
Cold Stress during Flowering Alters Plant Structure, Yield and Seed Quality of Different Soybean Genotypes

 ,
,  ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Experimental Conditions
2.3. Scope of Tests and Measurements
2.4. Statistical Analyses
3. Results
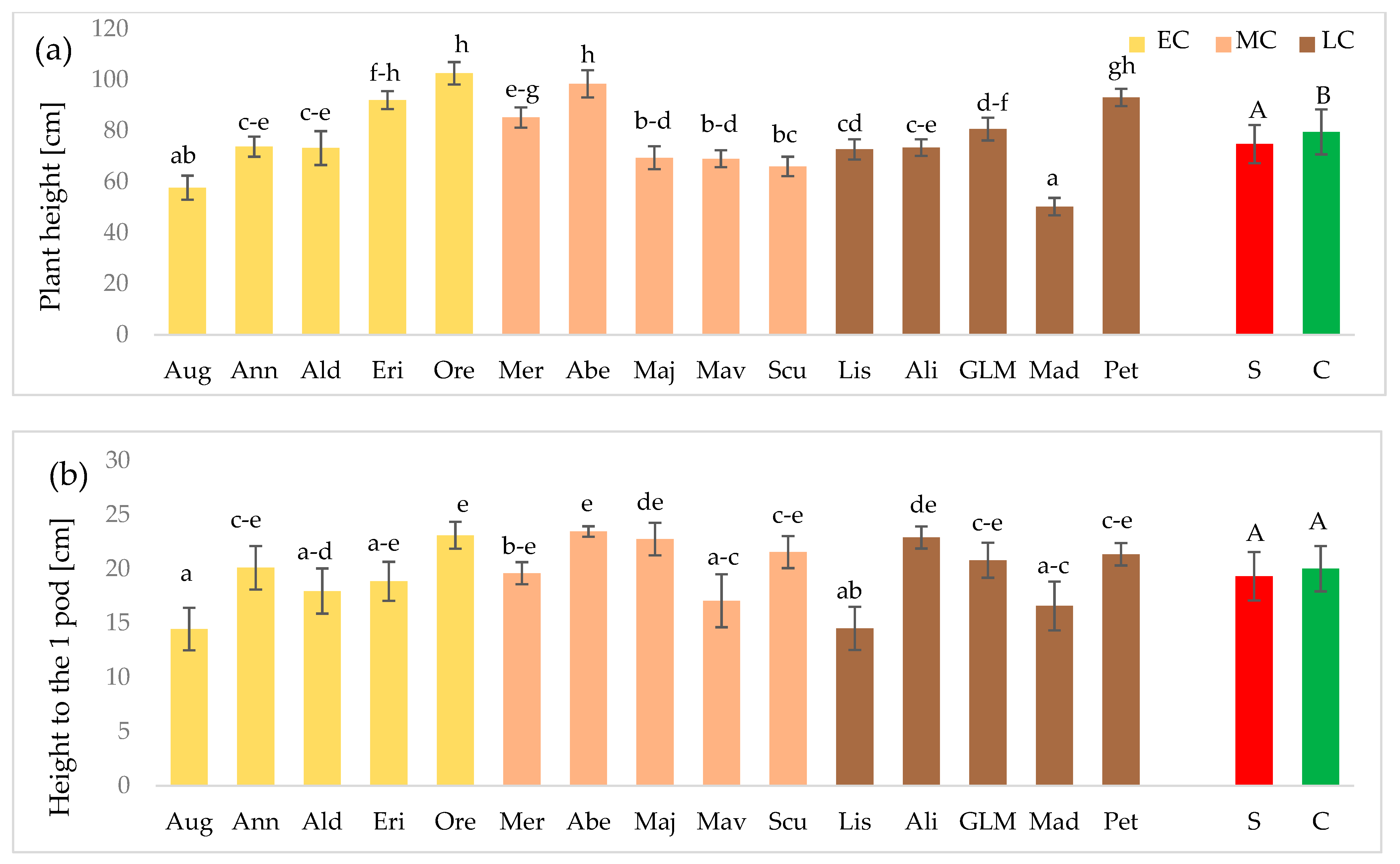
3.1. Plant Morphological Features and Elements of the Yield Structure
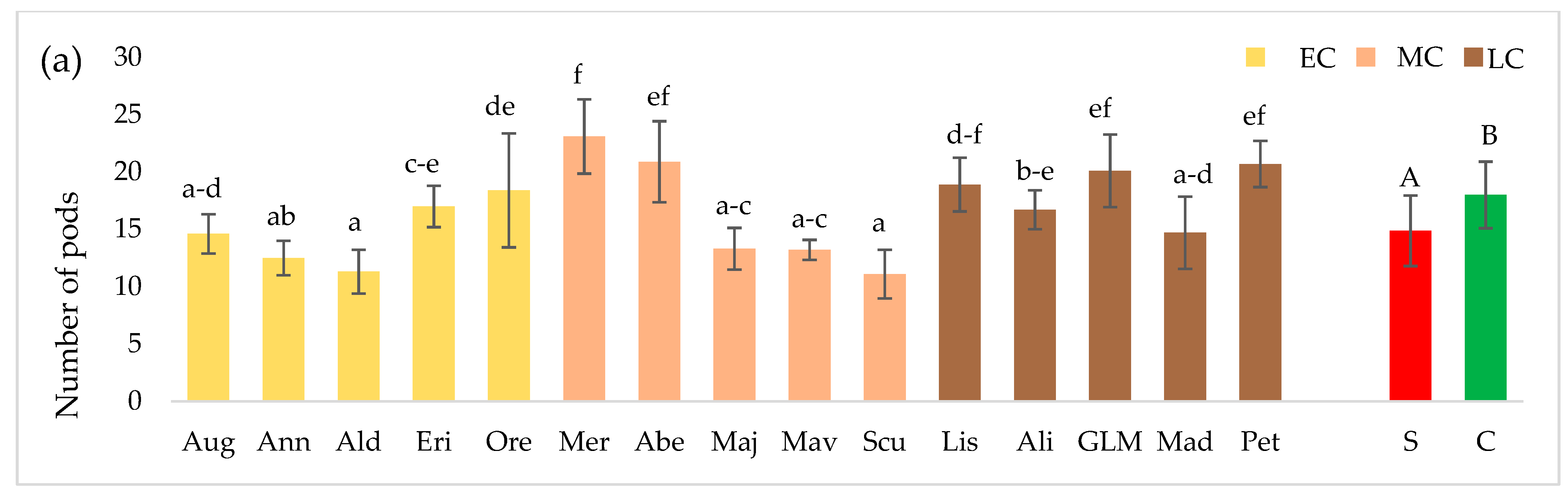
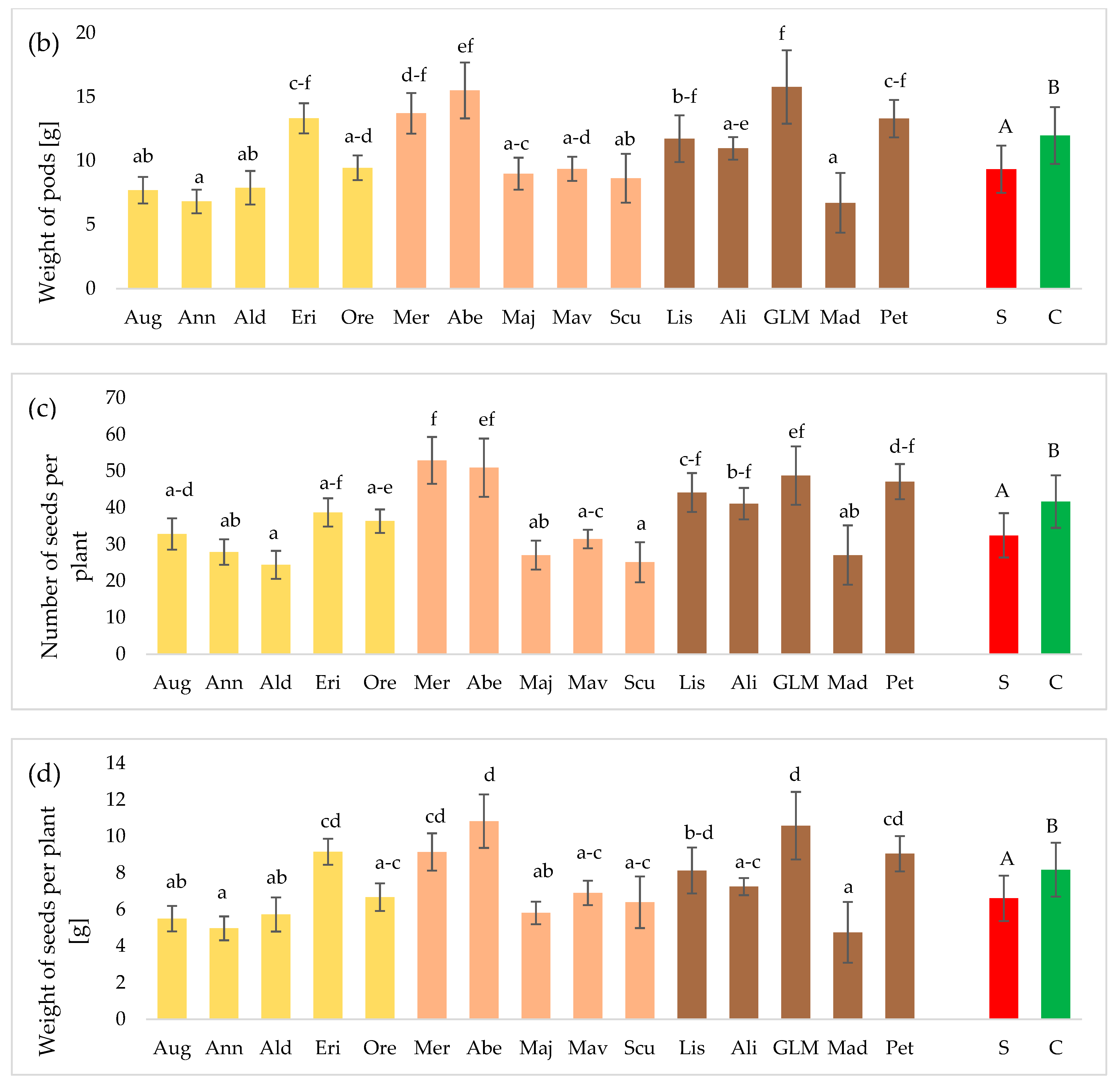
3.2. Seed Yield and 1000 Seed Weight (TSW)
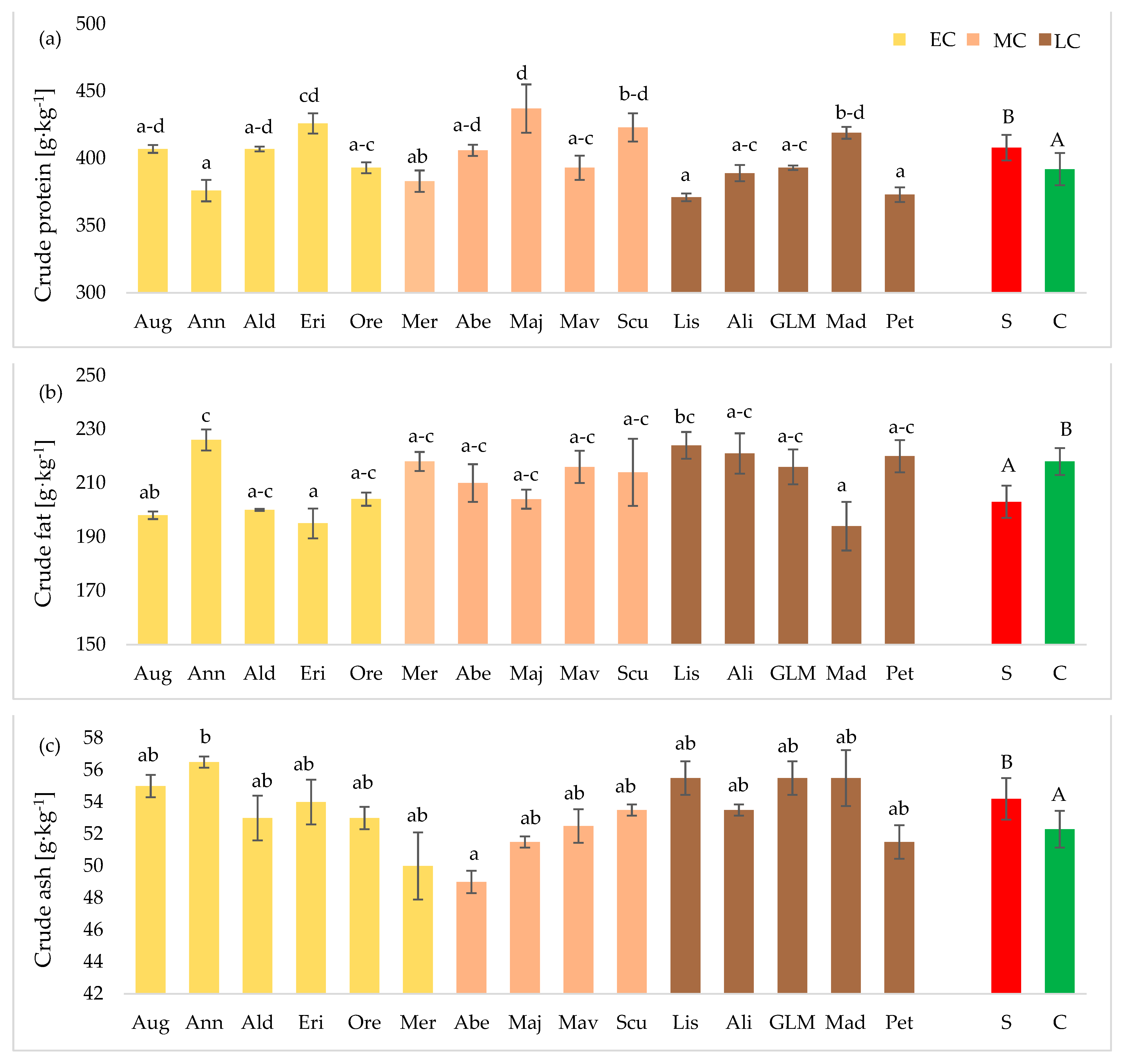
3.3. Chemical Composition of Seeds
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAOSTAT 2020. Available online: http://www.fao.org/faostat/en/-data/QC (accessed on 12 January 2021).
- Kotecki, A.; Lewandowska, S. (Eds.) Studies on the Cultivation of Soybean (Glycine max (L.) Merrill) in South-Western Poland; UP: Wrocław, Poland, 2020; p. 226. (In Polish) [Google Scholar]
- Bellaloui, N.; Bruns, H.A.; Abbas, H.K.; Mengistu, A.; Fisher, D.K.; Reddy, K.N. Agricultural practices altered soybean seed protein, oil, fatty acids, sugars, and minerals in the Midsouth USA. Front. Plant Sci. 2015, 6, 1–14. [Google Scholar] [CrossRef]
- Bezdicek, D.F.; Abede, E.B.; Evans, D.W.; Witters, R.E. Evaluation of peat and granular inoculum for soybean yield and N fixation under irrigation. J. Agron. 1978, 70, 865–868. [Google Scholar] [CrossRef]
- Gass, T.; Schori, A.; Fossati, A.; Soldati, A.; Stamp, P. Cold tolerance of soybean (Glycine Max (L.) Merr.) during the reproductive phase. Eur. J. Agron. 1996, 5, 71–88. [Google Scholar] [CrossRef]
- Gaynor, L.G.; Lawn, R.J.; James, A.T. Agronomic studies on irrigated soybean in southern New South Wales. I. Phenological adaptation of genotypes to sowing date. Crop Past. Sci. 2011, 62, 1056–1066. [Google Scholar] [CrossRef]
- Wu, T.; Li, J.; Wu, C.; Sun, S.; Mao, T.; Jiang, B.; Hou, W.; Han, T. Analysis of the independent- and interactive-photo-thermal effects on soybean flowering. J. Integr. Agric. 2015, 14, 622–632. [Google Scholar] [CrossRef] [Green Version]
- Câmara, G.M.S.; Sediyama, T.; Dourado-Neto, D.; Bernardes, M.S. Influence of photoperiod and air temperature on the growth, flowering and maturation of soybean (Glycine Max (L.) Merrill). Sci. Agric. 1997, 54, 149–154. [Google Scholar] [CrossRef]
- Rahman, M.M.; Hampton, J.G.; Hill, M.J. Soybean development under the cool temperate environment of Canterbury, New Zealand. J. New Seeds 2008, 4, 17–36. [Google Scholar] [CrossRef]
- Kumagai, E.; Sameshima, R. Genotypic differences in soybean yield responses to increasing temperature in a cool climate are related to maturity group. Agric. For. Meteorol. 2014, 198–199, 265–272. [Google Scholar] [CrossRef]
- Ohnishi, S.; Miyoshi, T.; Shirai, S. Low temperature stress at different flower developmental stages affects pollen development, pollination, and pod set in soybean. Environ. Exp. Bot. 2010, 69, 56–62. [Google Scholar] [CrossRef]
- Nawracała, J. Efektywność kwitnienia zróżnicowanych genotypów soi (Glycine max (L.) Merrill) w warunkach środowiskowych Wielkopolski. Oilseed Crop. 2001, 26, 27–44. (In Polish) [Google Scholar]
- Gibson, L.R.; Mullen, R.E. Influence of day and night temperature on soybean seed yield. Crop Sci. 1996, 36, 98–104. [Google Scholar] [CrossRef]
- Wheeler, T.R.; Craufurd, P.Q.; Ellis, R.H.; Porter, J.R.; Prasad, P.V.V. Temperature variability and the yield of annual crops. Agric. Ecosyst. Environ. 2000, 82, 159–167. [Google Scholar] [CrossRef]
- Schlenker, W.; Roberts, M.J. Estimating the Impact of Climate Change on Crop Yields: The Importance of Nonlinear Temperature Effects; Working Paper; National Bureau of Economic Research: Cambridge, MA, USA, 2008; p. 13799. [Google Scholar]
- Mourtzinis, S.; Specht, J.E.; Lindsey, L.E.; Wiebold, W.J.; Ross, J.; Nafziger, E.D.; Herman, K.; Nathan, M.; Devillez, P.L.; Arriaga, F.J.; et al. Climate-induced reduction in US-wide soybean yields underpinned by region and in season specific responses. Nat. Plants 2015, 1, 14026. [Google Scholar] [CrossRef] [PubMed]
- Gao, C. The future of CRISPR technologies agriculture. Nat. Rev. Mol. Cell Biol. 2018, 19, 275–276. [Google Scholar] [CrossRef] [PubMed]
- Jiang, B.; Khan, M.A.A.; Song, W.; Sun, S.; Zhang, C.; Wu, T.; Wu, C.; Zhang, S. Natural variations of FT family genes in soybean varieties covering a wide range of maturity groups. BMC Genom. 2019, 20, 230. [Google Scholar] [CrossRef]
- Kozyra, J.; Doroszewski, A.; Nieróbca, A. Zmiany klimatyczne i ich przewidywany wpływ na rolnictwo w Polsce. Studia I Rap. IUNG-PIB 2009, 14, 243–257. (In Polish) [Google Scholar] [CrossRef]
- Thompson, J.A.; Nelson, R.L. Utilization of diverse germplasm for soybean yield improvement. Crop Sci. 1998, 38, 1362–1368. [Google Scholar] [CrossRef]
- Mikel, M.A.; Diers, B.W.; Nelson, R.L.; Smith, H.H. Genetic diversity and agronomic improvement of North American soybean germplasm. Crop Sci. 2010, 50, 1219–1229. [Google Scholar] [CrossRef] [Green Version]
- Kusano, K.; Baxter, I.; Fukushima, A.; Oikawa, A.; Okazaki, Y.; Nakabayashi, R.; Bouvrette, D.J.; Achard, F.; Jakubowski, A.R.; Ballam, J.M.; et al. Assessing metabolomic and chemical diversity of a soybean lineage representing 35 years of breeding. Metabolomics 2015, 11, 261–270. [Google Scholar] [CrossRef]
- Holmberg, S.A. Soybeans for cool temperate climates. Agri. Horti. Genet. 1973, 31, 1–20. [Google Scholar]
- Schmid, J.; Keller, E.R. The behavior of three cold-tolerant and a standard soybean variety in relation to the level and the duration of a cold stress. Can. J. Plant Sci. 1980, 60, 821–829. [Google Scholar] [CrossRef]
- Hume, D.J.; Jackson, A.K.H. Pod formation in soybeans at low temperatures. Crop Sci. 1981, 21, 933. [Google Scholar] [CrossRef]
- Kurosaki, H.; Yumoto, S.; Matsukawa, I. Pod setting pattern during and after low temperature and the mechanism of cold-weather tolerance at the flowering stage in soybeans. Plant Prod. Sci. 2003, 6, 247–254. [Google Scholar] [CrossRef]
- Hou, G.; Ablett, G.R.; Pauls, K.P.; Rajcan, I. Environmental effects on fatty acid levels in soybean seed oil. J. Am. Oil Chem. Soc. 2006, 83, 759–763. [Google Scholar] [CrossRef]
- Shao, H.B.; Chu, L.Y.; Lu, Z.H.; Kang, C.M. Primary antioxidant free radical scavenging and redox signaling pathways in higher plant cells. Int. J. Biol. Sci. 2008, 4, 8–14. [Google Scholar] [CrossRef]
- Thomas, J.M.G.; Boote, K.J.; Allen, L.H.; Gallo-Meagher, M.; Davis, J.M. Elevated temperature and carbon dioxide effects on soybean seed composition and transcript abundance. Crop Sci. 2003, 43, 1548–1557. [Google Scholar] [CrossRef]
- Wilson, R.F. Seed composition. In Soybeans: Improvement, Production, and Uses; Boerma, H.R., Specht, J.E., Wilson, R.F., Eds.; The American Society of America, Crop Science Society of America, Soil Science Society of America: Madison, WI, USA, 2004; pp. 621–677. [Google Scholar] [CrossRef]
- Yaklich, R.W.; Vinyard, B.T. A method to estimate soybean seed protein and oil concentration before harvest. J. Am. Oil Chem. Soc. 2004, 81, 1021–1027. [Google Scholar] [CrossRef]
- Rinker, K.; Nelson, R.; Specht, J.; Sleper, D.; Cary, R.; Cianzio, S.; Casteel, S.; Conley, S.; Chen, P.; Davis, V.; et al. Genetic improvement of soybean in maturity groups II, III, and IV. Crop Sci. 2014, 54, 1–14. [Google Scholar] [CrossRef]
- Patil, G.; Mian, R.; Vuong, T.; Pantalone, V.; Song, Q.; Chen, P.; Shannon, J.; Carter, T.C.; Nguyen, H.T. Molecular mapping and genomics of soybean seed protein: A review and perspective for the future. Theor. Appl. Genet. 2017, 130, 1975–1991. [Google Scholar] [CrossRef] [Green Version]
- Mourtzinis, S.; Gaspar, A.P.; Naeve, S.L.; Conley, S.P. Planting date, maturity, and temperature effects on soybean seed yield and composition. J. Agron. 2017, 109, 2040–2049. [Google Scholar] [CrossRef] [Green Version]
- Kumar, V.; Rani, A.; Solanki, S.; Hussain, S.M. Influence of growing environment on the biochemical composition and physical characteristics of soybean seed. J. Food Compos. Anal. 2006, 19, 188–195. [Google Scholar] [CrossRef]
- EU Common Catalogue of Varieties of Agricultural Plant Species. Consolidated Version. 20 September 2019. Available online: https://ec.europa.eu/food/sites/food/files/plant/docs/plant_variety_catalogues_agriculturalplant- (accessed on 6 September 2021).
- Mourtzinis, S.; Conley, S.P. Delineating soybean maturity groups across the United States. Agron. J. 2017, 109, 1397–1403. [Google Scholar] [CrossRef] [Green Version]
- Kjeldahl, J.A. New method for the determination of nitrogen in organic matter. Z. Anal. Chem. 1883, 22, 366–382. [Google Scholar] [CrossRef] [Green Version]
- Soxhlet, F. The weight analytic determination of milk fat. Polytech. J. 1879, 232, 461–465. [Google Scholar]
- Official Methods of Analysis of the AOAC. Wyd. XX; Association of Official Analytical Chemists: Rockville, MD, USA, 2016.
- Nogala-Kałucka, M. (Ed.) Analiza Żywności. In Wybrane Metody Oznaczeń Jakościowych i Ilościowych Składników Żywności; UP: Poznan, Poland, 2016; p. 165. (In Polish) [Google Scholar]
- Jones, D.B. Factors for Converting Percentages of Nitrogen in Foods and Feed into Percentages of Proteins; Circular, 1941, No. 183 (Original Version, 1931); United States Department of Agriculture: Washington, DC, USA, 1931.
- Staniak, M.; Stępień-Warda, A.; Czopek, K.; Kocira, A.; Baca, E. Seeds quality and quantity of soybean [Glicine max (L.) Merr.] cultivars in response to cold stress. Agronomy 2021, 11, 520. [Google Scholar] [CrossRef]
- Kurosaki, H.; Yumoto, S. Effects of low temperature and shading during flowering on the yield components in soybeans. Plant Prod. Sci. 2003, 6, 17–23. [Google Scholar] [CrossRef]
- Sionit, N.; Strain, B.R.; Flint, E.P. Interaction of temperature and CO2 enrichment on soybean: Photosynthesis and seed yield. Can. J. Plant Sci. 1987, 67, 629–636. [Google Scholar] [CrossRef]
- Hatfield, J.L.; Boote, K.J.; Kimnball, B.A.; Ziska, L.H.; Izaurralde, R.C.; Ort, D.; Thomson, A.M.; Wolfe, D. Climate impacts on agriculture: Implications for crop production. Agron. J. 2011, 103, 351–370. [Google Scholar] [CrossRef] [Green Version]
- Salem, M.A.; Kakani, V.G.; Koti, S.; Reddy, K.R. Pollen-based screening of soybean genotypes for high temperatures. Crop Sci. 2007, 47, 219–231. [Google Scholar] [CrossRef] [Green Version]
- Kantolic, A.G.; Slafer, G.A. Development and seed number in indeterminate soybean as affected by timing and duration of exposure to long photoperiods after flowering. Ann. Bot. 2007, 99, 925–933. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kołodziej, J.; Pisulewska, E. Wpływ czynników meteorologicznych na plon nasion i tłuszczu oraz zawartość tłuszczu w nasionach dwóch odmian soi. Oilseed Crop. 2000, 21, 759–773. (In Polish) [Google Scholar]
- Rotundo, J.L.; Westgate, M.E. Meta-analysis of environmental effects on soybean seed composition. Field Crop. Res. 2009, 110, 147–156. [Google Scholar] [CrossRef]
- Wolf, R.B.; Cavins, J.F.; Kleiman, R.; Black, L.T. Effect of temperature on soybean seed constituents: Oil, protein, moisture, fatty acids, amino acid, and sugars. J. Am. Oil Chem. Soc. 1982, 59, 230–232. [Google Scholar] [CrossRef]
- Dornbos, D.L.; Mullen, R.E. Soybean seed protein and oil contents and fatty acid composition adjustments by drought and temperature. J. Am. Oil Chem. Soc. 1992, 69, 228–231. [Google Scholar] [CrossRef]
- Khan, A.Z.; Shah, P.; Khan, H.; Nigar, S.; Perveen, S.; Shah, M.K.; Amanullah; Khalil, S.K.; Munir, S.; Zubair, M. Seed quality and vigor of soybean cultivars as influenced by canopy temperature. Pak. J. Bot. 2011, 43, 643–648. [Google Scholar]
- Rotundo, J.L.; Miller-Garvin, J.E.; Naeve, S.L. Regional and temporal variation in soybean seed protein and oil across the United States. Crop Sci. 2016, 56, 797–808. [Google Scholar] [CrossRef]
- Alsajri, F.A.; Wijewardana, C.; Irby, J.T.; Bellaloui, N.; Krutz, L.J.; Golden, B.; Gao, W.; Reddy, K.R. Developing functional relationships between temperature and soybean yield and seed quality. J. Agron. 2020, 112, 194–204. [Google Scholar] [CrossRef] [Green Version]
- Nakagawa, A.C.; Ario, N.; Tomita, Y.; Tanaka, S.; Murayama, N.; Mizuta, C.; Iwaya-Inoue, M.; Ishibashi, Y. High temperature during soybean seed development differentially alters lipid and protein metabolism. Plant Prod. Sci. 2020, 23, 504–512. [Google Scholar] [CrossRef] [Green Version]
- Hou, A.; Chen, P.L.; Shi, A.; Alloatti, J.; Zhang, B.; Wang, Y. Sugar variation soybean seed associated with a rapid extraction and quantification method. Int. J. Agron. 2009, 2009, 484571. [Google Scholar] [CrossRef] [Green Version]
- Arendt, E.K.; Zannini, E. Cereal Grains for the Food and Beverage Industries; Woodhead Publishing: Cambridge, UK, 2013; pp. 409–438. [Google Scholar] [CrossRef]
- Piper, E.L.; Boote, K.J. Temperature and cultivar effects on soybean seed oil and protein concentrations. J. Am. Oil Chem. Soc. 1999, 76, 1233–1241. [Google Scholar] [CrossRef]
- Nascimento, M.; Finoto, E.L.; Sediyama, T.; Cruz, C.D. Adaptability and stability of soybean in terms of oil and protein content. Crop Breed. Appl. Biotechnol. 2010, 10, 48–54. [Google Scholar] [CrossRef] [Green Version]
- Biel, W.; Gawęda, D.; Łysoń, E.; Hury, G. Wpływ czynników genetycznych i agrotechnicznych na wartość odżywczą nasion soi. Acta Agroph. 2017, 24, 395–404. (In Polish) [Google Scholar]
- Kumar, V.; Rani, A.; Goyal, L.; Dixit, A.K.; Manjaya, J.G.; Dev, J.; Swamy, M. Sucrose and raffinose family oligosaccharides (RFOs) in soybean seeds as influenced by genotype and growing location. J. Agric. Food Chem. 2010, 58, 5081–5085. [Google Scholar] [CrossRef] [PubMed]
- Kozak, M.; Malarz, W.; Kotecki, A.; Černý, I.; Serafin-Andrzejewska, M. The effect of different sowing rate and Asahi SL biostimulator. Oilseed Crop. 2008, 29, 217–230. (In Polish) [Google Scholar]








{kind=link}
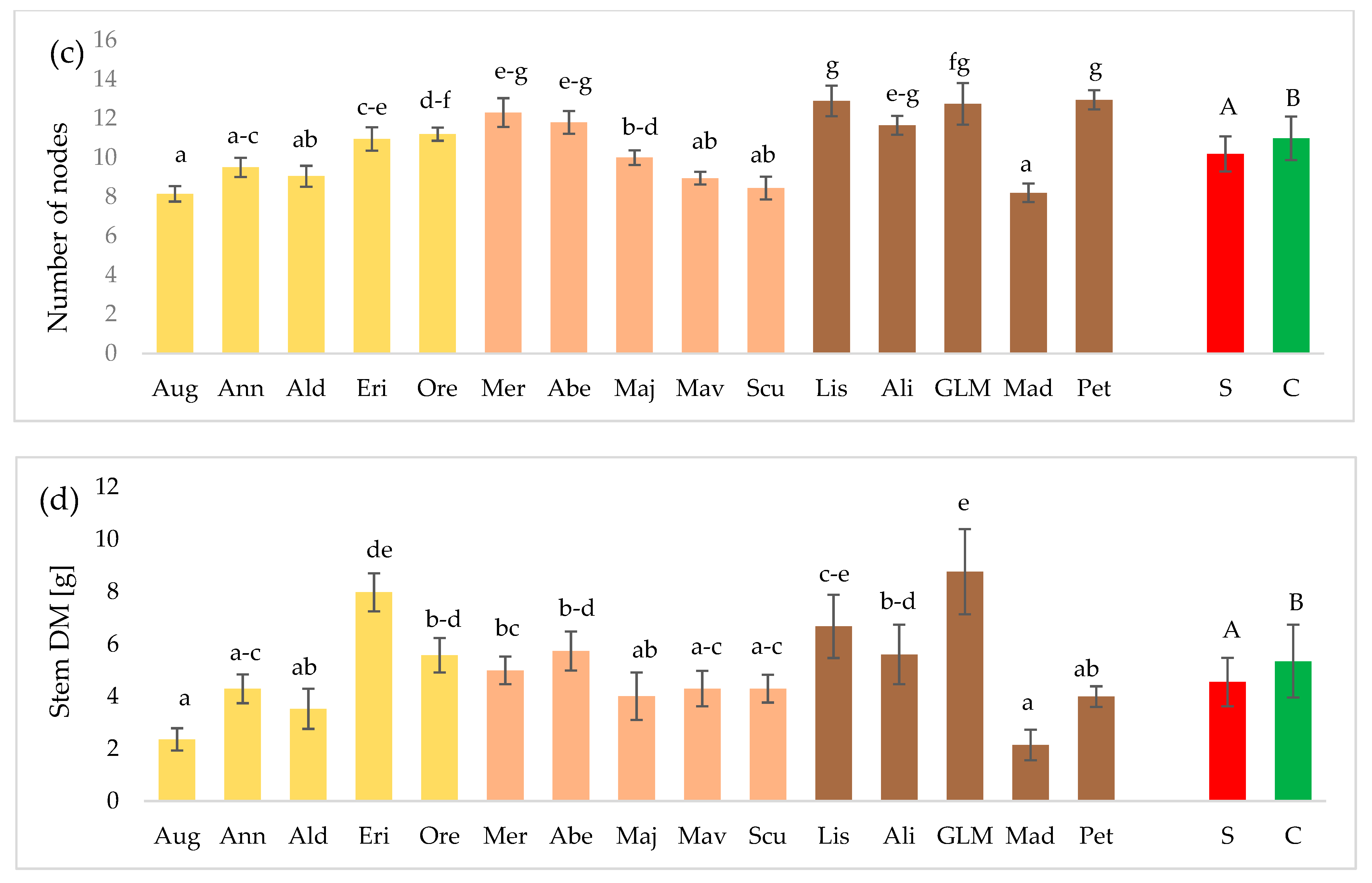{kind=link}
{kind=link}
{kind=link}
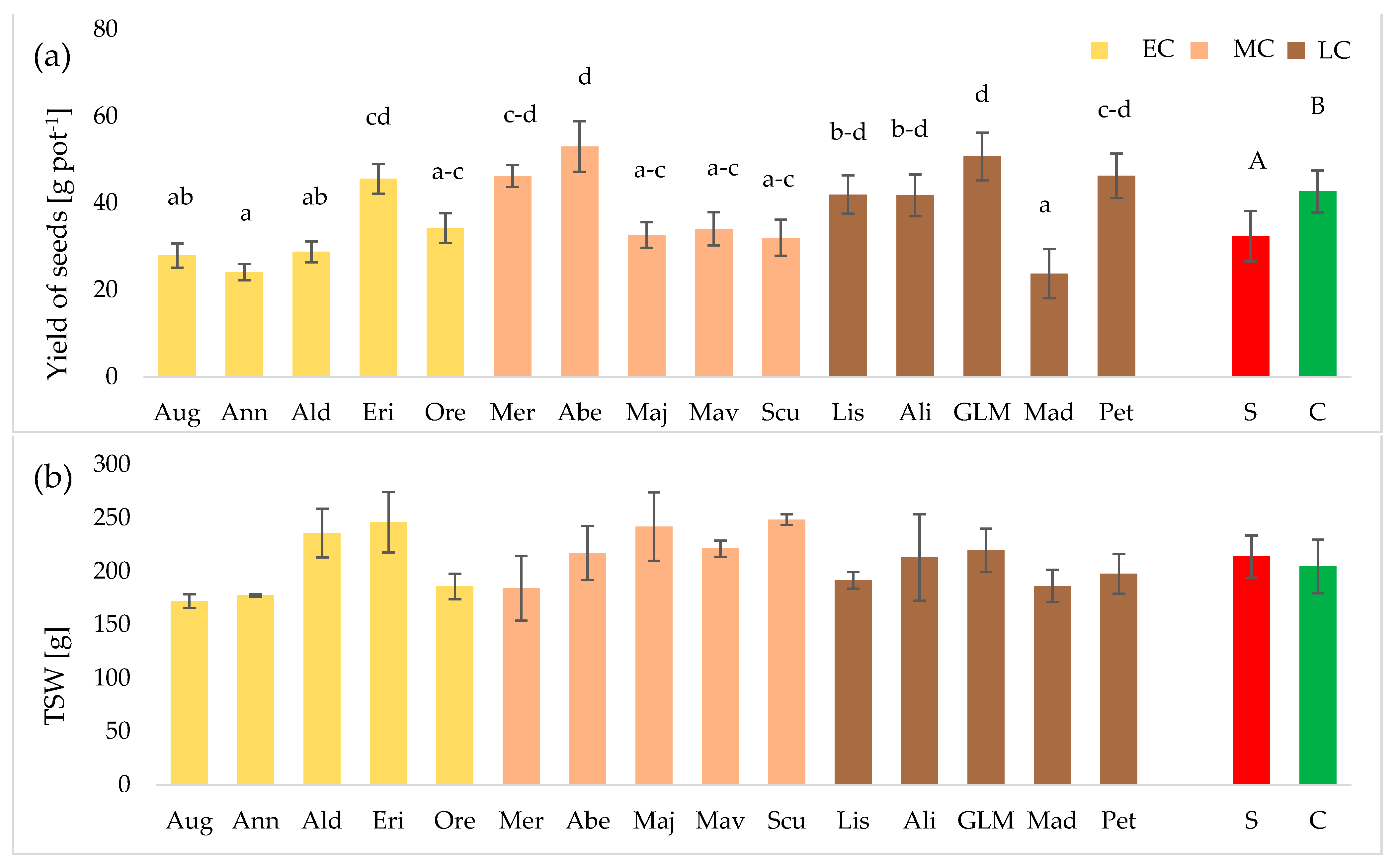{kind=link}
{kind=link}
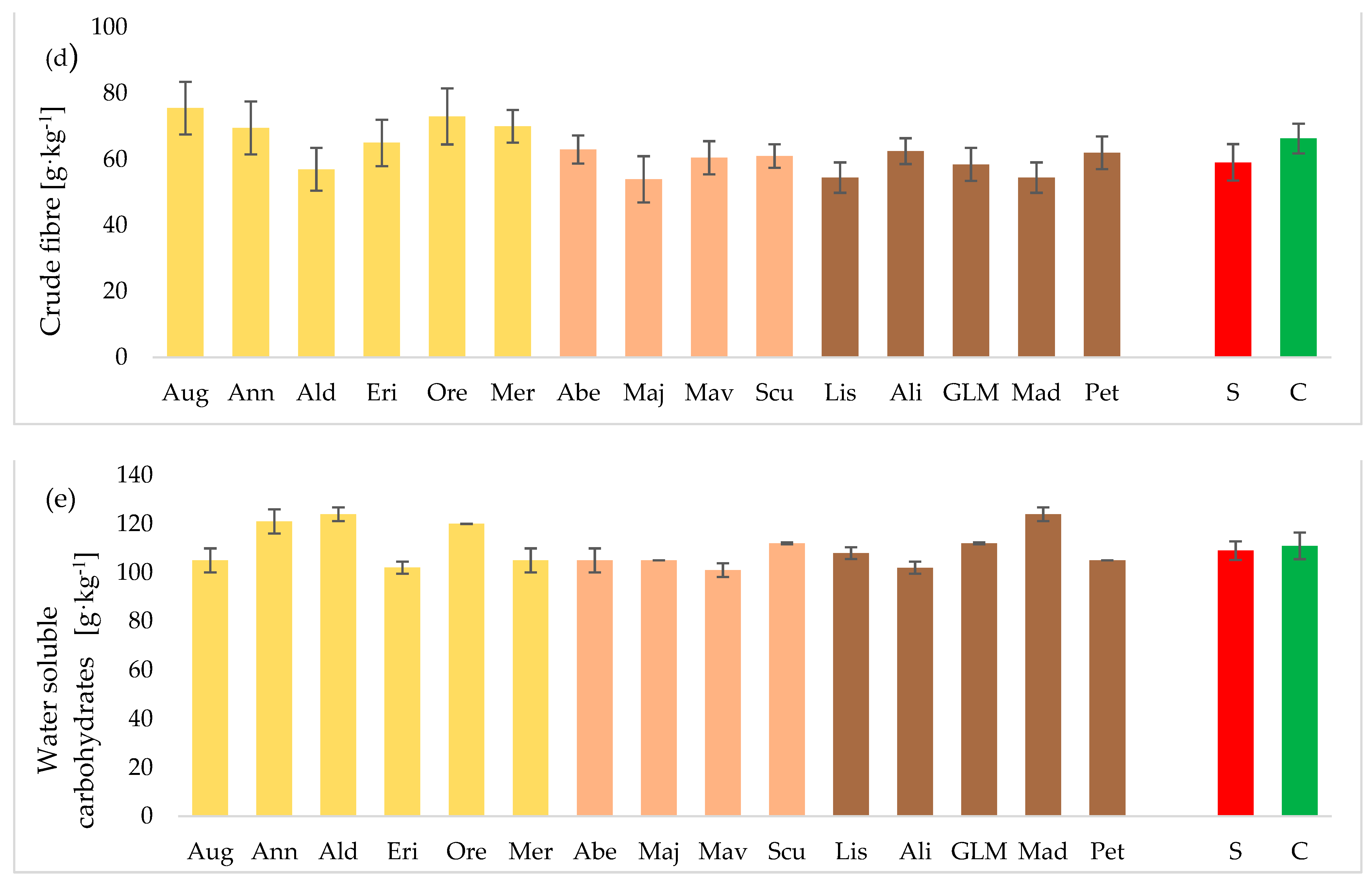{kind=link}
| Treatment | PH | H1P | NN | SDM | NP | WP | NS | WS | SY | TSW |
|---|---|---|---|---|---|---|---|---|---|---|
| Stress | ˂0.0001 | 0.1946 | ˂0.0001 | ˂0.0001 | ˂0.0001 | ˂0.0001 | ˂0.0001 | ˂0.0001 | ˂0.0001 | 0.4856 |
| Cultivar | ˂0.0001 | ˂0.0001 | ˂0.0001 | ˂0.0001 | ˂0.0001 | ˂0.0001 | ˂0.0001 | ˂0.0001 | ˂0.0001 | 0.4172 |
| Stress × Cultivar | 0.1052 | 0.2130 | 0.0430 | 0.1461 | 0.4762 | 0.7115 | 0.7802 | 0.5672 | 0.8385 | 0.9999 |
| Treatment | CP | CFa | WSC | CF | CA |
|---|---|---|---|---|---|
| Stress | 0.0006 | ˂0.0001 | 0.2586 | 0.0973 | 0.0050 |
| Cultivar | 0.0002 | 0.0029 | 0.0860 | 0.7155 | 0.0185 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Staniak, M.; Czopek, K.; Stępień-Warda, A.; Kocira, A.; Przybyś, M. Cold Stress during Flowering Alters Plant Structure, Yield and Seed Quality of Different Soybean Genotypes. Agronomy 2021, 11, 2059. https://doi.org/10.3390/agronomy11102059
Staniak M, Czopek K, Stępień-Warda A, Kocira A, Przybyś M. Cold Stress during Flowering Alters Plant Structure, Yield and Seed Quality of Different Soybean Genotypes. Agronomy. 2021; 11(10):2059. https://doi.org/10.3390/agronomy11102059
Chicago/Turabian StyleStaniak, Mariola, Katarzyna Czopek, Anna Stępień-Warda, Anna Kocira, and Marcin Przybyś. 2021. "Cold Stress during Flowering Alters Plant Structure, Yield and Seed Quality of Different Soybean Genotypes" Agronomy 11, no. 10: 2059. https://doi.org/10.3390/agronomy11102059
APA StyleStaniak, M., Czopek, K., Stępień-Warda, A., Kocira, A., & Przybyś, M. (2021). Cold Stress during Flowering Alters Plant Structure, Yield and Seed Quality of Different Soybean Genotypes. Agronomy, 11(10), 2059. https://doi.org/10.3390/agronomy11102059