
Negative Pressure Irrigation System Reduces Soil Nitrogen Loss for Lettuce during Greenhouse Production



Abstract
:1. Introduction
2. Materials and Methods
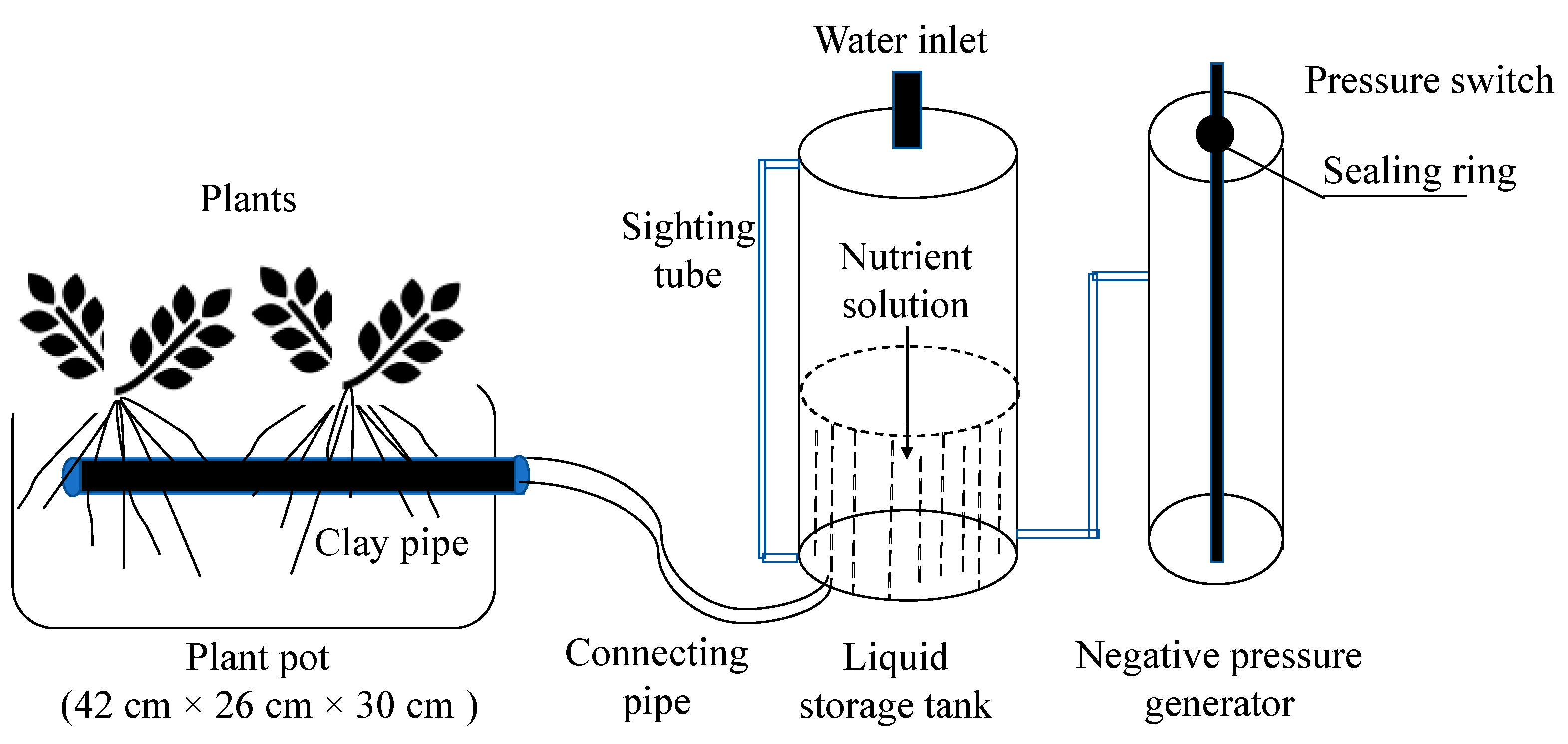
2.1. Materials
2.2. Experimental Design
2.2.1. Experiment 1
2.2.2. Experiment 2
2.3. Analyses
2.4. Statistical Analysis
3. Results
3.1. Performance of the Lettuce in Experiment 1
3.2. Yield and Quality of Lettuce in Experiment 2
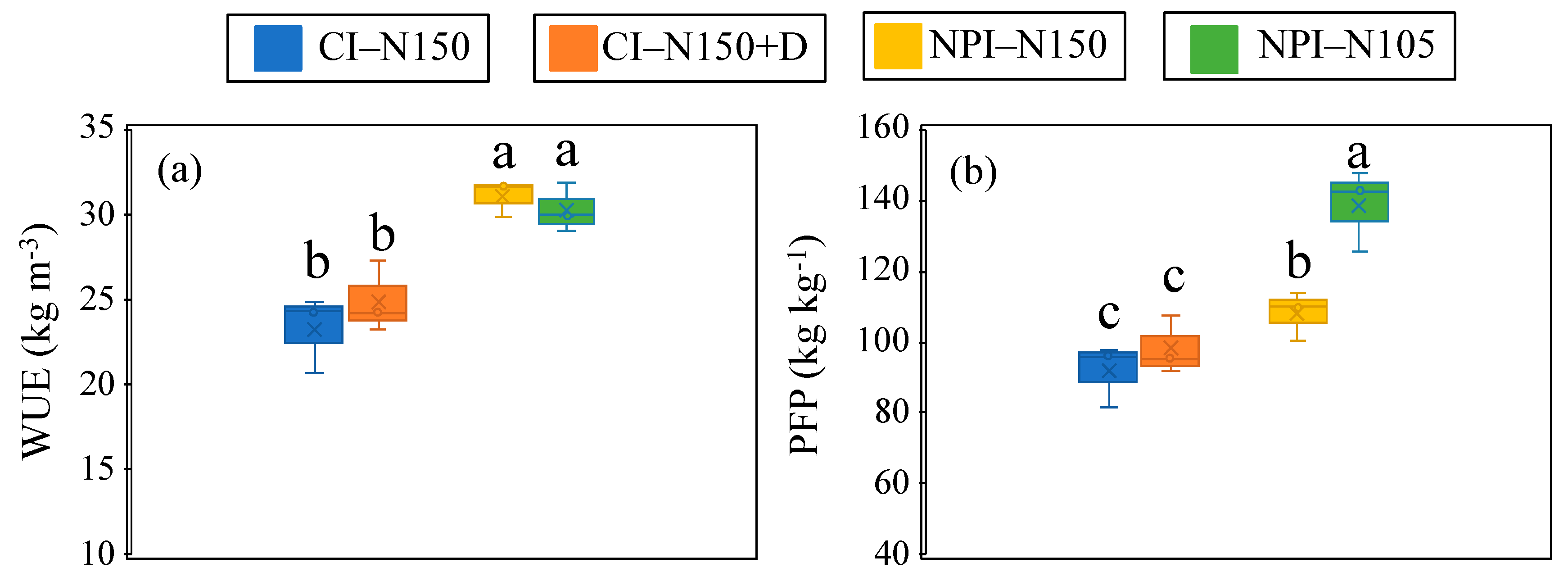
3.3. Water Use Efficiency and Partial Factor Productivity of Applied N
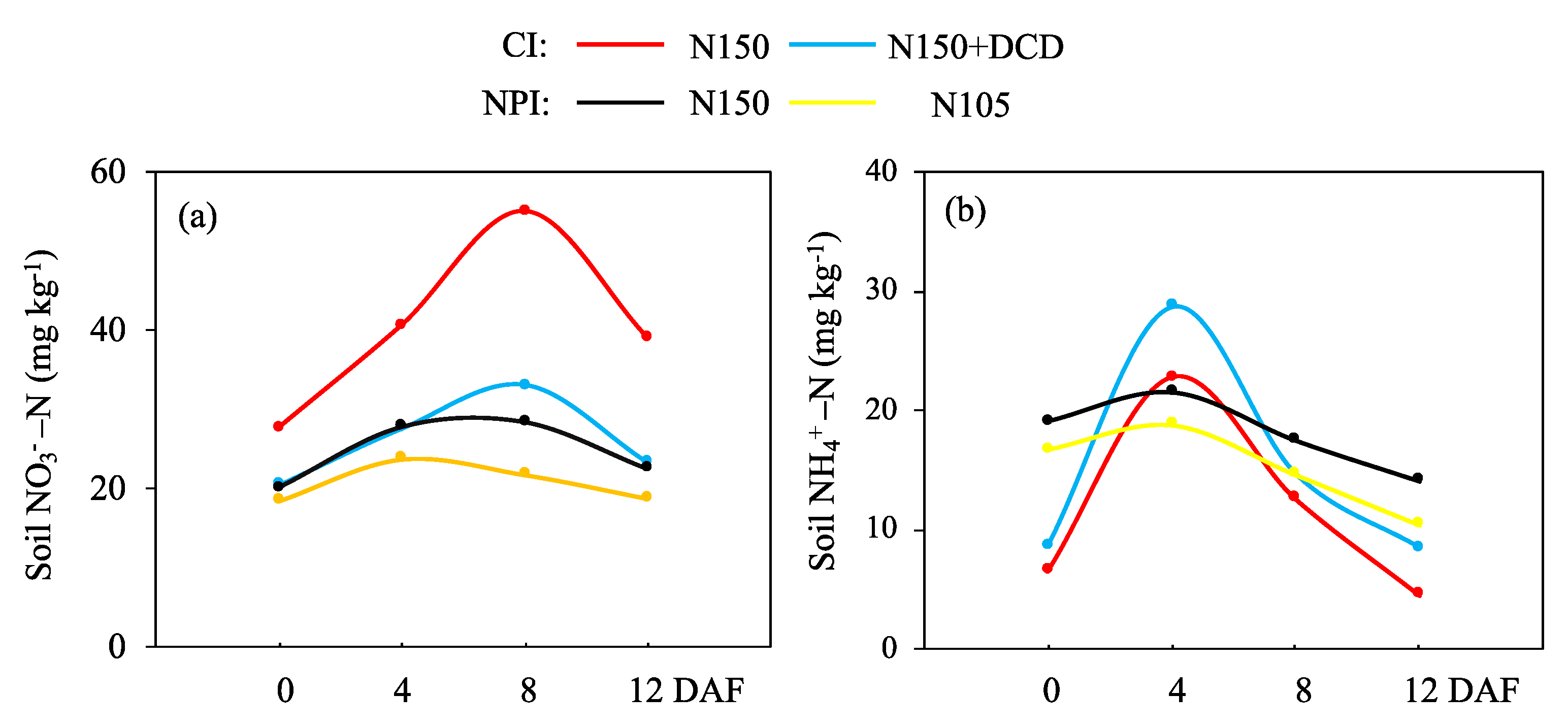
3.4. Dynamic Change of Soil NO3−-N and NH4+-N Content
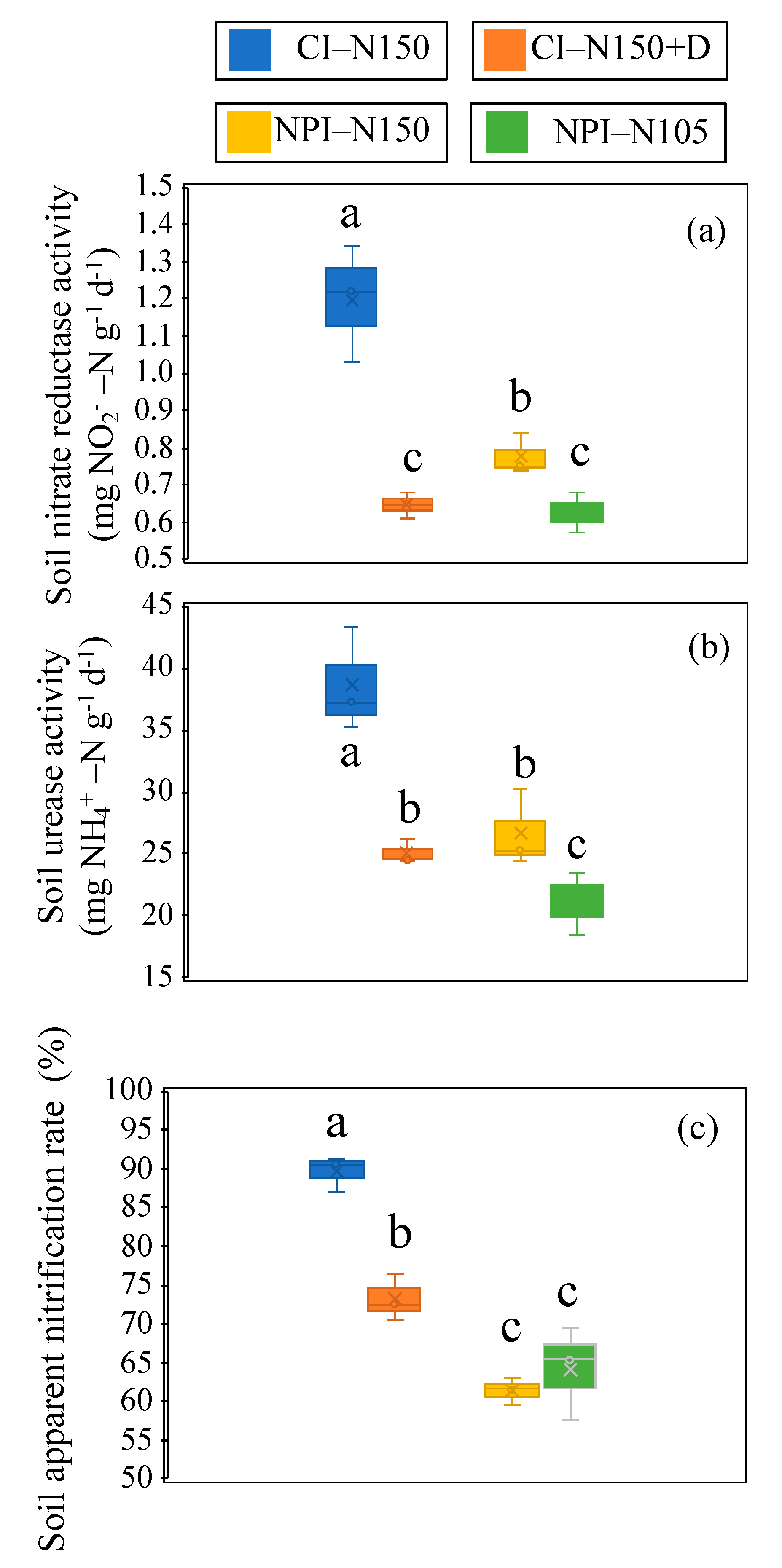
3.5. Soil Apparent Nitrification Rate, Nitrate Reductase and Urease Activities
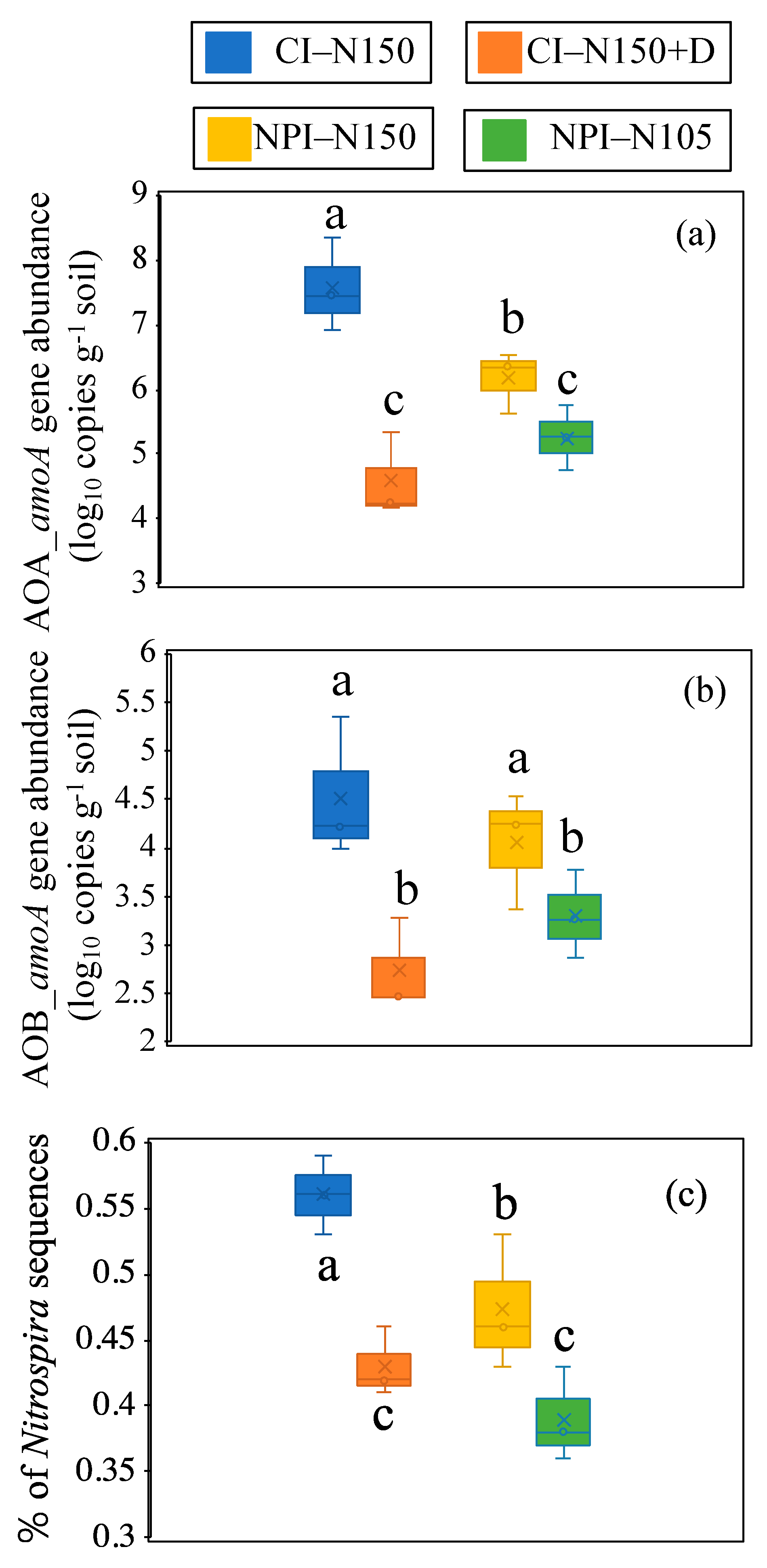
3.6. AOA and AOB Gene, Nitrospira Abundance
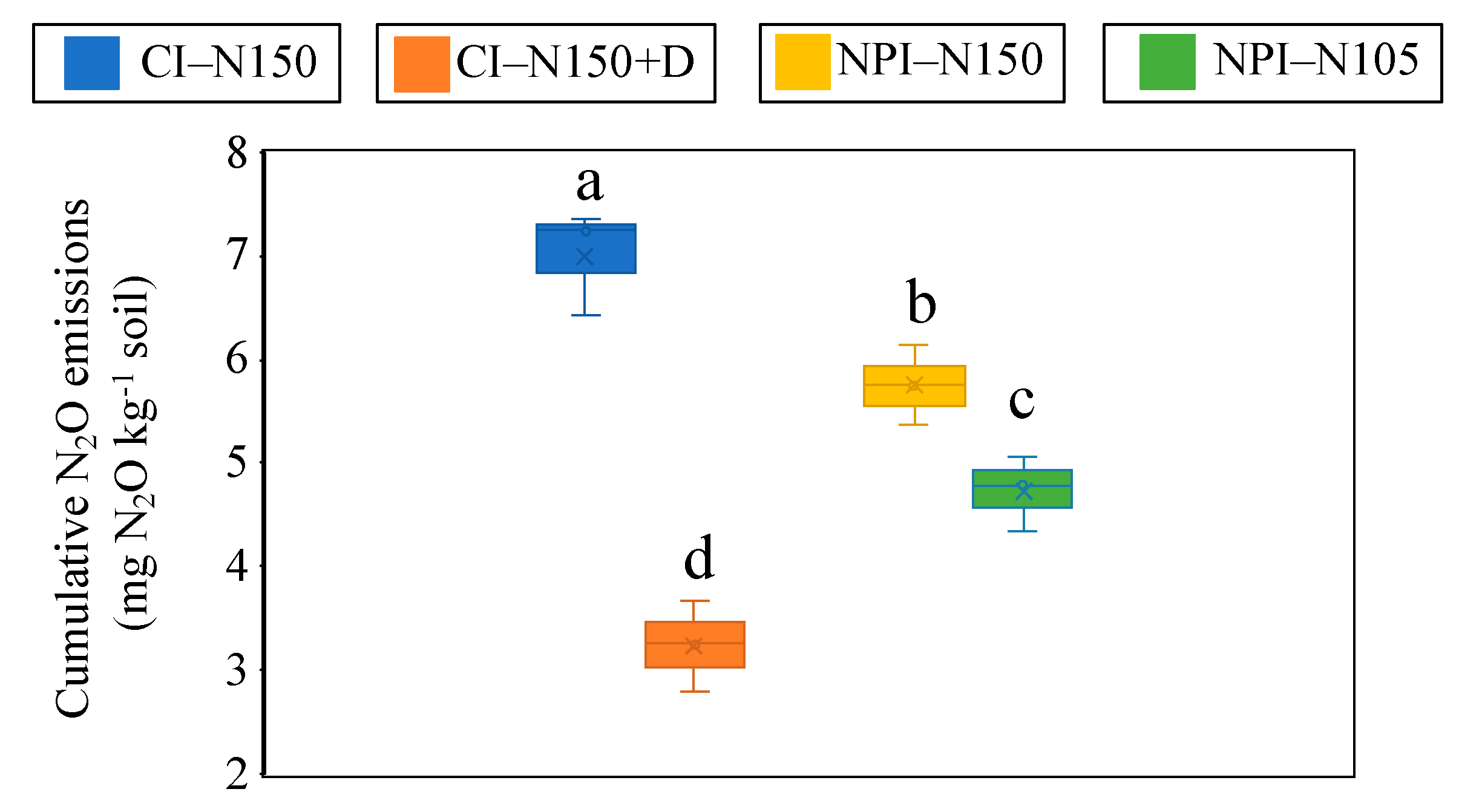
3.7. Cumulative N2O Emissions
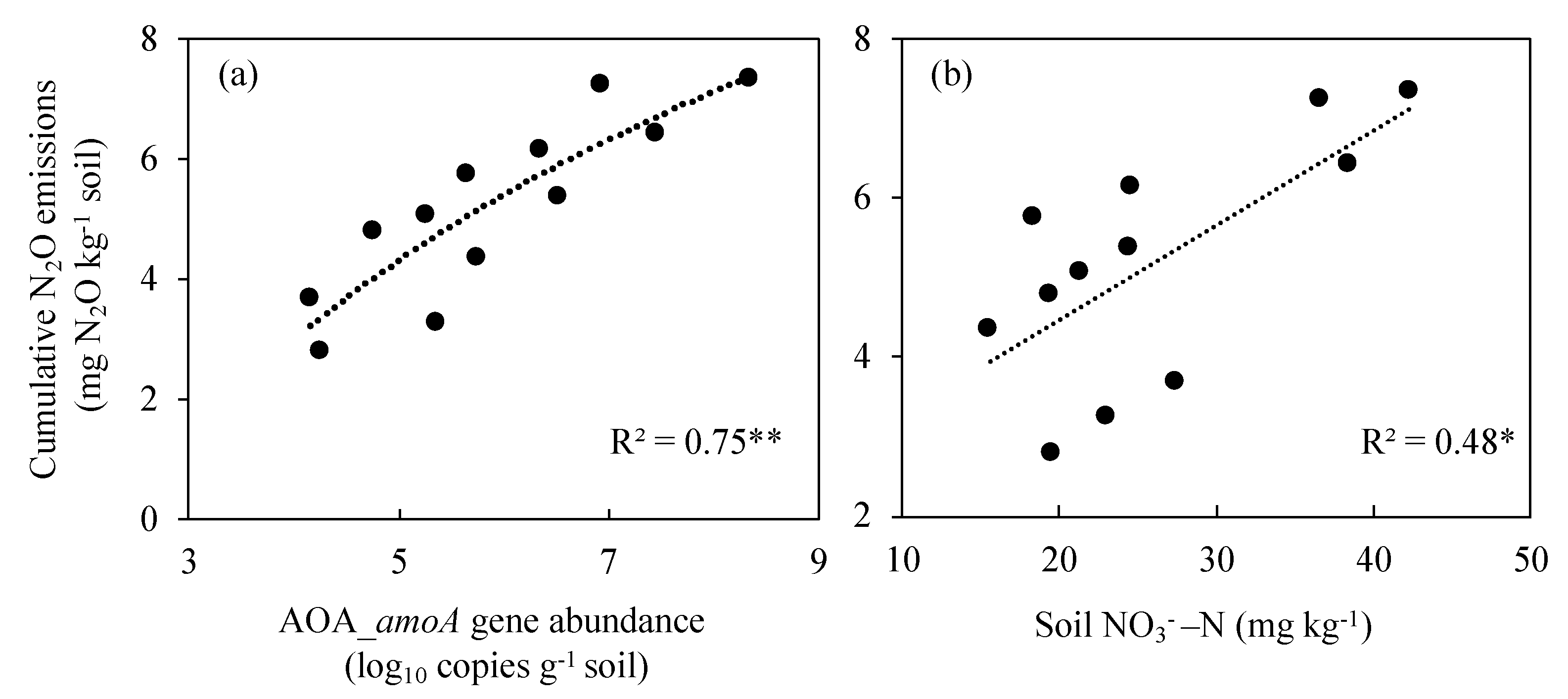
3.8. Relationship between N2O Emissions and AOA and Soil NO3−-N
4. Discussion
4.1. NPI Improved the Performance of Lettuce
4.2. Reduction of the Formation of NO3−-N with NPI
4.3. Mitigation of Nitrifying Activity and N2O Emissions
5. Conclusions
6. Patents
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Long, H.; Zhang, H.; Yue, X.; Zhang, R. Design and experiment of heavy-type negative pressure value used for negative pressure irrigation. Trans. Chin. Soc. Agric. Eng. 2018, 34, 85–92. [Google Scholar]
- Zhao, X.; Gao, X.; Zhang, S.; Long, H. Improving the growth of rapeseed (Brassica chinensis L.) and the composition of rhizosphere bacterial communities through negative pressure irrigation. Water Air Soil Poll. 2019, 239, 9. [Google Scholar] [CrossRef]
- Li, S.; Tan, D.; Wu, X.; Degre, A.; Long, H.; Zhang, S.; Lu, J.; Gao, L.; Zheng, F.; Liu, X.; et al. Negative pressure irrigation increases vegetable water productivity and nitrogen use efficiency by improving soil water and NO3−-N distributions. Agric. Water Manage. 2021, 251, 106853. [Google Scholar] [CrossRef]
- Wang, J.; Huang, Y.; Long, H. Water and salt movement in different soil textures under various negative irrigation pressures. J. Integr. Agric. 2016, 15, 1874–1882. [Google Scholar] [CrossRef] [Green Version]
- Gao, X.; Zhang, S.; Zhao, X.; Long, H. Evaluation of potassium application on tomato performance and rhizosphere bacterial communities under negative pressure irrigation of greenhouse-grown. J. Plant Nutr. 2020, 43, 317–326. [Google Scholar] [CrossRef]
- Long, H.; Wu, X.; Zhang, S.; Wang, J.; Drohan, P.J.; Zhang, R. Connotation and research progress of crop initiate water drawing technology. Trans. Chin. Soc. Agric. Eng. 2020, 36, 139–152. [Google Scholar]
- Gao, X.; Zhang, S.; Zhao, X.; Long, H. Stable water and fertilizer supply by negative pressure irrigation improve tomato production and soil bacterial communities. SN App. Sci. 2019, 1, 178. [Google Scholar] [CrossRef]
- Li, D.; Long, H.Y.; Zhang, S.X.; Wu, X.P.; Shao, H.Y.; Wang, P. Effect of continuous negative pressure water supply on the growth, development and physiological mechanism of Capsicum annuum L. J. Integr. Agric. 2017, 16, 1978–1989. [Google Scholar] [CrossRef]
- Wang, J.; Long, H.; Huang, Y.; Wang, X.; Cai, B.; Liu, W. Effects of different irrigation management parameters on cumulative water supply under negative pressure irrigation. Agric. Water Manage. 2019, 224, 105743. [Google Scholar] [CrossRef]
- Davison, E.A. The contribution of manure and fertilizer nitrogen to atmospheric nitrous oxide since 1860. Nat. Geosci. 2009, 2, 659–662. [Google Scholar] [CrossRef]
- Ding, J.; Fang, F.; Lin, W.; Qiang, X.; Xu, C.; Mao, L.; Li, Q.; Zhang, X.; Li, Y. N2O emissions and source partitioning using stable isotopes under furrow and drip irrigation in vegetable field of North China. Sci. Total Environ. 2019, 665, 709–717. [Google Scholar] [CrossRef]
- Subbarao, G.V.; Searchinger, T.D. Opinion: A “more ammonium solution” to mitigate nitrogen pollution and boost crop yields. Proc. Natl. Acad. Sci. USA 2021, 118, e2107576118. [Google Scholar] [CrossRef] [PubMed]
- Subbarao, G.V.; Arango, J.; Masahiro, K.; Hooper, A.M.; Yoshibashi, T.; Ando, Y.; Nakahara, K.; Deshpande, S.; Ortiz-Monasterio, I.; Ishitani, M.; et al. Genetic mitigation strategies to tackle agricultural GHG emissions: The case for biological nitrification inhibition technology. Plant Sci. 2017, 262, 165–168. [Google Scholar] [CrossRef]
- Qasim, W.; Xia, L.; Lin, S.; Wan, L.; Zhao, Y.; Butterbach-Bahl, K. Global greenhouse vegetable production systems are hotspots of soil N2O emissions and nitrogen leaching: A meta-analysis. Environ. Pollut. 2021, 272, 116371. [Google Scholar] [CrossRef] [PubMed]
- Tan, B.; Li, Y.; Liu, T.; Tan, X.; He, Y.; You, X.; Leong, K.; Liu, C.; Li, L. Response of plant rhizosphere microenvironment to water management in soil-and substrate-based controlled environment agriculture (ECA) systems: A review. Front. Plant Sci. 2021, 12, 691651. [Google Scholar] [CrossRef] [PubMed]
- Farneselli, M.; Benincasa, P.; Tosti, G.; Simonne, E.; Guiducci, M.; Tei, F. High fertigation frequency improves uptake and crop performance in processing tomato grown with high nitrogen and water supply. Agric. Water Manage. 2015, 154, 52–58. [Google Scholar] [CrossRef]
- Bai, X.; Gao, J.; Wang, S.; Cai, H.; Chen, Z.; Zhou, J. Excessive nutrient balance surpluses in newly built solar greenhouses over five years leads to high nutrient accumulations in soil. Agric. Ecosyst. Environ. 2020, 288, 106717. [Google Scholar] [CrossRef]
- Coskun, D.; Britto, D.T.; Shi, W.; Kronzucker, H.J. Nitrogen transformations in modern agriculture and role of biological nitrification inhibition. Nat. Plants 2017, 3, 17074. [Google Scholar] [CrossRef]
- Thompson, R.L.; Lassaletta, L.; Patra, P.K.; Wilson, C.; Wells, K.C.; Gressent, A.; Koffi, E.N.; Chipperfield, M.P.; Winiwarter, W.; Davidson, E.A.; et al. Acceleration of global N2O emissions seen from two decades of atmospheric inversion. Nat. Clim. Change 2019, 9, 993–998. [Google Scholar] [CrossRef]
- Subbarao, G.V.; Yoshihashi, T.; Worthington, M.; Nakahara, K.; Ando, Y.; Sahrawat, K.L.; Rao, I.M.; Lata, J.C.; Kishii, M.; Braun, H.J. Suppression of soil nitrification by plants. Plant Sci. 2015, 233, 155–164. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Dou, Z.; Shi, X.; Zou, C.; Liu, D.; Wang, Z.; Guan, X.; Sun, Y.; Wu, G.; Zhang, B.; et al. Innovative management programme reduces environmental impacts in Chinese vegetable production. Nat. Food 2021, 2, 47–53. [Google Scholar] [CrossRef]
- Zhang, M.; Zeng, H.; Afzal, M.R.; Gao, X.; Li, Y.; Subbarao, G.V.; Zhu, Y. BNI-release mechanisms in plant root system—Current status of understanding. Biol. Fert. Soils 2021. [Google Scholar] [CrossRef]
- Prosser, J.I.; Nicaol, G.W. Archaeal and bacterial ammonia-oxidisers in soil: The quest for niche specialization and differentiation. Trends Microbiol. 2012, 20, 523–531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beeckman, F.; Motte, H.; Beeckman, T. Nitrification in agricultural soils impact, actors and mitigation. Curr. Opin. Biotech. 2018, 50, 166–173. [Google Scholar] [CrossRef]
- Sarr, P.S.; Ando, Y.; Nakamura, S.; Deshpande, S.; Subbarao, G.V. Sorgoleone release from sorghum roots shapes the composition of nitrifying populations, total bacteria, and archaea and determines the level of nitrification. Biol. Fert. Soils 2020, 56, 145–166. [Google Scholar] [CrossRef] [Green Version]
- Afzal, M.R.; Zhang, M.; Jin, H.; Wang, G.; Zhang, M.; Ding, M.; Raza, S.; Hu, J.; Zeng, H.; Gao, X.; et al. Post-translation regulation of plasma membrane H+-ATPase is involved in the release of biological nitrification inhibitors from sorghum roots. Plant Soil 2020, 450, 357–372. [Google Scholar] [CrossRef]
- Min, J.; Sun, H.; Kronzucker, H.; Wang, Y.; Shi, W. Comprehensive assessment of the effects of nitrification inhibitor application on reactive nitrogen loss in intensive vegetable production systems. Agric. Ecosyst. Environ. 2021, 307, 107227. [Google Scholar] [CrossRef]
- Abalos, D.; Sanchez-Martin, L.; Garcia-Torres, L.; von Groenigen, J.W.; Vallejo, A. Management of irrigation frequency and nitrogen fertilization to mitigate GHG and NO emissions from drip-fertigated crops. Sci. Total Environ. 2014, 490, 880–888. [Google Scholar] [CrossRef] [Green Version]
- Liang, H.; Lv, H.; Batchelor, W.; Lian, X.; Wang, Z.; Lin, S.; Hu, K. Simulating nitrate and DON leaching to optimize water and N management practices for greenhouse vegetable production systems. Agric. Water Manage. 2020, 241, 106377. [Google Scholar] [CrossRef]
- Li, Y.K.; Wang, L.C.; Xue, X.Z.; Guo, W.Z.; Xu, F.; Li, Y.L.; Sun, W.T.; Chen, F. Comparison of drip fertigation and negative pressure fertigation on soil water dynamics and water use efficiency of greenhouse tomato grown in the North China Plain. Agric. Water Manage. 2017, 184, 1–8. [Google Scholar] [CrossRef]
- Lv, H.; Lin, S.; Wang, Y.; Lian, X.; Zhao, Y.; Li, Y.; Du, J.; Wang, Z.; Wang, J.; Butterbach-Bahl, K. Drip fertigation significantly reduces nitrogen leaching in solar greenhouse vegetable productions system. Environ. Pollut. 2019, 245, 694–701. [Google Scholar] [CrossRef]
- Yao, Z.; Yan, G.; Wang, R.; Zheng, X.; Liu, C.; Butterbach-bahl, K. Drip irrigation or reduced N-fertilizer rate can mitigate the high annual N2O + NO fluxes from a Chinese intensive vegetable systems. Atmos. Environ. 2019, 212, 183–193. [Google Scholar] [CrossRef]
- Singh, D.K.; Kumar, S. Nitrate reductase, arginine deaminase, urease and dehydrogenase activities in natural soil (ridges with forest) and in cotton soil after acetamiprid treatments. Chemosphere 2008, 71, 412–418. [Google Scholar] [CrossRef] [PubMed]
- Meinhardt, K.A.; Bertagnolli, A.; Pannu, M.W.; Stand, S.E.; Brown, S.L.; Stahl, D.A. Evaluation of revised polymerase chain reaction primers for more inclusive quantification of ammonia-oxidizing archaea and bacteria. Environ. Microbiol. Rep. 2015, 7, 354–363. [Google Scholar] [CrossRef] [PubMed]
- Hink, L.; Nicol, G.W.; Prosser, J. Archaea produce lower yield of N2O than bacteria during aerobic ammonia oxidation in soil. Environ. Microbiol. 2017, 19, 4829–4837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, P.; Drohan, P.; Long, H.; Bian, Y.; Bryant, R. Negative pressure irrigation on water use efficiency, yield and quality of B. chinensis L. J. Sci. Food Agric. 2021. [Google Scholar] [CrossRef] [PubMed]
- Fan, Z.; Lin, S.; Zhang, X.; Jiang, Z.; Yang, K.; Jian, D.; Chen, Y.; Li, J.; Chen, Q.; Wang, J. Conventional flooding irrigation causes an overuse of nitrogen fertilizer and low nitrogen use efficiency in intensively used solar greenhouse vegetable production. Agric. Water Manage. 2014, 144, 11–19. [Google Scholar] [CrossRef]
- Abel, A.; Mattes, R. Simulation of the dry matter production and seed yield of common beans under varying soil water and salinity conditions. Agric. Water Manage. 2001, 47, 55–68. [Google Scholar] [CrossRef]
- Patane, C.; Tringali, S.; Sortino, O. Effects of soil water on yield and quality of processing tomato under a Mediterranean climate conditions. Agric. Water Manage. 2010, 97, 131–138. [Google Scholar] [CrossRef]
- Wang, C.; Gu, F.; Chen, J.; Yang, H.; Jiang, J.; Du, T.; Zhang, J. Assessing the response of yield and comprehensive fruit quality of tomato grown in greenhouse to deficit irrigation and nitrogen application strategies. Agric. Water Manage. 2015, 161, 9–19. [Google Scholar] [CrossRef]
- Subbarao, G.V.; Kishii, M.; Bozal-Leorri, A.; Ortiz-Monasterio, I.; Gao, X.; Ibba, M.I.; Karwat, H.; Gonzalez-Moro, M.B.; Gonzalez-Murua, C.; Yoshihashi, T.; et al. Enlisting wild grass genes to combat nitrification in wheat farming: A nature-based solution. Proc. Natl. Acad. Sci. USA 2021, 118, e2106595118. [Google Scholar] [CrossRef]
- Fu, Q.; Abadie, M.; Blaud, A.; Carswell, A.; Misselbrook, T.; Clark, I.; Hirsch, P. Effects of urease and nitrification inhibitors on soil N, nitrifier abundance and activity in a sandy loam soil. Biol. Fert. Soils 2020, 25, 185–194. [Google Scholar] [CrossRef] [Green Version]
- Guo, Y.; Chen, Y.; Searchinger, T.; Zhou, M.; Pan, D.; Yang, J.; Wu, L.; Cui, Z.; Zhang, W.; Zhang, F.; et al. Air quality, nitrogen use efficiency and food security in China are improved by cost-effective agricultural nitrogen management. Nat. Food 2020, 1, 648–658. [Google Scholar] [CrossRef]
- Ganeteg, U. Uptake of organic nitrogen by plants. New Phytol. 2009, 182, 31–48. [Google Scholar] [CrossRef]
- Murphy, D.V.; Macdonald, A.J.; Stockdale, E.A.; Goulding, K.W.T.; Fortune, S.; Gaunt, J.L.; Poulton, P.R.; Wakefield, J.A. Soluble organic nitrogen in agricultural soils. Biol. Fert. Soils 2020, 30, 374–387. [Google Scholar] [CrossRef]
- Sun, Y.; Hu, K.; Zhang, K.; Jiang, L.; Xu, Y. Simulation of nitrogen fate for greenhouse cucumber grown under different water and fertilizer management using the EU-Rotate N model. Agric. Water Manage. 2012, 112, 21–32. [Google Scholar] [CrossRef]
- Hink, L.; Gubry-Rangin, C.; Nicol, G.W.; Prosser, J. The consequences of niche and physiological differentiation of archaeal and bacterial ammonia oxidisers for nitrous oxide emissions. ISME J. 2018, 12, 1084–1093. [Google Scholar] [CrossRef] [PubMed]
- Tosi, M.; Brown, S.; Machado, P.; Wagner-Riddle, C.; Dunfield, K. Short-term response of soil N-cycling genes and transcripts to fertilization with nitrification and urease inhibitors, and relationship with field-scale N2O emissions. Soil Biol. Biochem. 2020, 142, 107703. [Google Scholar] [CrossRef]
- Zhao, Y.; Lv, H.; Qasim, W.; Wan, L.; Wang, Y.; Lian, X.; Liu, Y.; Hu, J.; Wang, Z.; Li, G.; et al. Drip fertigation with straw incorporation significantly reduces N2O emission and N leaching while maintaining high vegetable yields in solar greenhouse production. Environ. Pollut. 2021, 273, 116521. [Google Scholar] [CrossRef]
- Stevens, R.J.; Laughlin, R.J. Measurement of nitrous oxide and dinitrogen emissions from agricultural soils. Nut. Cycl. Agroecosystems 1998, 52, 131–139. [Google Scholar] [CrossRef]
- Wang, X.; Bai, J.; Xie, T.; Wang, W.; Zhang, G.; Yin, S.; Wang, D. Effects of biological nitrification inhibitors on nitrogen use efficiency and greenhouse gas emissions in agricultural soils: A review. Ecotoxicol. Environ. Saf. 2021, 220, 112338. [Google Scholar] [CrossRef] [PubMed]
- Prosser, J.I.; Hink, L.; Rangin, C.G.; Nicol, G.W. Nitrous oxide production by ammonia oxidizers: Physiological diversity, niche differentiation and potential mitigation strategies. Glob. Change Biol. 2020, 26, 103–118. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Irrigation | N | Yield | Total N Content | Nitrate | WUE | PFP |
|---|---|---|---|---|---|---|
| Treatments | g pot−1 | g pot−1 | mg kg−1 | kg m−3 | Kg Kg−1 | |
| CI | N150 | 361 ± 16 b | 17.7 ± 0.9 b | 508 ± 12 a | 22.6 ± 1.0 b | 89 ± 4 d |
| N120 | 338 ± 20 bc | 14.5 ± 0.3 d | 485 ± 30 a | 21.1 ± 1.3 b | 104 ± 6 c | |
| N105 | 288 ± 20 de | 12.0 ± 0.8 e | 417 ± 24 b | 18.3 ± 1.2 c | 101 ± 7 c | |
| N90 | 263 ± 19 e | 10.3 ± 0.4 f | 391 ± 25 b | 16.4 ± 1.2 c | 108 ± 8 c | |
| NPI | N150 | 421 ± 12 a | 20.5 ± 0.8 a | 403 ± 17 b | 29.9 ± 1.2 a | 104 ± 3 c |
| N120 | 410 ± 18 a | 19.6 ± 0.7 a | 381 ± 15 b | 30.3 ± 1.8 a | 126 ± 6 b | |
| N105 | 403 ± 14 a | 18.9 ± 0.9 ab | 346 ± 14 c | 29.6 ± 0.6 a | 142 ± 5 a | |
| N90 | 327 ± 25 bcd | 16.4 ± 0.4 c | 319 ± 17 c | 26.9 ± 2.3 a | 135 ± 10 ab |
| Irrigation | N | Yield | Plant N | Nitrate | Soluble Sugar | VC |
|---|---|---|---|---|---|---|
| Treatments | g pot−1 | g pot−1 | mg kg−1 | mg g−1 | mg kg−1 | |
| CI | N150 | 373 ± 21 c | 18.8 ± 1.1 c | 493 ± 21 a | 25.8 ± 1.2 b | 217 ± 9 b |
| N150 + D | 399 ± 20 bc | 20.6 ± 1.1 b | 387 ± 14 b | 24.9 ± 0.9 b | 214 ± 6 b | |
| NPI | N150 | 439 ± 16 a | 23.4 ± 1.1 a | 394 ± 36 b | 27.2 ± 1.0 a | 233 ± 5 a |
| N105 | 412 ± 15 ab | 21.3 ± 1.0 b | 352 ± 20 c | 27.9 ± 0.3 a | 238 ± 8 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gao, X.; Zhang, S.; Song, Y.; Long, H. Negative Pressure Irrigation System Reduces Soil Nitrogen Loss for Lettuce during Greenhouse Production. Agronomy 2021, 11, 2380. https://doi.org/10.3390/agronomy11122380
Gao X, Zhang S, Song Y, Long H. Negative Pressure Irrigation System Reduces Soil Nitrogen Loss for Lettuce during Greenhouse Production. Agronomy. 2021; 11(12):2380. https://doi.org/10.3390/agronomy11122380
Chicago/Turabian StyleGao, Xiang, Shuxiang Zhang, Yanyan Song, and Huaiyu Long. 2021. "Negative Pressure Irrigation System Reduces Soil Nitrogen Loss for Lettuce during Greenhouse Production" Agronomy 11, no. 12: 2380. https://doi.org/10.3390/agronomy11122380
APA StyleGao, X., Zhang, S., Song, Y., & Long, H. (2021). Negative Pressure Irrigation System Reduces Soil Nitrogen Loss for Lettuce during Greenhouse Production. Agronomy, 11(12), 2380. https://doi.org/10.3390/agronomy11122380