
Repeated Manure Application for Eleven Years Stimulates Enzymatic Activities and Improves Soil Attributes in a Typic Hapludalf

 ,
,  , ,
, ,  , ,
, ,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
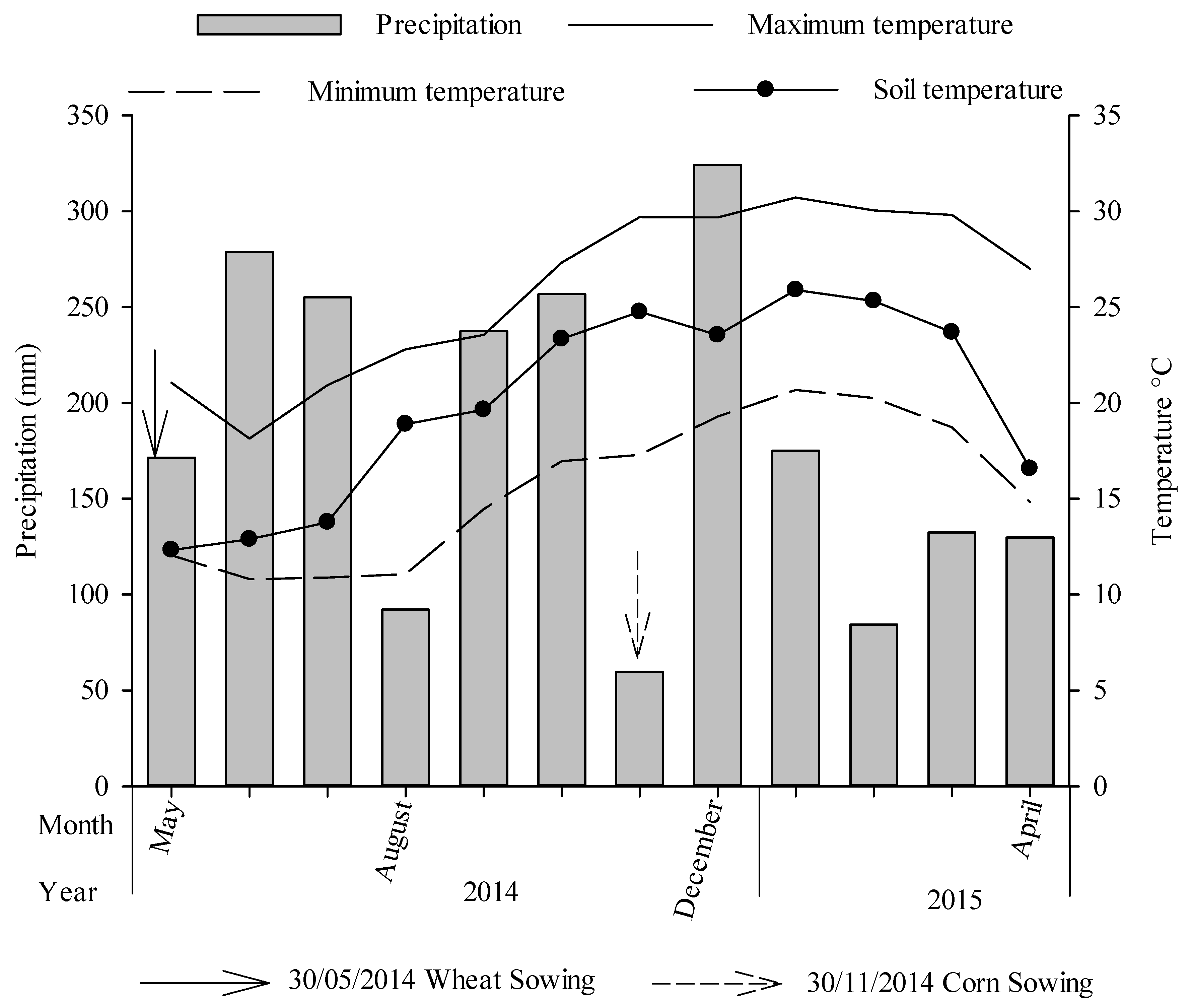
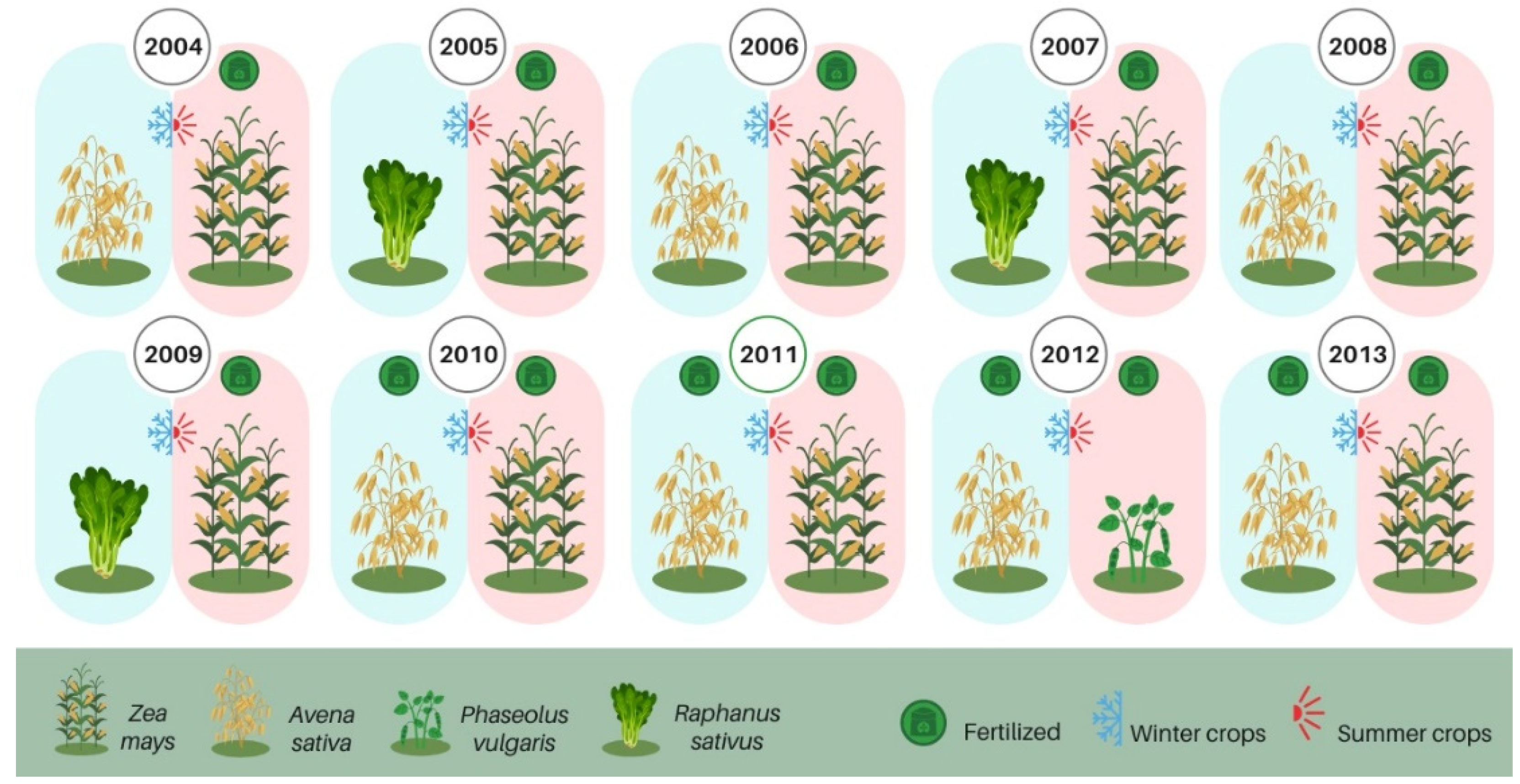
2.1. Long-Term Field Experiment
2.2. Soil and Plant Tissue Analyses and Grain Yield
2.3. Statistical Analysis
3. Results
3.1. Chemical and Physical Properties of the Soil
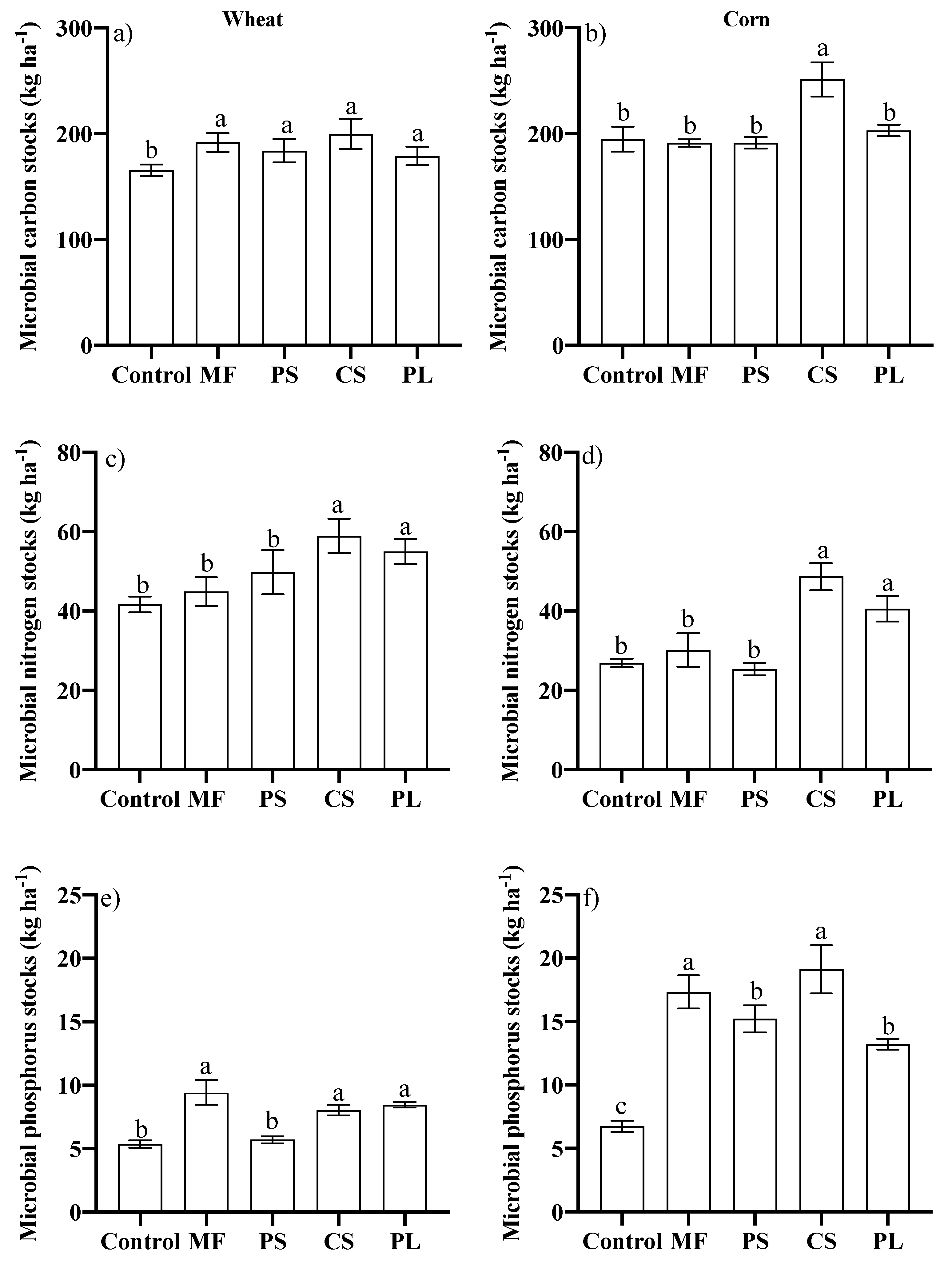
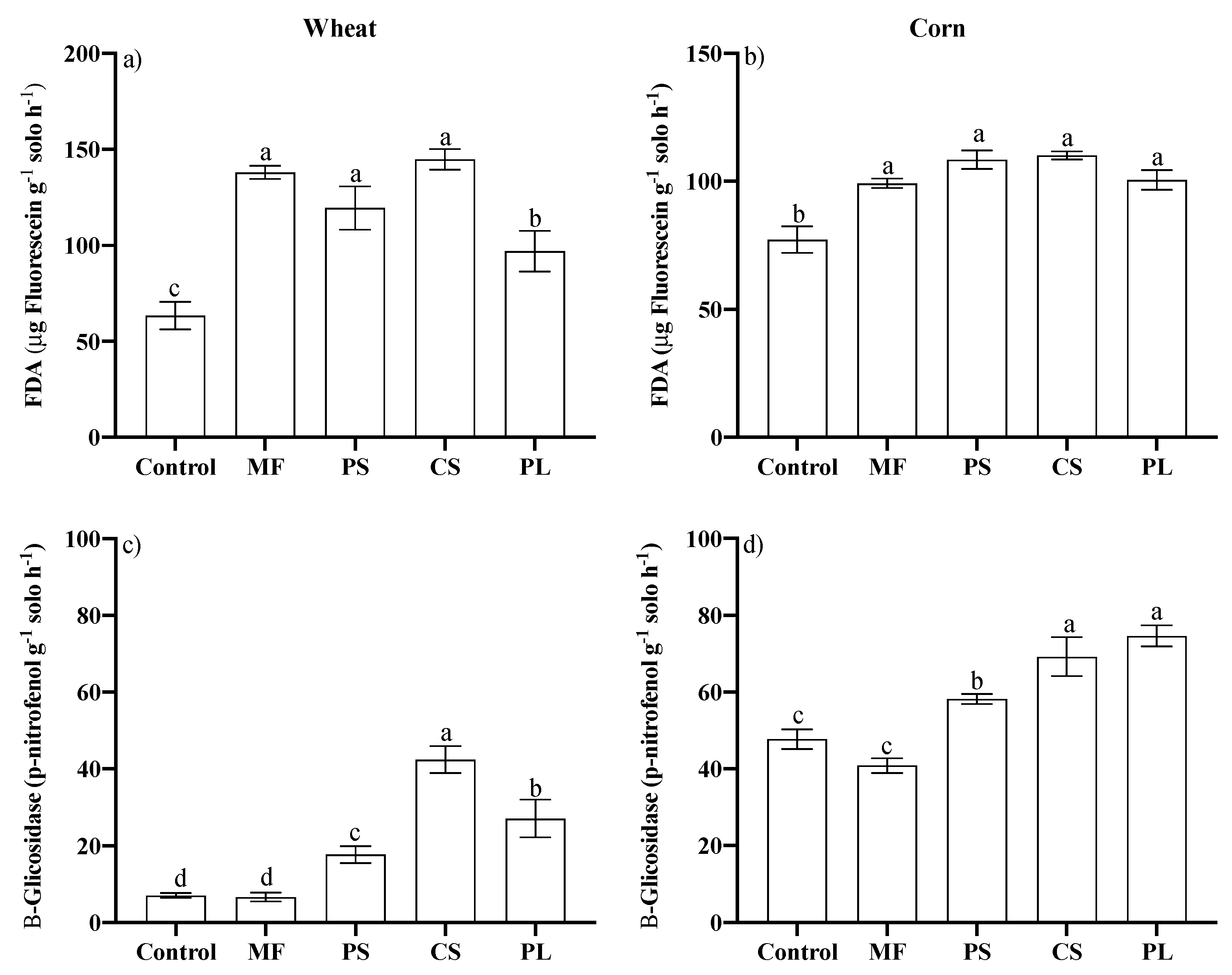
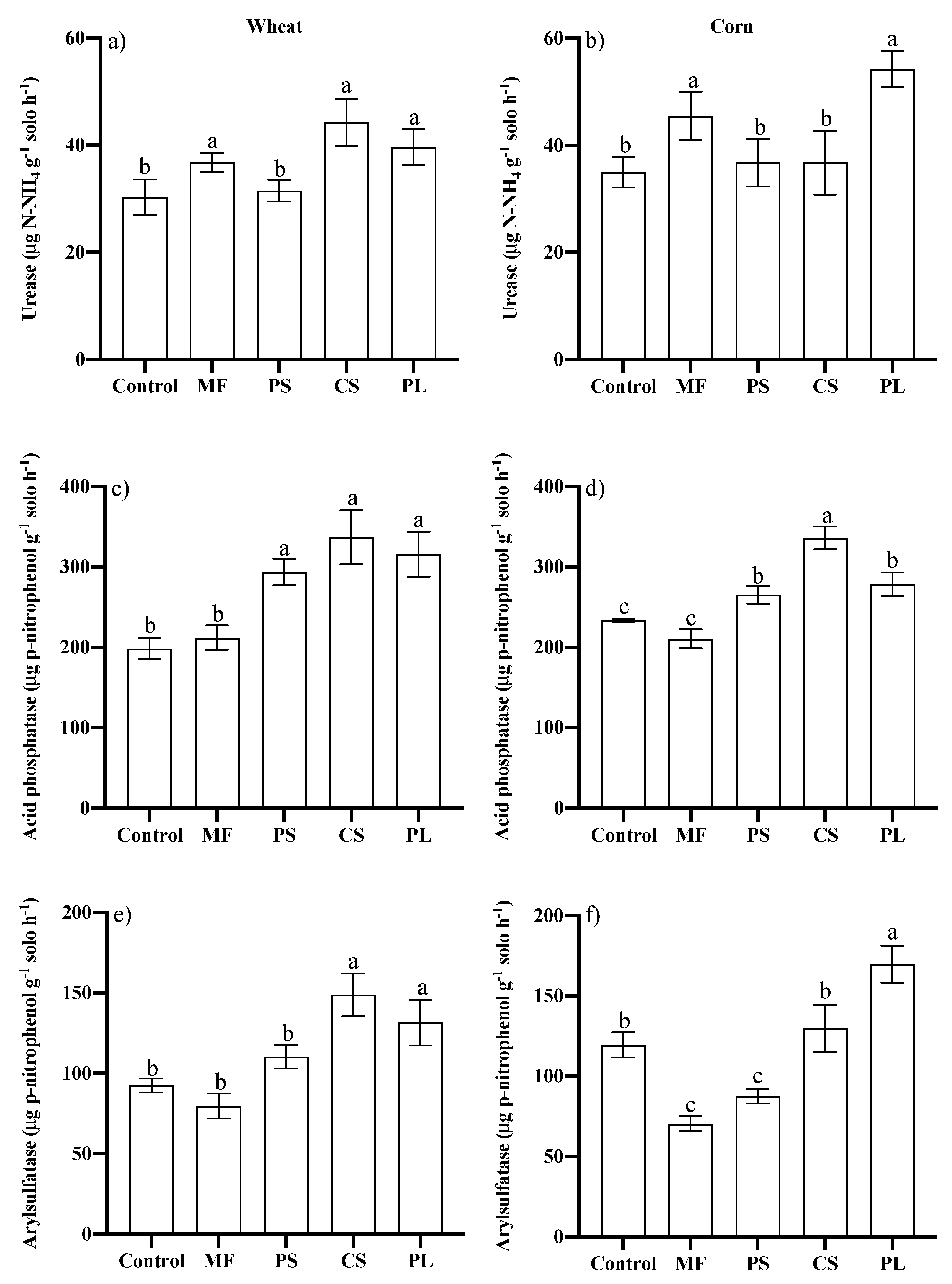
3.2. Microbiological Attributes of the Soil
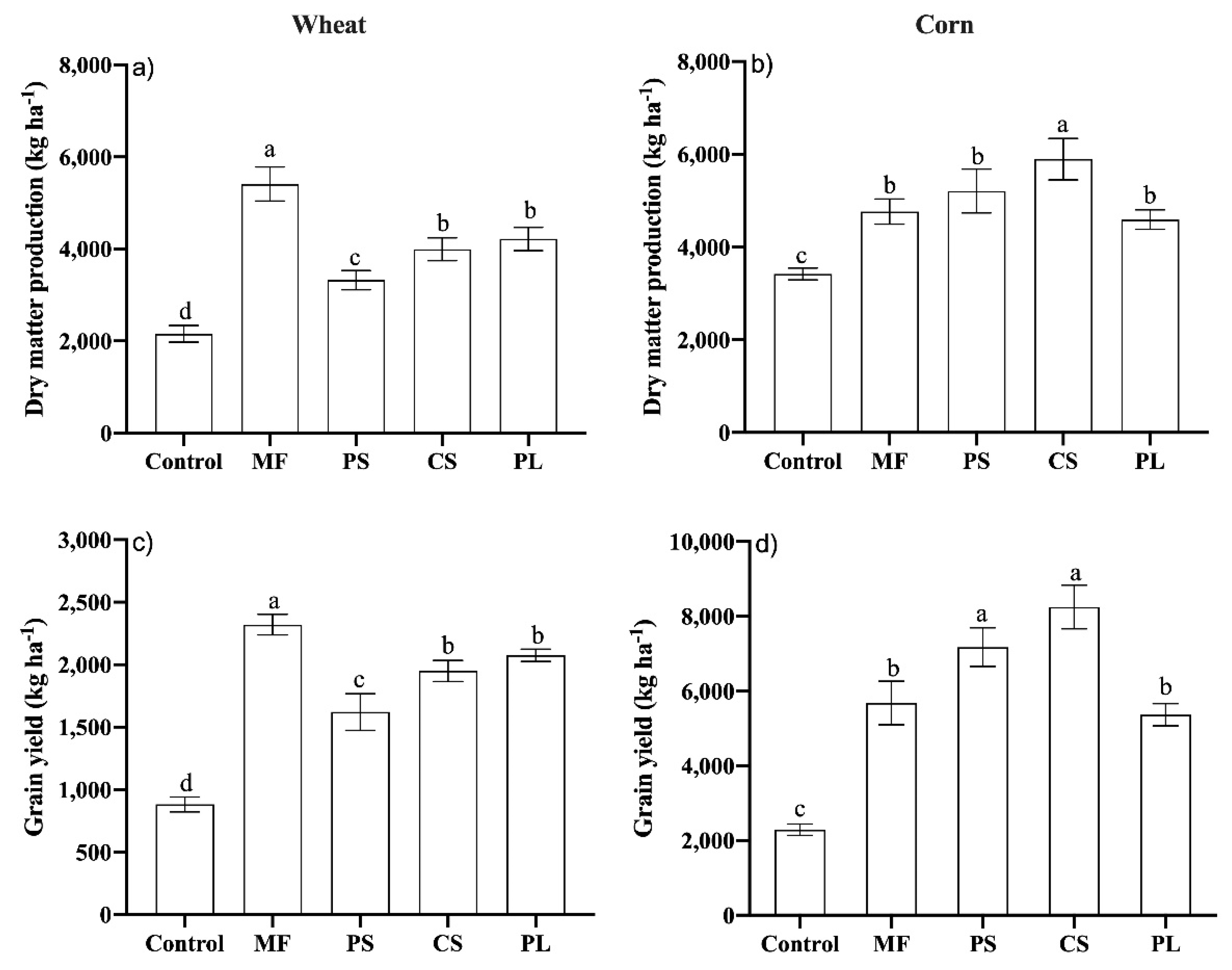
3.3. Dry Matter and Grain Yield of Wheat and Maize
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Zhang, X.; Fang, Q.; Zhang, T.; Ma, W.; Velthof, G.L.; Hou, Y.; Oenema, O.; Zhang, F. Benefits and trade-offs of replacing synthetic fertilizers by animal manures in crop production in China: A meta-analysis. Glob. Chang. Biol. 2020, 26, 888–900. [Google Scholar] [CrossRef] [PubMed]
- FAOSTAT-2021. Live Animals. Available online: http://www.fao.org/faostat/en/#data (accessed on 23 May 2021).
- Font-Palma, C. Methods for the treatment of cattle manure—A review. J. Carbon Res. 2019, 5, 27. [Google Scholar] [CrossRef] [Green Version]
- Losinger, W.C.; Sampath, R.K. Economies of scale in the production of swine manure. Arq. Bras. Med. Vet. E Zootec. 2000, 52, 285–294. [Google Scholar] [CrossRef]
- Bogaard, R.; Fraser, T.H.E.; Heaton, M.; Wallace, P.; Vaiglova, M.; Charles, G.; Jones, R.P.; Evershed, A.K.; Styring, N.H.; Andersen, R.M.; et al. Crop manuring and intensive land management by Europe’s first farmers. Proc. Natl. Acad. Sci. USA 2013, 110, 12589–12594. [Google Scholar] [CrossRef] [Green Version]
- Singh, B. Are nitrogen fertilizers deleterious to soil health? Agronomy 2018, 8, 48. [Google Scholar] [CrossRef] [Green Version]
- Nannipieri, P.; Trasar-Cepeda, C.; Dick, R.P. Soil enzyme activity: A brief history and biochemistry as a basis for appropriate interpretations and meta-analysis. Biol. Fertil. Soils 2018, 54, 11–19. [Google Scholar] [CrossRef]
- Meng, Q.; Ma, X.; Zhang, J.; Yu, Z. The long-term effects of cattle manure application to agricultural soils as a natural-based solution to combat salinization. Catena 2019, 175, 193–202. [Google Scholar] [CrossRef]
- Ren, F.; Sun, N.; Xu, M.; Zhang, X.; Wu, L.; Xu, M. Changes in soil microbial biomass with manure application in cropping systems: A meta-analysis. Soil Tillage Res. 2019, 194, 104291. [Google Scholar] [CrossRef] [Green Version]
- Du, Y.; Cui, B.; Zang, Q.; Wang, Z.; Sun, J.; Niu, W. Effects of manure fertilizer on crop yield and soil properties in China: A meta-analysis. Catena 2020, 193, 104617. [Google Scholar] [CrossRef]
- Liu, S.; Wang, J.; Pu, S.; Blagodatskaya, E.; Kuzyakov, Y.; Razavi, B.S. Impact of manure on soil biochemical properties: A global synthesis. Sci. Total Environ. 2020, 745, 141003. [Google Scholar] [CrossRef] [PubMed]
- Rayne, N.; Aula, L. Livestock manure and the impacts on soil health: A review. Soil Syst. 2020, 4, 64. [Google Scholar] [CrossRef]
- Ma, Q.; Wen, Y.; Wang, D.; Sun, X.; Hill, P.W.; Macdonald, A.; Chadwick, D.R.; Wu, L.; Jones, D.L. Farmyard manure applications stimulate soil carbon and nitrogen cycling by boosting microbial biomass rather than changing its community composition. Soil Biol. Biochem. 2020, 144, 107760. [Google Scholar] [CrossRef]
- Röös, E.; Mie, A.; Wivstad, M.; Salomon, E.; Johansson, B.; Gunnarsson, S.; Wallenbeck, A.; Hoffmann, R.; Nilsson, U.; Sundberg, C.; et al. Risks and opportunities of increasing yields in organic farming: A review. Agron. Sustain. Dev. 2018, 38, 14. [Google Scholar] [CrossRef] [Green Version]
- Benedet, L.; Ferreira, G.W.; Brunetto, G.; Loss, A.; Lovato, P.E.; Lourenzi, C.R.; Silva, S.H.G.; Curi, N.; Comin, J.J. Use of swine manure in agriculture in southern brazil: Fertility or potential contamination? In Soil Contamination-Threats and Sustainable Solutions; Larramendy, M.L., Soloneski, S., Eds.; IntechOpen: London, UK, 2020. [Google Scholar] [CrossRef]
- Tavares, R.L.M.; Assis, R.L.; Ferreira, R.V.; Menezes, J.F.S.; Simon, G.A.; Boldrin, P.F.; Cantão, V.C.G. Long term application of pig manure on the chemical and physical properties of Brazilian Cerrado soil. Carbon Manag. 2019, 10, 1–9. [Google Scholar] [CrossRef]
- Leip, A.; Ledgard, S.; Uwizeye, A.; Palhares, J.C.P.; Aller, M.F.; Amon, B.; Binder, M.; Cordovil, C.M.S.; De Camillis, C.; Dong, H.; et al. The value of manure—Manure as co-product in life cycle assessment. J. Environ. Manag. 2019, 241, 293–304. [Google Scholar] [CrossRef]
- Barth, G.; Gotz, L.F.; Favaretto, N.; Pauletti, V. Does dairy liquid manure complementary to mineral fertilization increase grain yield due to changes in soil fertility? Braz. Arch. Biol. Technol. 2020, 63, e20190537. [Google Scholar] [CrossRef]
- Bhunia, S.; Bhowmik, A.; Mallick, R.; Mukherjee, J. Agronomic efficiency of animal-derived organic fertilizers and their effects on biology and fertility of soil: A review. Agronomy 2021, 11, 823. [Google Scholar] [CrossRef]
- Heuck, C.; Weig, A.; Spohn, M. Soil microbial biomass C:N:P stoichiometry and microbial use of organic phosphorus. Soil Biol. Biochem. 2015, 85, 119–129. [Google Scholar] [CrossRef]
- Lazeris, T.S.; Souza, J.P.; Vezzani, F.M.; Matos, C.L.; Kaschuk, G. Carbon and phosphorus biogeochemical cycles in native forest and horticultural areas in the Metropolitan Region of Curitiba, Brazil. Colloq. Agrar. 2021, 17, 1–11. [Google Scholar] [CrossRef]
- Lupwayi, N.Z.; Zhang, Y.; Hao, X.; Ben, W.; Thomas, B.W.; Eastman, A.H.; Schwinghamer, T.D. Linking soil microbial biomass and enzyme activities to long-term manure applications and their nonlinear legacy. Pedobiologia 2019, 74, 34–42. [Google Scholar] [CrossRef]
- Lin, Y.; Ye, G.; Kuzyakov, Y.; Liu, D.; Fan, J.; Ding, W. Long-term manure application increases soil organic matter and aggregation, and alters microbial community structure and keystone taxa. Soil Biol. Biochem. 2019, 134, 187–196. [Google Scholar] [CrossRef]
- Herrero, M.A.; Palhares, J.C.P.; Salazar, F.J.; Charlón, V.; Tieri, M.P.; Pereyra, A.M. Dairy manure management perceptions and needs in South American Countries. Front. Sustain. Food Syst. 2018, 2, 22. [Google Scholar] [CrossRef]
- Tan, M.; Hou, Y.; Zhang, L.; Shi, S.; Long, W.; Ma, Y.; Oenema, O. Operational costs and neglect of end-users are the main barriers to improving manure treatment in intensive livestock farms. J. Clean. Prod. 2020, 289, 125149. [Google Scholar] [CrossRef]
- EMBRAPA. Sistema Brasileiro de Classificação de Solos—Humberto Gonçalves dos Santos, 3rd ed.; Embrapa: Brasília, Brazil, 2013; 353p. [Google Scholar]
- Comissao de Química e Fertilidade do Solo. Manual de Adubação e Calagem para os Estados do Rio Grande do Sul e de Santa Catarina, 10th ed.; SBCS/NRS/UFRGS: Porto Alegre, Brazil, 2004. [Google Scholar]
- Vance, E.D.; Brookes, P.C.; Jenkinson, D.S. An extraction method for measuring soil microbial biomass. Soil Biol. Biochem. 1987, 19, 703–707. [Google Scholar] [CrossRef]
- Brookes, P.C.; Powlson, D.S.; Jenkinson, D.S. Measurement of microbial biomass phosphorus in soil. Soil Biol. Biochem. 1982, 14, 319–329. [Google Scholar] [CrossRef]
- Anderson, J.P.; Domsch, K.H. The metabolic quotient of CO2 (q CO2) as a specific activity paramenter to assess the effects of environmental condition, such as pH, on the microbial of forest soil. Soil Biol. Biochem. 1993, 25, 393–395. [Google Scholar] [CrossRef]
- Green, V.S.; Stott, D.E.; Miack, M. Assay for fluorescein diacetate hydrolytic activity: Optimization for soil samples. Soil Biol. Biochem. 2006, 38, 693–701. [Google Scholar] [CrossRef]
- Verchot, L.V.; Borelli, T. Application of para-nitrophenol (pNP) enzyme assays in degraded tropical soils. Soil Biol. Biochem. 2005, 37, 625–633. [Google Scholar] [CrossRef]
- EMBRAPA. Manual de Métodos de Análise de Solo, 2nd ed.; Atual: Rio de Janeiro, Brazil, 1997; 212p. [Google Scholar]
- Ferreira, D.F. Sisvar: Um programa para análises e ensino de estatística. Rev. Symp. 2008, 6, 36–41. [Google Scholar]
- Fageria, N.K.; Nascente, A.S. Management of soil acidity of South American soils for sustainable crop production. Adv. Agron. 2014, 128, 221–275. [Google Scholar] [CrossRef]
- Jing, Z.; Min, L.; Mick, W. Phosphorus activators contribute to legacy phosphorus availability in agricultural soils: A review. Sci. Total Environ. 2018, 612, 522–537. [Google Scholar] [CrossRef] [Green Version]
- Abboud, F.Y.; Favaretto, N.; Motta, A.C.V.; Barth, G.; Goularte, G.D. Phosphorus mobility and degree of saturation in oxisol under no-tillage after long-term dairy liquid manure application. Soil Tillage Res. 2018, 177, 45–53. [Google Scholar] [CrossRef]
- Bousfield, S.W.; Favaretto, N.; Motta, A.C.V.; Barth, G.; Celante, L.S.; Cherobim, V.F. Environmental soil phosphorus threshold under no-tillage and swine manure application. Braz. Arch. Biol. Technol. 2020, 63. [Google Scholar] [CrossRef]
- Qin, X.; Guo, S.; Zhai, L.; Pan, J.; Khoshnevisan, B.; Wu, S.; Liu, H. How long-term excessive manure application affects soil phosphorous species and risk of phosphorous loss in fluvo-aquic soil. Environ. Pollut. 2020, 266, 115304. [Google Scholar] [CrossRef]
- Lourenzi, C.R.; Ceretta, C.A.; Silva, L.S.; Girotto, E.; Lorensini, F.; Tiecher, T.L.; De Conti, L.; Trentin, G.; Brunetto, G. Nutrients in layers of soil under no-tillage treated with successive applications of pig slurry. Rev. Bras. Ciênc. Do Solo 2013, 37, 157–167. [Google Scholar] [CrossRef]
- Lourenzi, C.R.; Ceretta, C.A.; Ciancio, N.H.R.; Tiecher, T.L.; da Silva, L.O.S.; De Conti, L.; Girotto, E.; Ferreira, P.A.A.; Vidal, R.F.; Scopel, G.; et al. Forms of nitrogen and phosphorus transfer by runoff in soil under no-tillage with successive organic waste and mineral fertilizers applications. Agric. Water Manag. 2021, 248, 106779. [Google Scholar] [CrossRef]
- Kaschuk, G.; Alberton, O.; Hungria, M. Three decades of soil microbial biomass studies in Brazilian ecosystems: Lessons learned about soil quality and indications for improving sustainability. Soil Biol. Biochem. 2010, 42, 1–13. [Google Scholar] [CrossRef]
- Lopes, A.A.C.; de Sousa, D.M.G.; Chaer, G.M.; dos Reis-Junior, F.B.; Goedert, W.J.; Mendes, I.C. Interpretation of microbial soil indicators as a function of crop yield and organic carbon. Soil Sci. Soc. Am. J. 2013, 77, 461–472. [Google Scholar] [CrossRef] [Green Version]
- Anzalone, R.A.; Vezzani, F.M.; Kaschuk, G.; Hungria, M.; Vargas, L.K.; Nogueira, M.A. Establishing reference values for soil microbial biomass-C in agroecosystems in the Atlantic Forest Biome in Southern Brazil. Ecol. Indic. 2020, 117, 106586. [Google Scholar] [CrossRef]
- Bonanomi, G.; De Filippis, F.; Zotti, M.; Idbella, M.; Cesarano, G.; Al-Rowaily, S.; Abd-ElGawad, A. Repeated applications of organic amendments promote beneficial microbiota, improve soil fertility and increase crop yield. Appl. Soil Ecol. 2020, 156, 103714. [Google Scholar] [CrossRef]
- Anderson, T.H.; Domsch, K.H. Soil microbial biomass: The eco-physiological approach. Soil Biol. Biochem. 2010, 42, 2039–2043. [Google Scholar] [CrossRef]
- Gamboa, C.H.; Vezzani, F.M.; Kaschuk, G.; Favaretto, N.; Cobos, J.Y.G.; Costa, G.A. Soil-root dynamics in maize-beans-eggplant intercropping system under organic management in a subtropical region. J. Soil Sci. Plant Nutr. 2020, 20, 1480–1490. [Google Scholar] [CrossRef]
- CONAB-Companhia Nacional de Abastecimento. Monitoramento Agrícola. Acompanhamento da Safra Brasileira de Grãos, v. 2–Safra 2014/15, n. 7–Sétimo Levantamento, abr. 2015. Available online: File:///C:/Users/User/Dropbox/Escrita%20em%20andamento/53_Artigo%20Max/Boletim_Graos_abril_2015.pdf. (accessed on 28 October 2021).
- Bacca, A.; Ceretta, C.A.; Kulmann, M.S.S.; Souza, R.O.S.; Ferreira, P.A.A.; Rodrigues, L.A.T.; Marchezan, C.; Garlet, L.P.; Brunetto, G. Residual and immediate effect after 16 applications of organic sources on yield and nitrogen use efficiency in black oat and corn. Rev. Bras. Ciênc. Do Solo 2020, 44, e0190013. [Google Scholar] [CrossRef]
- Plante, A.F.; Stone, M.M.; McGill, W.B. Chapter 9—The metabolic physiology of soil microorganisms. In Soil Microbiology, Ecology and Biochemistry, 4th ed.; Paul, E.A., Ed.; Academic Press: Cambridge, MA, USA, 2015; pp. 245–272. [Google Scholar] [CrossRef]
- Rheinheimer, D.S.; Anghinoni, I.; Conte, E. Fósforo da biomassa microbiana em solos sob diferentes sistemas de manejo. Rev. Bras. Ciênc. Do Solo 2000, 24, 589–597. [Google Scholar] [CrossRef] [Green Version]
- Srivastava, S.C.; Singh, J.S. Microbial C, N and P in dry tropical forest soils: Effects of alternate land-user and nutrient flux. Soil Biol. Biochem. 1991, 23, 117–124. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments 1 | Total Added by Fertilizers | Total Added by Crop Residue | ||||||
|---|---|---|---|---|---|---|---|---|
| TDM | TC | TN | TP | TDM | TC | TN | TP | |
| t ha−1 | ||||||||
| Control | - | - | - | - | 68 | 33.358 | 0.519 | 0.184 |
| MF | - | - | 1.120 | 0.576 | 106 | 54.252 | 0.874 | 0.348 |
| PS | 18.1 | 4.780 | 1.439 | 0.822 | 147 | 74.307 | 1.267 | 0.466 |
| CS | 67.7 | 15.258 | 1.942 | 0.765 | 123 | 63.415 | 1.120 | 0.413 |
| PL | 134.9 | 35.419 | 1.894 | 1.133 | 131 | 66.467 | 1.223 | 0.450 |
| Treatments 1 | Control † | MF | PS | CS | PL |
|---|---|---|---|---|---|
| Chemical attributes | |||||
| pH | 4.97 b | 4.30 c | 4.99 b | 5.10 b | 5.40 a |
| Ca—cmolc dm−3 | 1.22 d | 1.13 d | 2.61 c | 3.45 b | 5.52 a |
| Mg—cmolc dm−3 | 0.62 a | 0.85 a | 0.87 a | 1.12 a | 1.46 a |
| K—mg dm−3 | 23.10 d | 60.10 c | 73.40 b | 103.4 a | 111.0 a |
| P—mg dm−3 | 8.71 d | 41.37 c | 71.49 b | 75.94 b | 93.10 a |
| Al saturation—% | 44.18 b | 66.30 a | 27.40 c | 22.42 c | 9.55 d |
| V—% | 13.61 c | 9.16 c | 22.1 b | 27.91 b | 52.49 a |
| P—Total—mg kg−1 | 711 d | 856 c | 887 b | 995 b | 1257 a |
| P—Org—mg kg−1 | 4.52 d | 27.51 c | 45.17 b | 31.32 c | 104.60 a |
| COT—g kg−1 | 7.80 c | 9.40 b | 9.70 b | 12.60 a | 12.30 a |
| NT—g kg−1 | 0.70 c | 0.90 b | 1.00 b | 1.30 a | 1.20 a |
| COT/NT | 11.14 a | 10.44 a | 9.70 a | 9.96 a | 10.25 a |
| COT/P-org | 1725 a | 342 b | 215 c | 402 b | 118 d |
| Physical attributes | |||||
| DS—g cm−3 | 1.58 a | 1.51 a | 1.51 a | 1.44 a | 1.30 b |
| VTP—cm3 cm−3 | 0.40 c | 0.45 b | 0.45 b | 0.46 b | 0.56 a |
| Ma—cm3 cm−3 | 0.08 b | 0.12 a | 0.11 a | 0.09 b | 0.13 a |
| Mi—cm3 cm−3 | 0.32 c | 0.33 c | 0.34 c | 0.38 b | 0.43 a |
| DMG—mm | 0.49 a | 0.65 a | 0.69 a | 0.78 a | 0.76 a |
| Ksat—mm min−1 | 0.13 b | 0.68 a | 0.53 a | 0.75 a | 0.72 a |
| Kar—μm2 | 1.27 b | 7.60 a | 6.94 a | 8.01 a | 14.48 a |
| Wheat | ||||||
|---|---|---|---|---|---|---|
| Treatments 1 | C-BIO (mg kg−1 solo) | N-BIO (mg kg−1 solo) | P-BIO (mg kg−1 solo) | C-CO2 (mg C-CO2 kg−1 solo h−1) | qCO2 (μg C-CO2 mg−1 C-BIO h−1) | qMicro (%) |
| Control | 104.79 a | 26.45 b | 3.38 b | 0.25 b | 2.38 b | 1.34 a |
| MF | 121.22 a | 25.43 b | 5.96 a | 0.31 b | 2.59 b | 1.29 a |
| PS | 116.26 a | 31.49 a | 3.60 b | 0.31 b | 2.60 b | 1.20 a |
| CS | 126.40 a | 37.28 a | 5.09 a | 0.48 a | 3.79 a | 1.00 b |
| PL | 113.09 a | 34.77 a | 5.34 a | 0.49 a | 4.32 a | 0.93 b |
| Maize | ||||||
| C-BIO (mg kg−1 solo) | N-BIO (mg kg−1 solo) | P-BIO (mg kg−1 solo) | C-CO2 (mg C-CO2 kg−1 solo h−1) | qCO2 (μg C-CO2 mg−1 C-BIO h−1) | qMicro (%) | |
| Control | 123.10 b | 16.97 c | 4.25 c | 0.25 b | 2.12 c | 1.63 a |
| MF | 120.84 b | 19.08 c | 10.96 a | 0.57 a | 4.72 b | 1.32 b |
| PS | 120.96 b | 16.04 c | 9.62 a | 0.42 b | 3.42 b | 1.35 b |
| CS | 158.88 a | 32.86 a | 12.09 a | 0.65 a | 4.12 b | 1.36 b |
| PL | 128.95 b | 23.71 b | 8.35 b | 0.74 a | 5.83 a | 1.01 c |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ferreira, P.A.A.; Coronas, M.V.; Dantas, M.K.L.; Somavilla, A.; Brunetto, G.; Ceretta, C.A.; Giacomini, S.J.; Gubiani, P.I.; Boitt, G.; Soares, C.R.F.S.; et al. Repeated Manure Application for Eleven Years Stimulates Enzymatic Activities and Improves Soil Attributes in a Typic Hapludalf. Agronomy 2021, 11, 2467. https://doi.org/10.3390/agronomy11122467
Ferreira PAA, Coronas MV, Dantas MKL, Somavilla A, Brunetto G, Ceretta CA, Giacomini SJ, Gubiani PI, Boitt G, Soares CRFS, et al. Repeated Manure Application for Eleven Years Stimulates Enzymatic Activities and Improves Soil Attributes in a Typic Hapludalf. Agronomy. 2021; 11(12):2467. https://doi.org/10.3390/agronomy11122467
Chicago/Turabian StyleFerreira, Paulo A. A., Mariana V. Coronas, Max K. L. Dantas, André Somavilla, Gustavo Brunetto, Carlos A. Ceretta, Sandro J. Giacomini, Paulo I. Gubiani, Gustavo Boitt, Claudio R. F. S. Soares, and et al. 2021. "Repeated Manure Application for Eleven Years Stimulates Enzymatic Activities and Improves Soil Attributes in a Typic Hapludalf" Agronomy 11, no. 12: 2467. https://doi.org/10.3390/agronomy11122467
APA StyleFerreira, P. A. A., Coronas, M. V., Dantas, M. K. L., Somavilla, A., Brunetto, G., Ceretta, C. A., Giacomini, S. J., Gubiani, P. I., Boitt, G., Soares, C. R. F. S., Kaschuk, G., Bordallo, S. U., & Lourenzi, C. R. (2021). Repeated Manure Application for Eleven Years Stimulates Enzymatic Activities and Improves Soil Attributes in a Typic Hapludalf. Agronomy, 11(12), 2467. https://doi.org/10.3390/agronomy11122467