirical Relationships between Species Richness, Evenness, and Proportional Diversity. Am. Nat. 2001, 158, 286–299. [Google Scholar] [CrossRef]
- Tuomisto, H.; Ruokolainen, K. Environmental and the diversity of Pteridophytes and Melastomataceae in western Amazonia. Biol. Skr. 2005, 55, 37–56. [Google Scholar]
- Sullivan, P.T.; Sullivan, S.D. Plant and small mammal diversity in orchard versus non-crop habitats. Agric. Ecosyst. Environ. 2006, 116, 235–243. [Google Scholar] [CrossRef]
- Solomou, A.; Sfougaris, A. Comparing conventional and organic olive groves in central Greece: Plant and bird diversity and abundance. Renew. Agr. Food Syst. 2011, 26, 297–316. [Google Scholar] [CrossRef]
- Gough, L.; Shaver, G.R.; Carroll, J.; Rozer, D.L.; Laundre, J.A. Vascular plant species richness in Alaskan arctic tundra: The importance of soil pH. J. Ecol. 2000, 88, 54–66. [Google Scholar] [CrossRef]
- Chen, Q.; Zhang, X.; Zhang, H.; Christie, P.; Li, X.; Horlacher, D.; Liebig, H.P. Evaluation of current fertilizer practice and soil fertility in vegetable production in the Beijing region. Nutr. Cycl. Agroecosys. 2004, 69, 51–58. [Google Scholar] [CrossRef] [Green Version]
- Akkuzu, E. Impact of cultural practices on arthropod abundance in soybean fields. Bulg. J. Agric. Sci. 2006, 12, 501–513. [Google Scholar]
- Sánchez, P.A.; Palm, C.A.; Szott, L.T.; Cuevas, E.; Lal, R. Organic input management in tropical agroecosystems. In Dynamics of Soil Organic Matter in Tropical Ecosystems; Coleman, D.C., Oades, J.M., Uehara, G., Eds.; University of Hawaii Press: Honolulu, HI, USA, 1989; pp. 125–152. [Google Scholar]
- Hoogmoed, W.B. Tillage for Soil and Water Conservation in the Semi-Arid Tropics; Wageningen University: Wageningen, The Netherlands, 1999. [Google Scholar]
- Pleasant, J.; Schlater, K.J. Incidence of weed seed in cow manure and its importance as a weed source in cropland. Weed Technol. 1994, 8, 304–310. [Google Scholar] [CrossRef]
- Yang, Y.H.; Chen, Y.N.; Li, W.H. Relationship Between Soil Properties and Plant Diversity in a Desert Riparian Forest in the Lower Reaches of the Tarim River, Xinjiang, China. Arid. Land Res. Manag. 2009, 23, 283–296. [Google Scholar] [CrossRef]
- Cook, A.R.; Posner, J.L.; Baldock, J.O. Effects of Dairy Manure and Weed Management on Weed Communities in Corn on Wisconsin Cash-grain Farms. Weed Technol. 2007, 21, 389–395. [Google Scholar] [CrossRef]
- Preston, F.W. The canonical distribution of commonness and rarity. Ecology 1962, 43, 185–215. [Google Scholar] [CrossRef]
- Schoener, T.W. Patterns in terrestrial vertebrate versus arthropod communities: Do systematic differences in regularity exist. In Community Ecology; Diamond, C.J., Case, T.S., Eds.; Harper and Pow: New York, NY, USA, 1986; pp. 556–586. [Google Scholar]
- Jacquemyn, H.; Brys, R.; Hermy, M. Patch occupancy, population size and reproductive success of a forest herb (Primula elatior) in a fragmented landscape. Oecologia 2002, 130, 617–625. [Google Scholar] [CrossRef]
- Bruun, H.H. A field test of the relationship between habitat area and population size for five perennial plant species. Web Ecol. 2005, 5, 1–5. [Google Scholar] [CrossRef]
- Belfrage, K.; Bjorklund, J.; Salomonsson, L. The effects of farm size and organic farming on diversity of birds, pollinators, and plants in a Swedish landscape. Ambio 2005, 34, 582–588. [Google Scholar] [CrossRef] [PubMed]
- Marini, L.; Fontana, P.; Battisti, A.; Gaston, K.J. Agricultural management, vegetation traits and landscape drive orthopteran and butterfly diversity in a grassland–forest mosaic: A multi-scale approach. Insect Conserv. Divers. 2009, 2, 213–220. [Google Scholar] [CrossRef]
- Panagopoulou, A. Biogeochemical Study of the Origanum majorana plant aiming at the Protection of Public Health. Master’s Thesis, Department of Geology, University of Patras, Patras, Greece, 2011. [Google Scholar]
- Al-Mufti, M.M.; Sydes, C.L.; Furness, S.B.; Grime, J.P.; Band, S.R. A quantitative analysis of shoot phenology and dominance in herbaceous vegetation. J. Ecol. 1977, 65, 759–791. [Google Scholar] [CrossRef]
- Grime, J.P. Plant Strategies and Vegetation Processes; John Wiley and Son: New York, NY, USA, 1979; p. 222. [Google Scholar]
- Wheeler, B.D.; Shaw, S.C. Above-ground crop mass and species richness of the principal types of herbaceous rich-fen vegetation of lowland England and Wales. J. Ecol. 1991, 79, 285–301. [Google Scholar] [CrossRef]
- Grace, J.B. The factors controlling species density in herbaceous plant communities: An assessment. Perspect. Plant Ecol. Evol. Syst. 1999, 2, 1–28. [Google Scholar] [CrossRef]
- Zechmeister, H.G.; Schmitzberger, I.; Steurer, B.; Peterseil, J.; Wrbka, T. The influence of land-use practices and economics on plant species richness in meadows. Biol. Conserv. 2003, 114, 165–177. [Google Scholar] [CrossRef]
- Fattahi, B.; Reza Ildoromi, A. Effect of Some Environmental Factors on Plant Species Diversity in the Mountainous Grasslands (Case Study: Hamedan—Iran). Ecopersia 2011, 1, 45–52. [Google Scholar]
- Willems, J.H.; Peet, R.K.; Bik, L. Changes in chalk grassland structure and species richness resulting from selective nutrient additions. J. Veg. Sci. 1993, 4, 203–212. [Google Scholar] [CrossRef]
- Kirkham, F.W.; Mountford, J.O.; Wilkins, R.J. The effects of nitrogen, potassium and phosphorus addition on the vegetation of a Somerset peat moor under cutting management. J. Appl. Ecol. 1996, 33, 1013–1029. [Google Scholar] [CrossRef]
- Ellenberg, H. Indicator Values of Vascular Plants in Central Europe. Scripta Geobotanica 1974, 9, 1–97. [Google Scholar]
- Böhling, N.; Greuter, W.; Raus, T. Indicator values of the vascular plants in the Southern Aegean Greece. Braun Blanquetia 2002, 32, 108. [Google Scholar]
- Vasilakoglou, I. Weeds: Recognition and Management; Stamouli Publications SA: Athens, Greece, 2004; p. 303. [Google Scholar]
- Eleftherochorinos, E.G.; Giannopolitis, K.N. Weeds: Identification Guide; Agrotypos Publications: Athens, Greece, 2009; p. 270. [Google Scholar]
- Schmalfuss, H. World catalog of terrestrial isopods (Isopoda: Oniscidea). Stuttg. Beitr. Naturkd. A. 2003, 654, 1–341. [Google Scholar]
- Schmalfuss, H. The terrestrial isopods (Isopoda: Oniscidea) of Greece. 28th contribution: The genus Armadillidium (Armadillidiidae) on the central Greek mainland. Stuttg. Beitr. Naturkd. A.N. Ser. 2008, 5, 73–101. [Google Scholar]
- Graf, O. Die Regenwürmer Deutschlands. Schriftreihe der Forschungsanstalt für Landwirtschaft Braunschweig Volkenrode. Heft 7; Verlag, M.u. H Schaper: Hannover, Germany, 1955. [Google Scholar]
- Zicsi, A. Über die Regenwürmer Ungarns mit Bestimmungstabellen der Arten. Opusc. Zool. Budapest 1991, 24, 167–191. [Google Scholar]
- Christian, E.; Zicsi, A. Ein synoptischer Bestimmungsschlüssel der Regenwürmer Österreichs (Oligochaeta: Lumbricidae). Die Bodenkultur. Aust. J. Agric. Res. 1999, 50, 121–131. [Google Scholar]

| Organic | Conventional | |
|---|---|---|
| Average field size (ha) | 13.83 | 15.5 |
| Average number of olive trees per hectare | 200 | 200 |
| Age of olive groves (years) | ~150–170 | ~150–170 |
| Years of enrolment | 1997 | |
| Average olive production (kg/tree) | 48.20 | 51.00 |
| Manure (kg per tree) | 50 | |
| Inorganic fertilizer N (kg per tree) | 1.5–2 | |
| Inorganic fertilizer K (kg per tree) | 1.5–2 | |
| Organic fertilizer K (kg per tree)Weed control | 2–3Grass cutting | Herbicide |
| Irrigation application | No | No |
| Organic Olive Groves | ||||
|---|---|---|---|---|
| Variable | ||||
| Names or selection procedure | AIC | Residual deviance | D2 | Percentage change in D2 |
| Ed | 33.900 | 4.950 | 0.605 | ̶ |
| Ed + Man | 27.949 | 4.300 | 0.610 | 0.800 |
| All variables | 30.220 | 3.270 | 0.620 | ̶ |
| Conventional Olive Groves | ||||
| Variable | ||||
| Names or selection procedure | AIC | Residual deviance | D2 | Percentage change in D2 |
| Om | 50.008 | 7.726 | 0.685 | ̶ |
| Om + IfN | 25.548 | 7.167 | 0.690 | 0.875 |
| All variables | 40.854 | 6.894 | 0.725 | ̶ |
| Organic Olive Groves | ||||||||
|---|---|---|---|---|---|---|---|---|
| Robust | ||||||||
| Variables (X1, X2) | Coef. (Estimate) | SE | t | Adj.R2 | Likelihood Ratio χ2 | BIC | Wald Chi-Square | F |
| 0.889 | 24.524 | 29.159 | 37.157 | |||||
| (Intercept) | 14.675 | 4.933 | 2.974 * | 8.853 | ||||
| Ed | 0.034 | 0.009 | 3.777 * | 13.473 | ||||
| Man | 2.787 | 0.623 | 4.473 * | 19.970 | ||||
| Conventional Olive Groves | ||||||||
| Robust | ||||||||
| Variables (X1, X2) | Coef. (Estimate) | SE | t | Adj.R2 | Likelihood Ratio χ2 | BIC | Wald Chi-Square | F |
| 0.850 | 749.533 | 26.456 | 374.766 | |||||
| (Intercept) | 1.428 | 1.796 | 0.795 * | 0.632 | ||||
| Om | 1.857 | 0.818 | 2.270 * | 5.142 | ||||
| IfN | 4.674 | 0.236 | 19.805 * | 390.523 | ||||
| Organic Olive Groves | ||||
|---|---|---|---|---|
| Variables | ||||
| Names or selection procedure | AIC | Residual deviance | D2 | Percentage change in D2 |
| Sd | 48.840 | 3.530 | 0.750 | − |
| Sd + OrgK | 47.550 | 3.400 | 0.770 | 2.666 |
| Sd + OrgK + Man | 45.230 | 3.330 | 0.778 | 1.038 |
| Sd + OrgK + Man + Fs | 20.115 | 3.110 | 0.785 | 0.899 |
| All variables | 42.000 | 3.090 | 0.791 | − |
| Conventional Olive Groves | ||||
| Variables | ||||
| Names or selection procedure | AIC | Residual deviance | D2 | Percentage change in D2 |
| IfN | 56.020 | 5.330 | 0.533 | − |
| All variables | 54.000 | 4.100 | 0.635 | |
| Organic Olive Groves | ||||||||
|---|---|---|---|---|---|---|---|---|
| Robust | ||||||||
| Variables (X1, X2…) | Coef. (Estimate) | SE | t | Adj.R2 | Likelihood Ratio χ2 | BIC | Wald Chi-Square | F |
| 0.985 | 54.600 | 21.931 | 292.621 | |||||
| (Intercept) | 20.692 | 1.076 | 19.23 * | 369.538 | ||||
| Sd | 5.808 | 0.426 | 13.633 * | 185.533 | ||||
| OrgK | 0.109 | 0.02 | 5.45 * | 14.353 | ||||
| Man | 2.231 | 0.367 | 6.07 * | 36.874 | ||||
| Fs | 1.912 | 0.116 | 16.482 * | 267.427 | ||||
| Conventional Olive Groves | ||||||||
| Robust | ||||||||
| Variables (X1, X2…) | Coef. (Estimate) | SE | t | Adj.R2 | Likelihood Ratio χ2 | BIC | Wald Chi-Square | F |
| 0.456 | 7.268 | 56.928 | 8.548 | |||||
| (Intercept) | 57.703 | 17.800 | 3.241 * | 10.509 | ||||
| IfN | 4.301 | 1.778 | 2.419 * | 5.484 | ||||
| Species | IndVal (%) | Management System |
|---|---|---|
| Leontodon tuberosus | 97.5 | Olive grove management systems |
| Muscari comosum | 97.5 | Olive grove management systems |
| Ferulago nodosa | 92.5 | Olive grove management systems |
| Salvia verbenaca | 90 | Olive grove management systems |
| Raphanus raphanistrum | 87.5 | Olive grove management systems |
| Fumaria officinalis | 85 | Olive grove management systems |
| Cyclamen graecum | 75 | Olive grove management systems |
| Erodium cicutarium | 72.5 | Olive grove management systems |
| Anthemis arvensis | 70 | Olive grove management systems |
| Aegilops ovata | 67.5 | Olive grove management systems |
| Geranium robertianum | 67.5 | Olive grove management systems |
| Avena barbata | 60 | Olive grove management systems |
| Convolvulus althaeoides | 57.5 | Olive grove management systems |
| Pallenis spinosa | 57.5 | Olive grove management systems |
| Setaria verticillata | 73.77 | Organic olive groves |
| Medicago lupulina | 77.87 | Organic olive groves |
| Trifolium arvense | 77.67 | Organic olive groves |
| Hordeum bulbosum | 63.83 | Organic olive groves |
| Malva sylvestris | 74.84 | Organic olive groves |
| Papaver rhoeas | 57.55 | Organic olive groves |
| Trifolium campestre | 56 | Organic olive groves |
| Anagallis arvensis | 55.46 | Organic olive groves |
| Matricaria recutita | 89.41 | Organic olive groves |
| Sinapis arvensis | 51 | Organic olive groves |
| Sonchus oleraceous | 72.61 | Conventional olive groves |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Solomou, A.D.; Sfougaris, A. Contribution of Agro-Environmental Factors to Yield and Plant Diversity of Olive Grove Ecosystems (Olea europaea L.) in the Mediterranean Landscape. Agronomy 2021, 11, 161. https://doi.org/10.3390/agronomy11010161
Solomou AD, Sfougaris A. Contribution of Agro-Environmental Factors to Yield and Plant Diversity of Olive Grove Ecosystems (Olea europaea L.) in the Mediterranean Landscape. Agronomy. 2021; 11(1):161. https://doi.org/10.3390/agronomy11010161
Chicago/Turabian StyleSolomou, Alexandra D., and Athanassios Sfougaris. 2021. "Contribution of Agro-Environmental Factors to Yield and Plant Diversity of Olive Grove Ecosystems (Olea europaea L.) in the Mediterranean Landscape" Agronomy 11, no. 1: 161. https://doi.org/10.3390/agronomy11010161
APA StyleSolomou, A. D., & Sfougaris, A. (2021). Contribution of Agro-Environmental Factors to Yield and Plant Diversity of Olive Grove Ecosystems (Olea europaea L.) in the Mediterranean Landscape. Agronomy, 11(1), 161. https://doi.org/10.3390/agronomy11010161