Implications of the Symbiotic Nitrogen Fixation in Common Bean under Seasonal Water Stress


Abstract
:1. Introduction
2. Materials and Methods
3. Results and Discussion
3.1. Greenhouse Experiment
3.2. Field Experiment
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Duke, J.A. Handbook of Legumes of World Economic Importance; Plenum Press: New York, NY, USA, 1981. [Google Scholar]
- Wondatir, Z.; Mekasha, Y. Feed resources availability and livestock production in the central rift valley of Ethiopia. Int. J. Livest. Prod. 2014, 5, 30–35. [Google Scholar] [CrossRef] [Green Version]
- Beebe, S.E.; Rao, I.M.; Blair, M.W.; Acosta-Gallegos, J.A. Phenotyping common beans for adaptation to drought. Front. Physiol. 2013, 4, 1–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sánchez-Reinoso, A.D.; Ligarreto-Moreno, G.A.; Restrepo-Díaz, H. Evaluation of drought índices to identify tolerant geno-types in common bean bush (Phaseolus vulgaris, L.). J. Integr. Agric. 2020, 19, 99–107. [Google Scholar] [CrossRef]
- Asfaw, A.; Blair, M.W. Quantification of drought tolerance in Ethiopian common bean varieties. Agric. Sci. 2014, 5, 124–139. [Google Scholar] [CrossRef] [Green Version]
- White, J.W.; Singh, S.P. Breeding for adaptation to drought. In Common Beans: Research for Crop Improvement; Van Shoonhoven, A., Voysest, O., Eds.; CABI: Wallingford, UK; CIAT: Cali, Colombia, 1991; pp. 501–560. [Google Scholar]
- Cuellar-Ortiz, S.M.; Arrieta-Montiel, M.D.L.P.; Acosta-Gallegos, J.; Covarrubias, A.A. Relationship between carbohydrate partitioning and drought resistance in common bean. Plant Cell Environ. 2008, 31, 1399–1409. [Google Scholar] [CrossRef] [PubMed]
- Asfaw, A.; Demissie, D.A.; Shah, T.; Blair, M. Trait associations in diversity panels of the two common bean (Phaseolus vulgaris L.) gene pools grown under well-watered and water-stress conditions. Front. Plant Sci. 2017, 8, 733. [Google Scholar] [CrossRef] [PubMed]
- Miklas, P.N.; Kelly, J.D.; Beebe, S.E.; Blair, M.W. Common bean breeding for resistance against biotic and abiotic stresses: From classical to MAS breeding. Euphytica 2006, 147, 105–131. [Google Scholar] [CrossRef]
- Muñoz-Perea, C.G.; Terán, H.; Allen, R.G.; Wright, J.L.; Westermann, D.T.; Singh, S.P. Selection for Drought Resistance in Dry Bean Landraces and Cultivars. Crop. Sci. 2006, 46, 2111–2120. [Google Scholar] [CrossRef] [Green Version]
- Nielsen, D.C.; Nelson, N.O. Black Bean Sensitivity to Water Stress at Various Growth Stages. Crop Sci. 1998, 38, 422–427. [Google Scholar] [CrossRef]
- Yonts, C.D. Deficit Irrigation of Dry Edible Beans during Early, Mid and Late Season. In Proceedings of the 5th National Decennial Irrigation Conference Proceedings, Phoenix, AZ, USA, 5–8 December 2010; American Society of Agricultural and Biological Engineers (ASABE): St. Joseph, MI, USA, 2010; p. 1. [Google Scholar]
- Broughton, W.J.; Zhang, F.; Perret, X.; Staehelin, C. Signals exchanged between legumes and Rhizobium: Agricultural uses and perspectives. Plant Soil 2003, 252, 129–137. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Romero, E. Diversity of Rhizobium-Phaseolus vulgaris symbiosis: Overview and perspectives. Plant Soil 2003, 252, 11–23. [Google Scholar] [CrossRef]
- Kuykendall, L.D. Family I Rhizobiaceae. In Bergey’s Manual of Systematic Bacteriology, 2rd ed.; Garrity, G., Krieg, N.R., Holt, J.G., Eds.; Springer: New York, NY, USA, 2005; pp. 324–340. ISBN 9780387241456. [Google Scholar]
- Herrera-Cervera, J.A.; Caballero-Mellado, J.; Laguerre, G.; Tichy, H.-V.; Requena, N.; Amarger, N.; Martínez-Romero, E.; Olivares, J.; Sanjuán, J. At least five rhizobial species nodulate Phaseolus vulgaris in a Spanish soil. FEMS Microbiol. Ecol. 1999, 30, 87–97. [Google Scholar] [CrossRef]
- Drevon, J.J.; Alkama, N.; Bargaz, A.; Rodiño, A.P.; Sungthongwises, K.; Zaman-Allah, M. The Legume–Rhizobia Symbiosis. In Grain Legumes; De Ron, A.M., Ed.; Springer Science + Business Media: New York, NY, USA, 2015. [Google Scholar]
- Kaymakanova, M.; Mincheva, T. Salinity and its effects on the physiological response of bean (Phaseolus vulgaris L.). J. Cent. Eur. Agric. 2008, 9, 749–755. [Google Scholar]
- Wilmowicz, E.; Kucko, A.; Golinska, P.; Burchardt, S.; Przywieczerski, T.; Świdziński, M.; Brzozowska, P.; Kapuścińska, D. Abscisic acid and ethylene in the control of nodule-specific response on drought in yellow lupine. Environ. Exp. Bot. 2020, 169, 103900. [Google Scholar] [CrossRef]
- Bouhmouch, I.; Souad-Mouhsine, B.; Brhada, F.; Aurag, J. Influence of host cultivars and Rhizobium species on the growth and symbiotic performance of Phaseolus vulgaris under water stress. J. Plant Physiol. 2005, 162, 1103–1113. [Google Scholar] [CrossRef]
- Chemining, G.N.; Vessey, J.K. The abundance and efficacy of Rhizobium leguminosarum bv. viciae in cultivated soils of the eastern Canadian prairie. Soil Biol. Biochem. 2006, 38, 294–302. [Google Scholar]
- SAS Institute Inc. SAS/STAT 15.1; SAS Institute Inc.: Cary, NC, USA, 2018. [Google Scholar]
- Steel, R.G.D.; Torrie, J.H.; Dickey, D.A. Principle and Procedures of Statistics: A Biometrical Approach, 3rd ed.; Mc Graw Hill: New York, NY, USA, 1997; p. 666. ISBN 0070610282. [Google Scholar]
- Hungria, M.; Franchini, J.C.; Campo, R.J.; Crispino, C.C.; Moraes, J.Z.; Sibaldelli, R.N.; Mendes, I.C.; Arihara, J. Nitrogen nutrition of soybean in Brazil: Contributions of biological N2 fixation and N fertilizer to grain yield. Can. J. Plant Sci. 2006, 86, 927–939. [Google Scholar] [CrossRef] [Green Version]
- Rodiño, A.P.; De La Fuente, M.; De Ron, A.M.; Lema, M.J.; Drevon, J.-J.; Santalla, M. Variation for nodulation and plant yield of common bean genotypes and environmental effects on the genotype expression. Plant Soil 2011, 346, 349–361. [Google Scholar] [CrossRef]
- Van Kessel, C.; Hartley, C. Agricultural management of grain legumes: Has it led to an increase in nitrogen fixation? Field Crops Res. 2000, 65, 165–181. [Google Scholar] [CrossRef]
- Rengel, Z. Breeding for better symbiosis. Plant Soil 2002, 245, 147–162. [Google Scholar] [CrossRef]
- Dorcinvil, R.; Sotomayor-Ramírez, D.; Beaver, J. Agronomic performance of common bean (Phaseolus vulgaris L.) lines in an Oxisol. Field Crops Res. 2010, 118, 264–272. [Google Scholar] [CrossRef]
- Mostasso, L.; Mostasso, F.L.; Dias, B.G.; Vargas, M.A.; Hungria, M. Selection of bean (Phaseolus vulgaris L.) rhizobial strains for the Brazilian Cerrados. Field Crops Res. 2002, 73, 121–132. [Google Scholar] [CrossRef]
- Oka-Kira, E.; Kawaguchi, M. Long-distance signaling to control root nodule number. Curr. Opin. Plant Biol. 2006, 9, 496–502. [Google Scholar] [CrossRef] [PubMed]
- Bourion, V.; Laguerre, G.; Depret, G.; Voisin, A.S.; Salon, C.; Duc, G. Genetic variability in nodulation and root growth affects nitrogen fixation and accumulation in pea. Ann. Bot. 2007, 100, 589–598. [Google Scholar] [CrossRef] [PubMed]
- Schultze, M.; Kondorosi, A. Regulation of symbiotic root nodule development. Annu. Rev. Genet. 1998, 32, 33–57. [Google Scholar] [CrossRef] [PubMed]
- Sinclair, T.R.; Purcell, L.C.; King, C.A.; Sneller, C.H.; Chen, P.; Vadez, V. Drought tolerance and yield increase of soybean resulting from improved symbiotic N2 fixation. Field Crops Res. 2007, 101, 68–71. [Google Scholar] [CrossRef] [Green Version]
- Mahieu, S.; Germon, F.; Aveline, A.; Hauggaard-Nielsen, H.; Ambus, P.; Jense, E.S. The influence of water stress on biomass and N accumulation, N partitioning between above and below ground parts and on N rhizodeposition during reproductive growth of pea (Pisum sativum L.). Soil Biol. Biochem. 2008, 41, 380–387. [Google Scholar] [CrossRef]
- Voisin, A.S.; Munier-Jolain, G.; Salon, C. The nodulation process is tightly adjusted to plant growth an analysis using environmentally and genetically induced variation of nodule. Plant Soil 2010, 337, 399–412. [Google Scholar] [CrossRef]
- Voisin, A.-S.; Bourion, V.; Duc, G.; Salon, C. Using an Ecophysiological Analysis to Dissect Genetic Variability and to Propose an Ideotype for Nitrogen Nutrition in Pea. Ann. Bot. 2007, 100, 1525–1536. [Google Scholar] [CrossRef] [Green Version]
- Graham, P.H. Ecology of the Root-Nodule Bacteria of Legumes. In Nitrogen-Fixing Leguminous Symbioses; Springer: Berlin/Heidelberg, Germany, 2008; pp. 23–58. [Google Scholar] [CrossRef]
- Muchero, W.; Ehlers, J.; Close, T.J.; Roberts, P.A. Mapping QTL for drought stress-induced premature senescence and ma-turity in cowpea. Theor. Appl. Genet. 2009, 118, 849–863. [Google Scholar] [CrossRef]
- Muchero, W.; Ehlers, J.D.; Roberts, P.A. Restriction site polymorphism-based candidate gene mapping for seedling drought tolerance in cowpea [Vigna unguiculata (L.) Walp.]. Theor. Appl. Genet. 2010, 120, 509–518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mergaert, P.; Uchiumi, T.; Alunni, B.; Evanno, G.; Cheron, A.; Catrice, O.; Mausset, A.-E.; Barloy-Hubler, F.; Galibert, F.; Kondorosi, A.; et al. Eukaryotic control on bacterial cell cycle and differentiation in the Rhizobium-legume symbiosis. Proc. Natl. Acad. Sci. USA 2006, 103, 5230–5235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, S.O.; Coyne, D.P.; Jung, G.; Skroch, P.W.; Arnaud-Santana, E.; Steadman, J.R.; Ariyarathne, H.; Nienhuis, J. Mapping of QTL for Seed Size and Shape Traits in Common Bean. J. Am. Soc. Hortic. Sci. 2000, 125, 466–475. [Google Scholar] [CrossRef] [Green Version]
- González, A.; Rodiño, A.; Santalla, M.; De Ron, A.M. Genetics of intra-gene pool and inter-gene pool hybridization for seed traits in common bean (Phaseolus vulgaris L.) germplasm from Europe. Field Crops Res. 2009, 112, 66–76. [Google Scholar] [CrossRef] [Green Version]
- Michiels, J.; Dombrecht, B.; Vermeiren, N.; Xi, C.; Luyten, E.; Vanderleyden, J. Phaseolus vulgaris is a non-selective host for nodulation. FEMS Microbiol. Ecol. 1998, 26, 193–205. [Google Scholar] [CrossRef]
- Graham, P. Some problems of nodulation and symbiotic nitrogen fixation in Phaseolus vulgaris L.: A review. Field Crops Res. 1981, 4, 93–112. [Google Scholar] [CrossRef]
- Romdhane, S.B.; Aouani, M.E.; Trabelsi, M.; de Lajudie, P.; Mhamdi, R. Selection of high nitrogen-fixing Rhizobia modulating chickpea (Cicer arietinum) for Semi-Arid Tunisia. J. Agron. Crop Sci. 2008, 194, 413–420. [Google Scholar]
- Hungria, M.; Campo, R.J.; Mendes, I.C. Benefits of inoculation of the common bean (Phaseolus vulgaris) crop with efficient and competitive Rhizobium tropici strains. Biol. Fertil. Soils 2003, 39, 88–93. [Google Scholar] [CrossRef]
- Rennie, R.J.; Kemp, G.A. N2-Fixation in Field Beans Quantified by 15N Isotope Dilution. II. Effect of Cultivars of Beans 1. Agron. J. 1983, 75, 645–649. [Google Scholar] [CrossRef]
- Vessey, J.K. Cultivar differences in assimilate partitioning and capacity to maintain N2 fixation rate in pea during pod-filling. Plant Soil 1992, 139, 185–194. [Google Scholar] [CrossRef]
- Araya, T.; Noguchi, K.; Terashima, I. Effect of nitrogen nutrition on the carbohydrate repression of photosynthesis in leaves of Phaseolus vulgaris L. J. Plant Res. 2009, 123, 371–379. [Google Scholar] [CrossRef] [PubMed]
- Hardarson, G.; Atkins, G. Optimising biological N2 fixation by legumes in farming systems. Plant Soil 2003, 252, 41–54. [Google Scholar] [CrossRef]
- Mutschler, M.A.; Bliss, F.A. Inheritance of bean seed globulin content and its relationship to protein content on quality. Crop Sci. 1981, 21, 289–294. [Google Scholar] [CrossRef]
- Escribano, M.; Santalla, M.; De Ron, A.M. Genetic diversity in pod and seed quality traits of common bean populations from northwestern Spain. Euphytica 1997, 93, 71–81. [Google Scholar] [CrossRef]
- Vasilas, B.L.; Nelson, R.L.; Fuhrmann, J.J.; Evans, T.A. Relationship of Nitrogen Utilization Patterns with Soybean Yield and Seed-Fill Period. Crop Sci. 1995, 35, 809–813. [Google Scholar] [CrossRef]
- Kurdali, F.; Kalifa, K.; Al-Shamma, M. Cultivar differences in nitrogen assimilation, partitioning and mobilization in rain-fed grown lentil. Field Crops Res. 1997, 54, 235–243. [Google Scholar] [CrossRef]
- Naya, L.; Ladrea, R.; Ramos, J.; González, E.M.; Arrese-Igor, C.; Minchin, F.R.; Becana, M. The response of carbon metabolism and antioxidant defenses of alfalfa nodules to drought stress and to the subsequent recovery of plants. Plant Physiol. 2007, 144, 1104–1114. [Google Scholar] [CrossRef] [Green Version]
- Singh, S.P. Drought Resistance in the Race Durango Dry Bean Landraces and Cultivars. Agron. J. 2007, 99, 1219–1225. [Google Scholar] [CrossRef]
- Sadras, V.O. Evolutionary aspects of the trade-off between seed size and number in crops. Field Crops Res. 2007, 100, 125–138. [Google Scholar] [CrossRef]
- Harper, L.A.; Giddens, J.E.; Angdale, G.W.; Sharpe, R.R. Environmental effects on nitrogen dynamics in soybean under conservation and clean tillage systems. Agron. J. 1989, 81, 623–631. [Google Scholar] [CrossRef]
- Boutraa, T.; Sanders, F.E. Effects of interactions of moisture regime and nutrient addition on nodulation and carbon partitioning in two cultivars of bean (Phaseolus vulgaris L.). J. Agron. Crop Sci. 2001, 186, 229–237. [Google Scholar] [CrossRef]
- Silva, A.d.N.; Gerosa, M.L.; Ribeiro, W.Q.; Rodrigues, E.; Carvalhoda, P.; Andreade, C.; Cleo, C.; Vanderlei, M.A. Water stress alters physical and chemical quality in grains of common bean, triticale and wheat. Agric. Water Manag. 2020, 231, 106023. [Google Scholar] [CrossRef]
- Hungria, M.; Vargas, M.A. Environmental factors affecting N2 fixation in grain legumes in the tropics, with an emphasis on Brazil. Field Crops Res. 2000, 65, 151–164. [Google Scholar] [CrossRef]
- Mnasri, B.; Aouani, M.E.; Mhamdi, R. Nodulation and growth of common bean (Phaseolus vulgaris) under water deficiency. Soil Biol. Biochem. 2007, 39, 1744–1750. [Google Scholar] [CrossRef]
- Güereña, D.T.; Lehmann, J.; Thies, J.E.; Vanek, S.; Karanja, N.; Neufeldt, H. Nodulation of beans with inoculant carriers from pyrolyzed and non-pyrolyzed sugarcane bagasse in response to different pre-planting water availability. Appl. Soil Ecol. 2019, 143, 126–133. [Google Scholar] [CrossRef]
- Kipe-Nolt, J.A.; Vargas, H.; Giller, K.E. Nitrogen fixation in breeding lines of Phaseolus vulgaris L. Plant Soil 1993, 152, 103–106. [Google Scholar] [CrossRef]
- Sinclair, T.R.; Muchow, R.C.; Ludlow, M.M.; Leach, G.J.; Lawn, R.J.; Foale, M.A. Field and model analysis of the effect of water deficits on carbon and nitrogen accumulation by soybean, cowpea and black gram. Field Crops Res. 1987, 17, 121–140. [Google Scholar] [CrossRef]
- Pfeiffer, N.E.; Malik, N.S.A.; Wagner, F.W.; Wang, C.Y.; Adams, D.O. Reversible Dark-Induced Senescence of Soybean Root Nodules. Plant Physiol. 1983, 71, 393–399. [Google Scholar] [CrossRef]
- Kumarasinghe, K.S.; Danso, S.K.A.; Zapata, F. Field evaluation of fixation and N partitioning in climbing bean (Phaseolus vulgaris L.) using 15N. Biol. Fertil. Soils 1992, 13, 142–146. [Google Scholar]
- Ramos, M.L.G.; Gordon, A.J.; Minchin, F.R.; Sprent, J.I.; Parsons, R. Effect of water stress on nodule physiology and bio-chemistry of a drought tolerant cultivar of common bean (Phaseolus vulgaris L.). Ann. Bot. 1999, 83, 57–63. [Google Scholar] [CrossRef] [Green Version]
- Gálvez, L.; González, E.M.; Arrese-Igor, C. Evidence for carbon flux shortage and strong carbon/nitrogen interactions in pea nodules at early stages of water stress. J. Exp. Bot. 2005, 56, 2551–2561. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peña-Cabriales, J.J.; Castellanos, J.Z. Effects of water stress on N2 fixation and grain yield of Phaseolus vulgaris L. In Enhancement of Biological Nitrogen Fixation of Common Bean in Latin America; Springer: Dordrecht, The Netherlands, 1993; Volume 52, pp. 151–155. [Google Scholar] [CrossRef]
- Araújo, A.P.; Teixeira, M.G. Nitrogen and phosphorus harvest indices of common bean cultivars: Implications for yield quantity and quality. Plant Soil 2003, 257, 425–433. [Google Scholar] [CrossRef]
- Vadez, V.; Drevon, J.J. Genotypic variability in phosphorus use efficiency for symbiotic N2 fixation in common bean (Phaseolus vulgaris). Agronomie 2001, 21, 691–699. [Google Scholar] [CrossRef] [Green Version]
- Remans, R.; Croonenborghs, A.; Gutierrez, R.T.; Michiels, J.; Vanderleyden, J. Effects of plant growth-promoting rhizobacteria on nodulation of Phaseolus vulgaris L. are dependent on plant P nutrition. Eur. J. Plant Pathol. 2007, 119, 341–351. [Google Scholar] [CrossRef]
- Wakelin, S.A.; Macdonald, L.; Rogers, S.; Gregg, A.; Bolger, T.; Baldock, J. Habitat selective factors influencing the structural composition and functional capacity of microbial communities in agricultural soils. Soil Biol. Biochem. 2008, 40, 803–813. [Google Scholar] [CrossRef]





{kind=link}
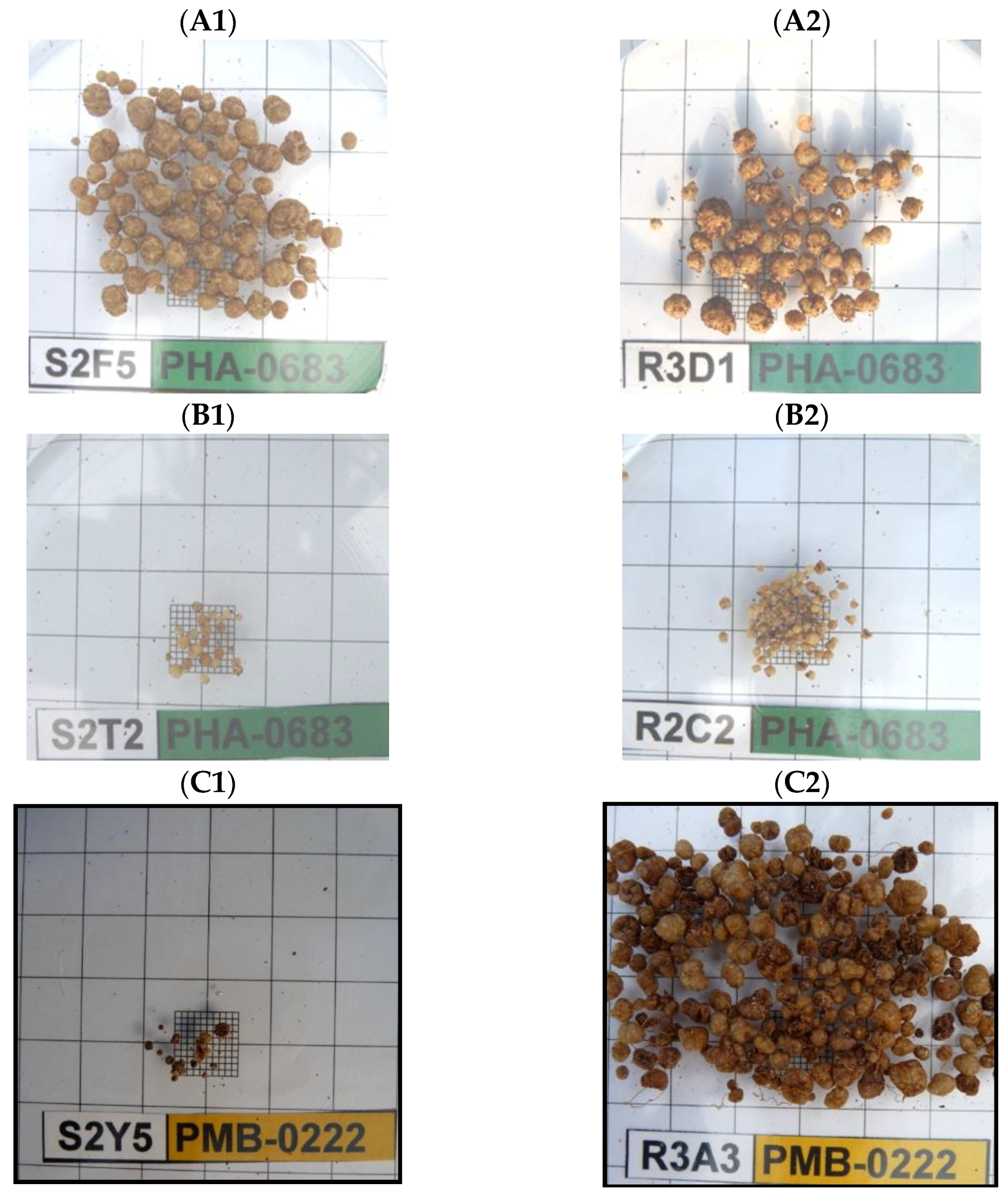{kind=link}
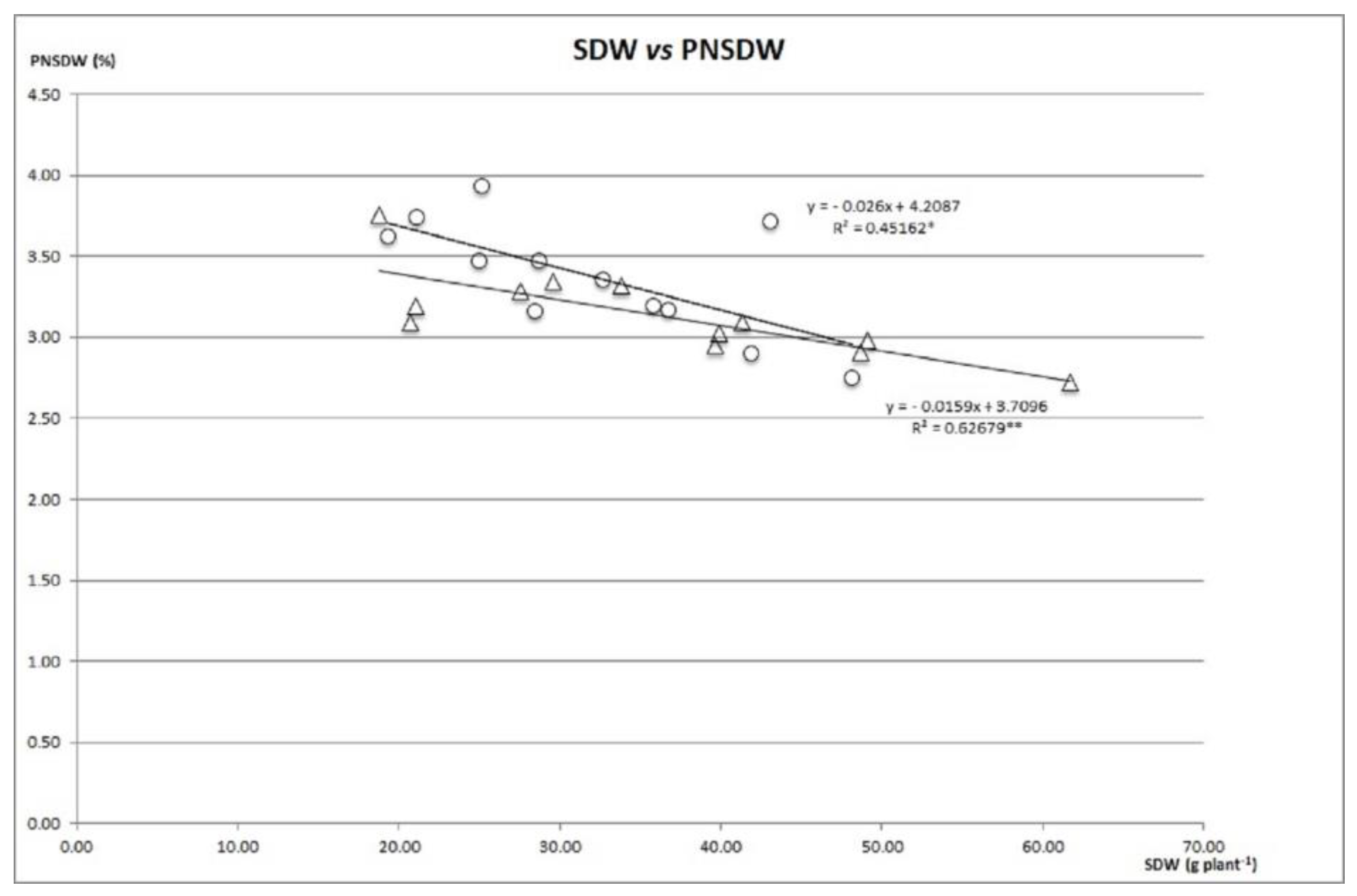{kind=link}
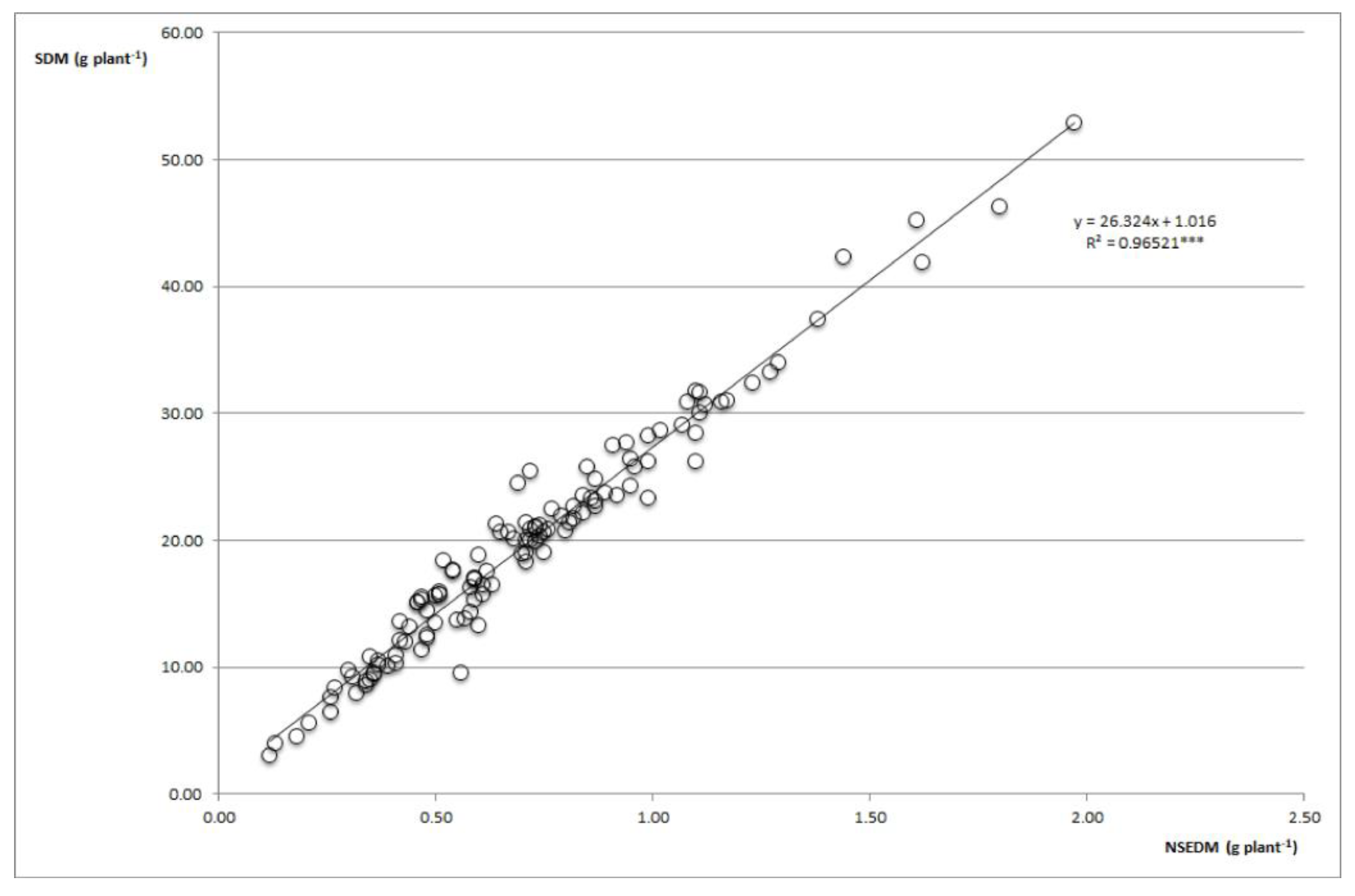{kind=link}
| A. Bean genotypes used in this study, name, commercial class and performance under drought. | |||
| Bean Genotypes | Name | Commercial Class | Drought Performance |
| PHA-0155 | local variety | White Kidney | unknown |
| PHA-0432 | local variety | Marrow | unknown |
| PHA-0471 | local variety | Navy | unknown |
| PHA-0483 | local variety | Guernikesa | unknown |
| PHA-0683 | local variety | Cranberry | unknown |
| PMB-0220 | Mattherhorn | Great Northern | tolerant |
| PMB-0222 | Almonga | Great Northern | unknown |
| PMB-0244 | Linex (R) | Cannellini | unknown |
| PMB-0285 | L88-18 (R) | Black Turtle | sensitive |
| PMB-0286 | L88-63 (R) | Black Turtle | tolerant |
| B. Rhizobia strains used. | |||
| Strains of Rhizobia | Type | ||
| SLL2 | local strain | ||
| EXIC | local strain | ||
| EXIB | local strain | ||
| EG | local strain | ||
| EF | local strain | ||
| EPOB | local strain | ||
| APAFI | local strain | ||
| LTMF | local strain | ||
| CIAT899 (R) | R. tropici | ||
| CFN42 (R) | R. etli | ||
| Traits Measured | Acronym | Units |
|---|---|---|
| Shoot dry weight, F, G | SDW | grams plant−1 |
| Root dry weight, G | RDW | grams plant−1 |
| Nodules number, F, G | NN | number plant−1 |
| Nodule dry weight, F, G | NDW | milligrams plant −1 |
| Medium nodular dry weight, F, G | MNDW | milligrams |
| 100 seed weight, F | 100SW | grams 100 seeds−1 |
| Seed dry matter, F | SDM | grams plant−1 |
| Percentage of nitrogen in shoot dry matter, F | PNSDW | % |
| Quantity of nitrogen in shoot dry matter, F | NSDW | grams plant−1 |
| Percentage of nitrogen in seed dry matter, F | PNSEDM | % |
| Nitrogen content in seed dry matter, F | NSEDM | grams plant−1 |
| Source of Variation | SDW | RDW | NN | NDW |
|---|---|---|---|---|
| G | *** | *** | ** | ** |
| T | *** | *** | ** | ns |
| I | ns | ns | ns | ns |
| G×T | *** | *** | * | ns |
| G×I | ns | * | ns | ns |
| T×I | ns | ns | ns | ns |
| G×T×I | ns | ns | ns | ns |
| SDW | NSDW | PNSDW | SDM | NSEDM | PNSEDM | NN | NDW | 100SW | |
|---|---|---|---|---|---|---|---|---|---|
| G | *** | *** | *** | *** | *** | *** | *** | *** | *** |
| T | *** | *** | *** | *** | *** | *** | *** | *** | *** |
| I | *** | *** | *** | *** | *** | * | ns | ns | *** |
| G×T | *** | *** | *** | ** | ** | *** | ** | ns | * |
| G×I | *** | *** | ns | ns | ns | ns | ns | ns | * |
| T×I | *** | ** | *** | *** | *** | * | ns | ns | *** |
| G×T×I | *** | *** | ns | ns | ns | ns | ns | ns | *** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rodiño, A.P.; Riveiro, M.; De Ron, A.M. Implications of the Symbiotic Nitrogen Fixation in Common Bean under Seasonal Water Stress. Agronomy 2021, 11, 70. https://doi.org/10.3390/agronomy11010070
Rodiño AP, Riveiro M, De Ron AM. Implications of the Symbiotic Nitrogen Fixation in Common Bean under Seasonal Water Stress. Agronomy. 2021; 11(1):70. https://doi.org/10.3390/agronomy11010070
Chicago/Turabian StyleRodiño, Ana P., Manuel Riveiro, and Antonio M. De Ron. 2021. "Implications of the Symbiotic Nitrogen Fixation in Common Bean under Seasonal Water Stress" Agronomy 11, no. 1: 70. https://doi.org/10.3390/agronomy11010070
APA StyleRodiño, A. P., Riveiro, M., & De Ron, A. M. (2021). Implications of the Symbiotic Nitrogen Fixation in Common Bean under Seasonal Water Stress. Agronomy, 11(1), 70. https://doi.org/10.3390/agronomy11010070