
Assessing and Modeling Ecosystem Carbon Exchange and Water Vapor Flux of a Pasture Ecosystem in the Temperate Climate-Transition Zone


Abstract
:
1. Introduction
2. Material and Methods
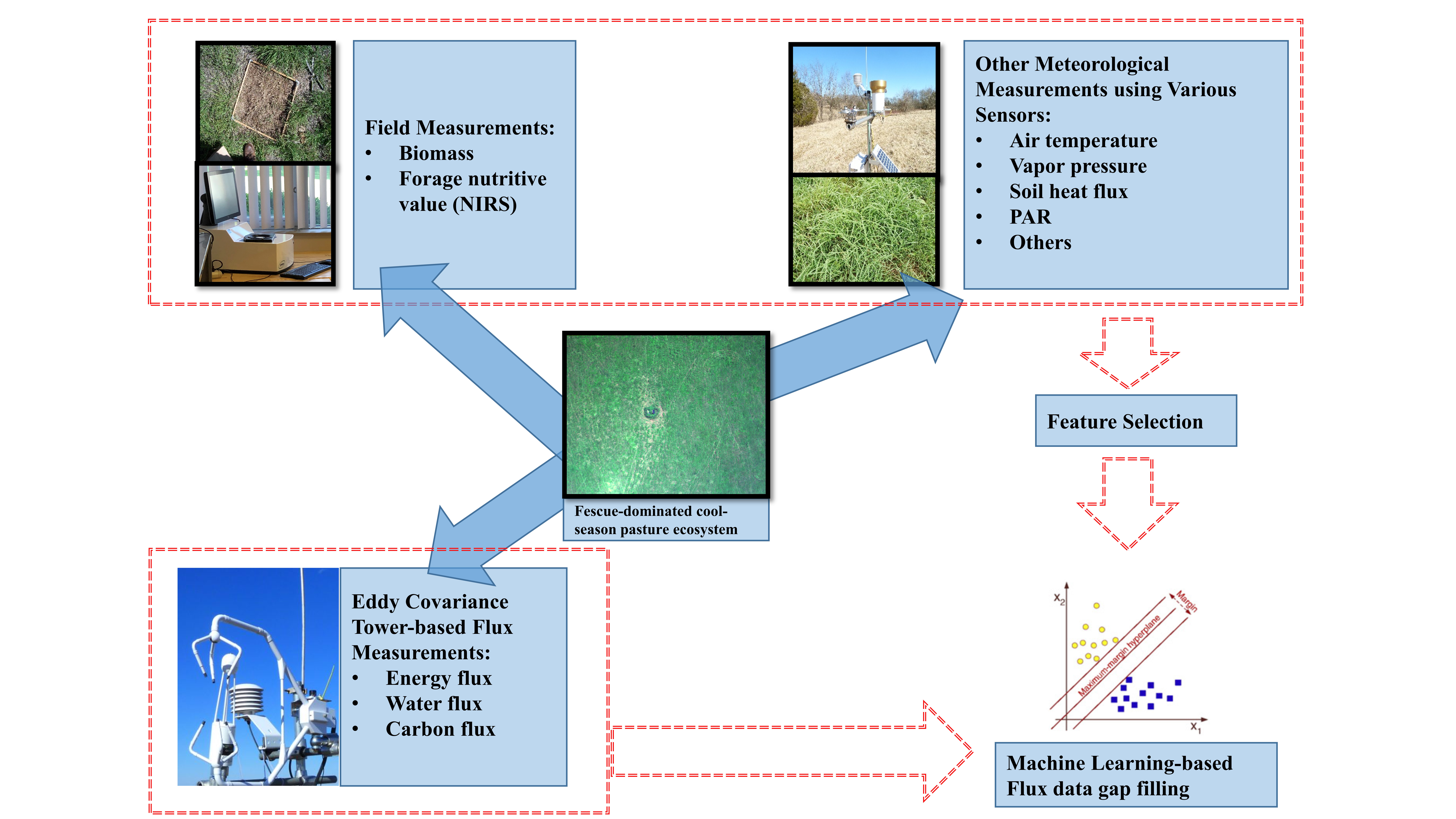
2.1. Study Site Description
2.2. Eddy Covariance Tower Establishment and Other Accessory Instruments
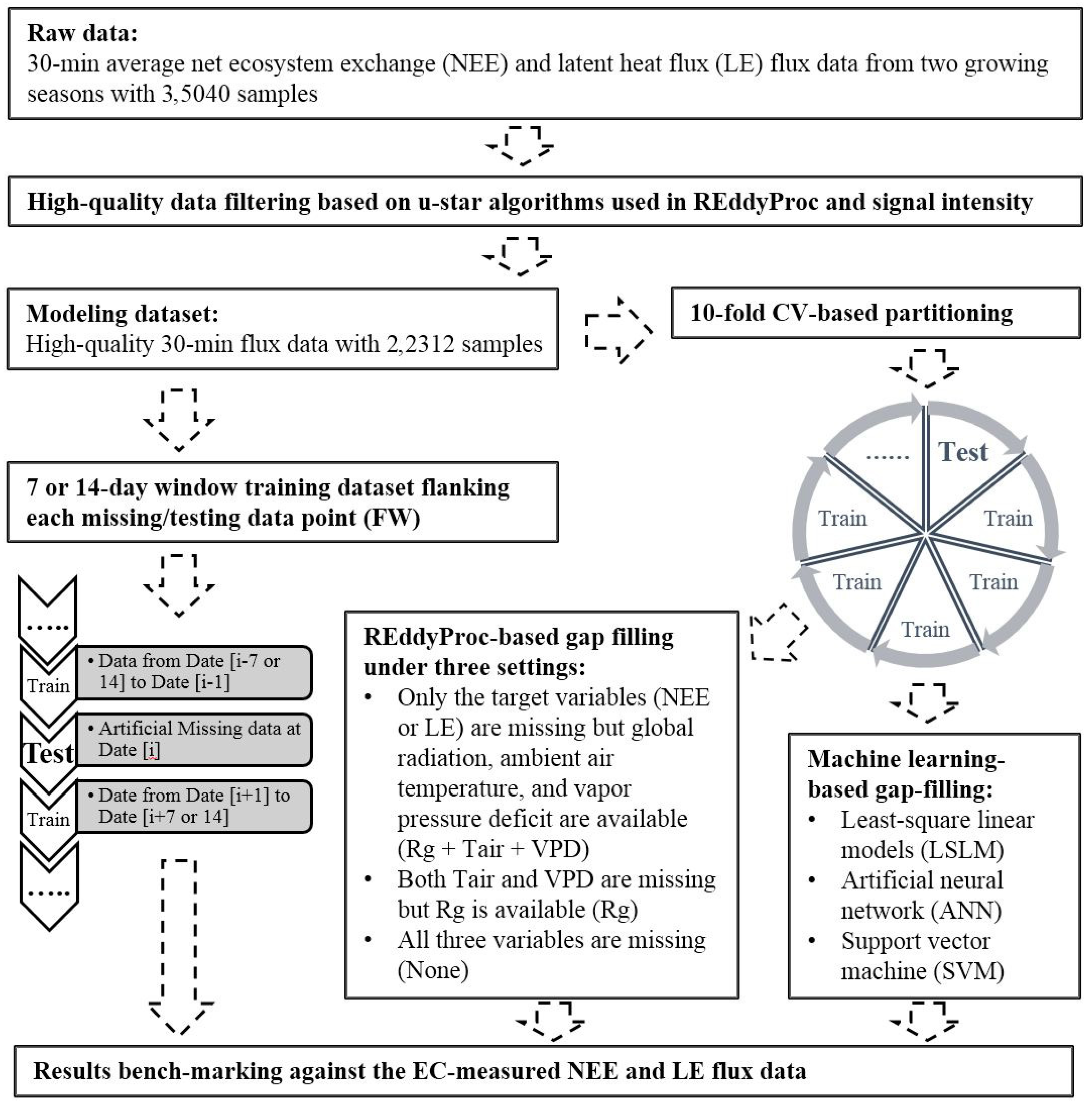
2.3. Flux Data Processing and Gap Filling
2.4. Plant Biomass Sampling and Nutritive Value Estimation
2.5. Modeling-Based Uncertainty Analysis and Benchmarking
3. Results
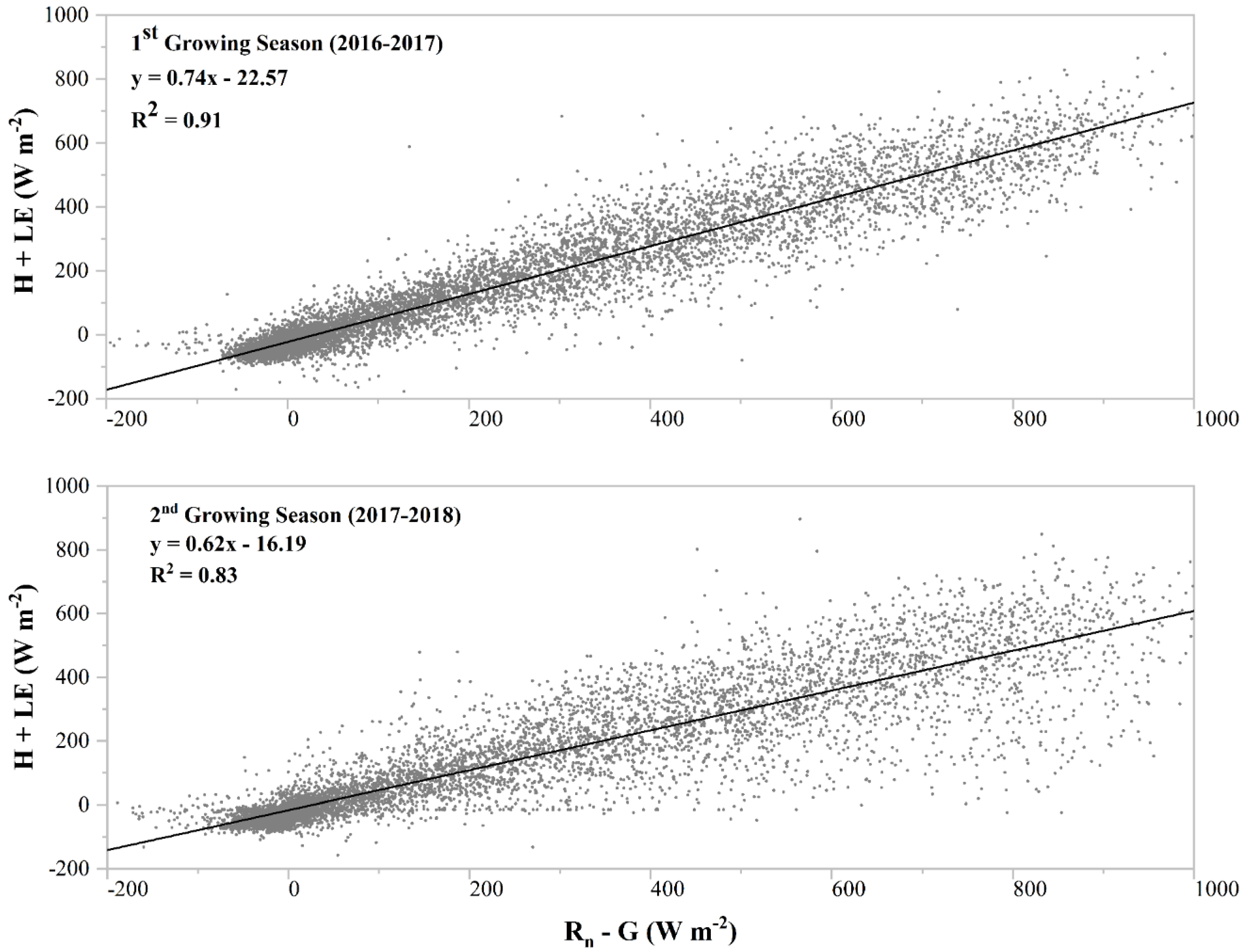
3.1. Energy Balance Analysis and Daily Cumulative Carbon Fluxes
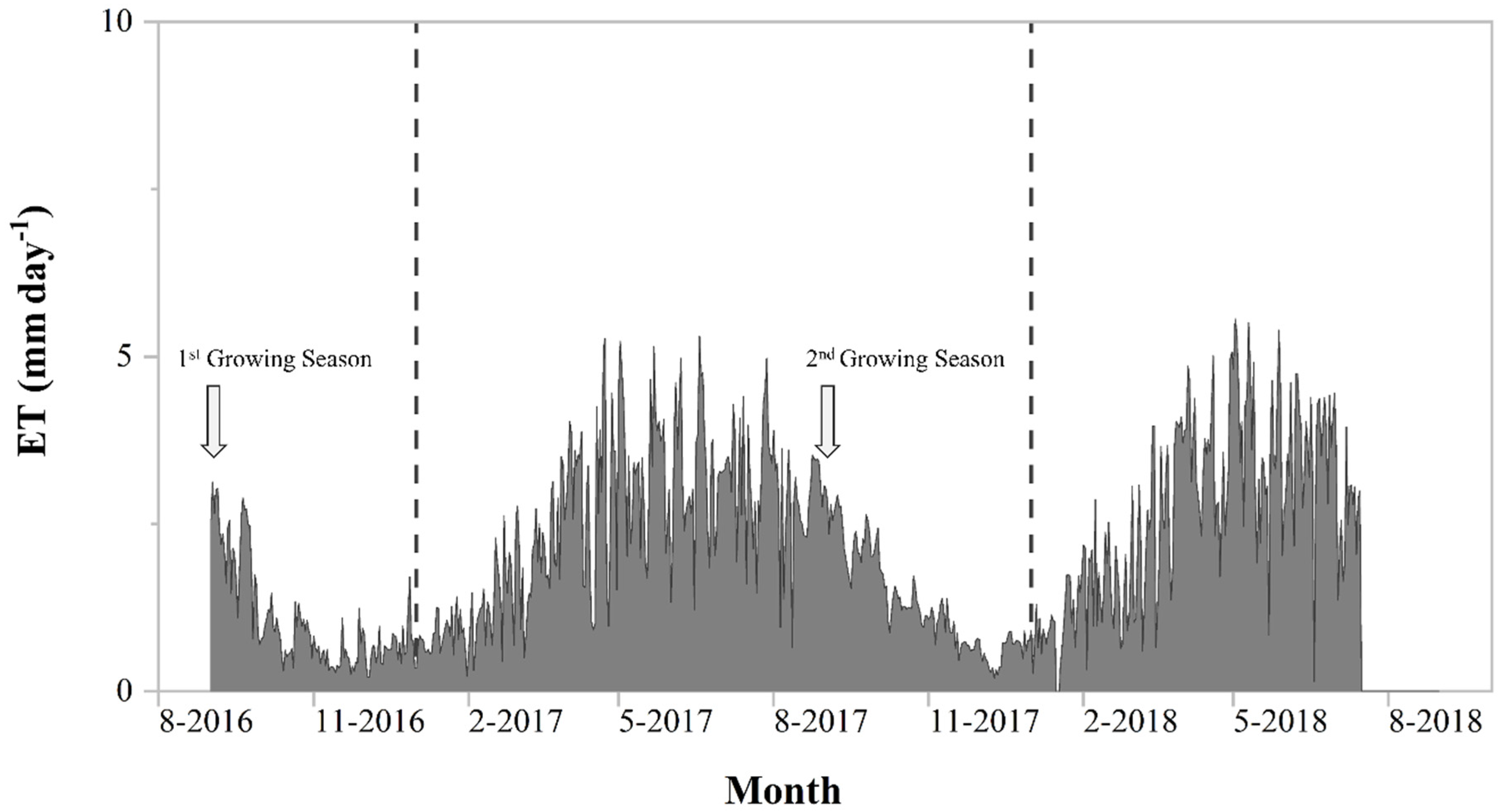
3.2. Evapotranspiration
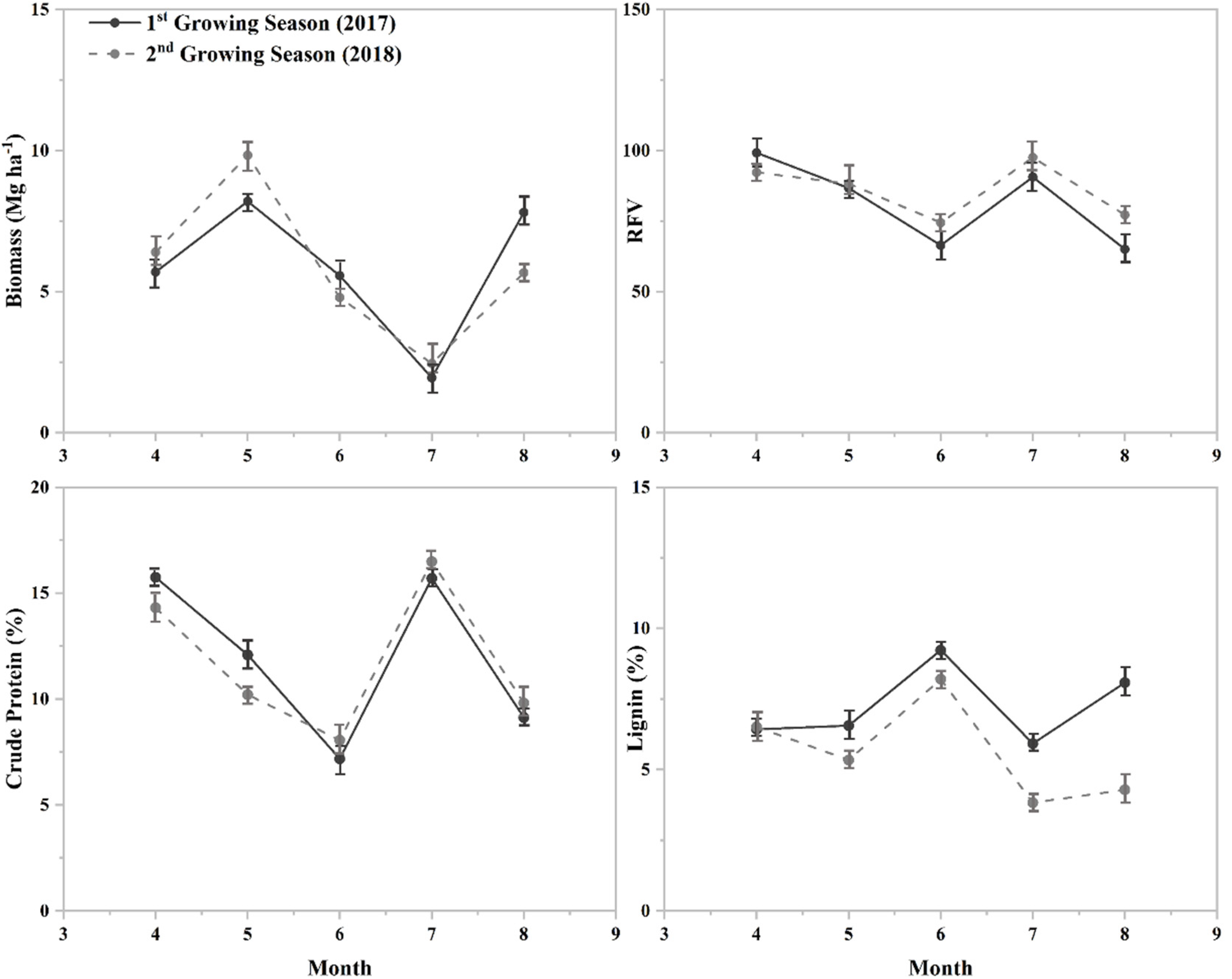
3.3. Plant Biomass and Nutritive Value
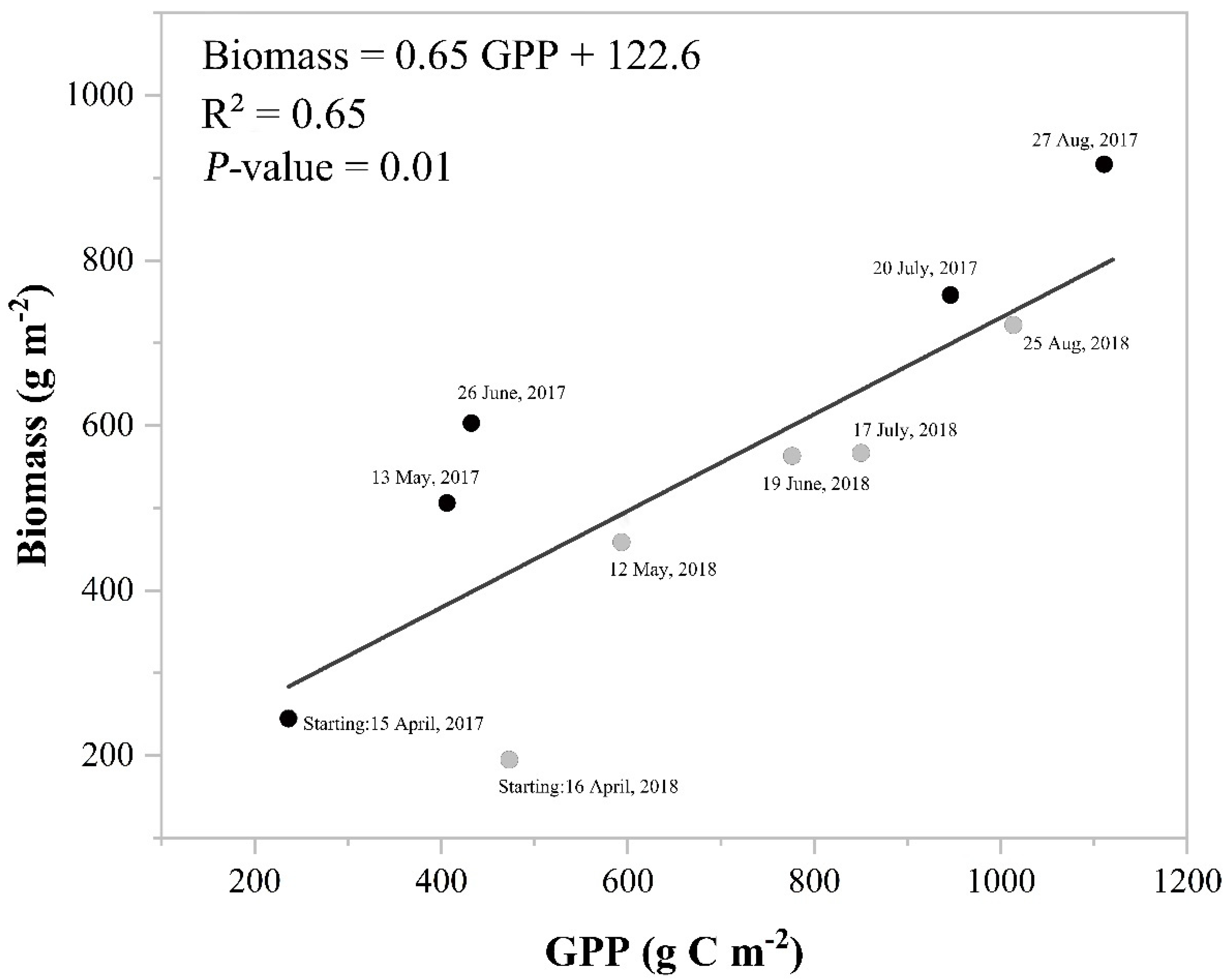
3.4. Flux Data Modeling, Uncertainty Analysis, and Benchmarking
4. Discussion
4.1. Energy Dynamics and Carbon Flux
4.2. Evapotranspiration and Ecosystem Water Use Efficiency
4.3. Biomass Productivity and Plant Nutritive Value
4.4. Model Performance
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Bond-Lamberty, B.; Thomson, A.M. Temperature-associated increases in the global soil respiration record. Nat. Cell Biol. 2010, 464, 579–582. [Google Scholar] [CrossRef] [PubMed]
- Cumming, G.; Buerkert, A.; Hoffmann, E.M.; Schlecht, E.; Von Cramon-Taubadel, S.; Tscharntke, T. Implications of agricultural transitions and urbanization for ecosystem services. Nat. Cell Biol. 2014, 515, 50–57. [Google Scholar] [CrossRef] [PubMed]
- Moss, R.H.; Edmonds, J.A.; Hibbard, K.A.; Manning, M.R.; Rose, S.K.; van Vuuren, D.P.; Carter, T.R.; Emori, S.; Kainuma, M.; Kram, T.; et al. The next generation of scenarios for climate change research and assessment. Nat. Cell Biol. 2010, 463, 747–756. [Google Scholar] [CrossRef] [PubMed]
- Kallenbach, R.; Roberts, C.; Lory, J.; Hamilton, S. Nitrogen Fertilization Rates Influence Stockpiled Tall Fescue Forage through Winter. Crop Sci. 2017, 57, 1732–1741. [Google Scholar] [CrossRef]
- Nelson, C.J.; Moore, K.J.; Collins, M. Forages and Grassland in a Changing World. In Forages, Volume 1: An Introduction to Grassland Agriculture, 7th ed.; Blackwell Publishing: Ames, IA, USA, 2018; pp. 3–18. [Google Scholar]
- Hunt, J.E.; Laubach, J.; Barthel, M.; Fraser, A.; Phillips, R.L. Carbon budgets for an irrigated intensively grazed dairy pasture and an unirrigated winter-grazed pasture. Biogeosciences 2016, 13, 2927–2944. [Google Scholar] [CrossRef] [Green Version]
- Leahy, P.; Kiely, G. The effect of introducing a winter forage rotation on CO2 fluxes at a temperate grassland. Agric. Ecosyst. Environ. 2012, 156, 49–56. [Google Scholar] [CrossRef] [Green Version]
- Lohila, A. Annual CO2exchange of a peat field growing spring barley or perennial forage grass. J. Geophys. Res. Space Phys. 2004, 109, 18116. [Google Scholar] [CrossRef]
- Skinner, H. Winter carbon dioxide fluxes in humid-temperate pastures. Agric. For. Meteorol. 2007, 144, 32–43. [Google Scholar] [CrossRef]
- Zhou, Y.; Xiao, X.; Wagle, P.; Bajgain, R.; Mahan, H.; Basara, J.; Dong, J.; Qin, Y.; Zhang, G.; Luo, Y.; et al. Examining the short-term impacts of diverse management practices on plant phenology and carbon fluxes of Old World bluestems pasture. Agric. For. Meteorol. 2017, 237–238, 60–70. [Google Scholar] [CrossRef] [Green Version]
- Kallenbach, R.L. Describing the Dynamic: Measuring and Assessing the Value of Plants in the Pasture. Crop Sci. 2015, 55, 2531–2539. [Google Scholar] [CrossRef]
- Bates, G.; Harper, C.; Allen, F. Forage & Field Crop Seeding Guide For Tennessee. The University of Tennessee Agricultural Extension Service, PB378 8/08 E12-5115-00-004-09 09-0041, 2008. Available online: http://trace.tennessee.edu/utk_agexcrop/41 (accessed on 12 July 2021).
- Reichstein, M.; Falge, E.; Baldocchi, D.; Papale, D.; Aubinet, M.; Berbigier, P.; Bernhofer, C.; Buchmann, N.; Gilmanov, T.; Granier, A.; et al. On the separation of net ecosystem exchange into assimilation and ecosystem respiration: Review and improved algorithm. Glob. Chang. Biol. 2005, 11, 1424–1439. [Google Scholar] [CrossRef]
- Papale, D.; Reichstein, M.; Aubinet, M.; Canfora, E.; Bernhofer, C.; Kutsch, W.; Longdoz, B.; Rambal, S.; Valentini, R.; Vesala, T.; et al. Towards a standardized processing of Net Ecosystem Exchange measured with eddy covariance technique: Algorithms and uncertainty estimation. Biogeosciences 2006, 3, 571–583. [Google Scholar] [CrossRef] [Green Version]
- Falge, E.; Baldocchi, D.; Olson, R.; Anthoni, P.; Aubinet, M.; Bernhofer, C.; Burba, G.; Ceulemans, R.J.; Clement, R.; Dolman, A.; et al. Gap filling strategies for defensible annual sums of net ecosystem exchange. Agric. For. Meteorol. 2001, 107, 43–69. [Google Scholar] [CrossRef] [Green Version]
- Abraha, M.; Gelfand, I.; Hamilton, S.K.; Shao, C.; Su, Y.-J.; Robertson, G.P.; Chen, J. Ecosystem Water-Use Efficiency of Annual Corn and Perennial Grasslands: Contributions from Land-Use History and Species Composition. Ecosystems 2016, 19, 1001–1012. [Google Scholar] [CrossRef]
- Rajan, N.; Maas, S.J.; Cui, S. Extreme Drought Effects on Carbon Dynamics of a Semiarid Pasture. Agron. J. 2013, 105, 1749–1760. [Google Scholar] [CrossRef] [Green Version]
- Zeeman, M.; Hiller, R.; Gilgen, A.K.; Michna, P.; Plüss, P.; Buchmann, N.; Eugster, W. Management and climate impacts on net CO2 fluxes and carbon budgets of three grasslands along an elevational gradient in Switzerland. Agric. For. Meteorol. 2010, 150, 519–530. [Google Scholar] [CrossRef] [Green Version]
- Wagle, P.; Kakani, V.G.; Huhnke, R. Evapotranspiration and Ecosystem Water Use Efficiency of Switchgrass and High Biomass Sorghum. Agron. J. 2016, 108, 1007–1019. [Google Scholar] [CrossRef]
- Wagle, P.; Gowda, P.H.; Moorhead, J.E.; Marek, G.W.; Brauer, D.K. Net ecosystem exchange of CO2 and H2O fluxes from irrigated grain sorghum and maize in the Texas High Plains. Sci. Total Environ. 2018, 637–638, 163–173. [Google Scholar] [CrossRef]
- Murray, I.; Cowe, I. Sample preparation. In Near-Infrared Spec-Troscopy in Agriculture; Roberts, C.A., Workman, J.J., Reeves, J.B., Eds.; ASA, CSSA, and SSSA: Madison, WI, USA, 2004; pp. 75–112. [Google Scholar]
- Wutzler, T.; Lucas-Moffat, A.; Migliavacca, M.; Knauer, J.; Sickel, K.; Šigut, L.; Menzer, O.; Reichstein, M. Basic and extensible post-processing of eddy covariance flux data with REddyProc. Biogeosciences 2018, 15, 5015–5030. [Google Scholar] [CrossRef] [Green Version]
- Kljun, N.; Calanca, P.; Rotach, M.W.; Schmid, H.P. A simple two-dimensional parameterisation for Flux Footprint Prediction (FFP). Geosci. Model Dev. 2015, 8, 3695–3713. [Google Scholar] [CrossRef] [Green Version]
- Foken, T. The Energy Balance Closure Problem: An Overview. Ecol. Appl. 2008, 18, 1351–1367. [Google Scholar] [CrossRef] [PubMed]
- Rajan, N.; Maas, S.J.; Cui, S. Extreme drought effects on summer evapotranspiration and energy balance of a grassland in the Southern Great Plains. Ecohydrology 2015, 8, 1194–1204. [Google Scholar] [CrossRef]
- Anapalli, S.S.; Fisher, D.K.; Reddy, K.N.; Krutz, J.L.; Pinnamaneni, S.R.; Sui, R. Quantifying water and CO2 fluxes and water use efficiencies across irrigated C3 and C4 crops in a humid climate. Sci. Total Environ. 2019, 663, 338–350. [Google Scholar] [CrossRef] [PubMed]
- Anapalli, S.S.; Fisher, D.K.; Reddy, K.N.; Wagle, P.; Gowda, P.H.; Sui, R. Quantifying soybean evapotranspiration using an eddy covariance approach. Agric. Water Manag. 2018, 209, 228–239. [Google Scholar] [CrossRef]
- Wilson, K.; Goldstein, A.; Falge, E.; Aubinet, M.; Baldocchi, D.; Berbigier, P.; Bernhofer, C.; Ceulemans, R.; Dolman, H.; Field, C.; et al. Energy balance closure at FLUXNET sites. Agric. For. Meteorol. 2002, 113, 223–243. [Google Scholar] [CrossRef] [Green Version]
- Collins, M.; Nelson, C.J. Grasses for Northern Areas. In Forages, Volume 1: An Introduction to Grassland Agriculture, 7th ed.; Collins, M., Nelson, C.J., Moore, K.J., Barnes, R.F., Eds.; Blackwell Publishing: Ames, IA, USA, 2018; pp. 131–132. [Google Scholar]
- Zeri, M.; Andersonteixeira, K.J.; Hickman, G.C.; Masters, M.D.; DeLucia, E.H.; Bernacchi, C. Carbon exchange by establishing biofuel crops in Central Illinois. Agric. Ecosyst. Environ. 2011, 144, 319–329. [Google Scholar] [CrossRef] [Green Version]
- Shurpali, N.J.; Hyvönen, N.P.; Huttunen, J.T.; Clement, R.J.; Reichstein, M.; Nykänen, H.; Biasi, C.; Martikainen, P.J. Cultivation of a perennial grass for bioenergy on a boreal organic soil—Carbon sink or source? GCB Bioenergy 2009, 1, 35–50. [Google Scholar] [CrossRef]
- Wu, J.; Bauer, M.E. Estimating Net Primary Production of Turfgrass in an Urban-Suburban Landscape with QuickBird Imagery. Remote Sens. 2012, 4, 849–866. [Google Scholar] [CrossRef] [Green Version]
- Cotten, D.L.; Zhang, G.; Leclerc, M.Y.; Raymer, P.; Steketee, C.J. Carbon dioxide fluxes from Tifway bermudagrass: Early results. Int. J. Biometeorol. 2017, 61, 103–113. [Google Scholar] [CrossRef]
- Skinner, R.H.; Adler, P.R. Carbon dioxide and water fluxes from switchgrass managed for bioenergy production. Agric. Ecosyst. Environ. 2010, 138, 257–264. [Google Scholar] [CrossRef]
- Cui, S.; Allen, V.G.; Brown, C.P.; Wester, D.B. Growth and Nutritive Value of Three Old World Bluestems and Three Legumes in the Semiarid Texas High Plains. Crop Sci. 2013, 53, 329–340. [Google Scholar] [CrossRef]
- Gelley, C.; Nave, R.L.G.; Bates, G. Forage Nutritive Value and Herbage Mass Relationship of Four Warm-Season Grasses. Agron. J. 2016, 108, 1603–1613. [Google Scholar] [CrossRef] [Green Version]
- Collins, M.; Newman, Y.C. Forage quality. In Forages, Volume 1: An Introduction to Grassland Agriculture, 7th ed.; Collins, M., Nelson, C.J., Moore, K.J., Barnes, R.F., Eds.; Blackwell Publishing: Ames, IA, USA, 2018; pp. 339–354. [Google Scholar]
- Gu, L.; Falge, E.M.; Boden, T.; Baldocchi, D.D.; Black, T.; Saleska, S.R.; Suni, T.; Verma, S.B.; Vesala, T.; Wofsy, S.C.; et al. Objective threshold determination for nighttime eddy flux filtering. Agric. For. Meteorol. 2005, 128, 179–197. [Google Scholar] [CrossRef] [Green Version]
- Acevedo, O.C.; Moraes, O.; Degrazia, G.A.; Fitzjarrald, D.R.; Manzi, A.O.; Campos, J.G. Is friction velocity the most appropriate scale for correcting nocturnal carbon dioxide fluxes? Agric. For. Meteorol. 2009, 149, 1–10. [Google Scholar] [CrossRef]
- Falge, E.; Baldocchi, D.; Olson, R.; Anthoni, P.; Aubinet, M.; Bernhofer, C.; Burba, G.; Ceulemans, R.; Clement, R.; Dolman, A.; et al. Gap filling strategies for long term energy flux data sets. Agric. For. Meteorol. 2001, 107, 71–77. [Google Scholar] [CrossRef] [Green Version]
- Hui, D.; Wan, S.; Su, B.; Katul, G.; Monson, R.; Luo, Y. Gap-filling missing data in eddy covariance measurements using multiple imputation (MI) for annual estimations. Agric. For. Meteorol. 2004, 121, 93–111. [Google Scholar] [CrossRef]
- Hsu, K.-L.; Gupta, H.; Gao, X.; Sorooshian, S.; Imam, B. Self-organizing linear output map (SOLO): An artificial neural network suitable for hydrologic modeling and analysis. Water Resour. Res. 2002, 38, 1302. [Google Scholar] [CrossRef] [Green Version]
- Kunwor, S.; Starr, G.; Loescher, H.W.; Staudhammer, C.L. Preserving the variance in imputed eddy-covariance measurements: Alternative methods for defensible gap filling. Agric. For. Meteorol. 2017, 232, 635–649. [Google Scholar] [CrossRef] [Green Version]
- Hayek, M.N.; Wehr, R.; Longo, M.; Hutyra, L.R.; Wiedemann, K.; Munger, J.W.; Bonal, D.; Saleska, S.R.; Fitzjarrald, D.R.; Wofsy, S.C. A novel correction for biases in forest eddy covariance carbon balance. Agric. For. Meteorol. 2018, 250–251, 90–101. [Google Scholar] [CrossRef]
- Dunn, A.L.; Barford, C.C.; Wofsy, S.C.; Goulden, M.L.; Daube, B.C. A long-term record of carbon exchange in a boreal black spruce forest: Means, responses to interannual variability, and decadal trends. Glob. Chang. Biol. 2007, 13, 577–590. [Google Scholar] [CrossRef] [Green Version]
- Moffat, A.M.; Papale, D.; Reichstein, M.; Hollinger, D.Y.; Richardson, A.D.; Barr, A.G.; Beckstein, C.; Braswell, B.; Churkina, G.; Desai, A.R.; et al. Comprehensive comparison of gap-filling techniques for eddy covariance net carbon fluxes. Agric. For. Meteorol. 2007, 147, 209–232. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target Variable | Algorithm | Training/Testing Partition Setting | Average | ||
|---|---|---|---|---|---|
| 10-Fold CV | 7-Day FW | 14-Day FW | |||
| NEE | LSLM | 0.57 (7.8) | 0.33 (17.7) | 0.42 (13.9) | 0.44 (13.3) |
| ANN | 0.64 (13.4) | 0.69 (13.4) | 0.02 (153.6) | 0.45 (60.1) | |
| SVM | 0.71 (12.7) | 0.77 (11.8) | 0.77 (11.8) | 0.75 (12.1) | |
| Feature Setting | |||||
| None | Rg | Rg + Tair + VPD | |||
| REddyProc | 0.21 (40.9) | 0.64 (15.7) | 0.56 (19.6) | 0.47 (25.4) | |
| Training/Testing Partition Setting | |||||
| 10-Fold CV | 7-Day FW | 14-Day FW | |||
| LE | LSLM | 0.61 (76.4) | 0.55 (132.8) | 0.57 (127.6) | 0.57 (112.2) |
| ANN | 0.77 (105.5) | 0.85 (91.7) | 0.86 (87.0) | 0.83 (94.7) | |
| SVM | 0.86 (86.8) | 0.90 (74.1) | 0.90 (73.8) | 0.89 (78.2) | |
| Feature Setting | |||||
| None | Rg | Rg + Tair + VPD | |||
| REddyProc | 0.79 (114.4) | 0.85 (95.6) | 0.87 (93.1) | 0.83 (101.0) | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Z.; Chen, C.; Nevins, A.; Pirtle, T.; Cui, S. Assessing and Modeling Ecosystem Carbon Exchange and Water Vapor Flux of a Pasture Ecosystem in the Temperate Climate-Transition Zone. Agronomy 2021, 11, 2071. https://doi.org/10.3390/agronomy11102071
Li Z, Chen C, Nevins A, Pirtle T, Cui S. Assessing and Modeling Ecosystem Carbon Exchange and Water Vapor Flux of a Pasture Ecosystem in the Temperate Climate-Transition Zone. Agronomy. 2021; 11(10):2071. https://doi.org/10.3390/agronomy11102071
Chicago/Turabian StyleLi, Zhou, Chao Chen, Andrew Nevins, Todd Pirtle, and Song Cui. 2021. "Assessing and Modeling Ecosystem Carbon Exchange and Water Vapor Flux of a Pasture Ecosystem in the Temperate Climate-Transition Zone" Agronomy 11, no. 10: 2071. https://doi.org/10.3390/agronomy11102071
APA StyleLi, Z., Chen, C., Nevins, A., Pirtle, T., & Cui, S. (2021). Assessing and Modeling Ecosystem Carbon Exchange and Water Vapor Flux of a Pasture Ecosystem in the Temperate Climate-Transition Zone. Agronomy, 11(10), 2071. https://doi.org/10.3390/agronomy11102071